
Carbon Dioxide, Methane and Nitrous Oxide Fluxes from Tree Stems in Silver Birch and Black Alder Stands with Drained and Naturally Wet Peat Soils
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
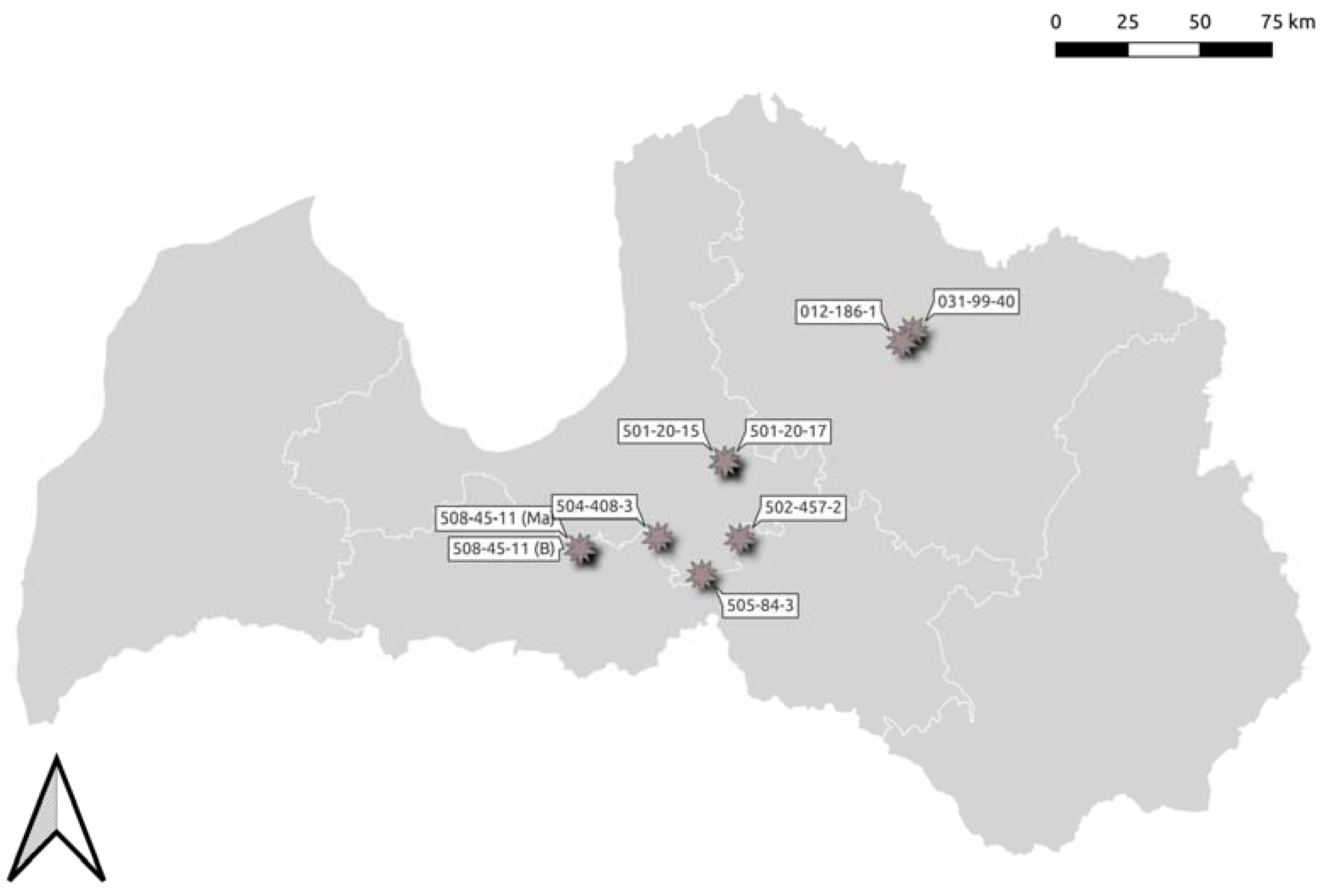
2.1. Study Sites
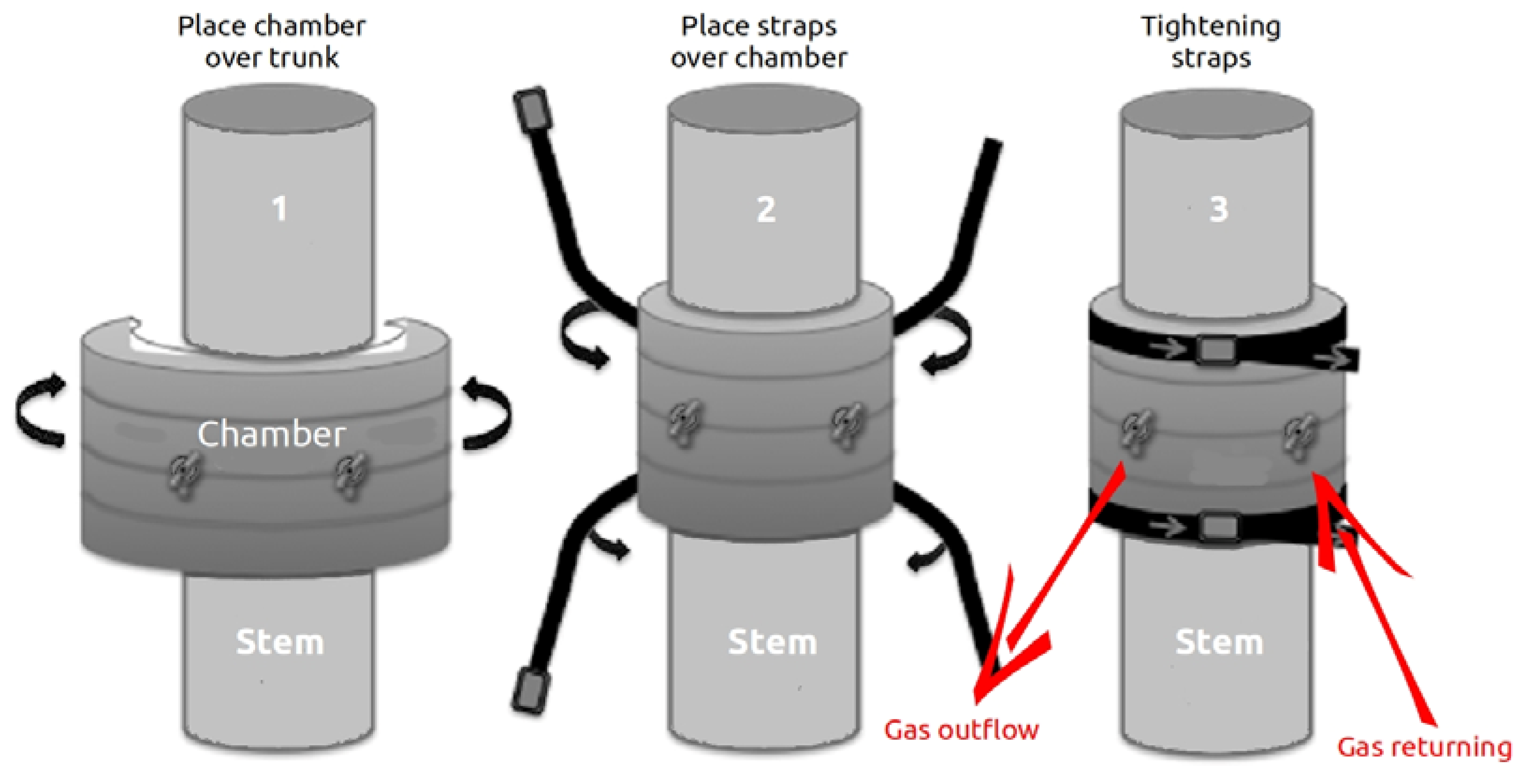
2.2. Closed Chamber Design and Gas Flux Exchange Measurements from Tree Stems
- P—air pressure in the chamber, assumed constant 101,300 Pa;
- M—molar mass of measured gases (CO2—44.01 g·mol−1; CH4—16.04 g·mol−1; 44.01 N2O—44.01 g·mol−1);
- δv—slope of regression representing gas concentration changes per hour;
- V—chamber volume, m3 (Table 3);
- —recalculation coefficient (0.27 for CO2, 0.75 for CH4 and 0.64 for N2O);
- A—chamber surface area, m2;
- T—soil temperature, K;
- t—measurement time, hours;
- R—universal gas constant (8.3143 m3·Pa·K−1·mol−1).
