Deadwood Characteristics in Mature and Old-Growth Birch Stands and Their Implications for Carbon Storage

 , and
, and 
Abstract
:1. Introduction
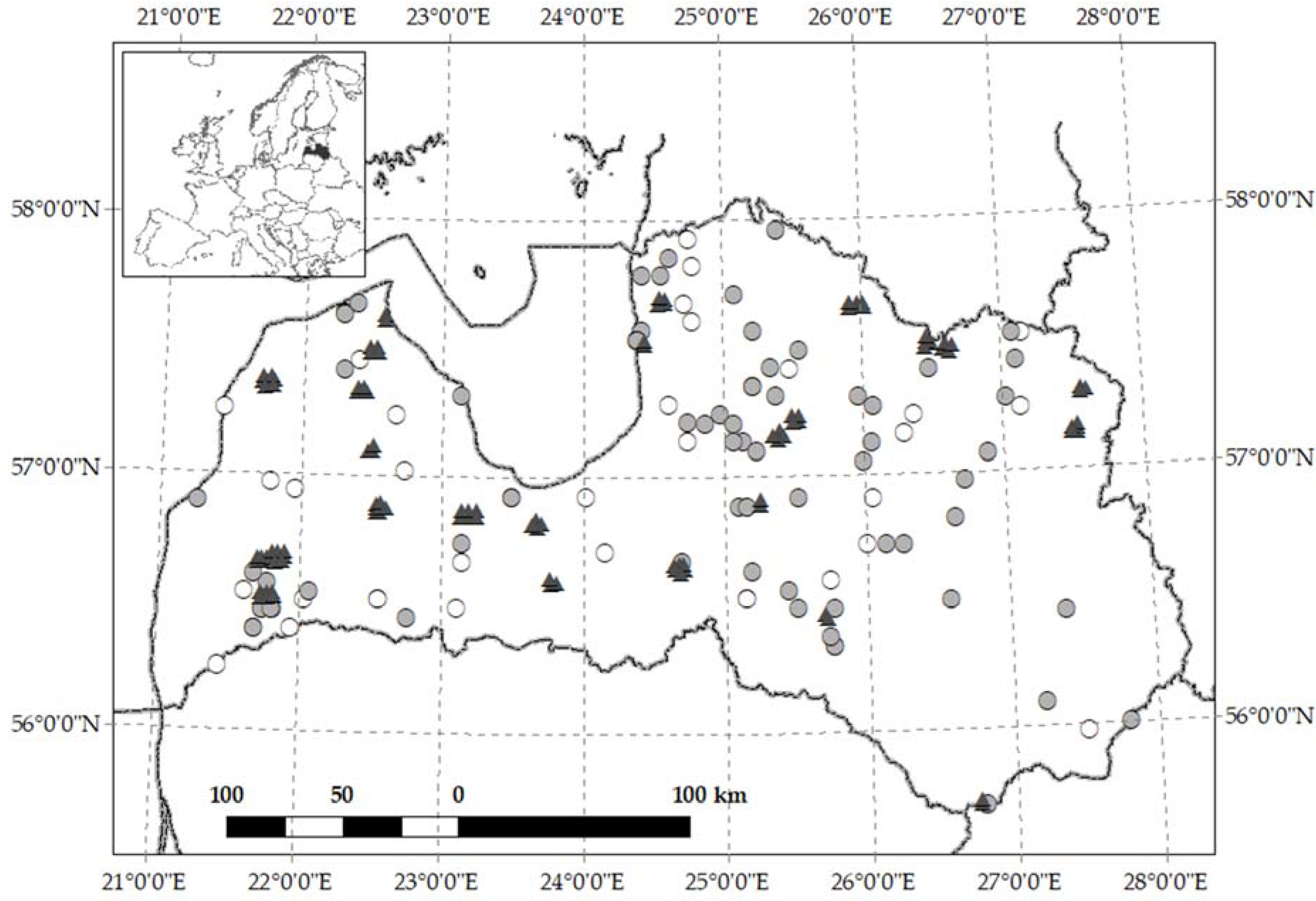
2. Materials and Methods
- V = Stump/snag volume,
- L = Length of the log or height of the stump, and
- dm = Mid-diameter of the log or the stump.
