
Biological Consequences of Marine Energy Development on Marine Animals




1
Pacific Northwest National Laboratory, Coastal Sciences Division, Sequim, WA 98382, USA
2
Pacific Northwest National Laboratory, Coastal Sciences Division, Seattle, WA 98109, USA
*
Author to whom correspondence should be addressed.
Energies 2021, 14(24), 8460; https://doi.org/10.3390/en14248460
Submission received: 10 November 2021
/
Revised: 9 December 2021
/
Accepted: 12 December 2021
/
Published: 15 December 2021
(This article belongs to the Collection Women's Research in Wind and Ocean Energy)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Marine energy devices harness power from attributes of ocean water to form a sustainable energy source. Knowledge gaps remain about whether marine energy systems can affect the environment, adding another threat to animal populations and habitats already under pressure from climate change and anthropogenic activities. To date, potential environmental effects have been studied under the scope of stressor–receptor interactions, where moving parts of, or emissions from, a system could harm the animals, habitats, and natural processes. While crucial for understanding effects and identifying knowledge gaps, this approach misses a holistic view of what animals may experience in the presence of marine energy systems. We look at six biological consequences and forces that drive the health of an animal population and the effects expected from marine energy development: success of early life stages; changes in competitive capabilities; growth and survival based on food availability; susceptibility to predators; injury or death; and reproductive success. We use case studies to develop this approach, focusing on a variety of marine animals. An approximate level of risk is assigned for each interaction based on the biological consequences. This work highlights the need to examine the effects of marine energy development on animal populations within their natural habitats.
1. Introduction
Marine energy development is the latest entry into the portfolio of low-carbon sustainable energy sources for many coastal nations [1]. Marine energy is derived from the harvest of power from the movement of ocean water in high-energy areas through tides, ocean currents, and waves, and from temperature and salinity differentials [2]. The marine energy industry is young and facing challenges associated with the need for further technology development and convergence, as well as the need to acclimate to a hostile ocean environment to reach commercial readiness [3]. In addition, remaining knowledge gaps and uncertainties about the potential effects of these technologies on marine animals, habitats, and oceanographic processes have slowed development [3]. Regulatory processes in many nations require that the risk to vulnerable species and habitats be evaluated, minimized, and mitigated where needed.
Researchers in nations with active marine energy development have been investigating risks to marine animals from collision with rotating turbine blades, effects of sound from devices on marine mammals and fish, effects of electromagnetic fields (EMFs) from power cables on sensitive species, changes in critical habitats associated to the installation and operation of the devices, and a host of other interactions. Multinational research efforts have been spawned from the European Union [4] and the International Energy Agency’s Ocean Energy Systems [2]. To date, most efforts to relate the potential effects of marine energy on the environment have been approached from the perspective of the stressors, or portions of a device and associated systems that could cause stress, injury, or death to marine animals, or irreparable harm to habitats [5]. The large number of groups and species of marine animals and varied habitats that could be at risk from marine energy devices have limited the number of studies that examine the interactions from the point of view of the receptors (the animals and habitats that might be harmed by the devices and systems), except in very localized and species-specific instances [6,7,8,9], as well as in a small number of review papers (e.g., [10]).
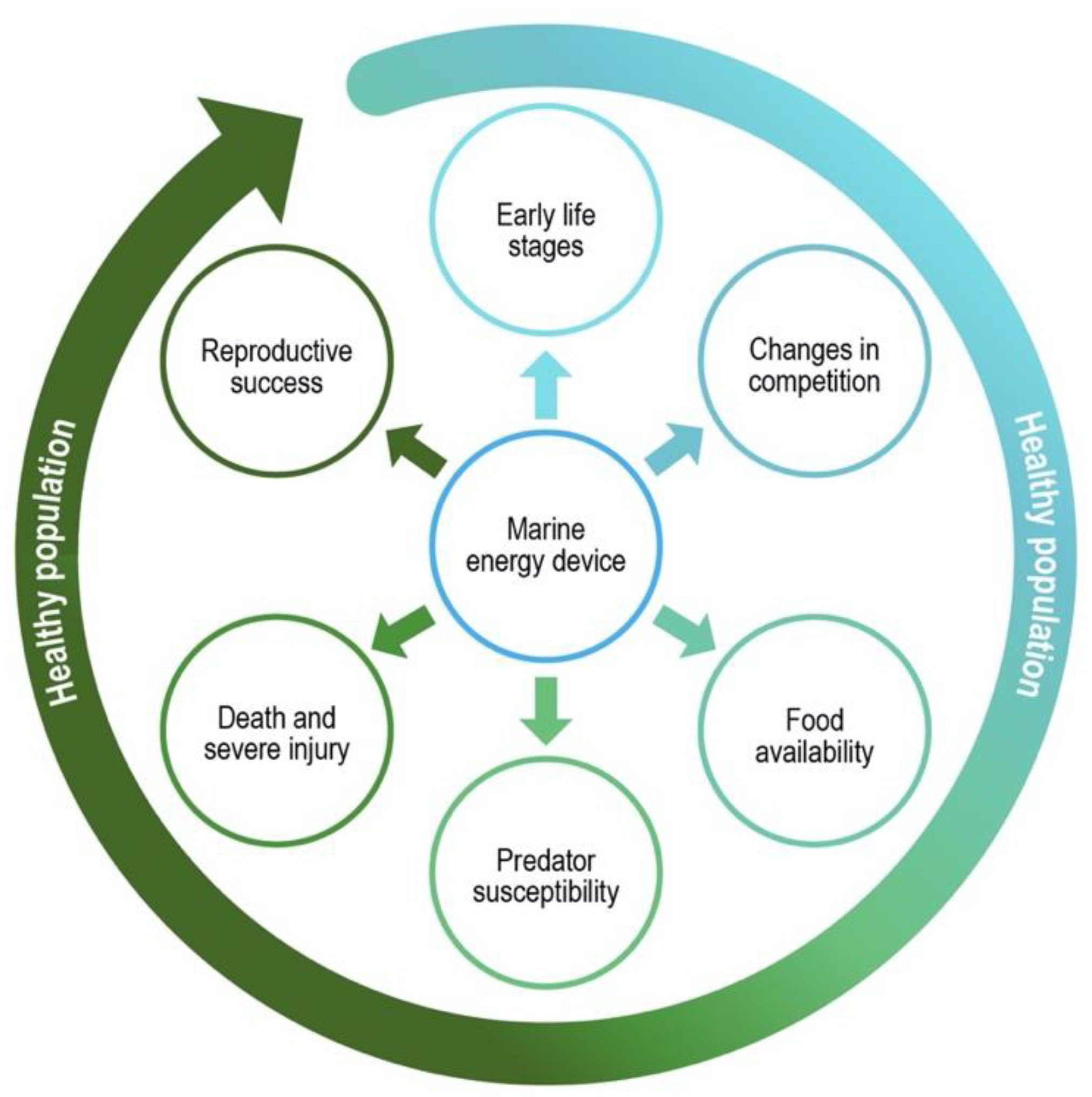
The present study examines how the risk of harm from marine energy development and operation might affect a broad range of receptor species. While the importance of acute harm to animals, such as collision with a rotating turbine blade severely injuring or killing an animal, can be easily understood, sublethal effects and chronic exposure to deleterious emissions are more difficult to assess. For example, the continuous low level of noise from an underwater marine energy device may affect stress levels in fish or marine mammals, and the cumulative exposure to EMFs from multiple power cables may affect the behavior of mobile invertebrates. To examine these sublethal effects, this paper evaluates outcomes of stressor–receptor interactions in terms of biological consequences, and various forces that cause them, that may potentially occur for specific receptor organisms. The biological consequences linked to marine energy used in this analysis are listed below and illustrated in Figure 1. These biological consequences are not an exhaustive list but cover most developmental, behavioral, and physiological changes that may impact population stability and maintenance. For each of these consequences, it is important to understand how interactions with marine energy devices might specifically affect the success of individuals from key species to contribute toward healthy populations. These consequences are:
- Success of early life stages, through development of larvae, juveniles, or young marine animals.
- Changes in competitive capabilities, through the ability of marine animals to effectively compete for mates, food, or space resources.
- Growth and survival based on food availability, allowing individuals and populations to thrive.
- Susceptibility to predators, through the ability of marine animals to avoid or escape predators.
- Injury or death from an encounter that may remove specific individuals from populations of marine animals.
- Reproductive success, through the ability of marine animals to mate and rear young to support their populations.
