Availability and Structure of Coarse Woody Debris in Hemiboreal Mature to Old-Growth Aspen Stands and Its Implications for Forest Carbon Pool

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
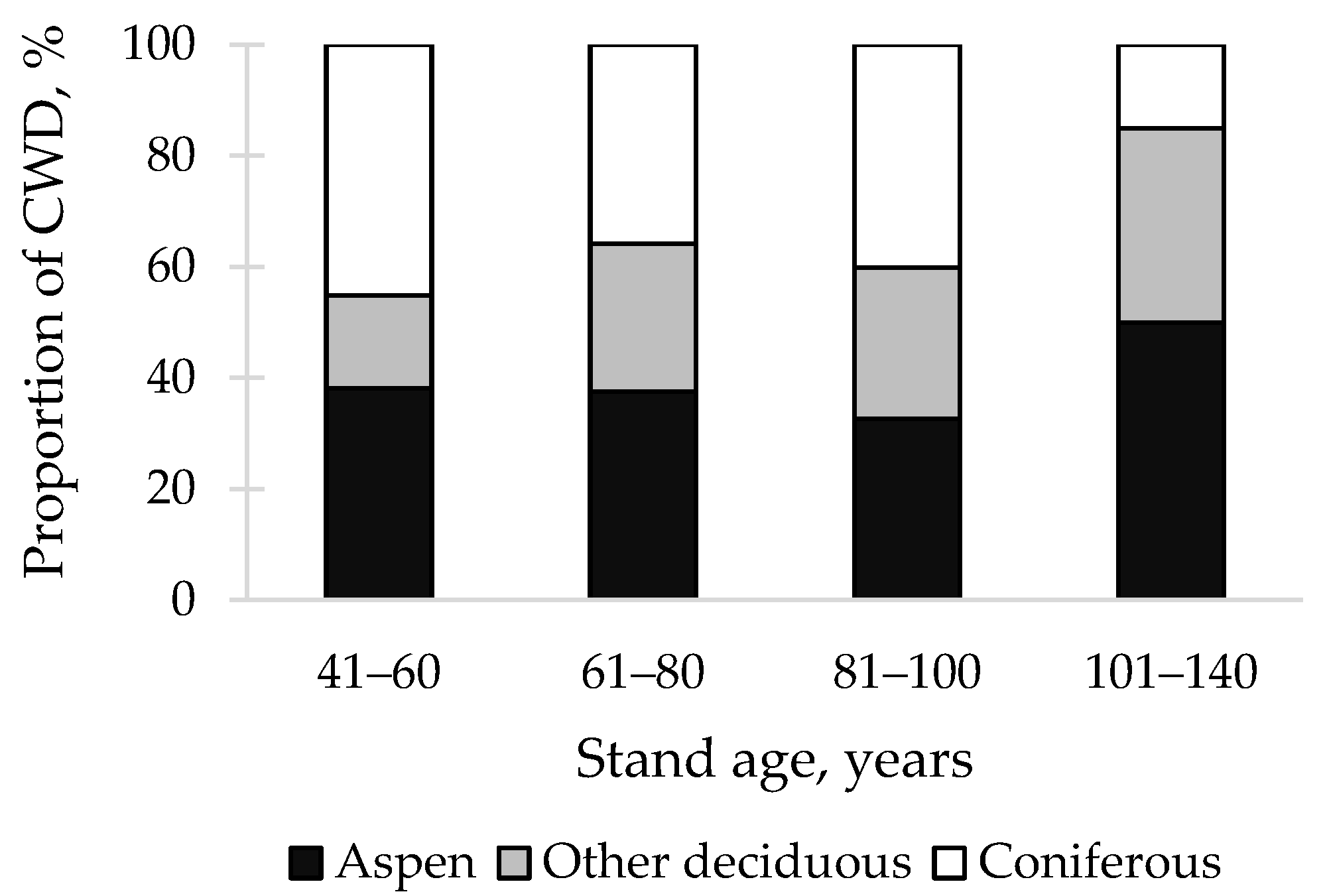
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Caudullo, G.; de Rigo, D. Populus tremula in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 138–139. [Google Scholar]
- Heräjärvi, H.; Junkkonen, R. Wood density and growth rate of European and hybrid aspen in Southern Finland. Balt. For. 2006, 12, 2–8. [Google Scholar]
- Global Forest Resources Assessment. Country Report, Sweden; Global Forest Resources Assessment: Rome, Italy, 2015. [Google Scholar]
- Rogers, P.C.; Pinno, B.D.; Šebesta, J.; Albrectsen, B.R.; Li, G.; Ivanova, N.; Kusbach, A.; Kuuluvainen, T.; Landhäusser, S.M.; Liu, H.; et al. A global view of aspen: Conservation science for widespread keystone systems. Glob. Ecol. Conserv. 2020, 21, e00828. [Google Scholar] [CrossRef]
- Sverdrup-Thygeson, A.; Ims, R.A. The effect of forest clearcutting in Norway on the community of saproxylic beetles on aspen. Biol. Conserv. 2002, 106, 347–357. [Google Scholar] [CrossRef]
- Kouki, J.; Arnold, K.; Martikainen, P. Long-term persistence of aspen—A key host for many threatened species—Is endangered in old-growth conservation areas in Finland. J. Nat. Conserv. 2004, 12, 41–52. [Google Scholar] [CrossRef]
- Kivinen, S.; Koivisto, E.; Keski-Saari, S.; Poikolainen, L.; Tanhuanpää, T.; Kuzmin, A.; Viinikka, A.; Heikkinen, R.K.; Pykälä, J.; Virkkala, R.; et al. A keystone species, European aspen (Populus tremula L.), in boreal forests: Ecological role, knowledge needs and mapping using remote sensing. For. Ecol. Manag. 2020, 462, 118008. [Google Scholar] [CrossRef]
- Siitonen, J.; Martikainen, P.; Punttila, P.; Rauh, J. Coarse woody debris and stand characteristics in mature managed and old-growth boreal mesic forests in southern Finland. For. Ecol. Manag. 2000, 128, 211–225. [Google Scholar] [CrossRef]
- Latva-Karjanmaa, T.; Penttilä, R.; Siitonen, J. The demographic structure of European aspen (Populus tremula) populations in managed and old-growth boreal forests in eastern Finland. Can. J. For. Res. 2007, 37, 1070–1081. [Google Scholar] [CrossRef]
- Shorohova, E.; Kapitsa, E. Influence of the substrate and ecosystem attributes on the decomposition rates of coarse woody debris in European boreal forests. For. Ecol. Manag. 2014, 315, 173–184. [Google Scholar] [CrossRef]
- Halme, P.; Purhonen, J.; Marjakangas, E.L.; Komonen, A.; Juutilainen, K.; Abrego, N. Dead wood profile of a semi-natural boreal forest-implications for sampling. Silva Fenn. 2019, 53, 10010. [Google Scholar] [CrossRef] [Green Version]
- Laarmann, D.; Korjus, H.; Sims, A.; Stanturf, J.A.; Kiviste, A.; Köster, K. Analysis of forest naturalness and tree mortality patterns in Estonia. For. Ecol. Manag. 2009, 258, S187–S195. [Google Scholar] [CrossRef]
- Doerfler, I.; Gossner, M.M.; Müller, J.; Seibold, S.; Weisser, W.W. Deadwood enrichment combining integrative and segregative conservation elements enhances biodiversity of multiple taxa in managed forests. Biol. Conserv. 2018, 228, 70–78. [Google Scholar] [CrossRef]