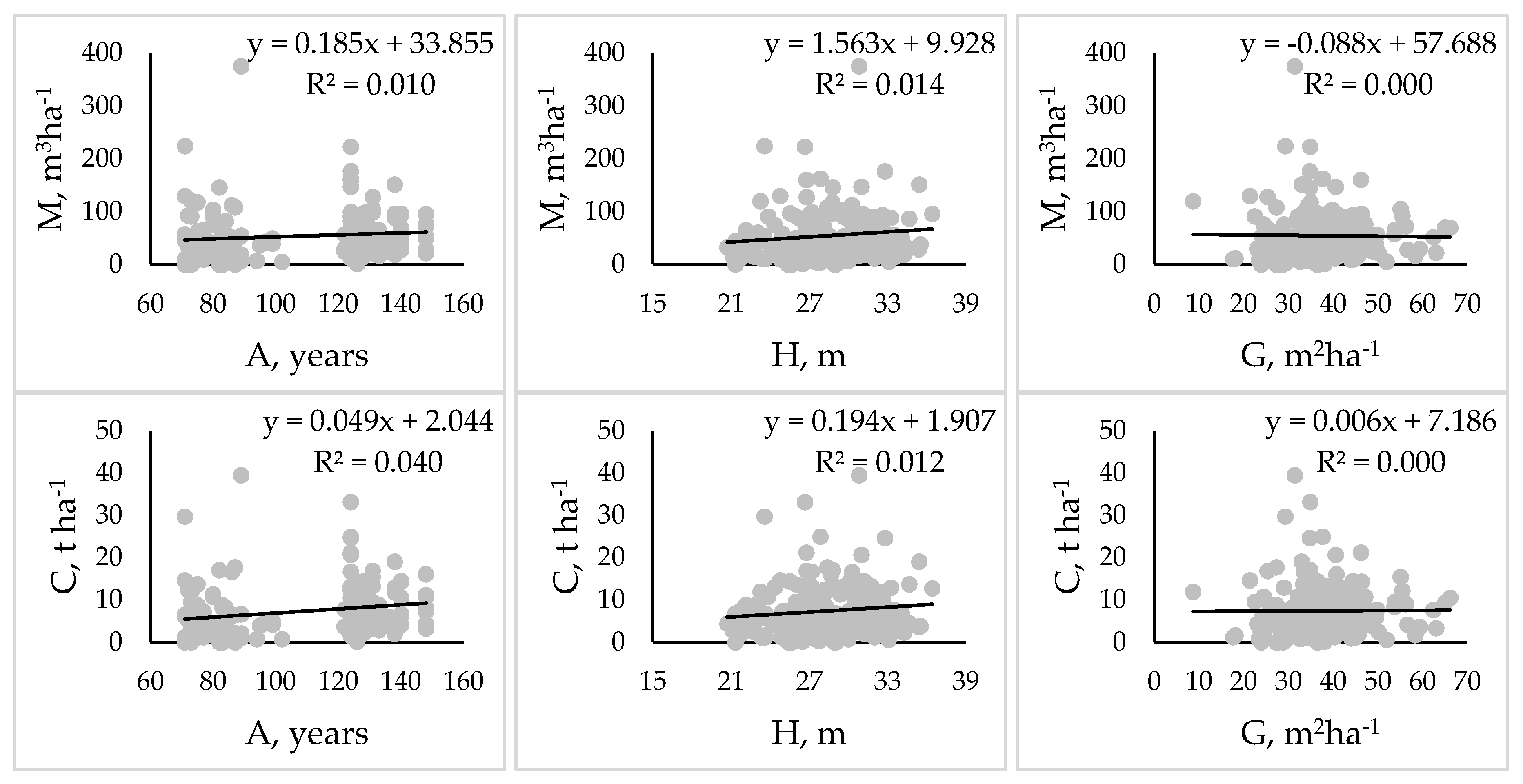
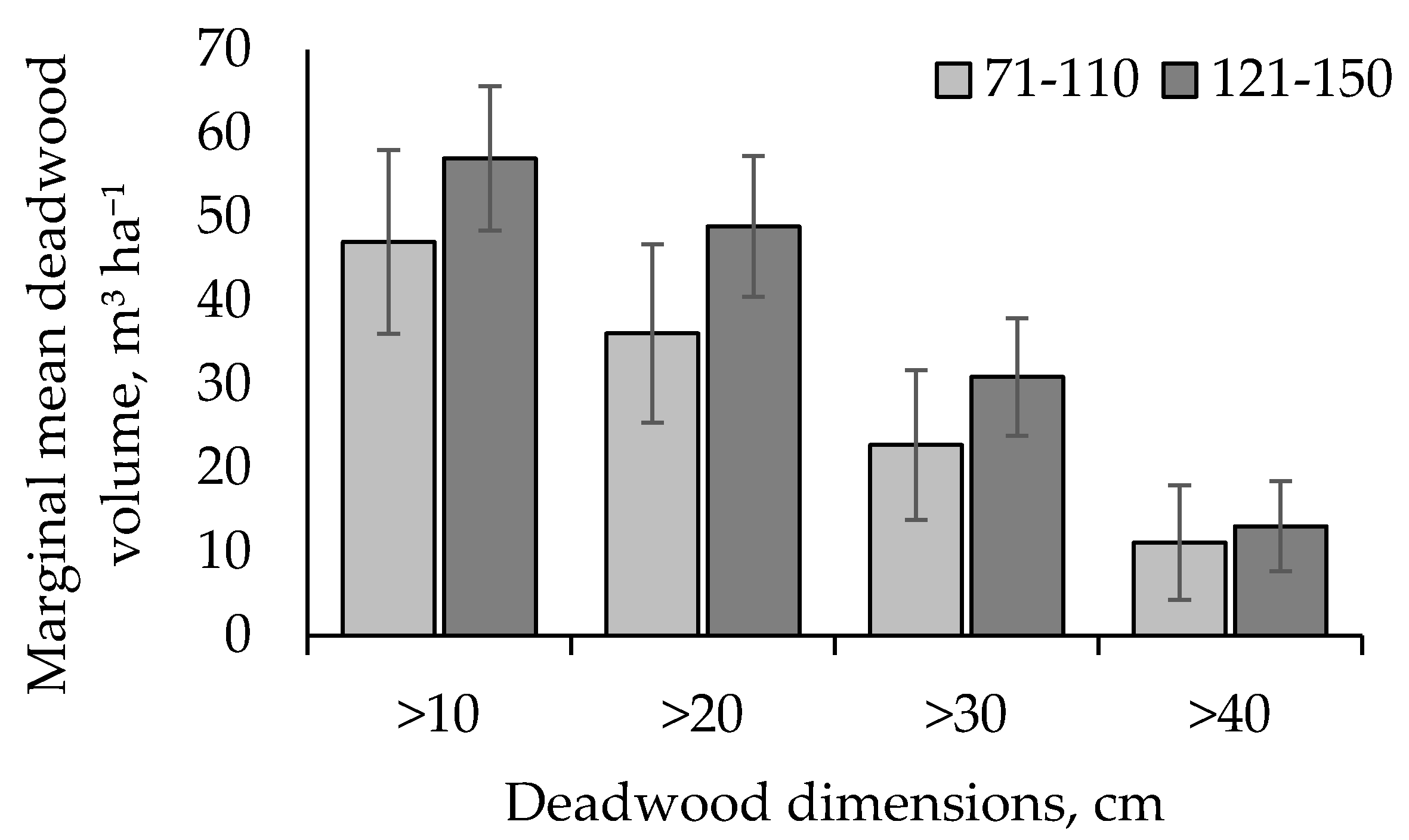
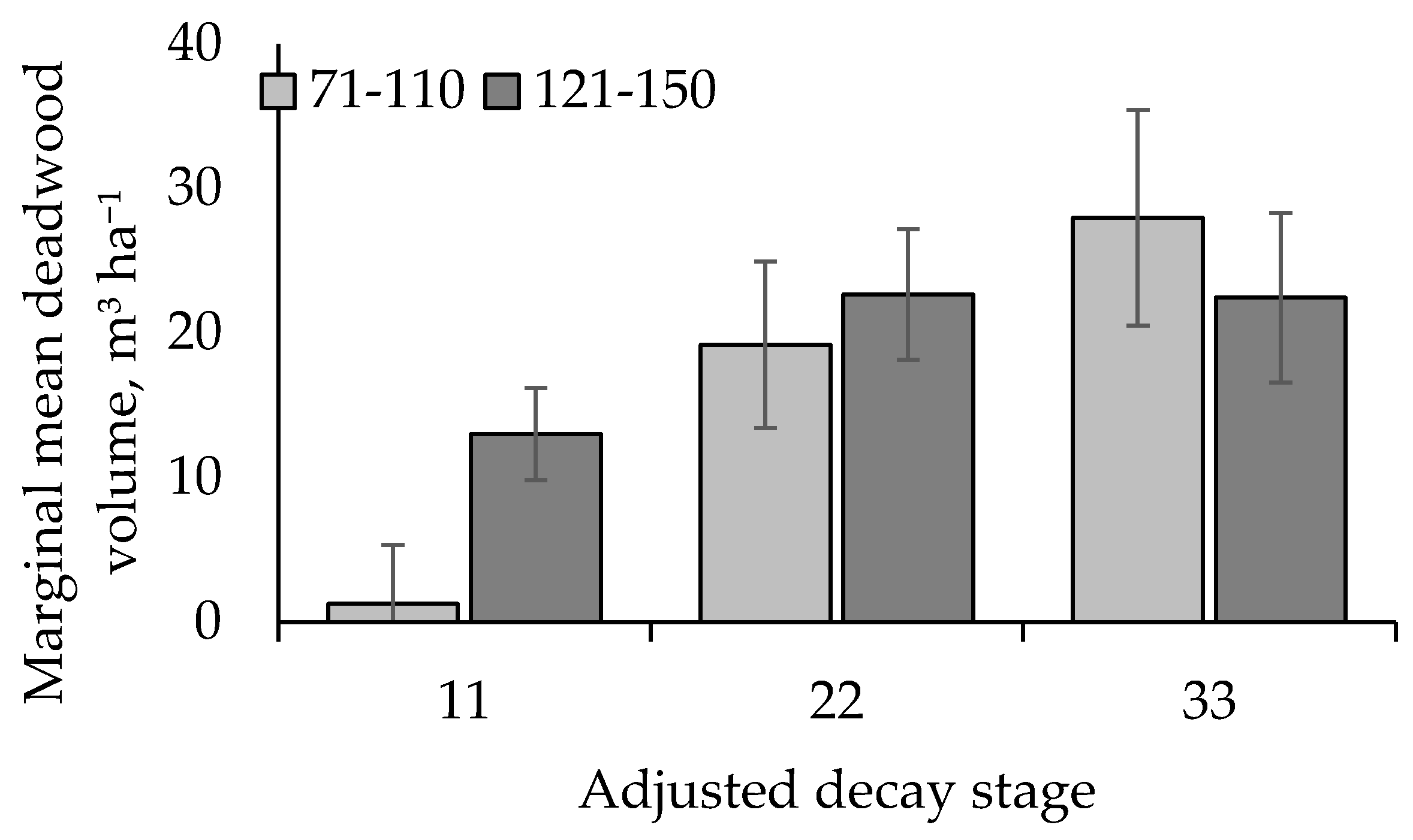
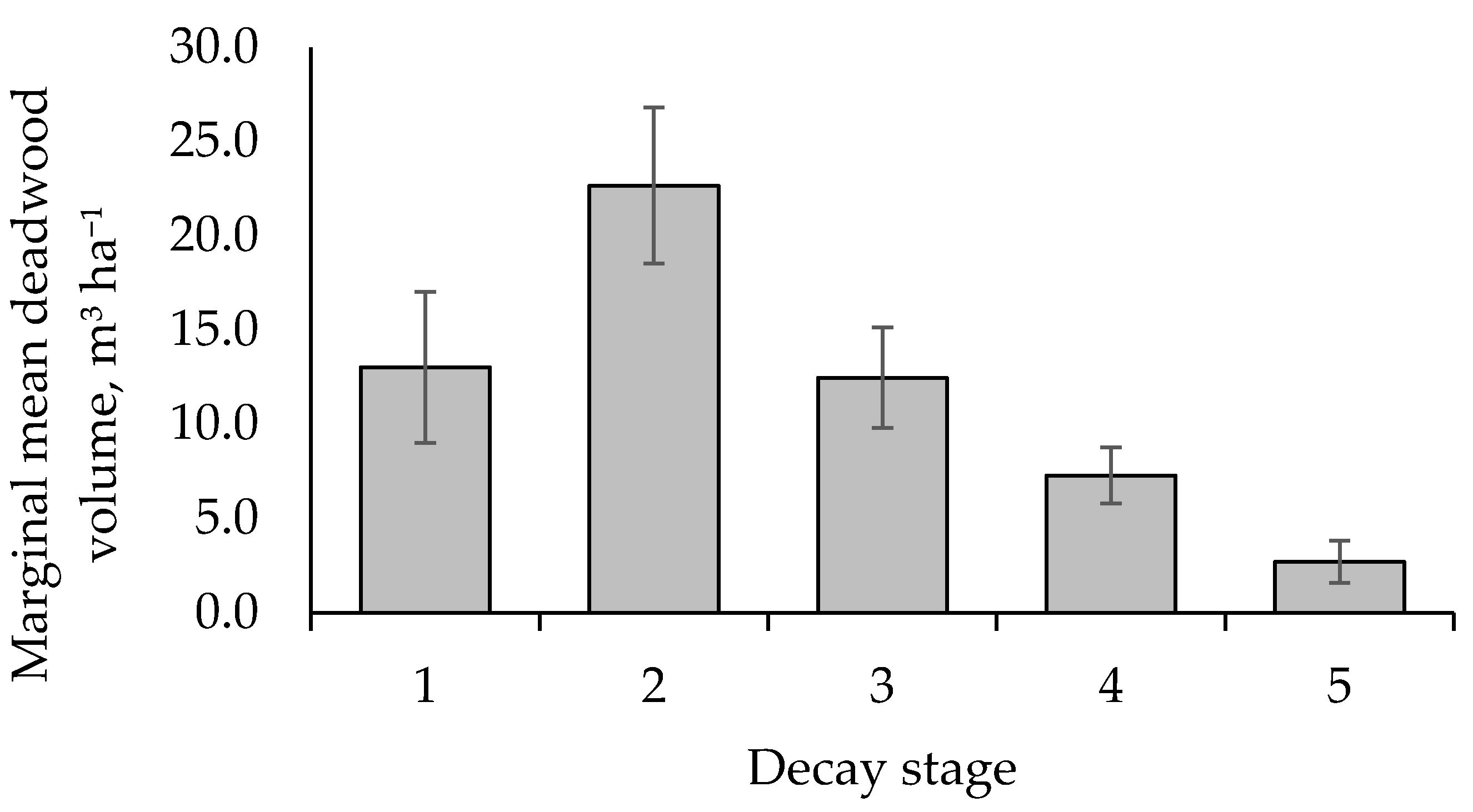
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hynynen, J.; Niemistö, P.; Viherä-Aarnio, A.; Brunner, A.; Hein, S.; Velling, P. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry 2010, 83, 103–119. [Google Scholar] [CrossRef]
- Felton, A.; Lindbladh, M.; Brunet, J.; Fritz, Ö. Replacing coniferous monocultures with mixed-species production stands: An assessment of the potential benefits for forest biodiversity in northern Europe. For. Ecol. Manag. 2010, 260, 939–947. [Google Scholar] [CrossRef]
- Renou-Wilson, F.; Pöllänen, M.; Byrne, K.; Wilson, D.; Farrell, E.P. The potential of birch afforestation as an after-use option for industrial cutaway peatlands. Suo 2010, 61, 59–76. [Google Scholar]
- Jõgiste, K.; Vares, A.; Sendrós, M. Restoration of former agricultural fields in Estonia: Comparative growth of planted and naturally regenerated birch. Forestry 2003, 76, 209–219. [Google Scholar] [CrossRef]
- Rudawska, M.; Kujawska, M.; Leski, T.; Janowski, D.; Karliński, L.; Wilgan, R. Ectomycorrhizal community structure of the admixture tree species Betula pendula, Carpinus betulus, and Tilia cordata grown in bare-root forest nurseries. For. Ecol. Manag. 2019, 437, 113–125. [Google Scholar] [CrossRef]
- Barantal, S.; Castagneyrol, B.; Durka, W.; Iason, G.; Morath, S.; Koricheva, J. Contrasting effects of tree species and genetic diversity on the leaf-miner communities associated with silver birch. Oecologia 2019, 189, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Pastorelli, R.; Paletto, A.; Agnelli, A.E.; Lagomarsino, A.; De Meo, I. Microbial communities associated with decomposing deadwood of downy birch in a natural forest in Khibiny Mountains (Kola Peninsula, Russian Federation). For. Ecol. Manag. 2020, 455, 117643. [Google Scholar] [CrossRef]
- Hägglund, R.; Hjältén, J. Substrate specific restoration promotes saproxylic beetle diversity in boreal forest set-asides. For. Ecol. Manag. 2018, 425, 45–58. [Google Scholar] [CrossRef]
- Jonsell, M.; Nittérus, K.; Stighäll, K. Saproxylic beetles in natural and man-made deciduous high stumps retained for conservation. Biol. Conserv. 2004, 118, 163–173. [Google Scholar] [CrossRef]
- Lindhe, A.; Lindelöw, Å. Cut high stumps of spruce, birch, aspen and oak as breeding substrates for saproxylic beetles. For. Ecol. Manag. 2004, 203, 1–20. [Google Scholar] [CrossRef]