2. Methods
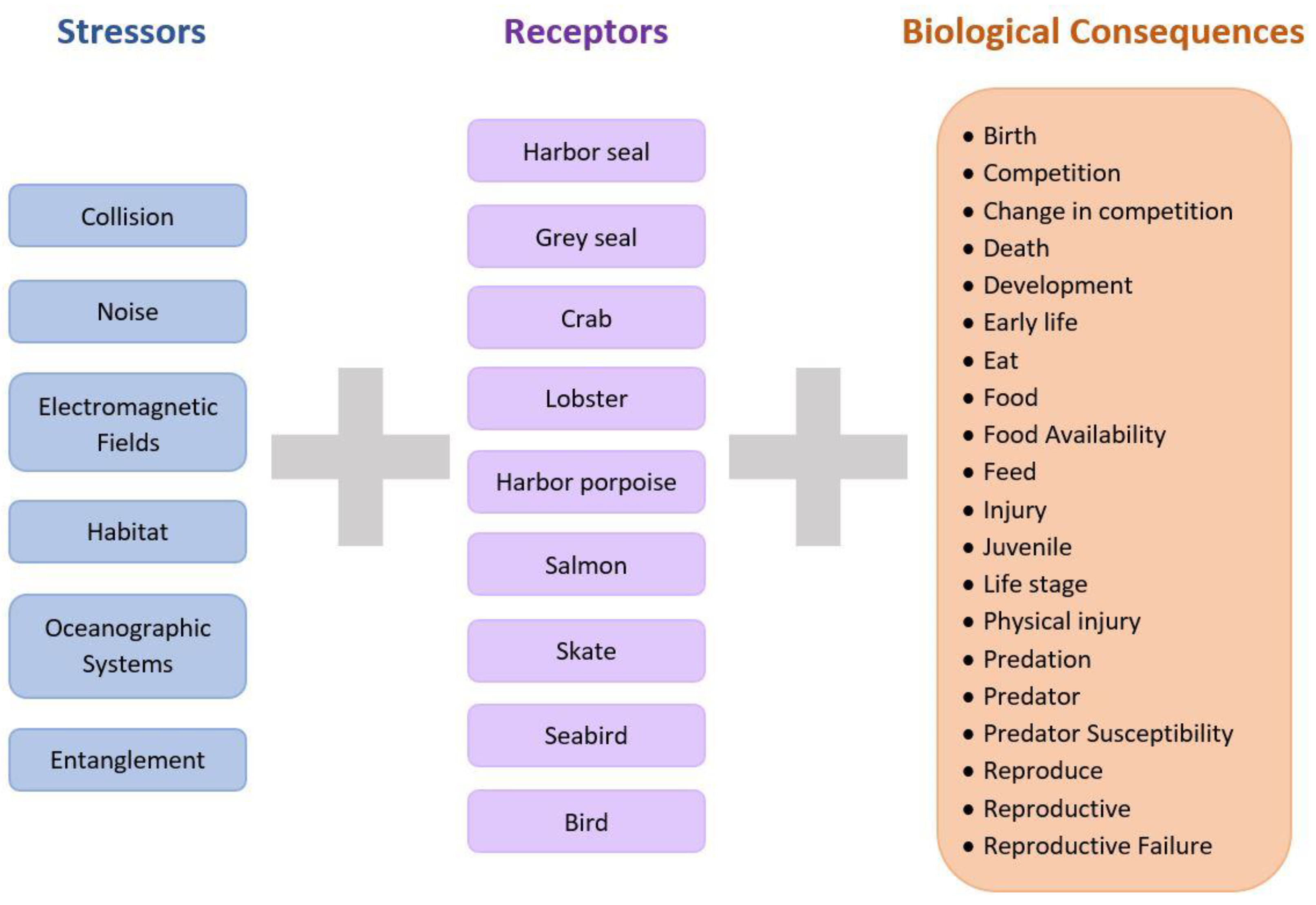
This paper is based on recent literature that addresses the interactions of marine animals with marine energy devices and systems, with a focus on the long-term operation of devices (i.e., multiple years), although aspects of construction and installation may also be relevant. Recent literature was collected using search engines, including Google and Google Scholar, and the dedicated collection of marine energy effects literature on the Tethys platform (https://tethys.pnnl.gov/) (accessed on 8 November 2021). Keywords and keyword combinations were used in the search to optimize the identification of relevant literature (Figure 2). All relevant literature included identifiers for marine energy, with key terms such as: wave energy, tidal device, marine renewable energy, or marine energy device, in addition to the stressors, receptors, and biological consequences.
The literature search found 99 documents, consisting mainly of journal articles, but also including conference papers, book chapters, magazine articles, dissertations, and reports. From each document, all relevant information about specific interactions, in addition to key information that describes specific biological consequences or forces for specific species (such as prey species, biological and physiological information, or habitat range), was collected. For each biological consequence, the existing literature was examined in the context of how specific stressors caused by marine energy may interact with the animal of concern, through the pathway of biological consequence. For example, how will the noise from an underwater turbine affect a harbor seal’s early life stages, reproductive success, susceptibility to predation, ability to compete for resources, and food availability, or result in a serious or fatal injury? The scientific literature that addresses the interactions of each species or group of species with marine energy devices was examined to determine the likely reactions of the animals to the marine energy systems, and the results were analyzed in the context of these biological consequences.
The groups of animals chosen for this study were those likely to be at risk from marine energy development around the world, and many can also be considered to be sentinel species for habitats common to high-energy areas (Figure 2). These groups include grey and harbor seals (Halichoerus grypus and Phoca vitulina, respectively), crabs and lobsters (e.g., European edible crab [Cancer pagurus] and American lobster [Homarus americanus]), harbor porpoise (Phocoena phocoena), salmon (e.g., Atlantic salmon [Salmo salar] and Chinook salmon [Oncorhynchus tshawytscha]), skates (e.g., little skates [Leucoraja erinacea]), and diving seabirds (e.g., common guillemot [Uria aalge] and great cormorants [Phalacrocorax carbo]). We focused on these six groups and ten species because they represent different components of a temperate marine ecosystem where marine energy devices may be deployed, and have been of greater interest to regulatory bodies because of their conservation and/or commercial status. We further developed two case studies (for harbor/grey seals and crabs/lobsters) because they are linked through a predator–prey relationship, which provided a multi-trophic-level representation.
3. Marine Energy Effects on Seals
Harbor seals are common marine mammals that live along the coastlines of the temperate northern hemisphere, whereas grey seals are only present along the shores of the North Atlantic in North America and Europe. Both species are often observed in estuaries and bays favorable to tidal energy development, and open coasts suitable for wave energy projects [11,12,13]. Although seals may be more sensitive than other marine animals to stressor–receptor interactions such as collision risk or underwater noise (Figure 3), other stressors such as changes in oceanographic systems and the emission of EMFs resulting from operational marine energy devices have no direct biological consequences for these animals. In addition, entanglement with mooring lines is not likely to be a threat to seals because lines associated with marine energy devices are taut and do not present loose ends [14,15]. However, lost or drifting fishing gear caught in devices’ mooring lines may cause a risk of secondary entanglement for seals [16].
Early life stages. Mother seals nurse their pups for 4 to 6 weeks then abandon them once they are weaned, leaving juvenile seals to discover the environment on their own at a relatively young age [17]. Marine energy devices may have undesirable effects on the seals’ early life stage development, especially through collision risks as inexperienced young seals approach tidal turbines. A report by Thompson [18] showed that young seals extensively use areas of high tidal currents during the first few months of their life, which could make them vulnerable to encountering the rotating blades of tidal turbines.
Change in competition. Competition among seals focuses on the availability of food resources and finding mates during the reproduction season. The ability of seals to adapt to changes in their environment due to the presence and operation of marine energy devices may have an effect on their competitive success. Seals display an exceptional ability to learn and adapt to their environments, based on acoustic cues [19]. Underwater sound cues from a marine energy device may alternately attract a seal to a successful foraging spot or warn of a hazardous location. The individuals who learn the appropriate reaction to these cues may be more successful competitors than their counterparts.
Food availability. Grey and harbor seals are opportunistic carnivores, feeding mainly on fish, as well as crustaceans and mollusks, and occasionally seabirds. Marine energy devices may act as artificial reefs, attracting fish and other mobile organisms such as crabs [20,21], and subsequently may increase food availability for seals, particularly because they may act to connect seal foraging habitats [22,23]. However, it has been shown that seals do not always forage around marine energy devices despite the prey availability, particularly in tidal channels where studies have also shown that pinnipeds forage more actively during flood than ebb tide [11]. In addition, seals may be able to recognize the acoustic cues of rotating tidal turbines and choose to forage elsewhere to avoid what they may perceive as a risk that the turbines may cause [24].
Death and severe injury. Even though monitoring around wave and tidal energy devices has reported few, if any, instances of severe injury or death of seals due to operational devices [25], risk of collision with the moving parts of turbines remains the most critical biological consequence for seals from operating marine energy devices. Similarly, the effects of underwater noise from devices could be harmful to the animals’ ability to navigate [25]. However, seals may use acoustic cues from an operating tidal turbine to avoid the immediate area of the turbine well before the levels of underwater noise may cause any temporary or permanent hearing loss or damage [26,27]. In addition, studies have shown that if a collision were to occur between a seal and a turbine blade, it is unlikely that a collision with the flat side of a blade would cause any injury, and impacts with the edge of a blade at the maximum effective speed would be unlikely to cause any skeletal damage [28,29,30]. While a head injury to a seal may lead to disorientation and drowning [31], results of modeling and field tests carried out with seal carcasses showed that severe injuries were limited to the thorax, and that the severity of the injuries decreased with increasing blubber thickness [29,32].
