
Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil
by

Liva Legzdina
1,2,*, 
Voldemars Spungis
2, 
Natalija Burnevica
1,
Talis Gaitnieks
1 and
Audrius Menkis
3 1
Latvian State Forest Research Institute “Silava”, 111 Rigas str., LV-2169 Salaspils, Latvia
2
Faculty of Biology, University of Latvia, 1 Jelgavas str., LV-1004 Riga, Latvia
3
Department of Forest Mycology and Plant Pathology, Swedish University of Agricultural Sciences, P.O. Box 7026, SE-75007 Uppsala, Sweden
*
Author to whom correspondence should be addressed.
Forests 2021, 12(8), 1100; https://doi.org/10.3390/f12081100
Submission received: 14 June 2021
/
Revised: 13 August 2021
/
Accepted: 16 August 2021
/
Published: 17 August 2021
(This article belongs to the Special Issue Forest Pathology and Entomology)
Abstract
:Heterobasidion spp. pathogenic fungi produce conspicuous fruitbodies on infected wood, which may represent a habitat for a range of organisms, including invertebrates. The aim was to: (i) assess and compare invertebrate diversity in Heterobasidion spp. fruitbodies, infected Picea abies wood and adjacent soil; (ii) test methods for collecting invertebrates from Heterobasidion spp. fruitbodies. A total of 69 Heterobasidion fruitbody samples, 46 wood samples and 19 soil samples were collected at two forest sites in Latvia. In total, 7311 invertebrate individuals were collected representing 62 different taxa, among which 55 were in fruitbodies, 47 in wood and 36 in soil. The dominant invertebrates in fruitbodies were Acari (82.4%), Collembola (6.2%), Coleoptera (4.7%); in wood were Acari (78.6%), Collembola (9.6%), Diplopoda (3.1%); in soil were Acari (90.6%), Collembola (3.4%) and Coleoptera (1.4%). Among all taxa, 17.7% were found exclusively in fruitbodies, 3.2% in wood, 4.8% in soil, while 48.4% were shared among different substrates. Although many invertebrate taxa were shared, their relative abundance differed substantially among different substrates. The sampling methods used provided valuable information on invertebrate communities. Further development of these methods may be needed to obtain more detailed and specific information about invertebrates inhabiting different habitats.
1. Introduction
Heterobasidion spp. pathogenic fungi cause considerable economic losses to forestry due to the infection of living trees and decay of wood. The impact of the pathogen is more severe in Norway spruce (Picea abies) stands as the fungal mycelia from the root system can expand several meters up in the stem, thereby causing rot in the most valuable part of the stem [1]. In Latvia, ca. 22% of P. abies stands are infected by Heterobasidion spp. and, on average, the economic loss at a final cutting exceeds EUR 4000/ha [2]. Heterobasidion spreads by basidiospores (primary infections) and by vegetative mycelia through root contacts (secondary infections) [3]. The abundance of fruitbodies of the pathogen in a particular stand has an impact on the primary infection by basidiospores. It is known that these fruitbodies mainly affect the rate of infection at the stand level but not in adjacent stands [4]. Both Heterobasidion parviporum Niemelä & Korhonen and Heterobasidion annosum (Fr.) Bref. are found in Latvia, and both of these species form fruitbodies on infected P. abies wood at a similar rate [5]. Studies conducted in Germany and Finland [6,7] have indicated that a large number of fruitbodies are formed on 3- to 4-year-old large dimension P. abies logs that are left in stands after forest operations. In Latvia, the area covered by fruitbodies on 3- to 4-year-old P. abies logs is on average 3759 cm2/m3, and can exceed 5000 cm2/m3 on logs in stands growing on drained peat soils [5].
Heterobasidion fruitbodies can produce between 35,000 and 151,000 basidiospores per cm2 of the fruitbody in one hour [8], and, in P. abies stands, the number of spores can exceed 1000 per m2 per hour [9]. This spore density is a high risk to freshly cut P. abies stumps during tree felling as fresh stumps are the primary substrate for colonisation by Heterobasidion spores. Therefore, to reduce the risk of infection, several studies have recommended the removal of large dimension P. abies logs that are infected by Heterobasidion spp. from forest stands [5,6,7]. However, other studies have indicated that large dimension P. abies logs are important as a habitat to many fungal and invertebrate species [10,11]. Fungal fruitbodies are known to be inhabited by a number of invertebrate species [12,13]. Fruitbodies provide not only a food source for invertebrates but also shelter and a place for reproduction, thus influencing the density of invertebrates in a given substrate [11]. Invertebrate fauna have been investigated in the fruitbodies of Fomitopsis pinicola (Fr.) Karst, a fungal species commonly found in conifer forests [14,15,16,17]. However, in most studies on invertebrate fauna inhabiting fungal fruitbodies, multiple fungal species were analysed, without differentiation. The focus has often been on macro arthropods that feed on fungi, typically Coleoptera and Diptera [17,18,19]. However, fungal fruitbodies are also known to be commonly inhabited by Acari and Collembola [20,21]. Some studies have also investigated Heterobasidion spp. fruitbodies, however, the majority of these have examined a small number of fruitbodies, from which only adult Coleoptera were collected as these are relatively easy to identify. In a study that included several fungal species, adult Coleoptera were found in only two H. parviporum fruitbodies from a total of 44 collected [22], while in another similar study, only half of examined H. parviporum fruitbodies were inhabited by Coleoptera [23]. Heterobasidion annosum was shown to be associated with several Coleoptera species from the family Ciidae [24], and reproduction in fruitbodies was confirmed for Cis lineatocribatus, with both adults and larvae identified [18]. The possible role of insect vectors for dissemination of Heterobasidion spp. has previously been highlighted by Stenlid [25] and Piri et al. [26]. Indeed, spores of Heterobasidion spp. are known to be disseminated by the large pine weevil (Hylobius abietis) [27,28], but the role of other insects is largely unknown. To determine the effect of the removal of infected P. abies wood from forest stands on invertebrate diversity, quantitative and qualitative assessment of invertebrate fauna in Heterobasidion spp. fruitbodies is required, particularly focussing on rare and endangered species. The aim of this study was to: (i) assess and compare invertebrate diversity in Heterobasidion spp. fruitbodies, infected P. abies wood and adjacent soil; (ii) develop and test methods for collecting invertebrate samples from Heterobasidion spp. fruitbodies.