- Parisi, F.; Pioli, S.; Lombardi, F.; Fravolini, G.; Marchetti, M.; Tognetti, R. Linking deadwood traits with saproxylic invertebrates and fungi in European forests-a review. iForest 2018, 11, 423–436. [Google Scholar] [CrossRef] [Green Version]
- Grove, S.J. Saproxylic insect ecology and the sustainable management of forests. Annu. Rev. Ecol. Syst. 2002, 33, 1–23. [Google Scholar] [CrossRef]
- Heilmann-Clausen, J.; Christensen, M. Does size matter? On the importance of various dead wood fractions for fungal diversity in Danish beech forests. For. Ecol. Manag. 2004, 201, 105–117. [Google Scholar] [CrossRef]
- Siitonen, J.; Saaristo, L. Habitat requirements and conservation of Pytho kolwensis, a beetle species of old-growth boreal forest. Biol. Conserv. 2000, 94, 211–220. [Google Scholar] [CrossRef]
- Pouska, V.; Svoboda, M.; Lepšová, A. The diversity of wood-decaying fungi in relation to changing site conditions in an old-growth mountain spruce forest, Central Europe. Eur. J. For. Res. 2010, 129, 219–231. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Ekström, M.; Esseen, P.A.; Grafström, A.; Ståhl, G.; Westerlund, B. Dead wood availability in managed Swedish forests–Policy outcomes and implications for biodiversity. For. Ecol. Manag. 2016, 376, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Ranius, T.; Kindvall, O.; Kruys, N.; Jonsson, B.G. Modelling dead wood in Norway spruce stands subject to different management regimes. For. Ecol. Manag. 2003, 182, 13–29. [Google Scholar] [CrossRef]
- Karjalainen, L.; Kuuluvainen, T. Amount and diversity of coarse woody debris within a boreal forest landscape dominated by Pinus sylvestris in Vienansalo wilderness, eastern Fennoscandia. Silva Fenn. 2002, 36, 147–167. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, S.G.; Niklasson, M.; Hedin, J.; Aronsson, G.; Gutowski, J.M.; Linder, P.; Ljungberg, H.; Mikusinski, G.; Ranius, T. Densities of large living and dead trees in old-growth temperate and boreal forests. For. Ecol. Manag. 2002, 161, 189–204. [Google Scholar] [CrossRef]
- Yatskov, M.; Harmon, M.E.; Krankina, O.N. A chronosequence of wood decomposition in the boreal forests of Russia. Can. J. For. Res. 2003, 33, 1211–1226. [Google Scholar] [CrossRef]
- Fridman, J.; Walheim, M. Amount, structure, and dynamics of dead wood on managed forestland in Sweden. For. Ecol. Manag. 2000, 131, 23–36. [Google Scholar] [CrossRef]