Knowledge gaps. Although the scientific literature provides some information about several biological consequences of marine energy devices for seals (i.e., early life stage development, change in competition, food availability, and death and severe injury), additional research is necessary to understand the full extent of the risk. In particular, it is uncertain whether interactions with marine energy devices may cause stress, leading to reproductive failure, or whether the presence and operation of devices may affect seals’ susceptibility to their natural predators, including orca and sharks, or capability to find their prey.
4. Marine Energy Effects on Crabs and Lobsters
Although seals feed mostly on fish, they do not hesitate to resort to eating crustaceans in the absence of their preferred prey. Crabs and lobsters are essential elements of benthic ecosystems, acting as scavengers and pursuing an opportunistic diet, and helping to clean up carcasses and other detritus in benthic habitats [33]. The installation and operation of marine energy devices may affect crabs and lobsters in various ways, throughout their different life stages (Figure 3). Risk of collision with turbine blades and entanglement in mooring lines do not affect crabs and lobsters because they reside on the seafloor and are very small in relation to marine energy devices, making it possible for them to wiggle their way out of potential entanglement hazards.
Early life stages. During the early life stages, crab and lobster larvae, the megalopae, may be vulnerable to underwater noise produced by marine energy devices. Megalopae use natural underwater sounds as cues to settle in benthic habitats, and anthropogenic noise may mask these cues [34]. Tidal turbine noise, along with other anthropogenic marine noise, has been shown to influence the settling behavior of megalopae in several estuarine crab species and delay their metamorphosis [34,35,36]. Although some stakeholders are concerned that EMFs emitted by marine energy devices and export cables may affect crustaceans, and especially young life stages, a laboratory experiment showed that anthropogenic EMFs at intensities similar to those of marine energy systems did not significantly affect juvenile lobster sheltering behavior [37]. Changes in oceanographic systems due to large numbers of operating marine energy devices are likely to modify the currents that larvae rely on for dispersal. In addition, new habitats provided by marine energy devices will likely increase the availability of shelter in which larvae can settle, and juvenile crabs and lobsters can hide from predators. However, there is a lack of research in these areas that undermines our understanding of these potential effects on early life stages.
Change in competition. Crabs and lobsters of the same species compete for food, mates, and shelter. Marine energy devices and their associated systems, such as foundations, mooring anchors, and power export cables, have been shown to provide additional structure in benthic environments and act as artificial reefs, offering new shelter and sources of food [21,38]. Crabs and/or lobsters were observed using nooks and crannies on device foundations, under cables, and on cable protection such as concrete mattresses [39,40,41]. These prime shelters may spark competition and physical encounters. A decrease in roaming behavior for the European edible crab (Cancer pagurus) was demonstrated in the presence of EMFs, causing the crab to limit their time foraging for food, or searching for shelter or mates. These actions amount to a decrease in potential opportunities for competition [42].
Food availability. Because crab and lobster are opportunistic feeders and will eat whatever prey or carcass they find, marine energy devices are unlikely to affect their food availability. As described above, marine energy devices act as artificial reefs and increase local biomass and biodiversity [38,43], providing more food opportunities for crabs and lobsters. Underwater noise and EMFs emitted by marine energy devices may disrupt crabs’ and lobsters’ foraging and feeding behavior. However, ship noise, which is much louder and more common in marine environments than marine energy device noise, has been shown to have no significant effect on crab foraging ability in a laboratory experiment, but to significantly affect their feeding behavior—crabs interrupt their meals more often when exposed to ship noise versus playback of ambient noise [44]. The effects of marine energy device noise on crab feeding behavior remain to be explored. An enclosure experiment over an energized cable showed significant changes in lobster behavior when exposed to EMFs compared to a control cable, suggesting an increase in foraging explorations [45].
Predator susceptibility. In addition to seals, crabs and lobsters are targeted by numerous predators such as other marine mammals, including sea otters, various fish species, sharks, and invertebrates, including octopus, sea stars, and other crabs and lobsters. The artificial reef and fish-aggregating effects of marine energy devices may increase predation on crabs and lobsters by attracting an increasing number of predators. Several reef-associated fish species such as sea bass have been shown to feed on crabs [46]. Excessive underwater noise can distract crabs and make them more susceptible to predators [44], although no study has been undertaken in the context of marine energy device noise. Laboratory experiments showed that crabs and lobsters buried themselves or took shelter over 75% of the time, even in the presence of EMFs [47], or exhibited a significant decrease in time spent roaming when exposed to EMFs [42], suggesting they may remain less exposed to predators.
Knowledge gaps. It is assumed that EMFs may affect crab and lobster behavior, but no laboratory experiment has demonstrated significant trauma or differences in survival between exposed and control settings [48]. In addition, crustaceans are known to experience pain and stress [49] but the extent to which EMFs, underwater noise, or any other interactions with marine energy devices may increase stress-related hormones in crabs and lobsters, which may lead to reproductive failure, remains unknown.
5. Marine Energy Effects on Other Receptors
Although our thorough literature review focused on seals and crabs/lobsters, a less in-depth approach was applied to various other receptors including cetaceans such as harbor porpoises, anadromous fish such as salmon, benthic fish such as skates, and diving seabirds.
5.1. Harbor Porpoise
Harbor porpoise live in most coastal and estuarine waters of the northern hemisphere, where they are likely to interact with anthropogenic activities at sea, including marine energy devices. Their ability to hear, locate, and respond to tidal turbines at a distance will modify their risk of collision, and fatal injuries are unlikely [8]. Collision risk and underwater noise from marine energy devices may cause severe injury or death to harbor porpoises; however, no increase in injured or dead porpoises has been observed near marine energy sites [6,50,51]. Recent research indicates that harbor porpoises have the ability to avoid tidal turbines in close proximity, which will limit their risk of collision [6]. Harbor porpoises feed on non-spiny fish such as herring, cod, whiting, mackerel, sardines, and occasionally squid or octopus [52]. Because marine energy devices act as artificial reefs, providing structure and habitat for many fish species and attracting small pelagic fish, harbor porpoise food availability may be increased in the presence of marine energy devices [20,21,23,53]. However, harbor porpoises’ ability to find their prey may be hindered in the near vicinity of devices due to the potential masking of acoustic cues [54]. Although predator susceptibility of harbor porpoises around operating marine energy devices has not been directly studied, no change in predation has been observed at wave and tidal energy sites to date (e.g., [55,56,57]). Harbor porpoise predators are mainly large sharks, orcas, and dolphins. While shark distribution in tidal channels remains largely unknown, some species forage in areas and at depths where tidal turbines may be placed [58,59]. Interactions between sharks and harbor porpoises are more likely along an open coast where wave energy converters (WECs) may be present; however, no such interactions around marine energy devices have been recorded to date. A handful of studies have focused on orcas [60] and dolphins [8,61,62] in high-energy environments, but it remains uncertain how their behavior around marine energy devices may increase their susceptibility to collision risk and underwater noise. Whether marine energy devices pose a greater risk for young porpoises than adults, influence competition among harbor porpoises, or lead to reproductive failure due to added stress remains to be investigated. Risk from EMFs, changes in oceanographic systems, and entanglement are less of a concern because harbor porpoises do not spend time in close proximity to the seafloor and are unlikely to be affected by taut mooring lines [16,55,63].
5.2. Salmon
To date, there have been few specific studies of how marine energy devices may lead to consequences for, or changes in, salmon early life stage development, competition, food availability, severe injury or death, predator susceptibility, or reproductive failure (e.g., [64]). Although salmon and other salmonid species (e.g., trout) are potentially at risk of physical injury or death if struck by blades when swimming around tidal and riverine turbines [10,65,66], the severity of the injuries and likely consequences are still uncertain. EMFs from marine energy cables may disrupt salmon foraging and swimming behavior due to their magneto-sensitivity [67,68], potentially affecting their competitive and reproductive success, but this interaction has not been studied. Tidal turbines create a reef effect and aggregate fish, and when placed in estuaries could provide new habitat for juvenile salmon [21,69], although any positive or negative effects on juvenile salmon have not been observed. Adult salmon are unlikely to detect the sound pressure element of underwater sound from tidal turbines [51,70,71]. Although no studies have examined the effect of the particle motion component of underwater sound on salmon, their sensitivity to this component may lead to adverse biological consequences, such as increased susceptibility to predators or diminished reproductive success. Changes in oceanographic systems and risk of entanglement with mooring systems are considered to be very low risk for salmon [16,72].