2. Materials and Methods
2.1. Sample Plots
Samples were collected from two sample plots in the autumn of 2016: (i) a 70-year-old forest stand consisting of 80% P. abies with an admixture of Betula pendula and Pinus sylvestris on peatland (forest type Oxalidosa turf.mel. [29]) near the Forest Research Station in the Kalsnava forest region (56°41′40.4″ N, 25°54′21.8″ E), further referred to as Kalsnava; (ii) a 50-year-old forest stand consisting of 90% P. abies with an admixture of B. pendula on mineral soil (forest type Oxalidosa [29]) in JSC Latvia’s State ForestsCentral Daugava region, Ogre division (56°50′50.8″ N, 24°48′17.8″ E), further referred to as Ogre. Sample plots contained infected P. abies logs of different sizes and decomposition stages, where Heterobasidion spp. fruitbodies of varying sizes and ages were previously identified.
2.2. Field Methods
To sample fruitbodies, 46 infected P. abies logs with large Heterobasidion spp. fruitbodies (Figure 1) were randomly selected in each plot. For each log, the root end diameter and length were measured. The length of logs ranged between 107 and 487 cm (average 274 cm); diameters ranged between 15 and 42 cm (average 25 cm). Fruitbody samples (100 cm2) were collected by cutting a 10 × 10 cm piece from the fruitbody closest to the root end of the log. During the collection of fruitbody samples, it was noted if the fruitbody was in contact with the soil and if any damage was visible. A similarly sized (100 cm2) wood sample was collected 20 cm from the fruitbody in the direction away from the root end of the log. One wood sample and at least one fruitbody sample was collected from each log. If multiple and well-separated fruitbodies were found on one log, two to three fruitbody samples were collected. Where the log was in contact with the soil, a 5 cm deep, 100 cm2 soil sample was collected. If multiple logs were found less than 10 m from each other (in log piles), only one soil sample was collected. After collection, all samples were placed into plastic bags, and kept at +4 °C until processing. A total of 69 Heterobasidion spp. fruitbodies, 46 P. abies wood samples and 19 soil samples were collected. In Kalsnava, 46 fruitbodies, 30 P. abies wood samples and 15 soil samples were collected, while in Ogre, 23 fruitbodies, 16 P. abies wood samples and 4 soil samples were collected.
2.3. Laboratory Methods
In the laboratory, each sample was weighed using a Kern EMB 200-2 balance (Balingen, Germany) before and after placing samples in Tullgren funnels (Figure 2), and macrofauna were collected (if present) using forceps. By weighing the samples before and after placing them in Tullgren funnels, the relative humidity of samples was calculated. The relative humidity of fruitbodies ranged from 40.6% to 85.6% with an average of 70.1%. In Ogre, the relative humidity of fruitbodies was on average 69.4%, but in Kalsnava it was on average 71.6%. The average weight of a fruitbody sample before drying was 68.7 g, maximal and minimal values being 151.0 g and 14.2 g, respectively. After drying, the weight varied between 5.1 g and 42.3 g with an average of 19.9 g.
The thickness of fruitbodies was measured with a ruler. Most of the collected fruitbodies were between 0.4 and 2.5 cm thick, but some fruitbodies that were collected close to the root collar were up to 5.0 cm thick. The majority of fruitbodies collected were at least 3 years old with some being between 6 and 7 years old.
After measurements, samples were placed in Tullgren funnels for at least 72 h. Tullgren funnels (Figure 2) are commonly utilized to obtain quantitative data about arthropods in soil [30], but this method is also effective to obtain data on invertebrates in fruitbodies and wood [14,31,32]. A 1:3 mixture of glycerine and 70% ethanol was placed in the collection vials to preserve the collected invertebrate individuals.
The collected invertebrates were examined using Leica NBO MSV266 (Wetzlar, Germany) and Optika B-383PH (Ponteranica, Italy) microscopes. Invertebrates were classified to the lowest possible taxonomic level, most to family level. Springtails (Collembola) were classified according to W. Dunger [33], mites (Acari) according to unpublished material by Dr U. Kagainis on the classification of oribatid mites (Oribatida) morphogroups in Latvia. Beetles (Coleoptera) were classified to the family level using the key by D. M. Unwin [34]. Taking into account the available information on Coleoptera of the Ciidae family inhabiting Heterobasidion spp. fruitbodies, these were classified to species level using the publicly available key by Lompes [35], which was developed from the results of studies by Reibnitz [24].
2.4. Statistical Analyses
To analyse invertebrate diversity in each substrate, the invertebrate data (actual observations) were used to construct accumulation curves [36], which was conducted in R [37] using the vegan package and specaccum function [38]. Differences in invertebrate taxa richness in different substrates (study sites as well as adults and larvae combined) were compared by nonparametric chi-square tests, taking into account the Bonferroni correction [39]. Shannon index values [40] were calculated for each substrate to determine the diversity in invertebrate communities. Values were calculated using invertebrate frequencies by taxa, i.e., the subclass or family taxonomic level. The invertebrate community structure in different substrates (fruitbodies, wood and soil) was analysed using principal component analysis in Canoco v.5.02 (Microcomputer Power, Ithaca, NY, USA).
3. Results
A total of 7311 invertebrate individuals were collected from all substrates and from both sample plots, including 7151 adults and 160 larvae. The number of invertebrates collected per individual fruitbody was between 1 and 396 (average 70). In wood and soil samples, invertebrate numbers ranged from 0 to 316 (average 35) and 0 to 192 (average 41), respectively. There were 55 different invertebrate taxa in 69 fruitbody samples, 47 taxa in 46 wood samples and 36 taxa in 19 soil samples. Consequently, the chi-square test showed that the absolute richness of invertebrate taxa differed significantly among all substrates, being highest in the soil, then in the wood and lowest in fungal fruitbodies (p < 0.004). A possible reason for the observed differences may be an uneven number of samples collected of different substrates. Indeed, when the same number of samples was taken from each substrate following the rarefaction analysis, the richness of invertebrate taxa did not differ significantly among different substrates (p > 0.05).