2.3. Soil Sampling and Analysis
2.4. Statistical Analysis
3. Results
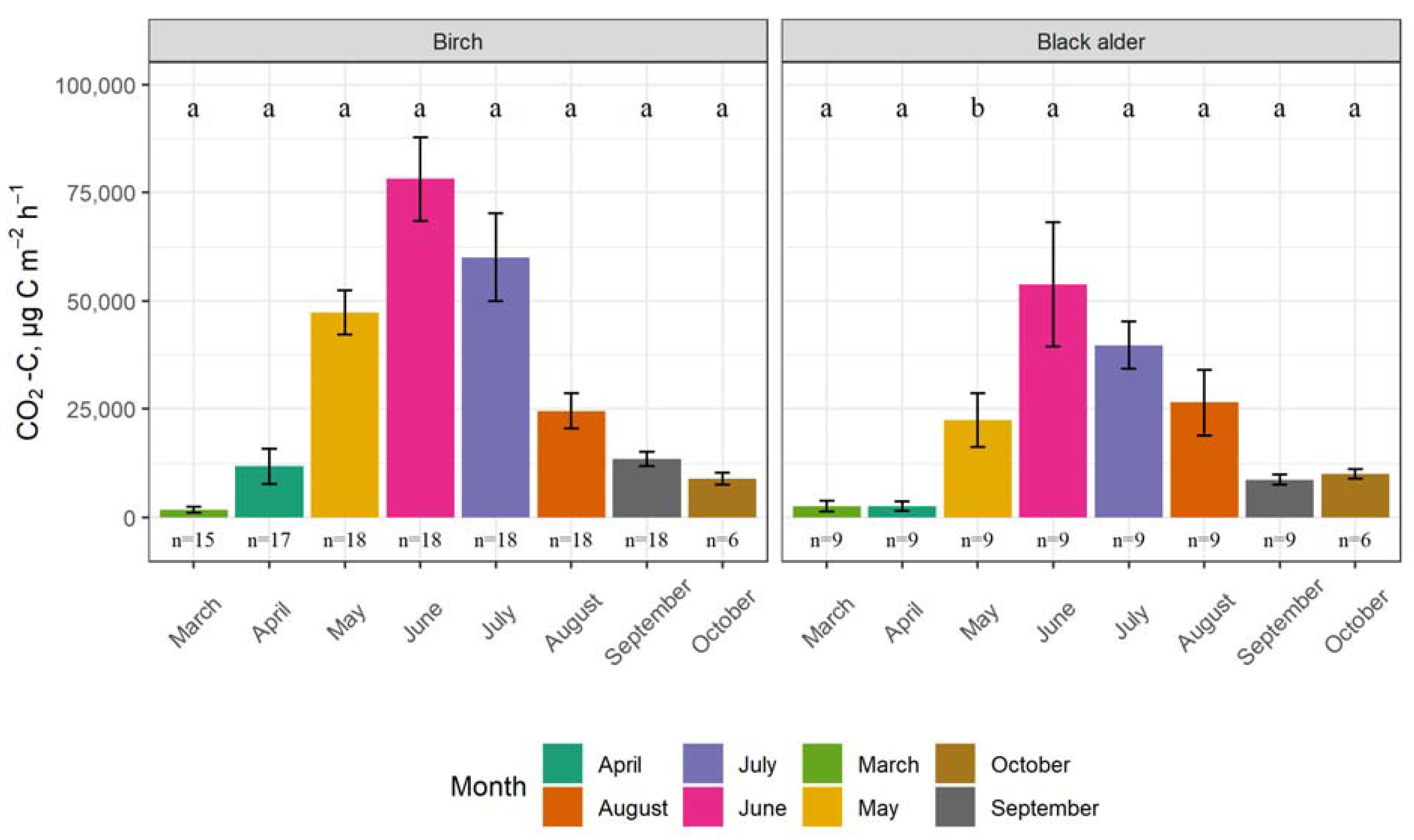
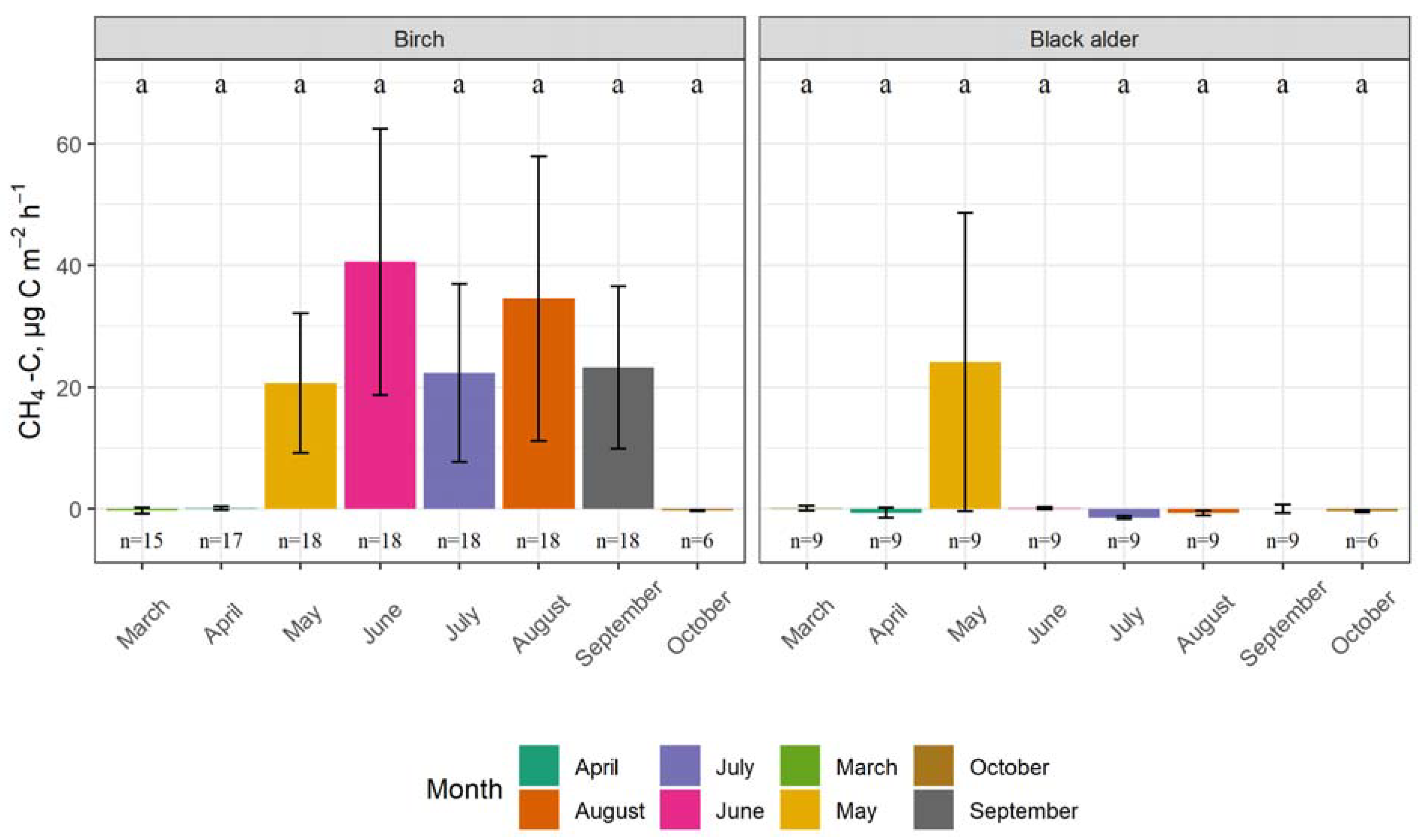