5.3. Skates
Skates are known to detect EMFs, particularly the fields induced by their prey [73]. Based on the skates’ sensory capability, stakeholders have expressed concern that EMFs emitted by marine energy cables may adversely affect skate populations. However, studies have shown negligible effects, including barrier effects, direct physical injuries, or adverse physiological effects, due to marine energy cables [74,75]. Although there have been no specific studies on changes in skate habitat due to marine energy devices, a large body of research suggests that loss of habitat is unlikely, and that there may be beneficial effects [51,76,77,78]. Skates feed on bottom-dwelling animals such as shrimp, crab, and clams. The artificial reef effect around marine energy devices could provide a suitable habitat for skates [21], while also increasing food availability [79] and opportunities for competition (Figure 3). The extent to which skates may use new habitats provided by devices and cables to attach their egg capsules is unknown. No studies have examined skates’ use of marine energy systems to lay their eggs, and it is unknown whether EMFs from the power cables and devices may affect the development of skate early life stages. The reef effect may also attract skate predators, including seals, but uncertainties remain about whether skates are more susceptible to predators around marine energy devices. However, seals are not known to frequently forage around marine energy devices, despite the prey availability [24,26,80]. Gaps remain in our knowledge of potential added stress and reproductive failure due to the installation and operation of marine energy devices.
5.4. Diving Seabirds
While seabirds do not reside in the oceans, they dive to various depths depending on the species, chasing fish and other prey. These diving forays may provide some level of risk for collision with the rotating blades of tidal turbines when diving depths overlap with turbine depths (Figure 3). Collisions of seabirds with turbine blades may lead to serious injuries or death, but current knowledge does not support the likelihood of this risk [81]. A lack of behavioral data for seabirds in tidal environments, poor underwater visibility at most sites that limits visual observations [82], and unknown consequences of collision limit our understanding of the risk. Whether young seabirds are more at risk for collisions with tidal turbines due to their lack of experience remains unknown. It is also unknown whether interactions with marine energy devices may influence seabirds’ susceptibility to predators or affect their reproductive success. Small pelagic fish are attracted to localized and persistent foraging hotspots, providing increased food availability for seabirds; these same areas that attract birds may be optimum for siting tidal turbines, causing an increased risk of collision [83]. Seabirds are unlikely to be affected by EMFs, changes in oceanographic systems, underwater noise, and entanglement with marine energy devices because of their water column use, dive patterns, and the tautness of mooring lines (e.g., [14,63,84]). Significant gaps remain in our knowledge of changes in competition among seabirds due to the presence and operation of marine energy devices.
6. Risk to Species from Marine Energy Development
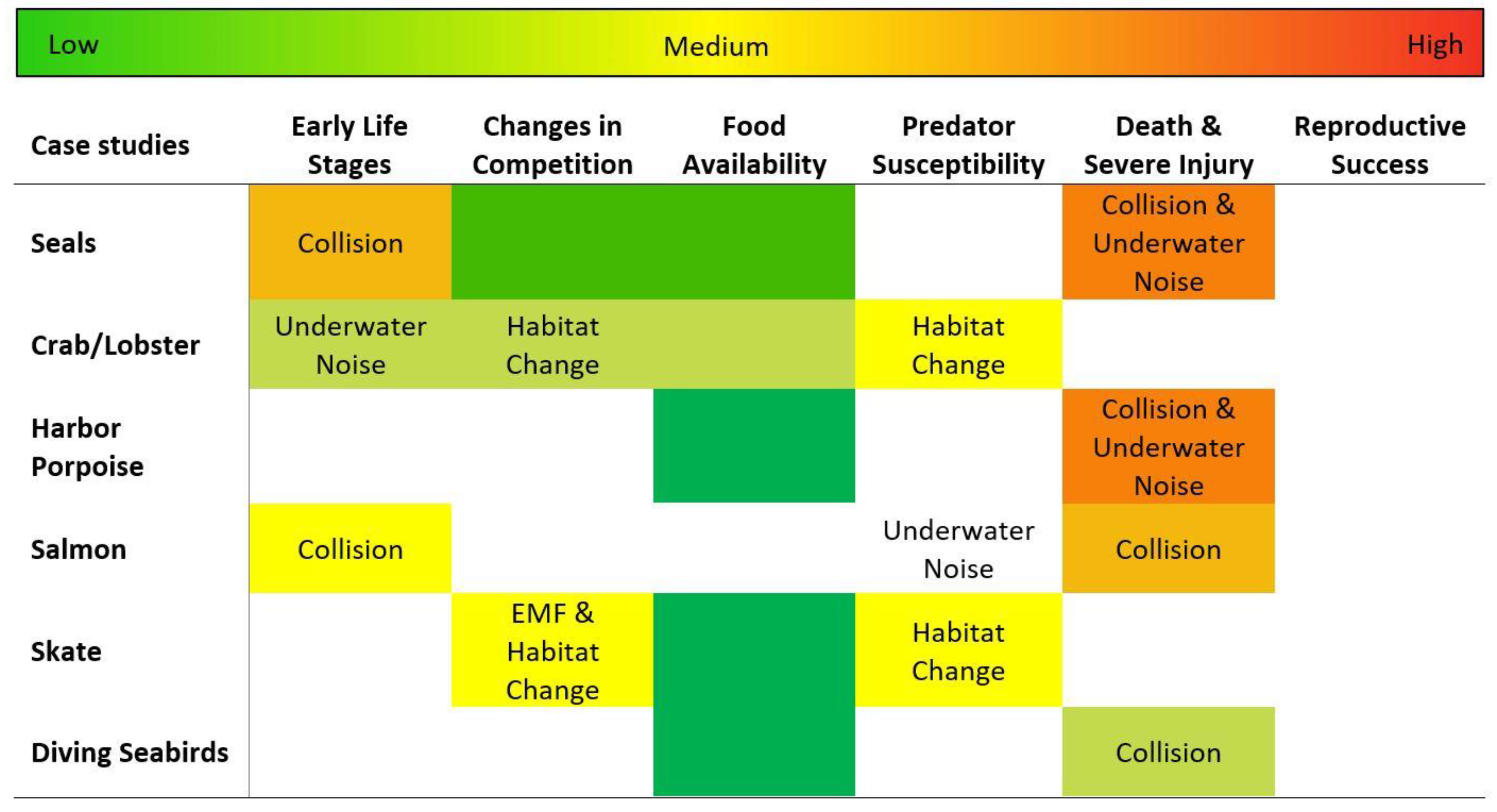
Relying on the existing literature, an approximate level of risk was assigned for each interaction of animals with marine energy devices or systems, based on each biological consequence (Figure 4). Risk was estimated as the intersection of the probability of a negative outcome, and the consequences if that event occurred [85]. Risk implies a negative outcome of the interaction, but in some cases, the interaction might create positive outcomes; for example, the presence of a marine energy device might provide increased food availability.
Each level of risk was assigned based on the following qualitative criteria:
- High risk = likely to interact with severe consequences
- Medium-high risk = high likelihood of interaction, outcomes could be severe
- Medium risk = reasonable likelihood of interaction, outcomes could be detrimental
- Medium-low risk = low likelihood of interaction, outcomes not likely to be severe
- Low risk = unlikely to interact
- Insufficient information to make an estimate.
The risk assignment took into consideration the additive effects of any relevant stressor–receptor interactions on the biological consequences, based on the information available in the literature. In many cases, the higher risks indicate some information about deleterious outcomes, but the uncertainty level remains high. For example, the risk of death and severe injury to seals and harbor porpoises remains relatively high as there are few definitive observations of collision or active evasion of turbine blades.
7. Discussion
The six biological consequences used in this study encompass most developmental, behavioral, and physiological changes that may have implications for population stability and maintenance. Growth of individuals is reflected through effects on early life stage development that includes the stages where significant growth takes place during an animal’s life cycle. Effects that may result in changes in competition reflect any situation where competition for mates, food resources, or shelter is affected by a physical, biological, or chemical change, resulting from an interaction with a marine energy device. The biological consequence of food availability incorporates any situation where prey abundance, food sources, or ability to detect prey may be altered by the presence of marine energy devices. The consequences of predator susceptibility describe any changes in the chances of survival of a prey animal due to the presence and operation of marine energy devices. Death and/or severe injury from interaction with a marine energy device will occur as the device, parts of the system, or emissions cause physical changes or injuries that result in the animal’s death or inability to maintain body integrity and continue its usual activities. Reproductive success incorporates any disturbance to an animal’s ability to reproduce, which can range from physical separation or injury caused by the device to biological or chemical changes that disrupt mating behavior, attraction between individuals, or other aspects of reproduction. The factors that increase the risk for the marine animals examined in this study are most influenced by the part of the water column that the species groups use and the ecological niche they occupy in the wild.