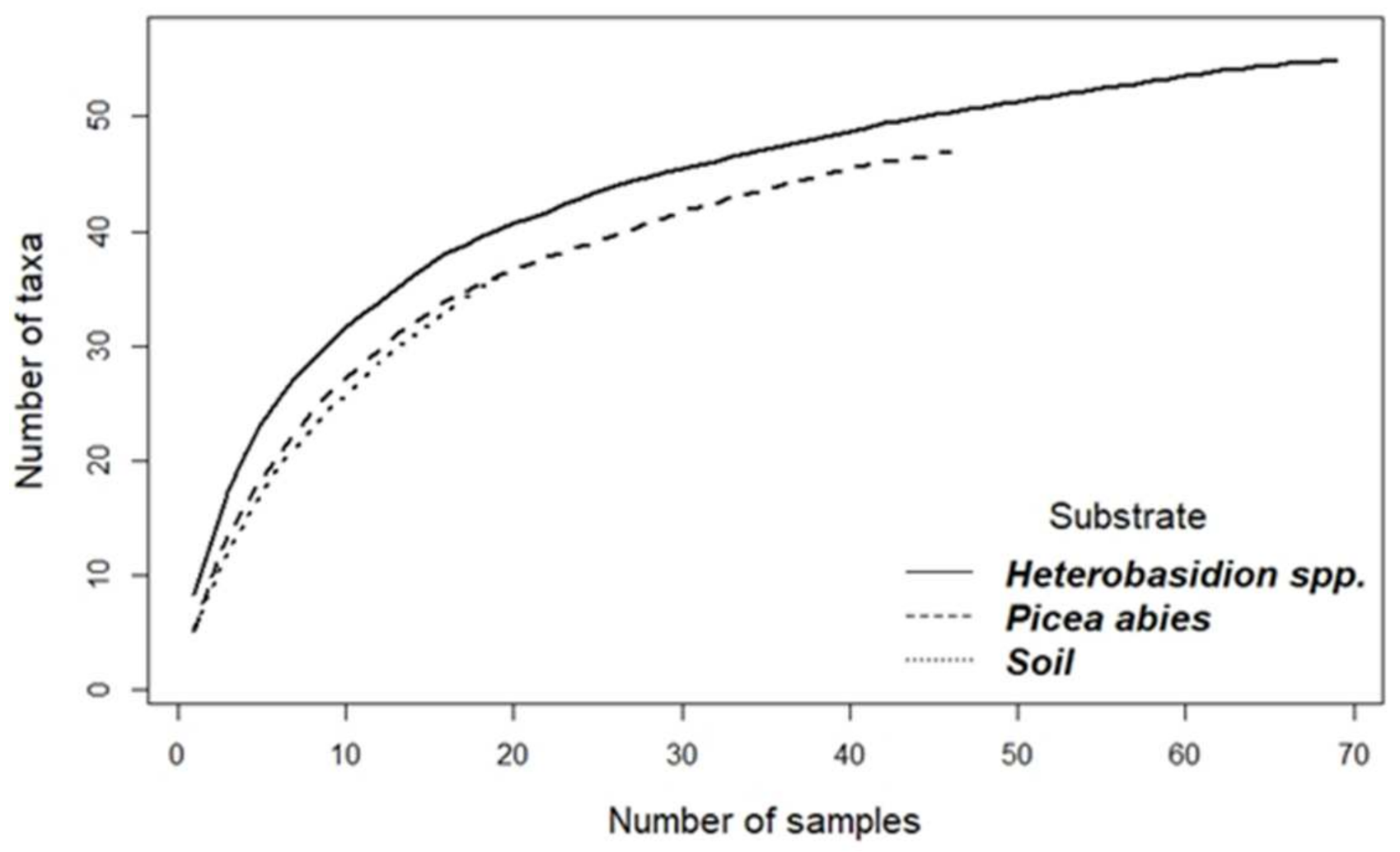
The accumulation curves for different substrates did not reach the asymptote (Figure 3).
The largest number of invertebrates was collected from Heterobasidion spp. fruitbodies—4886 adults and 101 larvae (Table 1). The dominant invertebrate taxa were Acari (82.4%), with the other invertebrates being Collembola (6.2%), Coleoptera (4.7%), Diplopoda (2.5%) and Diptera (1.8%). In addition, Haplotaxida (0.8%), Thysanoptera (0.7%), Hymenoptera (0.3%), Aranea (0.2%), Gastropoda (0.1%) and Chilopoda (0.1%) were found in fruitbodies. Several invertebrate groups were represented by very few individuals in fruitbodies—Lepidoptera, Isopoda and Hemiptera (0.04% each) and Protura and Opiliones (0.02% each).
Coleoptera of the Ciidae family were most prevalent in fruitbodies (86.8% of all beetles collected): Cis nitidus (27.6% of all beetles collected), C. castaneus (19.4%), C. lineatocribatus (18.8%), C. jacquemartii (14.7%) and C. glabratus (5.8%). Adult Phosphuga atrata, Daliopius marginatus and Oxypselaphus obscurus were observed, as well as larvae of several beetle families, i.e., Elateridae, Cantharidae, Ciidae and Staphylinidae. Larvae of these taxa were also found in wood samples; however, they were in a smaller number.
Three Diplopoda species were found in fruitbodies. The most prevalent species was Proteroiulus fuscus, which was twice as prevalent than in wood samples. One Cylindroiulus caeruleocinctus individual was found only in fruitbodies, as well as thirteen Polydesmus denticalatus individuals. Three earthworm species Lumbricus castaneus, L. rubellus and L. terrestris were found in fruitbodies. These species were also found in wood samples; however, they were in a smaller number.
The second largest number of invertebrates was collected from wood samples, i.e., 1570 adults and 54 larvae (Table 1). The dominant invertebrate taxa were Acari (78.6%), with the other invertebrates being Collembola (9.6%), Diplopoda (3.1%), Coleoptera (2.5%), Diptera (1.9%) and Haplotaxida (1.1%). Several invertebrate groups were represented by a few individuals in wood samples, which were Pseudoscorpionida and Thysanoptera (0.6% each), Aranea (0.5%), Hymenoptera (0.4%), Gastropoda (0.4%) Lepidoptera larvae (0.2%), Chilopoda (0.2%), Isopoda (0.1%) and Hemiptera (0.1%).
The smallest number of samples were collected from soil samples, i.e., 695 adults and five larvae (Table 1). As in the fruitbody and wood samples, Acari were the dominant invertebrate taxon (90.6%). Other invertebrates were present in small numbers: Collembola (3.4%), Coleoptera (1.4%), Haplotaxida (1.1%) and Hymenoptera (1.1%). Several invertebrate groups were represented by very few individuals in soil samples: Diptera (0.6%), Aranea (0.4%), Pseudoscorpionida (0.3%) and Hemiptera (0.3%), and Chilopoda, Diplopoda, Thysanoptera, Lepidoptera larvae and Gastropoda (0.14% each).
Shannon index values were 2.37 in wood samples, 1.63 in fruitbodies and 1.95 in soil samples.