3.1. Seasonality of GHG Fluxes and the Impact of Tree Species
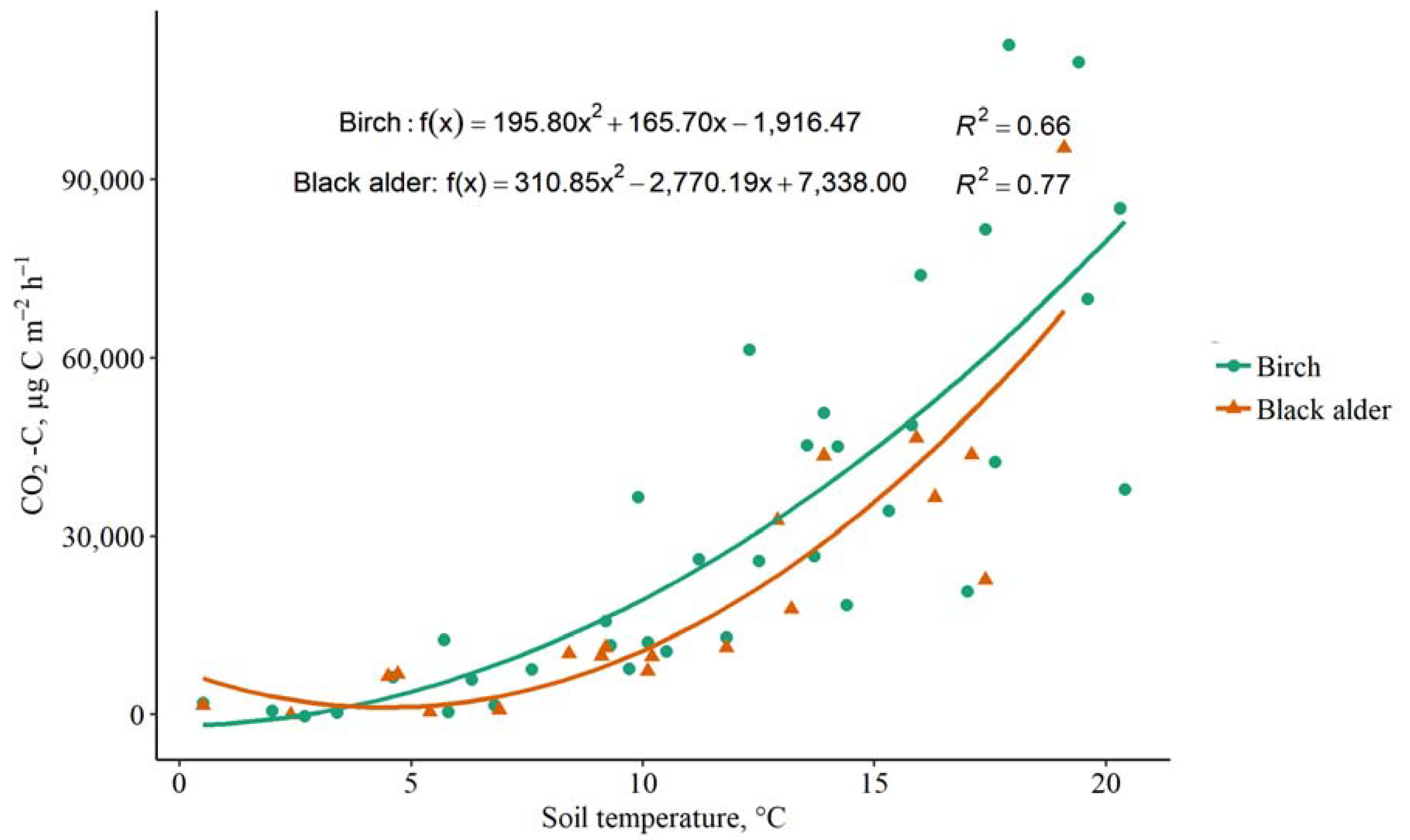
3.2. The Impact of Soil Temperature on