- Siitonen, J.; Martikainen, P.; Punttila, P.; Rauh, J. Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For. Ecol. Manag. 2000, 128, 211–225. [Google Scholar] [CrossRef]
- Ekbom, B.; Schroeder, L.M.; Larsson, S. Stand specific occurrence of coarse woody debris in a managed boreal forest landscape in central Sweden. For. Ecol. Manag. 2006, 221, 2–12. [Google Scholar] [CrossRef]
- Goers, L.; Ashton, M.S.; Tyrrell, M.L. Introduction. In Managing Forest Carbon in a Changing Climate; Ashton, M.S., Tyrrell, M.L., Spalding, D., Gentry, B., Eds.; Springer: Amsterdam, The Netherlands, 2012; pp. 1–4. [Google Scholar]
- Grassi, G.; House, J.; Dentener, F.; Federici, S.; den Elzen, M.; Penman, J. The key role of forests in meeting climate targets requires science for credible mitigation. Nat. Clim. Chang. 2017, 7, 220–226. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Gora, E.M.; Sayer, E.J.; Turner, B.L.; Tanner, E.V. Decomposition of coarse woody debris in a long-term litter manipulation experiment: A focus on nutrient availability. Funct. Ecol. 2018, 32, 1128–1138. [Google Scholar] [CrossRef] [Green Version]
- Carey, E.V.; Sala, A.; Keane, R.; Callaway, R.M. Are old forests underestimated as global carbon sinks? Glob. Chang. Biol. 2001, 7, 339–344. [Google Scholar] [CrossRef]
- Röser, C.; Montagnani, L.; Schulze, E.D.; Mollicone, D.; Kolle, O.; Meroni, M.; Papale, D.; Marchesini, L.B.; Federici, S.; Valentini, R. Net CO2 exchange rates in three different successional stages of the “Dark Taiga” of central Siberia. Tellus B 2002, 54, 642–654. [Google Scholar]
- Wardle, D.A.; Hörnberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar] [CrossRef]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Seedre, M.; Kopáček, J.; Janda, P.; Bače, R.; Svoboda, M. Carbon pools in a montane old-growth Norway spruce ecosystem in Bohemian Forest: Effects of stand age and elevation. For. Ecol. Manag. 2015, 346, 106–113. [Google Scholar] [CrossRef]
- Pukkala, T. Does management improve the carbon balance of forestry? Forestry 2017, 90, 125–135. [Google Scholar] [CrossRef]
- Nord-Larsen, T.; Vesterdal, L.; Bentsen, N.S.; Larsen, J.B. Ecosystem carbon stocks and their temporal resilience in a semi-natural beech-dominated forest. For. Ecol. Manag. 2019, 447, 67–76. [Google Scholar] [CrossRef]
- Hadden, D.; Grelle, A. Net CO2 emissions from a primary boreo-nemoral forest over a 10 year period. For. Ecol. Manag. 2017, 398, 164–173. [Google Scholar] [CrossRef]