To maintain a healthy population of marine animals, by definition, a large portion of individuals must succeed relative to all six of the biological consequences examined here. Failure in any portion may lead to a reduced population over time [86]. The presence and operation of marine energy devices apply to species groups through differing pathways of consequence. Effects on early life stages are most likely to be derived from disruption of the dispersal of pelagic larvae for benthic and demersal organisms such as crabs and lobsters, or skates, or from severe injuries to inexperienced young seals, porpoises, salmon, or seabirds if struck by turbine blades. Similarly, changes in competitive capabilities because of the presence of marine energy anchors and lines are likely to affect the organisms living on the seafloor who may suffer from loss of preferred habitat, although conversely, these same organisms may benefit from the formation of new habitat on the same infrastructure [87]. How these effects scale up with arrays of multiple devices remains to be fully understood. Growth and survival based on food availability are hard to link directly to the presence or operation of marine energy devices for the species groups examined, except for the possibility of providing additional small to medium-size fish and biofouling organisms in the vicinity of the devices that could provide predators and browsers with additional food [69,88,89]. Although the presence of marine energy devices is likely to attract prey species, it is difficult to determine whether this could adversely affect entire prey species populations. The likelihood of injury or death of marine mammals, fish, or potentially seabirds from turbine blade collision remains one of the least understood risks of marine energy [32,90]. If this were shown to be a significant issue with certain populations, the removal of sufficient individuals of reproductive age could limit the reproductive success of the population. Similarly, interference with the mating and rearing of young due to the presence of the devices could become a problem, though no evidence of this has been reported.
The location occupied by a species in the water column or on the seabed affects the susceptibility of that group to specific aspects of marine energy installation and operation. Benthic invertebrates and demersal fish living on or in proximity to the seafloor are more likely to be affected by the presence of a foundation or anchor points and lines and cables than those living higher in the water column, due to loss or alteration of critical habitat and exposure to EMFs from export cables. The intensity of the effects will depend on the size and number of foundations, anchors, and cables, and whether they are buried. The placement of bottom-based equipment for marine energy devices must be considered in the presence of bottom-living species, including those examined in this study: crabs, lobsters, and skates [37].
Animals examined in this study that live near the surface or throughout the water column, such as the harbor seal, harbor porpoise, or salmon, and to lesser extent diving seabirds, are less likely to be affected by the physical presence of a marine energy device because they can easily avoid a surface expression such as a WEC or float, the mooring lines that stretch through the water column, and inter-array power cables that span the space between marine energy devices at mid-depth or near the surface. The alteration or loss of pelagic habitat caused by well-sited marine energy will be small and animals are likely to adapt to these changes quickly. These pelagic animals are, however, more likely to be susceptible to injury or death from a rotating tidal turbine blade, although it is generally thought that collision with blades is likely to be very rare, based on the space available around devices in addition to the sensory, behavior, and swimming capabilities, such as evasion, of the pelagic species [6,81]. Additional observations and validated models of turbine blade encounter and collision are needed to better understand this dynamic, and mitigation strategies may be needed to lessen the likelihood or severity of such encounters.
Noise from the operation of marine energy devices is most likely to affect marine animals near the spreading source [54]. Marine energy generators are generally close to the surface (although some may be placed on the bottom or mid-water column). Thus, pelagic animals, including marine mammals, fish, diving seabirds, and sea turtles who may swim close to or even aggregate around devices, are most likely to be affected. There is also some evidence that planktonic larvae of crustaceans may be affected by underwater noise [35]. A few types of bottom-mounted wave devices may produce underwater noise in closer proximity to benthic invertebrates such as crabs or lobsters and demersal fish, although it is not known how many of these species are likely to be affected.
Changes in oceanographic conditions due to the presence and operation of marine energy devices are likely to affect only those marine animals with broadcast eggs and planktonic or small motile larvae. Estimates of the potential changes in flow and other oceanographic processes due to the development of marine energy are commonly provided by hydrodynamic models fit with simulated devices [91]. The model results indicate relatively small changes in flow fields that may cause some alterations in the distribution of floating and planktonic larvae, but they are unlikely to be at a magnitude that will alter population success [92]. None of these model outputs have been validated. Marine mammals, fish, sea turtles, and diving seabirds are unlikely to be adversely affected by changes in oceanographic conditions at the magnitudes predicted, even by large marine energy arrays.
Research on the interactions of marine energy devices and the marine environment has largely focused on specific interactions that are likely to trigger regulatory requirements or restrictions [3]. However, these studies have focused largely on the effects on individual animals, and little is known about how marine energy effects might radiate through populations. It is apparent that no attention has been paid to potential effects on the reproductive success of populations due to the presence and operation of marine energy devices (Figure 4). There has also been little attention paid to the intersection of multiple stressors from marine energy devices on species groups, or on how these effects might synergistically affect populations. For example, perhaps the sound from a tidal turbine attracts fish, placing them at greater risk for collision with a tidal blade; or the additional habitat created by a WEC foundation and anchor might attract crabs, placing them in closer proximity to EMF emissions from a power cable. The synergy of stressors from marine energy will require additional investigations, particularly as more devices are placed in the water and large arrays amass off coastlines.
Even less well studied are the potential indirect effects of marine energy device operation on the overall marine ecosystem, including changes that might occur over time to dissolved nutrients and gases, potentially altering primary productivity, and causing reverberations throughout the marine food web. Other indirect effects are possible, and the understanding of these effects is still in its infancy. It is important to note, however, that the shifting baselines of seawater chemistry and primary production due to climate change are already altering marine food webs as we know them [93]. Separating the effects of marine energy, first at a small scale, then at larger development scales, will be very challenging. Similarly, separating the effects of marine energy installation, operation, maintenance, and, finally, decommissioning from other anthropogenic effects of industries such as shipping, commercial fishing, oil and gas operations, and shoreline development, in addition to the cumulative effects of these various sources, will prove to be very complicated.
8. Conclusions
Investigations of the potential effects of marine energy development are a crucial part of moving this new industry forward. The value of providing low-carbon sustainable and renewable energy must not come at the cost of degradation of the marine environment and damage to fragile populations of marine animals and their habitats. To date, most investigations have focused on single-device stressors and individual animals or species. Although considerable research is still needed to understand these somewhat simplified interactions, there is also a need for new tools and approaches to understand what additional effects might occur, in order to proactively minimize deleterious outcomes. Examining potential interactions from the perspective of the receptors—the animals and habitats—provides an additional pathway to classifying, understanding, and ameliorating potential harm. The approach of investigating potential impacts on biological consequences and forces can provide a starting point for exploring a population’s ability to survive long-term effects of marine energy operation, in addition to ecosystem-wide effects of local disturbances. These case studies of major species groups of concern for marine energy development have provided a glimpse into how we might move beyond the need to examine the interaction of each stressor with each potential receptor species at all marine energy sites. In particular, this work can help apply the knowledge we have gained about the basic stressor–receptor interactions, and relate them to the location where animals live, in addition to the life histories of species groups. This increased understanding can be applied to careful siting and management of marine energy devices. As we better understand how species groups are likely to interact with operational marine energy, we can better project how large arrays will affect the overall marine environment over time, and begin to understand the cumulative effects of offshore renewables with other anthropogenic activities, all against the changing backdrop of climate change.
Author Contributions
Conceptualization, D.M.O., A.E.C., L.G.H.; methodology, D.M.O., L.G.H., A.E.C.; data curation, D.M.O.; writing, L.G.H., A.E.C., D.M.O.; project administration, A.E.C.; funding acquisition, A.E.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research has been made possible due to the financial support of the U.S. Department of Energy, Energy Efficiency and Renewable Energy Water Power Technologies Office to Pacific Northwest National Laboratory under contract DE-AC05-76RL01830.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We are grateful for the ongoing support and guidance from the U.S. Department of Energy (DOE), Water Power Technologies Office, and the collaboration with DOE staff, including Samantha Eaves. We also thank Susan Ennor for editorial assistance and Stephanie King for the illustrations.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- IRENA Renewable Energy Statistics 2019; The International Renewable Energy Agency: Abu Dhabi, United Arab Emirates, 2019; pp. 1–398. ISBN 978-92-9260-137-9.
- Ocean Energy Systems IEA-OES Annual Report: An Overview of Ocean Energy Activities in 2020; The Executive Committee of Ocean Energy Systems: Lisbon, Portugal, 2021; pp. 1–188.