CO2, CH4 and N2O Fluxes
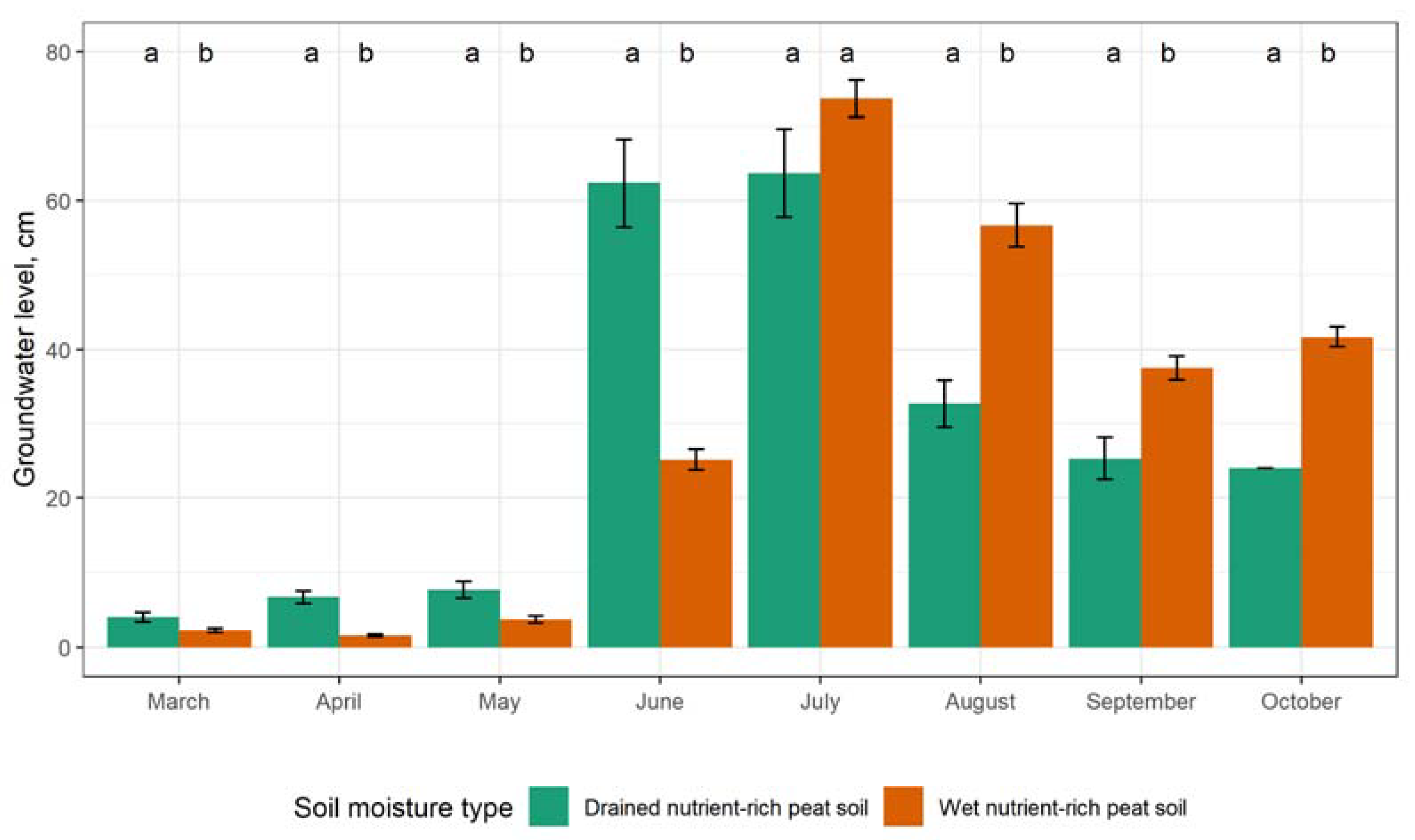