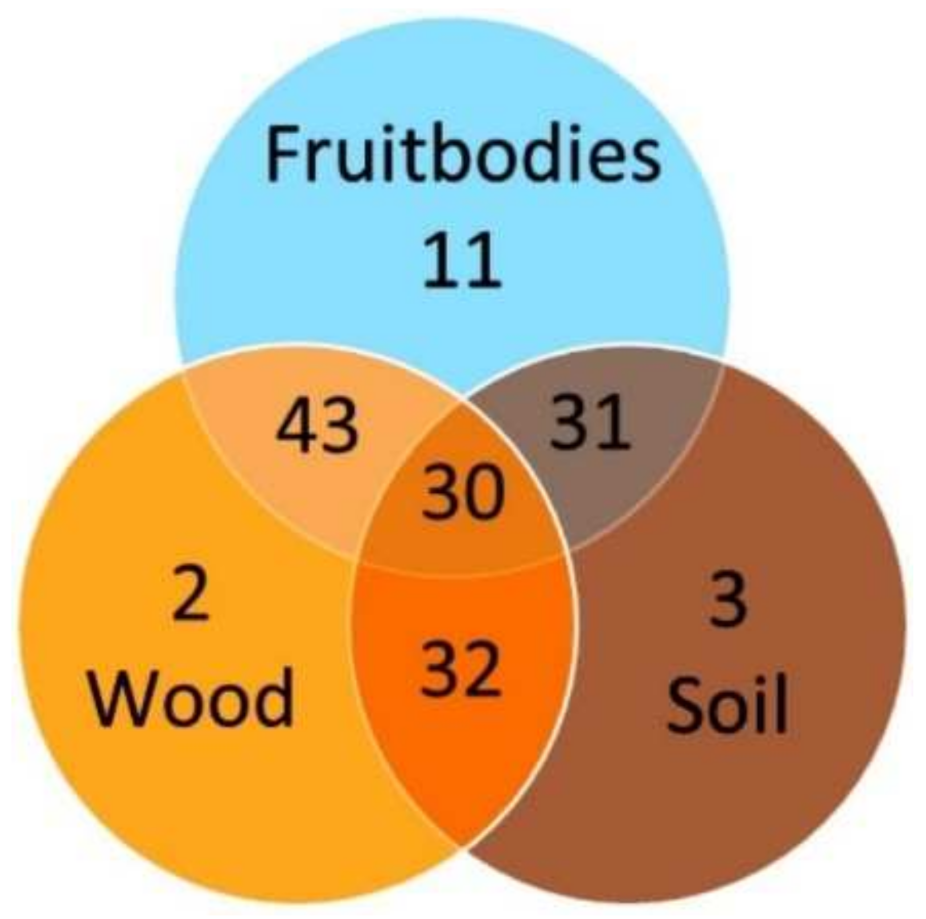
Dominance of one invertebrate taxonomic group was pronounced with Acari dominating in all substrates, and Collembola was the second most common group (Table 1). The third most prevalent taxonomic group varied between substrates, i.e., in Heterobasidion spp. samples it was Coleoptera, in wood it was Diplopoda and in soil it was Coleoptera and Haplotaxida. Among the 62 invertebrate taxa found in the present study, eleven (17.7%) were exclusively found in fruitbodies, two (3.2%) in the wood and three (4.8%) in the soil, while thirty (48.4%) were common to all substrates (Figure 4). There were 43 shared invertebrate taxa between fruitbodies and wood, 31 between fruitbodies and soil and 32 between wood and soil (Figure 4).
In principal component analysis (PCA) of invertebrate communities, axis 1 explained 81.7% of variation and axis 2 explained 18.3%. PCA showed that invertebrate communities from different substrates were separated from each other on axis 1, i.e., on the axis, which explained most of the variation (Figure 5).
4. Discussion
The taxonomic composition of invertebrates found in Heterobasidion spp. samples reflects the biology of fruitbodies. Fruitbodies develop on the part of the infected substrate that are in contact with the soil [7]. If the wood substrate is embedded in the soil, the fruitbody develops in a narrow line at the point of contact between the soil and the wood [5]. Therefore, the dominant invertebrate taxa in fruitbodies were also present in wood and soil samples (Table 1, Figure 4). Although the majority of invertebrate taxa were shared among different substrates, the PCA showed that invertebrate communities in different substrates were separated from each other, which was likely due to differences in the abundance data of different taxa (Table 1), thereby demonstrating the specificity and substrate preferences of different invertebrates. Our data show that Acari were the predominant invertebrate taxa found in Heterobasidion spp. fruitbodies (Table 1). This is consistent with other reports about fauna of invertebrates found in Heterobasidion spp. [32] and F. pinicola [14] fruitbodies. Some of the Acari that were not identified to family level belong to the order Mesostigmata, which was previously identified in fruitbodies from other species, such as F. pinicola [41]. Acari of the genus Carabodes were the most prevalent in fruitbodies. The presence of this genus in fruitbodies has been studied extensively in coniferous forests in Norway [14,42], and many taxa inhabiting soil also occupy fruitbodies as an additional food resource. This was also observed for Acari collected from Heterobasidion spp. [32].
Several studies have noted that Heterobasidion spp. fruitbody development is promoted by shade and moisture [5,43,44]. In this study, invertebrate groups that prefer moist environments, such as Haplotaxida, Diplopoda and Chilopoda, were also frequently detected in fruitbodies. However, these invertebrates are probably incidental and temporary inhabitants of fruitbodies even though their numbers in the analysed samples was relatively high (Table 1). Diplopoda and Chilopoda are prone to dehydration, and therefore seek moist places to forage [45]. The relative moisture content of the fruitbodies analysed in this study was 70%, and water drained from them when compressed. A portion of the Haplotaxida was collected from the oldest peripheries of the fruitbodies, where other invertebrates had already formed tunnels.
Heterobasidion spp. form perennial fruitbodies with a layered, cork-like structure [3]. Several Diptera adults and larvae were identified in fruitbodies, however the number of individuals collected was relatively small in comparison with the total number of invertebrates collected, which could be due the structure of fruitbodies. Diptera most often inhabit softer, fruitbodies of annual fungi that decay faster [46]. However, despite the structural differences of various fungal fruitbodies, our study is in agreement with the results from other studies [47,48], i.e., showing the low level of specialization between Diptera. Therefore, fungal fruitbodies can be colonized by various groups of Diptera. The number of amassed Diptera could be affected by the time of sampling as these insects are mostly inactive during the dormancy period.
One of the aims of this study was to compare invertebrate fauna inhabiting fruitbodies, rotting wood and soil. While these substrates are in close contact with each other in nature, differences in the invertebrate fauna inhabiting them were observed (Table 1, Figure 4 and Figure 5). These differences may be due to the season when samples were collected (October). Assessment of invertebrate fauna during the vegetation period may be more representative. Each substrate had a characteristic invertebrate fauna profile (Figure 5), and the number of individuals and taxa collected varied between different substrates. For example, Acari of the genus Carabodes were most prevalent in fruitbodies but were less common in wood and soil samples (Table 1). By contrast, some other Acari were most often found in wood and soil samples (Oppiidae and Ceratozetidae, respectively) but were less commonly found in fruitbodies. In total, 90% of invertebrate individuals collected from soil samples were Acari, while in fruitbody and wood samples the proportion of Acari was lower (82% and 78%, respectively). Although the majority of invertebrate taxa were found in all substrates, some characteristic taxa were only found in fruitbodies (e.g., Ciidae), but not in wood or soil samples (Figure 4).