- Copping, A.E.; Hemery, L.G. OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 1–327. [Google Scholar]
- Prittimaa, L.; Cruz, E. Ocean Energy and the Environment: Research and Strategic Actions. Report to the European Commission; European Technology and Innovation Platform for Ocean Energy (ETIP): Brussel, Belgium, 2020; p. 36. [Google Scholar]
- Boehlert, G.; Gill, A. Environmental and Ecological Effects of Ocean Renewable Energy Development—A Current Synthesis. Oceanography 2010, 23, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, D.; Palmer, L.; Macaulay, J.; Sparling, C.; Hastie, G. Harbour porpoises exhibit localized evasion of a tidal turbine. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 2459–2468. [Google Scholar] [CrossRef]
- Harwood, J.; King, S. The Sensitivity of UK Marine Mammal Populations to Marine Renewables Developments; Report No. SMRUL-NER-2012-027; SMRU Consulting: St Andrews, UK, 2014; pp. 1–53. [Google Scholar]
- Malinka, C.; Gillespie, D.; Macaulay, J.; Joy, R.; Sparling, C. First in situ passive acoustic monitoring for marine mammals during operation of a tidal turbine in Ramsey Sound, Wales. Mar. Ecol. Prog. Ser. 2018, 590, 247–266. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.J.; Blondel, P.; Williamson, L.D.; Scott, B.E. Application of a multibeam echosounder to document changes in animal movement and behaviour around a tidal turbine structure. ICES J. Mar. Sci. 2021, 78, 1253–1266. [Google Scholar] [CrossRef]
- Copping, A.E.; Hemery, L.G.; Viehman, H.; Seitz, A.C.; Staines, G.J.; Hasselman, D.J. Are fish in danger? A review of environmental effects of marine renewable energy on fishes. Biol. Conserv. 2021, 262, 109297. [Google Scholar] [CrossRef]
- Hastie, G.D.; Russell, D.J.F.; Benjamins, S.; Moss, S.; Wilson, B.; Thompson, D. Dynamic habitat corridors for marine predators; intensive use of a coastal channel by harbour seals is modulated by tidal currents. Behav. Ecol. Sociobiol. 2016, 70, 2161–2174. [Google Scholar] [CrossRef] [Green Version]
- Lieber, L.; Nimmo-Smith, W.A.M.; Waggitt, J.J.; Kregting, L. Fine-scale hydrodynamic metrics underlying predator occupancy patterns in tidal stream environments. Ecol. Indic. 2018, 94, 397–408. [Google Scholar] [CrossRef] [Green Version]
- SMRU Ltd. Utilisation of Space by Grey and Harbour Seals in the Pentland Firth and Orkney Waters; Scottish Natural Heritage: Oban, UK, 2011; p. 62. [Google Scholar]
- Benjamins, S.; Harnois, V.; Smith, H.; Johanning, L.; Greenhill, L.; Carter, C.; Wilson, B. Understanding the Potential for Marine Megafauna Entanglement Risk from Marine Renewable Energy Developments; Scottish Natural Heritage: Perth, UK, 2014; p. 95. [Google Scholar]
- Minesto Deep Green Holyhead Deep Project Phase I (0.5 MW)—Environmental Statement (Report No. L-100194-S14-EIAS-001); Minesto: Holyhead, UK, 2016; pp. 1–487.
- Garavelli, L. Encounters of Marine Animals with Marine Renewable Energy Device Mooring Systems and Subsea Cables. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 146–153. [Google Scholar] [CrossRef]
- Muelbert, M.M.C.; Bowen, W.D. Duration of lactation and postweaning changes in mass and body composition of harbour seal, Phoca vitulina, pups. Can. J. Zool. 1993, 71, 1405–1414. [Google Scholar] [CrossRef]
- Thompson, D. Annex I: Movements and Diving Behaviour of Juvenile Grey Seals in Areas of High Tidal Energy; RPS Group: Chepstow, UK, 2012; p. 55. [Google Scholar]
- Stansbury, A.L.; Götz, T.; Deecke, V.B.; Janik, V.M. Grey seals use anthropogenic signals from acoustic tags to locate fish: Evidence from a simulated foraging task. Proc. R. Soc. B Boil. Sci. 2015, 282, 20141595. [Google Scholar] [CrossRef] [Green Version]
- Fraser, S.; Williamson, B.J.; Nikora, V.; Scott, B.E. Fish distributions in a tidal channel indicate the behavioural impact of a marine renewable energy installation. Energy Rep. 2018, 4, 65–69. [Google Scholar] [CrossRef]
- Hemery, L. Changes in Benthic and Pelagic Habitats Caused by Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 105–125. [Google Scholar] [CrossRef]
- Arnould, J.P.Y.; Monk, J.; Ierodiaconou, D.; Hindell, M.A.; Semmens, J.; Hoskins, A.J.; Costa, D.P.; Abernathy, K.; Marshall, G.J. Use of Anthropogenic Sea Floor Structures by Australian Fur Seals: Potential Positive Ecological Impacts of Marine Industrial Development? PLoS ONE 2015, 10, e0130581. [Google Scholar] [CrossRef] [Green Version]
- Williamson, B.; Fraser, S.; Williamson, L.; Nikora, V.; Scott, B. Predictable changes in fish school characteristics due to a tidal turbine support structure. Renew. Energy 2019, 141, 1092–1102. [Google Scholar] [CrossRef]
- Hastie, G.D.; Lepper, P.; McKnight, J.C.; Milne, R.; Russell, D.J.F.; Thompson, D. Acoustic risk balancing by marine mammals: Anthropogenic noise can influence the foraging decisions by seals. J. Appl. Ecol. 2021, 58, 1854–1863. [Google Scholar] [CrossRef]
- Smith, K.; Simpson, N. Enabling Future Arrays in Tidal: Y1 Environmental Monitoring Report. EnFiat. 2018. Available online: https://tethys.pnnl.gov/sites/default/files/publications/Y1_Environmental_Monitoring_Report.pdf (accessed on 22 July 2021).
- Hastie, G.D.; Russell, D.J.; Lepper, P.; Elliott, J.; Wilson, B.; Benjamins, S.; Thompson, D. Harbour seals avoid tidal turbine noise: Implications for collision risk. J. Appl. Ecol. 2017, 55, 684–693. [Google Scholar] [CrossRef]
- Polagye, B.; Bassett, C. Risk to Marine Animals from Underwater Noise Generated by Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 67–86. [Google Scholar] [CrossRef]
- Band, B.; Sparling, C.; Thompson, D.; Onoufriou, J.; San Martin, E.; West, N. Refining Estimates of Collision Risk for Harbour Seals and Tidal Turbines; Report for Scottish Government; ABP Marine Environmental Research Ltd. (ABPmer); Marine Scotland Science: Edinburgh, UK, 2016; pp. 1–133. [Google Scholar]
- Copping, A.; Grear, M.; Jepsen, R.; Chartrand, C.; Gorton, A. Understanding the potential risk to marine mammals from collision with tidal turbines. Int. J. Mar. Energy 2017, 19, 110–123. [Google Scholar] [CrossRef]
- Thompson, D.; Onoufriou, J.; Brownlow, A.; Morris, C. Data Based Estimates of Collision Risk: An Example Based on Harbour Seal Tracking Data around a Proposed Tidal Turbine Array in the Pentland Firth; Natural Scottish Heritage: Perth, UK, 2016; pp. 1–41. [Google Scholar]
- Thompson, D.; Brownlow, A.; Onoufriou, J.; Moss, S. Collision Risk and Impact Study: Field Tests of Turbine Blade-Seal Carcass Collisions; Report to Scottish Government MR 7.2.3; Marine Scotland Science: Edinburgh, UK, 2015; pp. 1–16. [Google Scholar]
- Onoufriou, J.; Brownlow, A.; Moss, S.; Hastie, G.; Thompson, D. Empirical determination of severe trauma in seals from collisions with tidal turbine blades. J. Appl. Ecol. 2019, 56, 1712–1724. [Google Scholar] [CrossRef]
- Boudreau, S.A.; Worm, B. Ecological role of large benthic decapods in marine ecosystems: A review. Mar. Ecol. Prog. Ser. 2012, 469, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Pine, M. Underwater Anthropogenic Sound: Understanding the Potential Impacts on the Marine Environment and the Influence on Crab Larval Behaviour. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2013; pp. 1–152. [Google Scholar]
- Pine, M.K.; Jeffs, A.G.; Radford, C.A. Turbine Sound May Influence the Metamorphosis Behaviour of Estuarine Crab Megalopae. PLoS ONE 2012, 7, e51790. [Google Scholar] [CrossRef] [Green Version]
- Pine, M.K.; Jeffs, A.G.; Radford, C.A. Effects of Underwater Turbine Noise on Crab Larval Metamorphosis. In The Effects of Noise on Aquatic Life II; Popper, A.N., Hawkins, A., Eds.; Springer: New York, NY, USA, 2016; pp. 847–852. [Google Scholar]
- Taormina, B.; Di Poi, C.; Agnalt, A.-L.; Carlier, A.; Desroy, N.; Escobar-Lux, R.H.; D’eu, J.-F.; Freytet, F.; Durif, C.M.F. Impact of magnetic fields generated by AC/DC submarine power cables on the behavior of juvenile European lobster (Homarus gammarus). Aquat. Toxicol. 2020, 220, 105401. [Google Scholar] [CrossRef]
- Langhamer, O. The location of offshore wave power devices structures epifaunal assemblages. Int. J. Mar. Energy 2016, 16, 174–180. [Google Scholar] [CrossRef]
- Bicknell, A.W.J.; Sheehan, E.V.; Godley, B.J.; Doherty, P.D.; Witt, M.J. Assessing the impact of introduced infrastructure at sea with cameras: A case study for spatial scale, time and statistical power. Mar. Environ. Res. 2019, 147, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Langhamer, O.; Wilhelmsson, D. Colonisation of fish and crabs of wave energy foundations and the effects of manufactured holes—A field experiment. Mar. Environ. Res. 2009, 68, 151–157. [Google Scholar] [CrossRef] [Green Version]