- Koivula, M.; Vanha-Majamaa, I. Experimental evidence on biodiversity impacts of variable retention forestry, prescribed burning, and deadwood manipulation in Fennoscandia. Ecol. Process. 2020, 9, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Covey, K.R.; Bueno de Mesquita, C.; Oberle, B.; Maynard, D.S.; Bettigole, C.; Crowther, T.W.; Duguid, M.C.; Steven, B.; Zanne, A.E.; Lapin, M.; et al. Greenhouse trace gases in deadwood. Biogeochemistry 2016, 130, 215–226. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Ranius, T.; Kindvall, O.; Kruys, N.; Jonsson, B.G. Modelling dead wood in Norway spruce stands subject to different management regimes. For. Ecol. Manag. 2003, 182, 13–29. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Sass-Klaassen, U.; Poorter, L.; Van Geffen, K.; Van Logtestijn, R.S.P.; Van Hal, J.; Goudzwaard, L.; Sterck, F.J.; Klaassen, R.K.W.M.; Freschet, G.T.; et al. Controls on coarse wood decay in temperate tree species: Birth of the LOGLIFE experiment. Ambio 2012, 41, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Garbarino, M.; Marzano, R.; Shaw, J.D.; Long, J.N. Environmental drivers of deadwood dynamics in woodlands and forests. Ecosphere 2015, 6, 30. [Google Scholar] [CrossRef]
- Martin, A.; Dimke, G.; Doraisami, M.; Thomas, S. Carbon fractions in the world’s dead wood. Nat. Commun. 2021, 12, 889. [Google Scholar] [CrossRef]
- Carey, E.V.; Sala, A.; Keane, R.; Callaway, R.M. Are old forests underestimated as global carbon sinks? Glob. Chang. Biol. 2001, 7, 339–344. [Google Scholar] [CrossRef]
- Röser, C.; Montagnani, L.; Schulze, E.-D.; Mollicone, D.; Kolle, O.; Meroni, M.; Papale, D.; Marchesini, L.B.; Federici, S.; Valentini, R. Net CO₂ exchange rates in three different successional stages of the “Dark Taiga” of central Siberia. Tellus B Chem. Phys. Meteorol. 2002, 54, 642–654. [Google Scholar] [CrossRef]
- Wardle, D.A.; Hörnberg, G.; Zackrisson, O.; Kalela-Brundin, M.; Coomes, D.A. Long-term effects of wildfire on ecosystem properties across an island area gradient. Science 2003, 300, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Luyssaert, S.; Schulze, E.D.; Börner, A.; Knohl, A.; Hessenmöller, D.; Law, B.E.; Ciais, P.; Grace, J. Old-growth forests as global carbon sinks. Nature 2008, 455, 213–215. [Google Scholar] [CrossRef]
- Seedre, M.; Kopáček, J.; Janda, P.; Bače, R.; Svoboda, M. Carbon pools in a montane old-growth Norway spruce ecosystem in Bohemian Forest: Effects of stand age and elevation. For. Ecol. Manag. 2015, 346, 106–113. [Google Scholar] [CrossRef]
- Hadden, D.; Grelle, A. Net CO₂ emissions from a primary boreo-nemoral forest over a 10 year period. For. Ecol. Manag. 2017, 398, 164–173. [Google Scholar] [CrossRef]
- Pukkala, T. Does management improve the carbon balance of forestry? For. Int. J. For. Res. 2017, 90, 125–135. [Google Scholar] [CrossRef]
- Nord-Larsen, T.; Vesterdal, L.; Bentsen, N.S.; Larsen, J.B. Ecosystem carbon stocks and their temporal resilience in a semi-natural beech-dominated forest. For. Ecol. Manag. 2019, 447, 67–76. [Google Scholar] [CrossRef]
- Uri, V.; Varik, M.; Aosaar, J.; Kanal, A.; Kukumägi, M.; Lõhmus, K. Biomass production and carbon sequestration in a fertile silver birch (Betula pendula Roth) forest chronosequence. For. Ecol. Manag. 2012, 267, 117–126. [Google Scholar] [CrossRef]
- Uri, V.; Kukumägi, M.; Aosaar, J.; Varik, M.; Becker, H.; Morozov, G.; Karoles, K. Ecosystems carbon budgets of differently aged downy birch stands growing on well-drained peatlands. For. Ecol. Manag. 2017, 399, 82–93. [Google Scholar] [CrossRef]
- Varik, M.; Aosaar, J.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon and nitrogen accumulation in belowground tree biomass in a chronosequence of silver birch stands. For. Ecol. Manag. 2013, 302, 62–70. [Google Scholar] [CrossRef]