- Uri, V.; Varik, M.; Aosaar, J.; Kanal, A.; Kukumägi, M.; Lõhmus, K. Biomass production and carbon sequestration in a fertile silver birch (Betula pendula Roth) forest chronosequence. For. Ecol. Manag. 2012, 267, 117–126. [Google Scholar] [CrossRef]
- Varik, M.; Aosaar, J.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon and nitrogen accumulation in belowground tree biomass in a chronosequence of silver birch stands. For. Ecol. Manag. 2013, 302, 62–70. [Google Scholar] [CrossRef]
- Varik, M.; Kukumägi, M.; Aosaar, J.; Becker, H.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon budgets in fertile silver birch (Betula pendula Roth) chronosequence stands. Ecol. Eng. 2015, 77, 284–296. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Morozov, G.; Karoles, K. Ecosystems carbon budgets of differently aged downy birch stands growing on well-drained peatlands. For. Ecol. Manag. 2017, 399, 82–93. [Google Scholar] [CrossRef]
- EEA (European Environment Agency). European Forest Types. Categories and Types for Sustainable Forest Management Reporting and Policy; Technical Report; European Environment Agency: Copenhagen, Denmark, 2007. [Google Scholar]
- Buchwald, E. A hierarchical terminology for more or less natural forests in relation to sustainable management and biodiversity conservation. In Proceedings of the Third Expert Meeting on Harmonizing Forest-related Definitions, Rome, Italy, 11–19 January 2005. [Google Scholar]
- Bušs, K. Latvijas PSR meža tipoloģijas pamati; LRZTIPI: Rīga, Latvia, 1976. [Google Scholar]
- Mäkinen, H.; Hynynen, J.; Siitonen, J.; Sievänen, R. Predicting the decomposition of Scots pine, Norway spruce, and birch stems in Finland. Ecol. Appl. 2006, 16, 1865–1879. [Google Scholar] [CrossRef]
- Jansons, J.; Līcīte, I. Latvia. In National Forest Inventories: Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 341–350. [Google Scholar]
- Liepa, I. Pieauguma mācība; LLU: Jelgava, Latvia, 1996. [Google Scholar]
- Köster, K.; Metslaid, M.; Engelhart, J.; Köster, E. Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests. For. Ecol. Manag. 2015, 354, 35–42. [Google Scholar] [CrossRef]
- Woodall, C.W.; Westfall, J.A. Relationships between the stocking levels of live trees and dead tree attributes in forests of the United States. For. Ecol. Manag. 2009, 258, 2602–2608. [Google Scholar] [CrossRef]