- Taormina, B.; Laurans, M.; Marzloff, M.P.; Dufournaud, N.; Lejart, M.; Desroy, N.; Leroy, D.; Martin, S.; Carlier, A. Renewable energy homes for marine life: Habitat potential of a tidal energy project for benthic megafauna. Mar. Environ. Res. 2020, 161, 105131. [Google Scholar] [CrossRef]
- Scott, K.; Harsanyi, P.; Lyndon, A.R. Understanding the effects of electromagnetic field emissions from Marine Renewable Energy Devices (MREDs) on the commercially important edible crab, Cancer pagurus (L.). Mar. Pollut. Bull. 2018, 131, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Causon, P.D.; Gill, A.B. Linking ecosystem services with epibenthic biodiversity change following installation of offshore wind farms. Environ. Sci. Policy 2018, 89, 340–347. [Google Scholar] [CrossRef]
- Wale, M.A.; Simpson, S.D.; Radford, A.N. Noise negatively affects foraging and antipredator behaviour in shore crabs. Anim. Behav. 2013, 86, 111–118. [Google Scholar] [CrossRef]
- Hutchison, Z.L.; Gill, A.B.; Sigray, P.; He, H.; King, J.W. Anthropogenic electromagnetic fields (EMF) influence the behaviour of bottom-dwelling marine species. Sci. Rep. 2020, 10, 4219. [Google Scholar] [CrossRef] [Green Version]
- Leitão, F.; Santos, M.N.; Erzini, K.; Monteiro, C.C. The effect of predation on artificial reef juvenile demersal fish species. Mar. Biol. 2008, 153, 1233–1244. [Google Scholar] [CrossRef]
- Woodruff, D.; Cullinan, V.; Copping, A.; Marshall, K. Effects of Electromagnetic Fields on Fish and Invertebrates—FY2012 Progress Report (Report No. PNNL-22154); Report for US Department of Energy (DOE); Pacific Northwest National Laboratory (PNNL): Richland, WA, USA, 2013; pp. 1–62. [Google Scholar]
- Bochert, R.; Zettler, M.L. Long-term exposure of several marine benthic animals to static magnetic fields. Bioelectromagnetics 2004, 25, 498–502. [Google Scholar] [CrossRef]
- Appel, M.; Elwood, R.W. Motivational trade-offs and potential pain experience in hermit crabs. Appl. Anim. Behav. Sci. 2009, 119, 120–124. [Google Scholar] [CrossRef]
- Keenan, G.; Sparling, C.; Williams, H.; Fortune, F. SeaGen Environmental Monitoring Programme: Final Report; Royal Haskoning Enhancing Society: Edinburgh, UK, 2011; p. 81. [Google Scholar]
- Haskoning, R. 2012 Environmental Monitoring Report Final Draft; Report for Marine Current Turbines (MCT); Ocean Renewable Power Company (ORPC): Portland, ME, USA, 2013; p. 653. [Google Scholar]
- National Wildlife Federation. Harbor Porpoise. Available online: https://www.nwf.org/Home/Educational-Resources/Wildlife-Guide/Mammals/Harbor-Porpoise (accessed on 13 October 2021).
- Mikkelsen, L.; Mouritsen, K.N.; Dahl, K.; Teilmann, J.; Tougaard, J. Re-established stony reef attracts harbour porpoises Phocoena phocoena. Mar. Ecol. Prog. Ser. 2013, 481, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Lossent, J.; Lejart, M.; Folegot, T.; Clorennec, D.; Di Iorio, L.; Gervaise, C. Underwater operational noise level emitted by a tidal current turbine and its potential impact on marine fauna. Mar. Pollut. Bull. 2018, 131, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Oregon State University. Final License Application for the PacWave South Project; Oregon State University: Corvallis, OR, USA, 2019; p. 1681. [Google Scholar]
- Robertson, F.; Wood, J.; Joslin, J.; Joy, R.; Polagye, B. Marine Mammal Behavioral Response to Tidal Turbine Sound; University of Washington: Seattle, WA, USA, 2018; pp. 1–65. [Google Scholar]
- Tollit, D.; Joy, R.; Wood, J.; Redden, A.; Booth, C.; Boucher, T.; Porskamp, P.; Oldreive, M. Baseline Presence of and Effects of Tidal Turbine Installation and Operations on Harbour Porpoise in Minas Passage, Bay of Fundy, Canada. J. Ocean Technol. 2019, 14, 22–48. [Google Scholar]
- Roberts, L.; Harding, H.R.; Voellmy, I.; Bruintjes, R.; Simpson, S.D.; Radford, A.N.; Breithaupt, T.; Elliott, M. Exposure of benthic invertebrates to sediment vibration: From laboratory experiments to outdoor simulated pile-driving. Proc. Meet. Acoust. 2016, 27, 010029. [Google Scholar] [CrossRef] [Green Version]
- Staines, G.; Zydlewski, G.B.; Viehman, H.A.; Kocik, R. Applying Two Active Acoustic Technologies to Document Presence of Large Marine Animal Targets at a Marine Renewable Energy Site. J. Mar. Sci. Eng. 2020, 8, 704. [Google Scholar] [CrossRef]
- Bassett, C. Ambient Noise in an Urbanized Tidal Channel. Ph.D. Thesis, University of Washington, Seattle, WA, USA, 2013. Available online: https://ui.adsabs.harvard.edu/abs/2013PhDT.......269B/abstract (accessed on 14 June 2021).
- Cruz, E.; Simas, T. Discussion of the Effects of the Underwater Noise Radiated by a Wave Energy Device-Portugal. In Proceedings of the 11th European Wave and Tidal Energy Conference, Nantes, France, 6–11 September 2015; pp. 1–5. [Google Scholar]
- Palmer, L.; Gillespie, D.; Macaulay, J.; Onoufriou, J.; Sparling, C.; Thompson, D.; Hastie, G. Marine Mammals and Tidal Energy: Annual Report to Scottish Government-MRE Theme; Sea Mammal Research Unit, University of St. Andrews: St Andrews, UK, 2019; pp. 1–32. [Google Scholar]
- Gill, A.B.; Desender, M. Risk to Animals from Electromagnetic Fields Emitted by Electric Cables and Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 86–103. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Haro, A. Survival and Behavioral Effects of Exposure to a Hydrokinetic Turbine on Juvenile Atlantic Salmon and Adult American Shad. Estuaries Coasts 2015, 38, 203–214. [Google Scholar] [CrossRef]
- Bevelhimer, M.S.; Pracheil, B.M.; Fortner, A.M.; Saylor, R.; Deck, K.L. Mortality and injury assessment for three species of fish exposed to simulated turbine blade strike. Can. J. Fish. Aquat. Sci. 2019, 76, 2350–2363. [Google Scholar] [CrossRef]
- Xodus Group. Brims Tidal Array Collision Risk Modelling—Atlantic Salmon; Report No. A-100242-S02-TECH-001; Xodus Group: London, UK, 2016; pp. 1–11. [Google Scholar]
- Snyder, D.; Bailey, W.; Palmquist, K.; Cotts, B.; Olsen, K. Evaluation of Potential EMF Effects on Fish Species of Commercial or Recreational Fishing Importance in Southern New England; U.S. Department of the Interior, Bureau of Ocean Energy Management: Sterling, VA, USA, 2019; p. 62. [Google Scholar]
- Wyman, M.T.; Klimley, A.P.; Battleson, R.D.; Agosta, T.V.; Chapman, E.D.; Haverkamp, P.J.; Pagel, M.D.; Kavet, R. Behavioral responses by migrating juvenile salmonids to a subsea high-voltage DC power cable. Mar. Biol. 2018, 165, 134. [Google Scholar] [CrossRef]
- Kramer, S.H.; Hamilton, C.D.; Spencer, G.C.; Ogston, H.O. Evaluating the Potential for Marine and Hydrokinetic Devices to Act as Artificial Reefs or Fish Aggregating Devices. Based on Analysis of Surrogates in Tropical, Subtropical, and Temperate U.S. West Coast and Hawaiian Coastal Waters; U.S. Department of Energy Efficiency and Renewable Energy: Golden, CO, USA, 2015; p. 90. [Google Scholar] [CrossRef]
- Bevelhimer, M.S.; Deng, Z.D.; Scherelis, C. Characterizing large river sounds: Providing context for understanding the environmental effects of noise produced by hydrokinetic turbines. J. Acoust. Soc. Am. 2016, 139, 85–92. [Google Scholar] [CrossRef]
- Halvorsen, M.B.; Carlson, T.J.; Copping, A.E. Effects of Tidal Turbine Noise on Fish Hearing and Tissues—Draft Final Report—Environmental Effects of Marine and Hydrokinetic Energy; Pacific Northwest National Lab. (PNNL-20786): Richland, WA, USA, 2011; pp. 1–48. [Google Scholar]
- Guerra, M.; Thomson, J. Wake measurements from a hydrokinetic river turbine. Renew. Energy 2019, 139, 483–495. [Google Scholar] [CrossRef]
- Camperi, M.; Tricas, T.C.; Brown, B.R. From Morphology to Neural Information: The Electric Sense of the Skate. PLoS Comput. Biol. 2007, 3, e113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fundy Ocean Research Center for Energy (FORCE). Environmental Effects Monitoring Report 2011–2013; FORCE: Parrsboro, NS, Canada, 2014; pp. 1–44. [Google Scholar]
- Hutchison, Z.; Sigray, P.; He, H.; Gill, A.; Gibson, C. Electromagnetic Field (EMF) Impacts on Elasmobranch (Shark, Rays, and Skates) and American Lobster Movement and Migration from Direct Current Cables; U.S. Department of the Interior, Bureau of Ocean Energy Management: Sterling, VA, USA, 2018; pp. 1–254. [Google Scholar]