- Varik, M.; Kukumägi, M.; Aosaar, J.; Becker, H.; Ostonen, I.; Lõhmus, K.; Uri, V. Carbon budgets in fertile silver birch (Betula pendula Roth) chronosequence stands. Ecol. Eng. 2015, 77, 284–296. [Google Scholar] [CrossRef]
- Šēnhofa, S.; Jaunslaviete, I.; Šņepsts, G.; Jansons, J.; Liepa, L.; Jansons, A. Deadwood characteristics in mature and old-growth birch stands and their implications for carbon storage. Forests 2020, 11, 536. [Google Scholar] [CrossRef]
- Gao, B.; Taylor, A.R.; Searle, E.B.; Kumar, P.; Ma, Z.; Hume, A.M.; Chen, H.Y.H. Carbon Storage Declines in Old Boreal Forests Irrespective of Succession Pathway. Ecosystems 2017, 21, 1168–1182. [Google Scholar] [CrossRef]
- European Environment Agency. European Forest Types. Categories and Types for Sustainable Forest Management Reporting and Policy; Technical Report; European Environment Agency: Copenhagen, Denmark, 2007. [Google Scholar]
- Bušs, K. Forest ecosystem classification in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 1997, 51, 204–218. [Google Scholar]
- Buchwald, E. A hierarchical terminology for more or less natural forests in relation to sustainable management and biodiversity conservation. In Proceedings of the Proceedings of the Third Expert Meeting on Harmonizing Forest-related Definitions, Rome, Italy, 11–19 January 2005. [Google Scholar]
- Mäkinen, H.; Hynynen, J.; Siitonen, J.; Sievänen, R. Predicting the decomposition of scots pine, norway spruce, and birch stems in Finland. Ecol. Appl. 2006, 16, 1865–1879. [Google Scholar] [CrossRef]
- Köster, K.; Metslaid, M.; Engelhart, J.; Köster, E. Dead wood basic density, and the concentration of carbon and nitrogen for main tree species in managed hemiboreal forests. For. Ecol. Manag. 2015, 354, 35–42. [Google Scholar] [CrossRef]
- Jansons, J.; Līcīte, I. Latvia. In National Forest Inventories: Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 341–350. [Google Scholar]
- Liepa, I. Pieauguma mācība; LLU: Jelgava, Latvia, 1996. [Google Scholar]
- Bobiec, A. The influence of gaps on tree regeneration: A case study of the mixed lime-hornbeam (Tilio-Carpinetum Tracz. 1962) communities in the Białowieża Primeval Forest. Polish J. Ecol. 2007, 55, 441–455. [Google Scholar]
- Jonášová, M.; Matějková, I. Natural regeneration and vegetation changes in wet spruce forests after natural and artificial disturbances. Can. J. For. Res. 2007, 37, 1907–1914. [Google Scholar] [CrossRef]
- Vehmas, M.; Kouki, J.; Eerikainen, K. Long-term spatio-temporal dynamics and historical continuity of European aspen (Populus tremula L.) stands in the Koli National Park, eastern Finland. Forestry 2009, 82, 135–148. [Google Scholar] [CrossRef] [Green Version]
- Kull, O.; Niinemets, Ü. Distribution of leaf photosynthetic properties in tree canopies: Comparison of species with different shade tolerance. Funct. Ecol. 1998, 12, 472–479. [Google Scholar] [CrossRef]
- Yatskov, M.; Harmon, M.E.; Krankina, O.N. A chronosequence of wood decomposition in the boreal forests of Russia. Can. J. For. Res. 2003, 33, 1211–1226. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Ekström, M.; Esseen, P.A.; Grafström, A.; Ståhl, G.; Westerlund, B. Dead wood availability in managed Swedish forests—Policy outcomes and implications for biodiversity. For. Ecol. Manag. 2016, 376, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Similä, M.; Kouki, J.; Martikainen, P. Saproxylic beetles in managed and seminatural Scots pine forests: Quality of dead wood matters. For. Ecol. Manag. 2003, 174, 365–381. [Google Scholar] [CrossRef]