- Christensen, M.; Hahn, K.; Mountford, E.P.; Odor, P.; Standovár, T.; Rozenbergar, D.; Diaci, J.; Wijdeven, S.; Meyer, P.; Winter, S.; et al. Dead wood in European beech (Fagus sylvatica) forest reserves. For. Ecol. Manag. 2005, 210, 267–282. [Google Scholar] [CrossRef]
- Pedlar, J.H.; Pearce, J.L.; Venier, L.A.; McKenney, D.W. Coarse woody debris in relation to disturbance and forest type in boreal Canada. For. Ecol. Manag. 2002, 158, 189–194. [Google Scholar] [CrossRef]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood: A baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Debeljak, M. Coarse woody debris in virgin and managed forest. Ecol. Indic. 2006, 6, 733–742. [Google Scholar] [CrossRef]
- Dieler, J.; Uhl, E.; Biber, P.; Müller, J.; Rötzer, T.; Pretzsch, H. Effect of forest stand management on species composition, structural diversity, and productivity in the temperate zone of Europe. Eur. J. For. Res. 2017, 136, 739–766. [Google Scholar] [CrossRef]
- Liepa, L.; Straupe, I.; Miezīte, O.; Jansons, Ā. Structural diversity of dead wood in small-scaled protected forest parcels in Latvia. Res. Rural. Dev. 2019, 1, 12–17. [Google Scholar]
- Adermann, V. Estonia. In National Forest Inventories: Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 171–184. [Google Scholar]
- Parviainen, J.; Västilä, S. State of Finland’s Forests 2011. Based on the Criteria and Indicators of Sustainable Forest Management; Ministry of Agriculture and Forestry: Helsinki, Finland, 2011. [Google Scholar]
- Similä, M.; Kouki, J.; Martikainen, P. Saproxylic beetles in managed and seminatural Scots pine forests: Quality of dead wood matters. For. Ecol. Manag. 2003, 174, 365–381. [Google Scholar] [CrossRef]
- Köster, K.; Jõgiste, K.; Tukia, H.; Niklasson, M.; Möls, T. Variation and ecological characteristics of coarse woody debris in Lahemaa and Karula National Parks, Estonia. Scand. J. For. Res. 2005, 20, 102–111. [Google Scholar] [CrossRef]
- Lõhmus, A.; Kraut, A. Stand structure of hemiboreal old-growth forests: Characteristic features, variation among site types, and a comparison with FSC-certified mature stands in Estonia. For. Ecol. Manag. 2010, 260, 155–165. [Google Scholar] [CrossRef]
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar]
- Framstad, E.; de Wit, H.; Mäkipää, R.; Larjavaar, M.; Vesterdal, L.; Karltun, E. Biodiversity, Carbon Storage and Dynamics of Old Northern Forests; Nordic Council of Ministers: Copenhagen, Denmark, 2013. [Google Scholar]