In studies of polypore inhabitants, usually the entire fruitbody is collected [17,32]. Studies in Latvia on the development of Heterobasidion spp. fruitbodies on decaying P. abies wood indicate that the size of fruitbodies can vary widely, and older fruitbodies coalesce with each other [5]. Therefore, in this study, only partial fragments of fruitbodies were collected. Although Heterobasidion spp. form large fruitbodies on windthrown trees and large dimension logging debris in favourable conditions [3,5], collection of a large number of 10 cm × 10 cm fragments of Heterobasidion spp. fruitbodies is difficult. Although the area of collected fruitbodies was uniform (100 cm2), the volume of fruitbodies varied. This was due to differing thicknesses of the collected fruitbodies, which is influenced by the age of fruitbodies. In addition, the dry weight of collected fruitbodies varied between 5.1 g and 47.3 g, which has also been reported to influence invertebrate numbers and diversity in H. annosum fruitbodies [32]. The obtained data indicate that in further research, a larger number of samples of smaller size could be collected.
The Shannon diversity index values indicated that invertebrate diversity in fruitbodies was moderate. As mentioned before, the results of the study were influenced by the season when samples were collected. Heterobasidion spp. sporulate throughout the year, however, the maximum rate is between June and August [49,50]. Collection of fruitbodies at this time may identify invertebrate taxa that feed on spores at some developmental stage or taxa that are inactive during colder periods of the year. Analysis of invertebrates present in fruitbodies during peak sporulation times may identify potential vectors that can influence the primary infection pathways of this pathogen [51]. Previous studies on the role of different invertebrates as vectors of Heterobasidion have mainly focused on fungal conidia [3,52], but further studies are required to better understand the nature of such associations.
In this study, samples were placed in Tullgren funnels to collect invertebrates for three days. In total, 95% of the individuals collected were collected within the first 24 h. Some reports [45,53] indicate that in order to identify certain invertebrates, samples need to be held in Tullgren funnels for a longer period, for example, to allow development of larvae. Some Diptera and Coleoptera larvae were collected in this study, however, the proportion of larvae of the total number of individuals collected was low (2% in fruitbodies). To obtain a more comprehensive assessment of invertebrate fauna inhabiting Heterobasidion spp., methods that allow the development of larvae should be considered, by not preserving collected individuals in glycerine and placing collected larvae in rearing chambers, or by placing fruitbodies in vials and collecting emerged insects. In addition, Tullgren funnels are only partially effective for the collection of mobile invertebrates such as Collembola and should be used in conjunction with other collection methods [54], for example, by covering funnels. In this study, Collembola were observed on the surface of the samples during both sample sorting and collection in Tullgren funnels.
The stage of wood decay also influences invertebrate numbers and diversity, similarly to the decay stage of fruitbodies. The majority of organisms associated with polypores utilise fruitbodies in the later succession stages [11]. During the sterile phase, when the fungal hymenium layer is developing, perennial polypore species produce chemicals that protect them against invertebrate damage, which ceases when the fruitbody starts to decay [14]. Invertebrates may only start to inhabit fruitbodies after several years. In addition, fruitbodies on a single log do not start to decay simultaneously, ensuring suitable conditions for invertebrates that colonise decaying fruitbodies, even over several decades. Studies in Latvia have identified Heterobasidion spp. fruitbodies in various stages of decay on more than ten-year-old rotting spruce wood [5].
Ciidae was the most prevalent Coleoptera family collected from fruitbodies. This is a functionally important group that inhabits fruitbodies during both larval and adult stages, and several successive generations are often found in a single fruitbody. The diversity of Ciidae species is significantly influenced by host organism incidence within a territory, as well as fruitbody developmental stages [17,55]. Cis bidentatus was the only Coleoptera species collected from H. parviporum fruitbodies in Finland, and that colonised decaying fruitbodies or decaying parts of still living fruitbodies [23]. However, this species was not identified in this study.
While a comparatively small number of Cis glabratus individuals were identified in this study, this species has been reported to be prevalent in H. annosum [24] and F. pinicola fruitbodies [15,17]. In addition to species collected in this study (C. lineatocribatus, C. nitidus, C. glabratus and C. castaneus), another study also reported the presence of C. bidentatus and Ennearthron cornutum on Heterobasidion spp. [24].
According to the European Union Council directive 92/43/EEK (Protecting Europe’s biodiversity (Natura 2000)), no rare or protected species of invertebrates were identified in this study.
Further research is needed to reveal the effect of the decay stage of wood and fruitbodies, as well as the effect of wood dimensions on the invertebrate fauna found in Heterobasidion spp. fruitbodies. Differences in the invertebrate fauna inhabiting decaying wood in areas of mineral and peat soils could be assessed by collecting samples from forest stands of different forest types.
5. Conclusions
The results demonstrated the presence of a taxa-rich community of invertebrates inhabiting Heterobasidion fruitbodies, decayed P. abies logs and adjacent soil. Although a large number of invertebrates were shared among different substrates, their relative abundance varied substantially showing certain specificity and substrate preferences of particular taxa.
Author Contributions
Conceptualization, L.L., V.S. and T.G.; methodology, V.S. and L.L.; software, L.L. and A.M.; validation, T.G.; formal analysis, L.L. and A.M.; investigation, L.L., V.S., N.B. and T.G.; resources, T.G.; data curation, L.L. and T.G.; writing—original draft preparation, L.L. and T.G.; writing—review and editing, A.M., V.S. and L.L.; visualization, L.L., A.M. and N.B.; supervision, T.G.; project administration, T.G.; funding acquisition, T.G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was carried out as a part of National forest monitoring (Monitoring of damages by diseases and insects) according with The Regulation of the Cabinet of Ministers No. 238, and in accordance with the contract No. 1.2.1.1/18/A/004 between ‘Forest Sector Competence Centre of Latvia’ Ltd. and the Central Finance and Contracting Agency. The study ‘Development of chemical preparation for reducing root rot caused losses in Norway spruce stands on peat soils’ is conducted by LSFRI Silava with support from the European Regional Development Fund (ERDF) within the framework of the project ‘Forest Sector Competence Centre of Latvia’. AM was supported by the Swedish Research Council Formas (project no. 2019-00597).
Data Availability Statement
The data is available upon request from the corresponding author.