- Adermann, V. Estonia. In National Forest Inventories: Pathways for Common Reporting; Tomppo, E., Gschwantner, T., Lawrence, M., McRoberts, R.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 171–184. [Google Scholar]
- Siitonen, J. Forest management, coarse woody debris and saproxylic organisms: Fennoscandian boreal forests as an example. Ecol. Bull. 2001, 49, 11–41. [Google Scholar] [CrossRef]
- Rouvinen, S.; Kuuluvainen, T.; Karjalainen, L. Coarse woody debris in old Pinus sylvestris dominated forests along a geographic and human impact gradient in boreal Fennoscandia. Can. J. For. Res. 2002, 32, 2184–2200. [Google Scholar] [CrossRef]
- Aakala, T. Coarse woody debris in late-successional Picea abies forests in northern Europe: Variability in quantities and models of decay class dynamics. For. Ecol. Manag. 2010, 260, 770–779. [Google Scholar] [CrossRef]
- Lee, P. Dynamics of snags in aspen-dominated midboreal forests. For. Ecol. Manag. 1998, 105, 263–272. [Google Scholar] [CrossRef]
- Vasiliauskas, R.; Vasiliauskas, A.; Stenlid, J.; Matelis, A. Dead trees and protected polypores in unmanaged north-temperate forest stands of Lithuania. For. Ecol. Manag. 2004, 193, 355–370. [Google Scholar] [CrossRef]
- Bujoczek, L.; Bujoczek, M.; Zięba, S. How much, why and where? Deadwood in forest ecosystems: The case of Poland. Ecol. Indic. 2021, 121, 107027. [Google Scholar] [CrossRef]
- Krankina, O.N.; Treyfeld, R.F.; Harmon, M.E.; Spycher, G.; Povarov, E.D. Coarse Woody Debris in the Forests of the St. Petersburg Region, Russia. Ecol. Bull. 2001, 93–107. [Google Scholar] [CrossRef]
- Lee, P.C.; Crites, S.; Nietfeld, M.; Nguyen, H.V.; Stelfox, J.B. Characteristics and origins of deadwood material in aspen-dominated boreal forests. Ecol. Appl. 1997, 7, 691–701. [Google Scholar] [CrossRef]
- Köster, K.; Jõgiste, K.; Tukia, H.; Niklasson, M.; Möls, T. Variation and ecological characteristics of coarse woody debris in Lahemaa and Karula National Parks, Estonia. Scand. J. For. Res. 2005, 20, 102–111. [Google Scholar] [CrossRef]
- Nilsson, S.G.; Niklasson, M.; Hedin, J.; Aronsson, G.; Gutowski, J.M.; Linder, P.; Ljungberg, H.; Mikusinski, G.; Ranius, T. Densities of large living and dead trees in old-growth temperate and boreal forests. For. Ecol. Manag. 2002, 161, 198–204. [Google Scholar] [CrossRef]
- Framstad, E.; de Wit, H.; Mäkipää, R.; Larjavaar, M.; Vesterdal, L.; Karltun, E. Biodiversity, Carbon Storage and Dynamics of Old Northern Forests; Nordic Council of Ministers: Copenhagen, Denmark, 2013. [Google Scholar]
- Chen, H.Y.H.; Popadiouk, R.V. Dynamics of North American boreal mixedwoods. Environ. Rev. 2002, 10, 137–166. [Google Scholar] [CrossRef]
- Liepiņa, L. Apses. In Meža Enciklopēdija; Broks, J., Ed.; Zelta Grauds: Rīga, Latvia, 2005. [Google Scholar]
- Lankia, H.; Wallenius, T.; Várkonyi, G.; Kouki, J.; Snäll, T. Forest fire history, aspen and goat willow in a Fennoscandian old-growth landscape: Are current population structures a legacy of historical fires? J. Veg. Sci. 2012, 23, 1159–1169. [Google Scholar] [CrossRef]
- Müller, J.; Bütler, R. A review of habitat thresholds for dead wood: A baseline for management recommendations in European forests. Eur. J. For. Res. 2010, 129, 981–992. [Google Scholar] [CrossRef]
- Brin, A.; Brustel, H.; Jactel, H. Species variables or environmental variables as indicators of forest biodiversity: A case study using saproxylic beetles in Maritime pine plantations. Ann. For. Sci. 2009, 66, 306. [Google Scholar] [CrossRef] [Green Version]