- Johnson, E.A.; Miyanishi, K. Testing the assumptions of chronosequences in succession. Ecol. Lett. 2008, 11, 419–431. [Google Scholar] [CrossRef] [PubMed]
- Lindhe, A.; Åsenblad, N.; Toresson, H.G. Cut logs and high stumps of spruce, birch, aspen and oak–nine years of saproxylic fungi succession. Biol. Conserv. 2004, 119, 443–454. [Google Scholar] [CrossRef]
- Hottola, J.; Ovaskainen, O.; Hanski, I. A unified measure of the number, volume and diversity of dead trees and the response of fungal communities. J. Ecol. 2009, 97, 1320–1328. [Google Scholar] [CrossRef]
- Hein, S.; Winterhalter, D.; Wilhelm, G.J.; Kohnle, U. Wertholzproduktion mit der Sandbirke (Betula pendula Roth): Waldbauliche Möglichkeiten und Grenzen. Allg. Forst. Jagdztg. 2009, 180, 206–219. [Google Scholar]
- Zviedre, A.; Mangalis, I. Bērzs. In Meža Enciklopēdija; Broks, J., Ed.; Apgāds Zelta Grauds: Riga, Latvia, 2003. [Google Scholar]
- Ali, A.; Yan, E.R. Relationships between biodiversity and carbon stocks in forest ecosystems: A systematic literature review. Trop. Ecol. 2017, 58, 1–14. [Google Scholar]
- Huston, M.A.; Marland, G. Carbon management and biodiversity. J. Environ. Manag. 2003, 67, 77–86. [Google Scholar] [CrossRef]
- Schmid, A.V.; Vogel, C.S.; Liebman, E.; Curtis, P.S.; Gough, C.M. Coarse woody debris and the carbon balance of a moderately disturbed forest. For. Ecol. Manag. 2016, 361, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Harmon, M.E.; Ferrell, W.K.; Franklin, J.F. Effects on carbon storage of conversion of old-growth forests to young forests. Science 1990, 247, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.R.; Seedre, M.; Brassard, B.W.; Chen, H.Y. Decline in net ecosystem productivity following canopy transition to late-succession forests. Ecosystems 2014, 17, 778–791. [Google Scholar] [CrossRef]
- Ķēniņa, L.; Elferts, D.; Bāders, E.; Jansons, Ā. Carbon pools in a hemiboreal over-mature Norway spruce stands. Forests 2018, 9, 435. [Google Scholar] [CrossRef] [Green Version]
- Nabuurs, G.J.; Lindner, M.; Verkerk, P.J.; Gunia, K.; Deda, P.; Michalak, R.; Grassi, G. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Chang. 2013, 3, 792–796. [Google Scholar] [CrossRef]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Chang. Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Harmon, M.E.; Bond-Lamberty, B.; Tang, J.; Vargas, R. Heterotrophic respiration in disturbed forests: A review with examples from North America. J. Geophys. Res. Biogeosci. 2011, 116, G00K04. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Deadwood Characteristics | Deadwood Position | Stand Type | Marginal Mean | Standard Error | 95% Confidence Interval | |