Acknowledgments
We are grateful to Lauma Bruna and Diana Meiere for help with the field work and collecting the material, Dagmara Cakstina for help with the laboratory work and Dainis Edgars Rungis for language revision.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Garbelotto, M.; Gonthier, P. Biology, epidemiology, and control of Heterobasidion species worldwide. Annu. Rev. Phytopathol. 2013, 51, 39–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaitnieks, T.; Arhipova, N.; Donis, J.; Stendlid, J.; Vasaitis, R. Butt rot incidence and related losses in Latvian Picea abies (L.) Kasrt. stands. In Proceedings of the 12th International Conference on Root and Butt Rots, 12–19 August 2007; The University of California: Berkley, CA, USA, 2008; pp. 177–179. [Google Scholar]
- Korhonen, K.; Stenlid, J. Biology of Heterobasidion annosum. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 43–70. [Google Scholar]
- Stenlid, J. Regional differentiation in Heterobasidion annosum. In Proceedings of the Root and Butt Rots: Proceedings of the International Conference on Root and Butt Rots (IUFRO) Working Party S2.06.01, Wik, Sweden and Haikko, Finland, 9–16 August 1993; Johansson, M., Stenlid, J., Eds.; Swedish University of Agricultural Sciences: Uppsala, Sweden, 1994; pp. 243–248. [Google Scholar]
- Gaitnieks, T.; Bruna, L.; Zaluma, A.; Burnevica, N.; Klavina, D.; Legzdina, L.; Jansons, J.; Piri, T. Development of Heterobasidion spp. fruit bodies on decayed Piecea abies. For. Ecol. Manag. 2021, 482, 118835. [Google Scholar] [CrossRef]
- Schütt, P.; Schuck, H.J.; Lautenschlager, K.; Prestle, W.; Stimm, B. Fomes annosus sporocarps—Their abundance on decayed logs left in the forest. Eur. J. For. Pathol. 1979, 9, 57–61. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.M.; Heinonen, J.; Korhonen, K. Occurrence of Heterobasidion basidiocarps on cull pieces of Norway spruce left on cutting areas and in mature spruce stands. For. Pathol. 2007, 37, 374–386. [Google Scholar] [CrossRef]
- Möykkynen, T.; Von Weissenberg, K.; Pappinen, A. Estimation of dispersal gradients of S- and P-type basidiospores of Heterobasidion annosum. Eur. J. For. Pathol. 1997, 27, 291–300. [Google Scholar] [CrossRef]
- Gonthier, P.; Garbelotto, M.M.; Nicolotti, G. Seasonal patterns of spore deposition of Heterobasidion species in four forests of the western Alps. Phytopathology 2005, 95, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Penttilä, R.; Siitonen, J.; Kuusinen, M. Polypore diversity in managed and old-growth boreal Picea abies forests in southern Finland. Biol. Conserv. 2004, 117, 271–283. [Google Scholar] [CrossRef]
- Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Hanski, I. Fungivory: Fungi, Insects and Ecology. In Insect Fungus Interactions, Proceedings of the 14th Symposium of the Royal Entomological Society of London in Collabaration with the British Mycological Society, London, UK, 16–17 September 1989; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 25–61. [Google Scholar]
- Hammond, P.; Lawrence, J.F. Appendix. Mycophagy in insects: A summary. In Insect Fungus Interactions, Proceedings of the 14th Symposium of the Royal Entomological Society of London in Collabaration with the British Mycological Society, London, UK, 16–17 September 1989; Wilding, N., Collins, N.M., Hammond, P.M., Webber, J.F., Eds.; Academic Press: London, UK, 1989; pp. 275–324. [Google Scholar]
- Hågvar, S.; Steen, R. Succession of beetles (genus Cis) and oribatid mites (genus Carabodes) in dead sporocarps of the red-banded polypore fungus Fomitopsis pinicola. Scand. J. For. Res. 2013, 28, 436–444. [Google Scholar] [CrossRef]
- Jonsell, M.; Nordlander, G. Insects in polypore fungi as indicator species: A comparison between forest sites differing in amounts and continuity of dead wood. For. Ecol. Manag. 2002, 157, 101–118. [Google Scholar] [CrossRef]
- Komonen, A.; Jonsell, M.; Okland, B.; Sverdrup-Thygeson, A.; Thunes, K. Insect assemblage associated with the polypore Fomitopsis pinicola: A comparison across Fennoscandia. Entomol. Fenn. 2004, 15, 102–112. [Google Scholar] [CrossRef]
- Thunes, K.H.; Midtgaard, F.; Gjerde, I. Diversity of coleoptera of bracket fungus Fomitopsis pinicola in a Norweigian spruce forest. Biodivers. Conserv. 2000, 9, 833–852. [Google Scholar] [CrossRef]
- Benick, L. Pilzkäfer und Käferpilze. Ökologische und statistische Untersuchungen. Acta Zool. Fenn. 1952, 70, 1–250. [Google Scholar]
- Okland, B. Insect fauna compared between six polypore species in a southern Norwegian spruce forest. Fauna Nor. Ser. B 1995, 42, 21–26. [Google Scholar]
- Graves, R.C.; Graves, A.C.F. The Insects and Other Inhabitants of Shelf Fungi in the Southern Blue Ridge Region of Western North Carolina. I. Introduction, Description of the Habitat, and the Host Fungi. Ann. Entomol. Soc. Am. 1966, 59, 381–390. [Google Scholar] [CrossRef]
- Takahashi, K.H.; Tuno, N.; Kagaya, T. Abundance of mycophagous arthropods present on different species of fungi in relation to resource abundance at different spatial scales. Eur. J. Entomol. 2005, 102, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Schigel, D.S.; Niemelä, T.; Kinnunen, J. Polypores of western Finnish Lapland and seasonal dynamics of polypore beetles. Karstenia 2006, 46, 37–64. [Google Scholar] [CrossRef] [Green Version]
- Schigel, D.S. Polypore–beetle associations in Finland. Fenn. Ann. Zool. Fenn. 2011, 48, 319–348. [Google Scholar] [CrossRef]
- Reibnitz, J. Verbreitung und Lebensräume der Baumschwammfresser Südwestdeutschlands (Coleoptera: Cisidae). Mitt. Entomol. Ver. Stuttg. 1999, 34, 1–76. [Google Scholar]
- Stenlid, J. Population structure of Heterobasidion annosum as determined by somatic incompatibility, sexual incompatibility, and isoenzyme patterns. Can. J. Bot. 1985, 63, 2268–2273. [Google Scholar] [CrossRef]
- Piri, T.; Korhonen, K.; Sairanen, A. Occurrence of Heterobasidion annosum in Pure and Mixed Spruce Stands in Southern Finland. Scand. J. For. Res. 1990, 5, 113–125. [Google Scholar] [CrossRef]
- Kadlec, Z.; Stary, P.; Zumr, V. Field evidence for the large pine weevil, Hylobius abietis as a vector of Heterobasidion annosum. Eur. J. For. Pathol. 1992, 22, 316–318. [Google Scholar] [CrossRef]
- Drenkhan, T.; Sibul, I.; Kasanen, R.; Vainio, E.J. Viruses of Heterobasidion parviporum persist within their fungal host during passage through the alimentary tract of Hylobius abietis. For. Pathol. 2013, 43, 317–323. [Google Scholar] [CrossRef]
- Bušs, K. Forest ecosystem classification in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 1997, 51, 204–218. [Google Scholar]
- Crossley, D.A.; Blair, J.M. A high-efficiency, “low-technology” Tullgren-type extractor for soil microarthropods. Agric. Ecosyst. Environ. 1991, 34, 187–192. [Google Scholar] [CrossRef]
- Lindquist, E.E. Remarkable convergence between two taxa of ascid mites (Acari: Mesostigmata) adapted to living in pore tubes of bracket fungi in North America, with description of Mycolaelaps new genus. Can. J. Zool. 1995, 73, 104–128. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, T.; Bolger, T. Fungal fruiting bodies and the structure of fungus-micro-arthropod assemblages. Biol. Environ. 1997, 97, 249–262. [Google Scholar]
- Dunger, W.; Fiedler, H.J. Methoden der Bodenbiologie; Gustav Fischer Verlag: Stuttgart, Germany, 1997. [Google Scholar]
- Unwin, D.M. A key to the families of British Beetles. Field Stud. Counc. 1988, 166, 149–197. [Google Scholar]
- Lompe, A. Käfer Europas. Familir Ciidae. Available online: http://www.coleonet.de/coleo/texte/ciidae.htm (accessed on 13 May 2017).