- Hottola, J.; Ovaskainen, O.; Hanski, I. A unified measure of the number, volume and diversity of dead trees and the response of fungal communities. J. Ecol. 2009, 97, 1320–1328. [Google Scholar] [CrossRef]
- Lassauce, A.; Paillet, Y.; Jactel, H.; Bouget, C. Deadwood as a surrogate for forest biodiversity: Meta-analysis of correlations between deadwood volume and species richness of saproxylic organisms. Ecol. Indic. 2011, 11, 1027–1039. [Google Scholar] [CrossRef]
- Hekkala, A.M.; Ahtikoski, A.; Päätalo, M.L.; Tarvainen, O.; Siipilehto, J.; Tolvanen, A. Restoring volume, diversity and continuity of deadwood in boreal forests. Biodivers. Conserv. 2016, 25, 1107–1132. [Google Scholar] [CrossRef]
- Liu, Q.; Hytteborn, H. Gap structure, disturbance and regeneration in a primeval Picea abies forest. J. Veg. Sci. 1991, 2, 391–402. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Allison, S.D.; Bauhus, J.; Eggleton, P.; Preston, C.M.; Scarff, F.; Weedon, J.T.; Wirth, C.; Zanne, A.E. Plant traits and wood fates across the globe: Rotted, burned, or consumed? Glob. Chang. Biol. 2009, 15, 2431–2449. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J.; Spatz, H.-C. Worldwide correlations of mechanical properties and green wood density. Am. J. Bot. 2010, 97, 1587–1594. [Google Scholar] [CrossRef]
- Jacobsen, R.M.; Sverdrup-Thygeson, A.; Kauserud, H.; Mundra, S.; Birkemoe, T. Exclusion of invertebrates influences saprotrophic fungal community and wood decay rate in an experimental field study. Funct. Ecol. 2018, 32, 2571–2582. [Google Scholar] [CrossRef]
- DeLong, S.C.; Daniels, L.D.; Heemskerk, B.; Storaunet, K.O. Temporal development of decaying log habitats in wet spruce-fir stands in east-central British Columbia. Can. J. For. Res. 2005, 35, 2841–2850. [Google Scholar] [CrossRef]
- Tarasov, M.E.; Birdsey, R.A. Decay rate and potential storage of coarse woody debris in the Leningrad region. Ecol. Bull. 2001, 137–147. [Google Scholar] [CrossRef]
- Jonsson, B.G. Availability of coarse woody debris in a boreal old-growth Picea abies forest. J. Veg. Sci. 2000, 11, 51–56. [Google Scholar] [CrossRef]
- Hararuk, O.; Kurz, W.A.; Didion, M. Dynamics of dead wood decay in Swiss forests. For. Ecosyst. 2020, 7, 36. [Google Scholar] [CrossRef]
- Moroni, M.T.; Morris, D.M.; Shaw, C.; Stokland, J.N.; Harmon, M.E.; Fenton, N.J.; Merganičová, K.; Merganič, J.; Okabe, K.; Hagemann, U. Buried Wood: A Common Yet Poorly Documented Form of Deadwood. Ecosystems 2015, 18, 605–628. [Google Scholar] [CrossRef]
- Tikkanen, O.-P.; Martikainen, P.; Hyvärinen, E.; Junninen, K.; Kouki, J. Red-listed boreal forest species of Finland: Associations with forest structure, tree species, and decaying wood. Ann. Zool. Fennici 2006, 43, 373–383. [Google Scholar] [CrossRef]
- Ranius, T.; Martikainen, P.; Kouki, J. Colonisation of ephemeral forest habitats by specialised species: Beetles and bugs associated with recently dead aspen wood. Biodivers. Conserv. 2011, 20, 2903–2915. [Google Scholar] [CrossRef] [Green Version]
- Shorohova, E.; Kushnevskaya, H.; Ruokolainen, A.; Polevoi, A.; Borovichev, E. Behavior in a wide range of choices: Substrate preferences of threatened wood-inhabiting species in a mixed old-growth boreal forest. In Proceedings of the ECCB2018 5th European Congress of Conservation Biology, Jyväskylä, Finland, 12–15 June 2018; Open Science Centre, University of Jyväskylä: Jyväskylä, Finland, 2018. [Google Scholar]
- Puletti, N.; Canullo, R.; Mattioli, W.; Gawryś, R.; Corona, P.; Czerepko, J. A dataset of forest volume deadwood estimates for Europe. Ann. For. Sci. 2019, 76, 1–8. [Google Scholar] [CrossRef]