- Henkel, S. Assessment of Benthic Effects of Anchor Presence and Removal; Report for Oregon Wave Energy Trust (OWET); Northwest National Marine Renewable Energy Center (NNMREC): Corvallis, OR, USA, 2016; pp. 1–22. [Google Scholar]
- Kregting, L.; Elsaesser, B.; Kennedy, R.; Smyth, D.; O’Carroll, J.; Savidge, G. Do Changes in Current Flow as a Result of Arrays of Tidal Turbines Have an Effect on Benthic Communities? PLoS ONE 2016, 11, e0161279. [Google Scholar] [CrossRef] [Green Version]
- Kuhnz, L.; Barry, J.; Buck, B.; Whaling, P. Potential Impacts of the Monterey Accelerated Research System (MARS) Cable on the Seabed and Benthic Faunal Assemblages; Monterey Bay Aquarium Research Institute: Moss Landing, CA, USA, 2011; pp. 1–58. [Google Scholar]
- Packer, D.; Zetlin, C.; Vitaliano, J. Essential Fish and Habitat Source Document: Thorny Skate, Amblyraja Radiata, Life History and Habitat Characteristics; U.S. Department of Commerce National Oceanic and Atmospheric Administration: Woods Hole, MA, USA, 2003; pp. 1–50. [Google Scholar]
- Hastie, G.; Lepper, P.; McKnight, C.; Milne, R.; Russell, D.; Thompson, D. Risk Balancing by a Marine Mammal: Man-Made Noise Reduces the Foraging Success of Seals [Abstract]. In Proceedings of the World Marine Mammal Conference (WMMC2019), Barcelona, Spain, 9–12 December 2019; pp. 1–2. [Google Scholar]
- Sparling, C.E.; Seitz, A.C.; Masden, E.; Smith, K. Collision Risk for Animals around Turbines. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 29–65. [Google Scholar] [CrossRef]
- RPS Group. A Review of the Potential Use of Sonar to Observe the Underwater Behaviour of Diving Birds near Tidal Energy Devices (Report No. Report 051); Report for Scottish Natural Heritage; RPS Group: Chepstow, UK, 2010; pp. 1–26. [Google Scholar]
- Lieber, L.; Nimmo-Smith, W.A.M.; Waggitt, J.J.; Kregting, L. Localised anthropogenic wake generates a predictable foraging hotspot for top predators. Commun. Biol. 2019, 2, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCluskie, A.; Langston, A.; Wilkinson, N. Birds and Wave & Tidal Stream Energy: An Ecological Review; The Royal Society for the Protection of Birds (RSPB): Sandy, UK, 2012; p. 124. [Google Scholar]
- Gibbs, M.T.; Browman, H.I. Risk assessment and risk management: A primer for marine scientists. ICES J. Mar. Sci. 2015, 72, 992–996. [Google Scholar] [CrossRef] [Green Version]
- Holt, E.; Miller, S. Bioindicators: Using Organisms to Measure Environmental Impacts. Nat. Educ. Knowl. 2011, 3, 8. [Google Scholar]
- Bender, A.; Langhamer, O.; Sundberg, J. Colonisation of wave power foundations by mobile mega- and macrofauna—A 12 year study. Mar. Environ. Res. 2020, 161, 105053. [Google Scholar] [CrossRef]
- Langhamer, O. Artificial Reef Effect in Relation to Offshore Renewable Energy Conversion: State of the Art. Sci. World J. 2012, 2012, 386713. [Google Scholar] [CrossRef]
- Wilhelmsson, D.; Langhamer, O. The Influence of Fisheries Exclusion and Addition of Hard Substrata on Fish and Crustaceans. In Humanity and the Sea: Marine Renewable Energy Technology and Environmental Interactions; Springer: Berlin/Heidelberg, Germany, 2014; pp. 49–60. [Google Scholar]
- Copping, A.E.; Grear, M.E. Applying a simple model for estimating the likelihood of collision of marine mammals with tidal turbines. Int. Mar. Energy J. 2018, 1, 27–33. [Google Scholar] [CrossRef]
- Whiting, J.M.; Chang, G. Changes in Oceanographic Systems Associated with Marine Renewable Energy Devices. In OES-Environmental 2020 State of the Science Report: Environmental Effects of Marine Renewable Energy Development Around the World; Copping, A.E., Hemery, L.G., Eds.; Ocean Energy Systems: Seattle, WA, USA, 2020; pp. 127–145. [Google Scholar] [CrossRef]
- Wang, T.; Yang, Z.; Copping, A. A Modeling Study of the Potential Water Quality Impacts from In-Stream Tidal Energy Extraction. Estuaries Coasts 2015, 38, 173–186. [Google Scholar] [CrossRef]
- Nagelkerken, I.; Goldenberg, S.U.; Ferreira, C.M.; Ullah, H.; Connell, S.D. Trophic pyramids reorganize when food web architecture fails to adjust to ocean change. Science 2020, 369, 829–832. [Google Scholar] [CrossRef]
Figure 1.
In the presence of marine energy devices, specific forces and biological consequences integrate throughout the life cycle of a marine animal to condition its success in contributing toward a healthy population. (Illustration by Stephanie King/Pacific Northwest National Laboratory).
Figure 1.
In the presence of marine energy devices, specific forces and biological consequences integrate throughout the life cycle of a marine animal to condition its success in contributing toward a healthy population. (Illustration by Stephanie King/Pacific Northwest National Laboratory).

Figure 2.
Keyword search terms used in identifying relevant literature in Tethys, Google, and Google Scholar included all stressors (left column), the selected receptors (middle column), and biological consequences (right column) in various combinations. For documents where “harbor seal” or “harbor porpoise” was searched, the British English spelling “harbour” was also included.
Figure 2.
Keyword search terms used in identifying relevant literature in Tethys, Google, and Google Scholar included all stressors (left column), the selected receptors (middle column), and biological consequences (right column) in various combinations. For documents where “harbor seal” or “harbor porpoise” was searched, the British English spelling “harbour” was also included.

Figure 3.
Interactions of seal, crab, and lobster with marine energy devices, including a horizontal axis tidal turbine (lower right) and a point absorber wave energy converter (background left). The organisms are shown in their natural habitats, including interactions with their prey and predators. (Illustration by Stephanie King/Pacific Northwest National Laboratory).
Figure 3.
Interactions of seal, crab, and lobster with marine energy devices, including a horizontal axis tidal turbine (lower right) and a point absorber wave energy converter (background left). The organisms are shown in their natural habitats, including interactions with their prey and predators. (Illustration by Stephanie King/Pacific Northwest National Laboratory).

Figure 4.
Estimated levels of risk for interactions of animals with marine energy devices, as described in the case studies. Where information is available, the predominant interaction driving the risk is noted. The cells are color coded according to the color bar on the top—from green indicating a fairly well-resolved low risk, through yellow, orange, and red, indicating increasingly higher levels of risk. White cells indicate a lack of information and continuing uncertainty about risk. Research to date supports the highest level of certainty around the low-risk interactions (green cells) indicated in this table. The level of uncertainty associated with medium and high risks (yellow to orange cells) increases considerably as there are fewer studies and greater challenges in observing these interactions.
Figure 4.
Estimated levels of risk for interactions of animals with marine energy devices, as described in the case studies. Where information is available, the predominant interaction driving the risk is noted. The cells are color coded according to the color bar on the top—from green indicating a fairly well-resolved low risk, through yellow, orange, and red, indicating increasingly higher levels of risk. White cells indicate a lack of information and continuing uncertainty about risk. Research to date supports the highest level of certainty around the low-risk interactions (green cells) indicated in this table. The level of uncertainty associated with medium and high risks (yellow to orange cells) increases considerably as there are fewer studies and greater challenges in observing these interactions.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hemery, L.G.; Copping, A.E.; Overhus, D.M. Biological Consequences of Marine Energy Development on Marine Animals. Energies 2021, 14, 8460. https://doi.org/10.3390/en14248460
AMA Style
Hemery LG, Copping AE, Overhus DM. Biological Consequences of Marine Energy Development on Marine Animals. Energies. 2021; 14(24):8460. https://doi.org/10.3390/en14248460
Chicago/Turabian StyleHemery, Lenaïg G., Andrea E. Copping, and Dorian M. Overhus. 2021. "Biological Consequences of Marine Energy Development on Marine Animals" Energies 14, no. 24: 8460. https://doi.org/10.3390/en14248460
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.