- Colwell, R.K.; Chang, X.M.; Jing, C. Interpolating, extrapolating and comparing incidence-based species accumulation curves. Ecology 2004, 85, 2717–2727. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2015. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. “Package ‘Vegan’ ” Community Ecology Package, Version 2.5–7; 2020. Available online: http://CRAN.R-project.org/package=vegan (accessed on 25 January 2021).
- Magurran, A.E. Ecological Diversity and Its Measurement; Princeton University Press: Princeton, NJ, USA, 1988. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Makarova, O.L. Gamasid mites (Parasitiformes, Mesostigmata) dwelling in bracket fungi at the pechoro-ilychskii reserve (Republic of Komi). Zool. Zhurnal 2004, 83, 1335–1340. [Google Scholar]
- Hågvar, S.; Amundsen, T.; Økland, B. Mites of the genus Carabodes (Acari, Oribatida) in Norwegian coniferous forests: Occurrence in different soils, vegetation types and polypore hosts. Scand. J. For. Res. 2014, 29, 629–638. [Google Scholar] [CrossRef]
- Negruckii, S.F. Kornevaya Gubka, 2nd ed.; Agropromizdat: Moscow, Russian, 1986. [Google Scholar]
- Vasiliauskas, A.P. Root Fungus and Resistance of Coniferous Forests Ecosystems; Mokslas Publishers: Vilnius, Lithuania, 1989. [Google Scholar]
- Graves, R.C. Ecological observation on the insects and other inhabinants of woody shelf fungi (Basidiomycetes: Polyporaceae) in the Chichago area. Ann. Entomol. Soc. Amer. 1960, 53, 61–78. [Google Scholar] [CrossRef]
- Buxton, P.A. British Diptera associated with fungi. 111: Plies of all families reared from about 150 species of fungi. Entomol. Mon. 1960, 96, 61–94. [Google Scholar]
- Põldmaa, K.; Jürgenstein, S.; Bahram, M.; Teder, T.; Kurina, O. Host diversity and trophic status as determinants of species richness and community composition of fungus gnats. Basic Appl. Ecol. 2015, 16, 46–53. [Google Scholar] [CrossRef]
- Põldmaa, K.; Kaasik, A.; Tammaru, T.; Kurina, O.; Jürgenstein, S.; Teder, T. Polyphagy on unpredictable resources does not exclude host specialization: Insects feeding on mushrooms. Ecology 2016, 97, 2824–2833. [Google Scholar] [CrossRef]
- Brandtberg, P.; Johansson, M.; Seeger, P. Effects of season and urea treatment on infection of stumps of Picea abies by Heterobasidion annosum in stands on former arable land. Scand. J. For. Res. 1996, 11, 261–268. [Google Scholar] [CrossRef]
- Gonthier, P.; Garbelotto, M.; Varese, G.C.; Nicolotti, G. Relative abundance and potential dispersal range of intersterility groups of Heterobasidion annosum in pure and mixed forests. Can. J. Bot. 2001, 79, 1057–1065. [Google Scholar] [CrossRef]
- Vollbrecht, G.; Johansson, U.; Eriksson, H.; Stendlid, J. Butt rot incidence, yield and growth pattern in a tree species experiment in southwestern Sweden. For. Ecol. Manag. 1995, 76.1, 87–93. [Google Scholar] [CrossRef]
- Greig, B.J.W. Field recognition and diagnosis of Heretobasidion annisum. In Heterobasidion Annosum: Biology, Ecology, Impact and Control; Woodward, S., Stenlid, J., Karjalainen, R., Hüttermann, A., Eds.; CAB International: Wallingford, UK, 1998; pp. 35–92. [Google Scholar]
- Schigel, D.S. Collecting and rearing fungivorous Coleoptera. Rev. d’Ecol. 2008, 63, 15–20. [Google Scholar]
- Querner, P. Combining pitfall traps and soil samples to collect Collembola for site scale biodiversity assessments. Appl. Soil Ecol. 2010, 45, 293–297. [Google Scholar] [CrossRef]
- Thorn, S.; Müller, J.; Bässler, C.; Gminder, A.; Brandl, R.; Heibl, C. Host abundance, durability, basidiome form and phylogenetic isolation determine fungivore species richness. Biol. J. Linn. Soc. 2015, 114, 699–708. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Large Heterobasidion spp. fruitbody.

Figure 2.
Tullgren funnels utilized for the collection of invertebrates.

Figure 3.
Accumulation curves showing the relationship between the cumulative number of invertebrate taxa and the number of collected samples.
Figure 3.
Accumulation curves showing the relationship between the cumulative number of invertebrate taxa and the number of collected samples.

Figure 4.
Venn diagram showing the diversity and overlap of invertebrate taxa in different substrates (sites as well as adults and larvae are combined).
Figure 4.
Venn diagram showing the diversity and overlap of invertebrate taxa in different substrates (sites as well as adults and larvae are combined).

Figure 5.
Ordination diagram based on principal component analysis of invertebrate communities from Heterobasidion spp. fruitbodies, Picea abies wood and soil samples. The size of each point reflects the relative richness of invertebrate taxa.
Figure 5.
Ordination diagram based on principal component analysis of invertebrate communities from Heterobasidion spp. fruitbodies, Picea abies wood and soil samples. The size of each point reflects the relative richness of invertebrate taxa.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Occurrence and relative abundance (%) of invertebrate taxa identified in Heterobasidion fruitbodies, Picea abies wood and soil samples. Absolute numbers for higher taxonomic groups are shown in parentheses.
Table 1.