- Priewasser, K.; Brang, P.; Bachofen, H.; Bugmann, H.; Wohlgemuth, T. Impacts of salvage-logging on the status of deadwood after windthrow in Swiss forests. Eur. J. For. Res. 2013, 132, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Kahl, T.; Baber, K.; Otto, P.; Wirth, C.; Bauhus, J. Drivers of CO2 Emission Rates from Dead Wood Logs of 13 Tree Species in the Initial Decomposition Phase. Forests 2015, 6, 2484–2504. [Google Scholar] [CrossRef] [Green Version]
- Błońska, E.; Lasota, J.; Tullus, A.; Lutter, R.; Ostonen, I. Impact of deadwood decomposition on soil organic carbon sequestration in Estonian and Polish forests. Ann. For. Sci. 2019, 76, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Seibold, S.; Bässler, C.; Brandl, R.; Büche, B.; Szallies, A.; Thorn, S.; Ulyshen, M.D.; Müller, J. Microclimate and habitat heterogeneity as the major drivers of beetle diversity in dead wood. J. Appl. Ecol. 2016, 53, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Ķēniņa, L.; Mača, S.; Jaunslaviete, I.; Jansons, Ā. Carbon pools in old-growth Scots pine stands on organic soils and its concentration in deadwood: Cases study in Latvia. In Proceedings of the 9th International Scientific Conference “Rural Development 2019”, Kaunas, Lithuania, 26–28 September; VDU: Kaunas, Lithuania, 2019; pp. 284–288. [Google Scholar]
- Stakėnas, V.; Varnagirytė-Kabašinskienė, I.; Sirgedaitė-Šėžienė, V.; Armolaitis, K.; Araminienė, V.; Muraškienė, M.; Žemaitis, P. Dead wood carbon density for the main tree species in the Lithuanian hemiboreal forest. Eur. J. For. Res. 2020, 139, 1045–1055. [Google Scholar] [CrossRef]
- Witt, C. Characteristics of aspen infected with heartrot: Implications for cavity-nesting birds. For. Ecol. Manag. 2010, 260, 1010–1016. [Google Scholar] [CrossRef]
- Lõhmus, A. Aspen-inhabiting aphyllophoroid fungi in a managed forest landscape in Estonia. Scand. J. For. Res. 2011, 26, 212–220. [Google Scholar] [CrossRef]
- Andelic, M.; Tangnaes, M.-J. The Effect of Secondary Metabolites, Nutrients and Invertebrates on Fungal Establishment and Decomposition Rates in European Aspen (Populus tremula). Master’s Thesis, Norwegian University of Life Sciences, Ås, Norway, 2019. [Google Scholar]
- Wirth, C.; Lichstein, J.W. The Imprint of Species Turnover on Old-Growth Forest Carbon Balances—Insights From a Trait-Based Model of Forest Dynamics; Springer: Berlin/Heidelberg, Germany, 2009; pp. 81–113. [Google Scholar]
- Schmid, A.V.; Vogel, C.S.; Liebman, E.; Curtis, P.S.; Gough, C.M. Coarse woody debris and the carbon balance of a moderately disturbed forest. For. Ecol. Manag. 2016, 361, 38–45. [Google Scholar] [CrossRef] [Green Version]
- Harmon, M.E.; Bond-Lamberty, B.; Tang, J.; Vargas, R. Heterotrophic respiration in disturbed forests: A review with examples from North America. J. Geophys. Res. Biogeosci. 2011, 116, G00K04. [Google Scholar] [CrossRef]
- Nabuurs, G.J.; Lindner, M.; Verkerk, P.J.; Gunia, K.; Deda, P.; Michalak, R.; Grassi, G. First signs of carbon sink saturation in European forest biomass. Nat. Clim. Chang. 2013, 3, 792–796. [Google Scholar] [CrossRef]
- Senf, C.; Seidl, R. Mapping the forest disturbance regimes of Europe. Nat. Sustain. 2021, 4, 63–70. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | N | Overstory | Understory | CWD Volume, m3 ha−1 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Species | A | DBH, cm | Height, m | Growing Stock, m3 ha−1 | Species | Growing Stock, m3 ha−1 | ||||