|---|---|---|---|---|---|---|
| Min | Max | |||||
| Deadwood volume, m3·ha−1 | Standing | all mature | 13.8 | 2.4 | 9.2 | 18.5 |
| unmanaged mature | 16.3 | 2.7 | 11.1 | 21.6 | ||
| old-growth | 17.2 | 1.7 | 13.8 | 20.6 | ||
| Lying | all mature | 29.7 | 5.5 | 18.8 | 40.5 | |
| unmanaged mature | 34.9 | 6.2 | 22.6 | 47.2 | ||
| old-growth | 37.2 | 4.1 | 29.2 | 45.2 | ||
| Total | all mature | 43.5 | 6.4 | 31.0 | 56.0 | |
| unmanaged mature | 51.3 | 7.1 | 37.3 | 65.3 | ||
| old-growth | 54.4 | 4.6 | 45.3 | 63.5 | ||
| Deadwood carbon pool, t·ha−1 | Standing | all mature | 1.9 | 0.3 | 1.2 | 3.0 |
| unmanaged mature | 2.2 | 0.4 | 1.4 | 2.9 | ||
| old-growth | 2.6 | 0.3 | 2.1 | 3.0 | ||
| Lying | all mature | 3.5 | 0.6 | 2.1 | 4.8 | |
| unmanaged mature | 4.1 | 0.8 | 2.6 | 5.6 | ||
| old-growth | 5.3 | 0.5 | 4.3 | 6.3 | ||
| Total | all mature | 5.4 | 0.8 | 3.8 | 7.0 | |
| unmanaged mature | 6.3 | 0.9 | 4.5 | 8.1 | ||
| old-growth | 7.9 | 0.6 | 6.7 | 9.0 | ||
| Stand Age, Years | Species | Proportion of Deadwood, % | Standard Error | 95% Confidence Interval | |
|---|---|---|---|---|---|
| Min | Max | ||||
| 71–110 | Birch | 35.8 | 4.0 | 28.8 | 44.6 |
| Other deciduous | 25.4 | 3.1 | 19.9 | 32.3 | |
| Coniferous | 38.8 | 4.2 | 31.5 | 48.1 | |
| 121–150 | Birch | 37.9 | 3.0 | 31.9 | 43.9 |
| Other deciduous | 9.1 | 2.4 | 4.4 | 13.8 | |
| Coniferous | 53.0 | 3.2 | 46.7 | 59.2 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šēnhofa, S.; Jaunslaviete, I.; Šņepsts, G.; Jansons, J.; Liepa, L.; Jansons, Ā. Deadwood Characteristics in Mature and Old-Growth Birch Stands and Their Implications for Carbon Storage. Forests 2020, 11, 536. https://doi.org/10.3390/f11050536
Šēnhofa S, Jaunslaviete I, Šņepsts G, Jansons J, Liepa L, Jansons Ā. Deadwood Characteristics in Mature and Old-Growth Birch Stands and Their Implications for Carbon Storage. Forests. 2020; 11(5):536. https://doi.org/10.3390/f11050536
Chicago/Turabian StyleŠēnhofa, Silva, Ieva Jaunslaviete, Guntars Šņepsts, Jurģis Jansons, Līga Liepa, and Āris Jansons. 2020. "Deadwood Characteristics in Mature and Old-Growth Birch Stands and Their Implications for Carbon Storage" Forests 11, no. 5: 536. https://doi.org/10.3390/f11050536