Occurrence and relative abundance (%) of invertebrate taxa identified in Heterobasidion fruitbodies, Picea abies wood and soil samples. Absolute numbers for higher taxonomic groups are shown in parentheses.
| Invertebrate Taxa | Heterobasidion | Picea abies Wood | Soil | |||
|---|---|---|---|---|---|---|
| Fruitbodies | ||||||
| Subclass/Family | Adults | Larvae | Adults | Larvae | Adults | Larvae |
| Acari | (4111) | (1277) | (634) | |||
| Hypochthoniidae | 0.6 | - | 2.6 | - | 1.4 | - |
| Pthiracaridae | 0.9 | - | 4.8 | - | 2.2 | - |
| Euphthiracaridae | 0.2 | - | 5.9 | - | 0.3 | - |
| Crotonioidae | 0.7 | - | 0.1 | - | 1.4 | - |
| Nanhermanniidae | 0.1 | - | 0.3 | - | 0.4 | - |
| Damaeidae | 0.2 | - | 0.1 | - | 0.7 | - |
| Eremaeidae | 0.1 | - | 0.1 | - | - | - |
| Tenuialidae | 0.04 | - | 0.1 | - | 0.3 | - |
| Xenillidae | 0.02 | - | 0.2 | - | - | - |
| Carabodidae | 65.9 | - | 12.4 | - | 11.4 | - |
| Oppiidae | 7.3 | - | 39.2 | - | 32.7 | - |
| Scheloribatidae | - | - | - | - | 0.6 | - |
| Ceratozetidae | 0.8 | - | 5.3 | - | 34.9 | - |
| Galumnidae | 0.04 | - | 0.1 | - | 0.6 | - |
| Other 1 | 7.1 | - | 10.3 | - | 4.3 | - |
| Collembola | (309) | (156) | (24) | |||
| Hypogastruridae | 0.2 | - | 0.2 | - | 0.1 | - |
| Neanuridae | 4.2 | - | 6.4 | - | 2.3 | - |
| Onychiuridae | 0.8 | - | 1.5 | - | 0.4 | - |
| Entomobryidae | 0.1 | - | 1.2 | - | 0.3 | - |
| Isotomidae | 0.7 | - | 0.1 | - | 0.3 | - |
| Tomoceridae | 0.2 | - | 0.4 | - | - | - |
| Sminthuridae | 0.02 | - | 0.1 | - | - | - |
| Coleoptera | (170) | (62) | (12) | (29) | (7) | (3) |
| Anobiidae | 0.02 | - | - | - | - | - |
| Cantharidae | - | 17.8 | - | 16.7 | - | 40.0 |
| Carabidae | 0.1 | - | - | - | - | - |
| Ciidae | 3.0 | 11.9 | - | 1.9 | - | - |
| Cryptophagidae | - | - | - | - | 0.1 | - |
| Elateridae | 0.02 | 23.8 | - | 33.3 | - | - |
| Pselaphidae | - | - | 0.1 | - | 0.1 | - |
| Ptilidae | 0.1 | - | 0.5 | - | - | - |
| Silphidae | 0.1 | - | 0.1 | - | - | - |
| Staphylinidae | 0.2 | 7.9 | 0.1 | 1.9 | 0.7 | 20.0 |
| Diplopoda | (126) | (50) | (1) | |||
| Blaniulidae | 2.3 | - | 2.9 | - | 0.1 | - |
| Julidae | 0.02 | - | - | - | - | - |
| Polydesmidae | 0.3 | - | 0.3 | - | - | - |
| Diptera | (50) | (39) | (10) | (22) | (3) | (1) |
| Bibionidae | 0.02 | 0.9 | - | - | - | - |
| Brachycera | 0.1 | 15.8 | - | 7.4 | - | 20.0 |
| Cecidomyiidae | 0.04 | 2.9 | - | - | - | - |
| Ceratopogonidae | 0.1 | - | - | 3.7 | - | - |
| Chiromidae | 0.4 | 14.8 | 0.3 | 25.9 | - | - |
| Drosophilidae | - | - | 0.1 | - | - | - |
| Limoniidae | - | - | - | 3.7 | - | - |
| Psychodidae | 0.02 | - | - | - | - | - |
| Rhagionidae | - | - | - | - | 0.1 | - |
| Sciaridae | 0.3 | 0.9 | 0.3 | - | 0.3 | - |
| Stratiomyidae | 0.02 | 2.9 | - | - | - | - |
| Haplotaxida | (41) | (18) | (8) | |||
| Lumbricidae | 0.4 | - | 0.6 | - | 0.7 | - |
| Enchytraeidae | 0.4 | - | 0.5 | - | 0.4 | - |
| Thysanoptera | 0.7 (35) | - | 0.6 (10) | - | 0.1 (1) | - |
| Hymenoptera | (15) | (7) | (8) | |||
| Parasitica | 0.3 | - | 0.4 | - | 1.2 | - |
| Formicidae | 0.02 | - | - | - | - | - |
| Aranea | 0.2 (10) | - | 0.5 (8) | - | 0.4 (3) | - |
| Gastropoda | (6) | (6) | (1) | |||
| Arionidae | 0.02 | - | - | - | - | - |
| Clausiliidae | 0.02 | - | - | - | 0.1 | - |
| Zonitiidae | 0.1 | - | 0.4 | - | - | - |
| Chilopoda | 0.1 (5) | - | 0.2 (3) | - | 0.1 (1) | - |
| Hemiptera | 0.04 (2) | - | 0.1 (1) | - | 0.3 (2) | - |
| Lepidoptera | (2) | (3) | (1) | |||
| Tineidae | 0.04 | - | - | 5.5 | - | 20.0 |
| Isopoda | (2) | (2) | ||||
| Trichoniscidae | 0.04 | - | 0.1 | - | - | - |
| Opiliones | 0.02 (1) | - | - | - | - | |
| Protura | 0.02 (1) | - | - | - | - | - |
| Pseudoscorpionida | - | - | 0.6 (10) | - | 0.3 (2) | - |
1 Acari individuals that were not identified to family level.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Legzdina, L.; Spungis, V.; Burnevica, N.; Gaitnieks, T.; Menkis, A. Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil. Forests 2021, 12, 1100. https://doi.org/10.3390/f12081100
AMA Style
Legzdina L, Spungis V, Burnevica N, Gaitnieks T, Menkis A. Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil. Forests. 2021; 12(8):1100. https://doi.org/10.3390/f12081100
Chicago/Turabian StyleLegzdina, Liva, Voldemars Spungis, Natalija Burnevica, Talis Gaitnieks, and Audrius Menkis. 2021. "Invertebrates in Fruitbodies of Heterobasidion spp., Infected Picea abies Logs and Adjacent Soil" Forests 12, no. 8: 1100. https://doi.org/10.3390/f12081100
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.