| Aspen | Other | |||||||||
| 1 | 6 | 10 | – | 109 | 49.0 ± 1.6 | 39.4 ± 0.1 | 687 ± 99 | 5S5L | 118.3 ± 28.6 | 123.5 ± 9.7 |
| 2 | 6 | 9 | 1S | 114 | 57.1 ± 3.2 | 38.2 ± 0.3 | 555 ± 40 | 5S5L | 116.3 ± 20.7 | 142.2 ± 20.5 |
| 3 | 6 | 9 | 1S | 118 | 48.0 ± 1.7 | 38.1 ± 0.2 | 602 ± 42 | 10S | 168.8 ± 14.9 | 28.1 ± 10.1 |
| 4 | 6 | 9 | 1B | 118 | 44.0 ± 2.2 | 38.9 ± 0.5 | 620 ± 83 | 10S | 78.0 ± 14.4 | 111.2 ± 10.5 |
| 5 | 6 | 10 | – | 108 | 46.0 ± 1.9 | 39.5 ± 0.2 | 812 ± 55 | 10S | 111.8 ± 16.6 | 79.6 ± 15.4 |
| 6 | 8 | 7 | 1B1P1S | 109 | 41.0 ± 2.7 | 32.1 ± 0.6 | 623 ± 73 | 9S1Ba | 126.8 ± 12.8 | 75.2 ± 15.2 |
| 7 | 8 | 8 | 2B | 106 | 47.0 ± 2.7 | 32.3 ± 0.5 | 471 ± 31 | 8S1O1B | 81.3 ± 13.2 | 50.3 ± 13.0 |
| 8 | 3 | 9 | 1B | 134 | 53.5 ± 0.8 | 35.2 ± 0.1 | 579 ± 90 | 9S1A | 224.4 ± 21.0 | 61.0 ± 27.8 |
| 9 | 6 | 8 | 1S1B | 114 | 44.9 ± 2.0 | 36.7 ± 0.2 | 628 ± 50 | 10S | 109.4 ± 11.4 | 113.4 ± 10.4 |
| 10 | 6 | 9 | 1S | 104 | 55.6 ± 2.8 | 39.2 ± 0.7 | 783 ± 66 | 10S | 193.7 ± 17.8 | 137.8 ± 29.0 |
| 11 | 6 | 8 | 1B1S | 113 | 56.1 ± 2.9 | 38.7 ± 0.2 | 753 ± 35 | 10S | 138.9 ± 13.9 | 74.9 ± 13.7 |
| 12 | 6 | 9 | 1S | 109 | 47.3 ± 1.7 | 38.4 ± 0.1 | 770 ± 51 | 6S3L1A | 67.8 ± 11.5 | 59.7 ± 11.0 |
| 13 | 6 | 8 | 2S | 104 | 46.3 ± 2.2 | 37.2 ± 0.7 | 601 ± 52 | 7S3L | 105.8 ± 19.0 | 60.2 ± 9.1 |
| 14 | 6 | 9 | 1S | 104 | 49.3 ± 2.0 | 39.3 ± 0.2 | 712 ± 71 | 9S1L | 95.4 ± 12.1 | 174.1 ± 22.9 |
| 15 | 6 | 8 | 2S | 117 | 47.7 ± 2.1 | 33.0 ± 0.5 | 598 ± 52 | 10S | 122.5 ± 20.3 | 104.5 ± 21.8 |
| 16 | 6 | 8 | 1S | 117 | 54.2 ± 1.2 | 36.2 ± 0.1 | 761 ± 69 | 5S4L1B | 46.2 ± 3.1 | 70.0 ± 15.4 |
| 17 | 6 | 9 | 1B | 116 | 52.0 ± 2.4 | 39.1 ± 0.2 | 721 ± 64 | 7S2L1M | 85.4 ± 9.5 | 26.6 ± 10.7 |
| 18 | 6 | 9 | 1S | 111 | 50.1 ± 3.6 | 39.9 ± 0.3 | 950 ± 77 | 10S | 90.3 ± 8.6 | 94.6 ± 14.9 |
| 19 | 6 | 10 | – | 118 | 45.2 ± 1.1 | 37.1 ± 0.2 | 764 ± 48 | 9S1L | 185.3 ± 27.5 | 147.4 ± 27.6 |
| 20 | 6 | 7 | 2S1B | 107 | 52.0 ± 2.9 | 37.4 ± 0.4 | 622 ± 60 | 10S | 99.9 ± 9.6 | 132.5 ± 35.4 |
| 21 | 6 | 7 | 2S1B | 113 | 51.6 ± 2.2 | 36.4 ± 0.3 | 709 ± 71 | 10S | 106.6 ± 14.9 | 112.3 ± 18.7 |
| 22 | 4 | 9 | 1S | 107 | 50.4 ± 2.9 | 33.8 ± 0.2 | 381 ± 37 | 5Ga3As1Bc1S | 14.1 ± 3.3 | 71.0 ± 34.9 |
| 23 | 2 | 5 | 4P1O | 117 | 37.8 ± 10.1 | 27.6 ± 3.2 | 446 ± 31 | 9L1S | 5.3 ± 0.0 | 10.7 ± 3.8 |
| 24 | 3 | 9 | 1B | 104 | 41.4 ± 1.5 | 29.4 ± 0.3 | 617 ± 61 | 9S1B | 77.0 ± 14.0 | 56.8 ± 42.2 |
| 25 | 8 | 8 | 1S1B | 118 | 46.7 ± 2.0 | 37.7 ± 0.2 | 599 ± 47 | 9S1M | 131.9 ± 14.5 | 106.5 ± 24.6 |
| 26 | 6 | 9 | 1S | 108 | 55.6 ± 2.1 | 38.1 ± 0.1 | 689 ± 50 | 8S2M | 102.5 ± 7.2 | 99.3 ± 20.6 |
| Decay Class | Description | Basic Density, kg m3 | Carbon Concentration, % | |
|---|---|---|---|---|
| 1 | Recently dead | Wood hard, knife blade penetrates a few millimeters. Bark attached to the stem. | 391.3 | 47.2 |
| 2 | Weakly decayed | The outer layer of wood starts to soften, knife blade penetrates 1–2 cm. Loose bark, branches present. | 330.6 | 47.4 |
| 3 | Moderately decayed | The wood of outer layers of stem soft, the core still hard, knife blade penetrates <5 cm. Loose, fragmented bark. | 230.6 | 47.4 |
| 4 | Very decayed | Wood soft through the log, knife blade penetrates the wood in its entirety. No branches, most of the surface covered with mosses. | 161.1 | 46.6 |
| 5 | Almost completely decomposed | Lost consistency of wood, breaks up easily. Surface covered with lichens, mosses, and dwarf shrubs. | 60.7 | 46.3 |
| Characteristics | Stand Type and Age | |||||
|---|---|---|---|---|---|---|
| Managed | Unmanaged | |||||
| 41–60 | 61–80 | 41–60 | 61–80 | 81–100 | 101–140 | |
| Stand age, years | 50.1 ± 1.3 | 67.9 ± 1.1 | 53.1 ± 1.0 | 70.1 ± 1.2 | 86.9 ± 1.1 | 111.8 ± 0.5 |
| Site index, m | 28.0 ± 1.0 | 27.2 ± 1.0 | 28.8 ± 0.4 | 27.3 ± 0.7 | 25.7 ± 0.8 | 25.4 ± 0.2 |
| DBH, cm | 36.2 ± 2.2 | 42.8 ± 2.7 | 37.1 ± 1.5 | 39.5 ± 1.3 | 50.4 ± 2.2 | 49.0 ± 0.6 |
| Height, m | 28.0 ± 1.1 | 31.9 ± 1.1 | 29.7 ± 0.4 | 32.4 ± 0.7 | 33.8 ± 0.9 | 36.8 ± 0.2 |
| Total basal area, m2 ha−1 | 29.2 ± 2.5 | 34.2 ± 3.0 | 32.2 ± 1.4 | 39.5 ± 2.3 | 41.8 ± 1.9 | 50.3 ± 0.9 |
| Total growing stock, m3 ha−1 | 330 ± 30 | 447 ± 48 | 377 ± 21 | 526 ± 36 | 577 ± 37 | 774 ± 16 |
| Total number of trees, ha−1 | 1388 ± 245 | 1049 ± 241 | 1421 ± 173 | 1087 ± 143 | 942 ± 108 | 1134 ± 67 |
| Overstory basal area, m2 ha−1 | 21.8 ± 1.7 | 26.1 ± 2.6 | 24.3 ± 1.5 | 29.9 ± 2.3 | 30.4 ± 1.7 | 38.9 ± 0.8 |
| Overstory growing stock, m3 ha−1 | 278 ± 26 | 383 ± 45 | 320 ± 21 | 443 ± 36 | 470 ± 34 | 664 ± 14 |
| Overstory number of trees, ha−1 | 308 ± 34 | 262 ± 29 | 366 ± 34 | 345 ± 33 | 229 ± 25 | 246 ± 7 |
| Aspen basal area, m2 ha−1 | 14.0 ± 2.0 | 17.6 ± 2.4 | 14.6 ± 1.2 | 20.3 ± 2.3 | 21.9 ± 2.1 | 32.1 ± 0.8 |
| Aspen growing stock, m3 ha−1 | 187 ± 25 | 276 ± 42 | 205 ± 18 | 316 ± 38 | 355 ± 39 | 566 ± 15 |
| Aspen number of trees, ha−1 | 363 ± 139 | 162 ± 35 | 296 ± 71 | 273 ± 68 | 126 ± 18 | 202 ± 18 |
| Objects | 19 | 19 | 30 | 26 | 17 | 26 |
| Sample plots | 19 | 19 | 30 | 26 | 17 | 150 |
| CWD Pose | Stand Type | Stand Age, Years | Number of Sample Plots | CWD Volume, m3·ha−1 | CWD Carbon Pool, t·ha−1 | ||
|---|---|---|---|---|---|---|---|
| Marginal Mean | SE | Marginal Mean | SE | ||||
| Standing | Managed | 41–60 | 19 | 7.6 | 6.0 | 1.0 | 0.9 |
| 61–80 | 19 | 5.3 | 6.0 | 0.7 | 0.9 | ||
| Unmanaged | 41–60 | 30 | 16.6 | 4.8 | 2.2 | 0.7 | |
| 61–80 | 26 | 13.8 | 5.2 | 1.9 | 0.7 | ||
| 81–100 | 17 | 26.1 | 6.4 | 3.5 | 0.9 | ||
| 101–140 | 150 | 31.9 | 2.2 | 4.6 | 0.3 | ||
| Lying | Managed | 41–60 | 19 | 12.5 | 12.0 | 1.4 | 1.5 |
| 61–80 | 19 | 31.0 | 12.0 | 4.1 | 1.5 | ||
| Unmanaged | 41–60 | 30 | 53.3 | 9.6 | 6.6 | 1.2 | |
| 61–80 | 26 | 53.5 | 10.3 | 6.4 | 1.3 | ||
| 81–100 | 17 | 47.5 | 12.9 | 5.3 | 1.6 | ||
| 101–140 | 150 | 60.5 | 4.3 | 8.0 | 0.5 | ||
| Total | Managed | 41–60 | 19 | 20.1 | 14.2 | 2.4 | 1.8 |
| 61–80 | 19 | 36.3 | 14.2 | 4.8 | 1.8 | ||
| Unmanaged | 41–60 | 30 | 69.9 | 11.3 | 8.7 | 1.5 | |
| 61–80 | 26 | 67.3 | 12.1 | 8.2 | 1.6 | ||
| 81–100 | 17 | 73.5 | 15.3 | 8.8 | 2.0 | ||
| 101–140 | 150 | 92.4 | 5.1 | 12.5 | 0.7 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šēnhofa, S.; Šņepsts, G.; Bičkovskis, K.; Jaunslaviete, I.; Liepa, L.; Straupe, I.; Jansons, Ā. Availability and Structure of Coarse Woody Debris in Hemiboreal Mature to Old-Growth Aspen Stands and Its Implications for Forest Carbon Pool. Forests 2021, 12, 901. https://doi.org/10.3390/f12070901
Šēnhofa S, Šņepsts G, Bičkovskis K, Jaunslaviete I, Liepa L, Straupe I, Jansons Ā. Availability and Structure of Coarse Woody Debris in Hemiboreal Mature to Old-Growth Aspen Stands and Its Implications for Forest Carbon Pool. Forests. 2021; 12(7):901. https://doi.org/10.3390/f12070901
Chicago/Turabian StyleŠēnhofa, Silva, Guntars Šņepsts, Kārlis Bičkovskis, Ieva Jaunslaviete, Līga Liepa, Inga Straupe, and Āris Jansons. 2021. "Availability and Structure of Coarse Woody Debris in Hemiboreal Mature to Old-Growth Aspen Stands and Its Implications for Forest Carbon Pool" Forests 12, no. 7: 901. https://doi.org/10.3390/f12070901