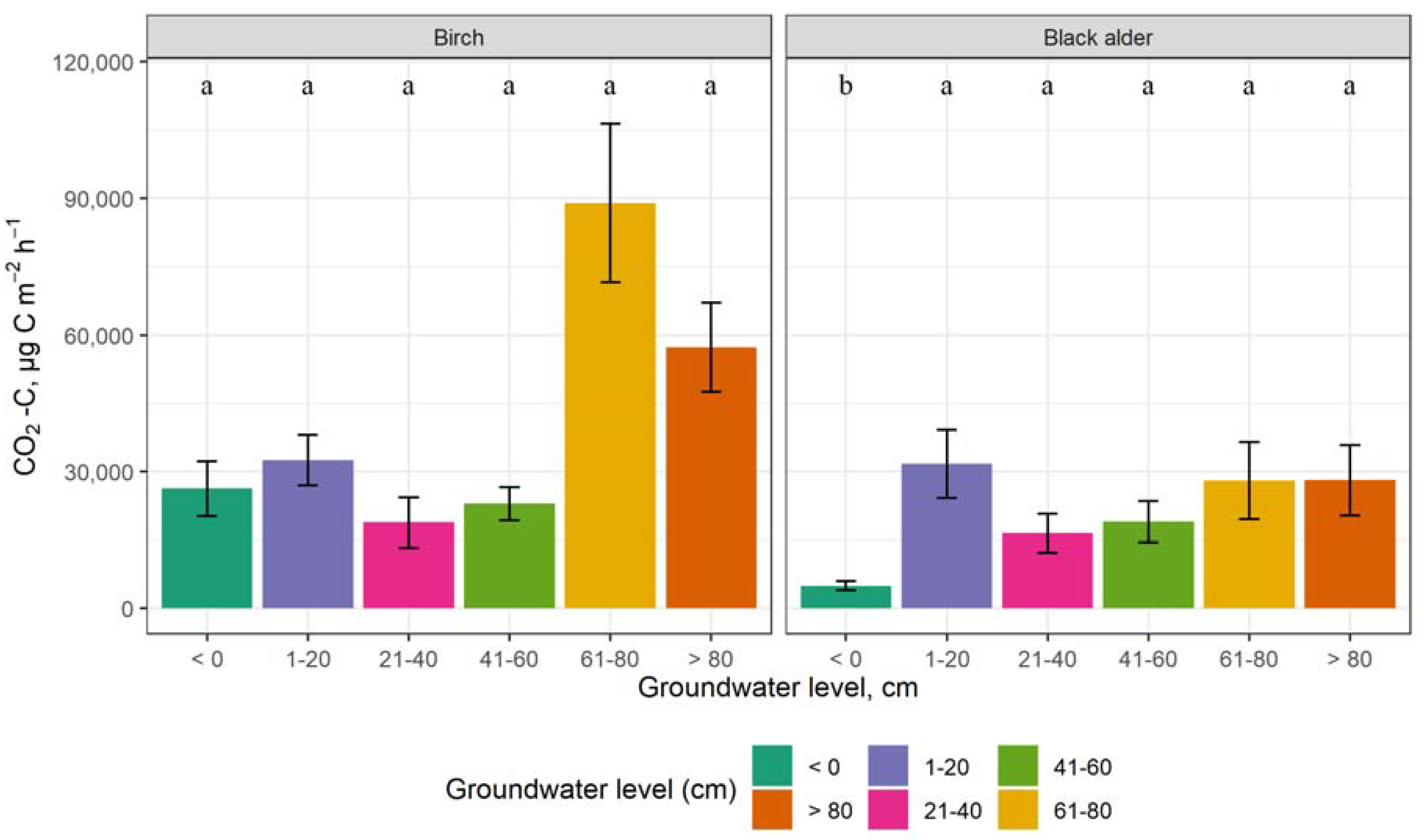
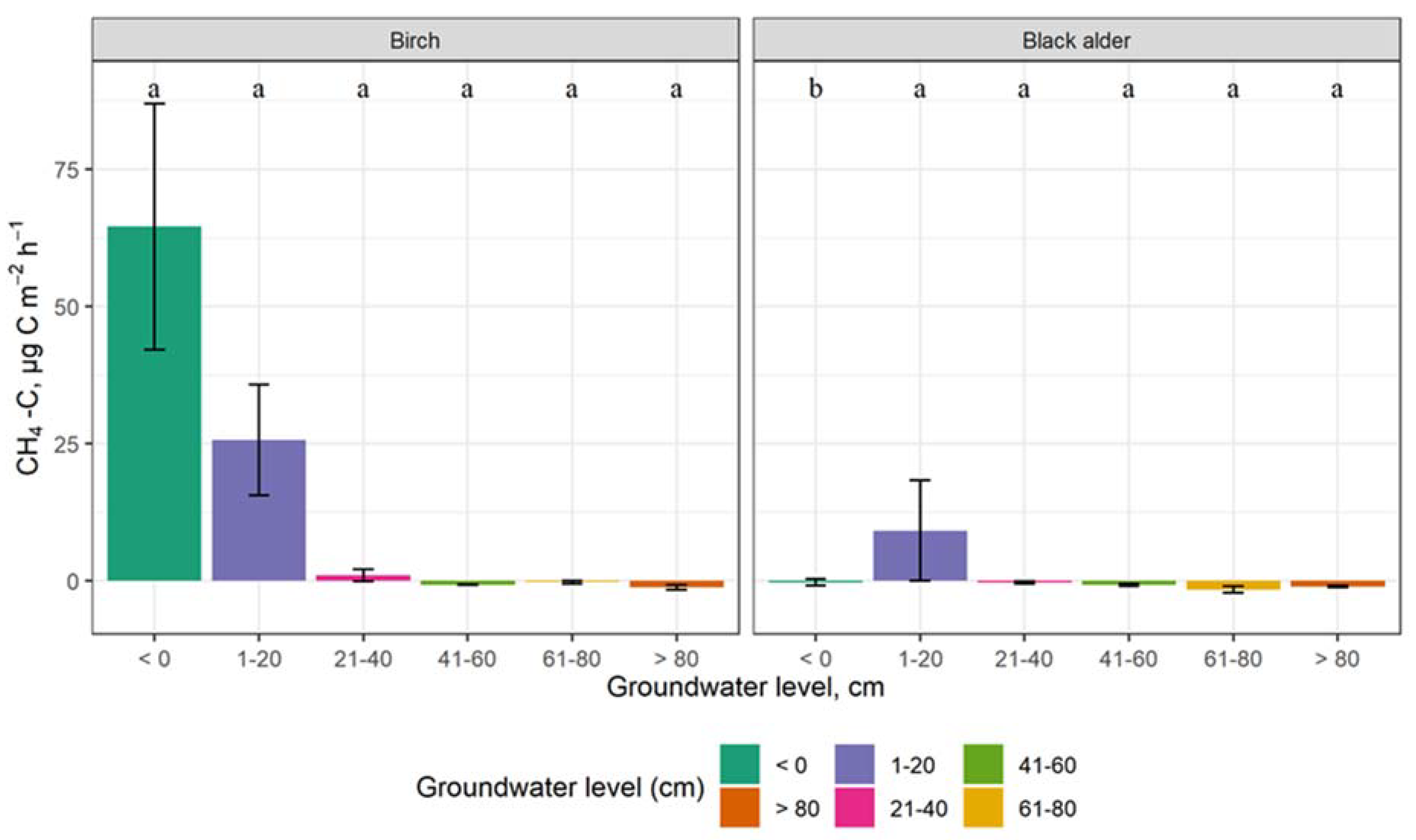
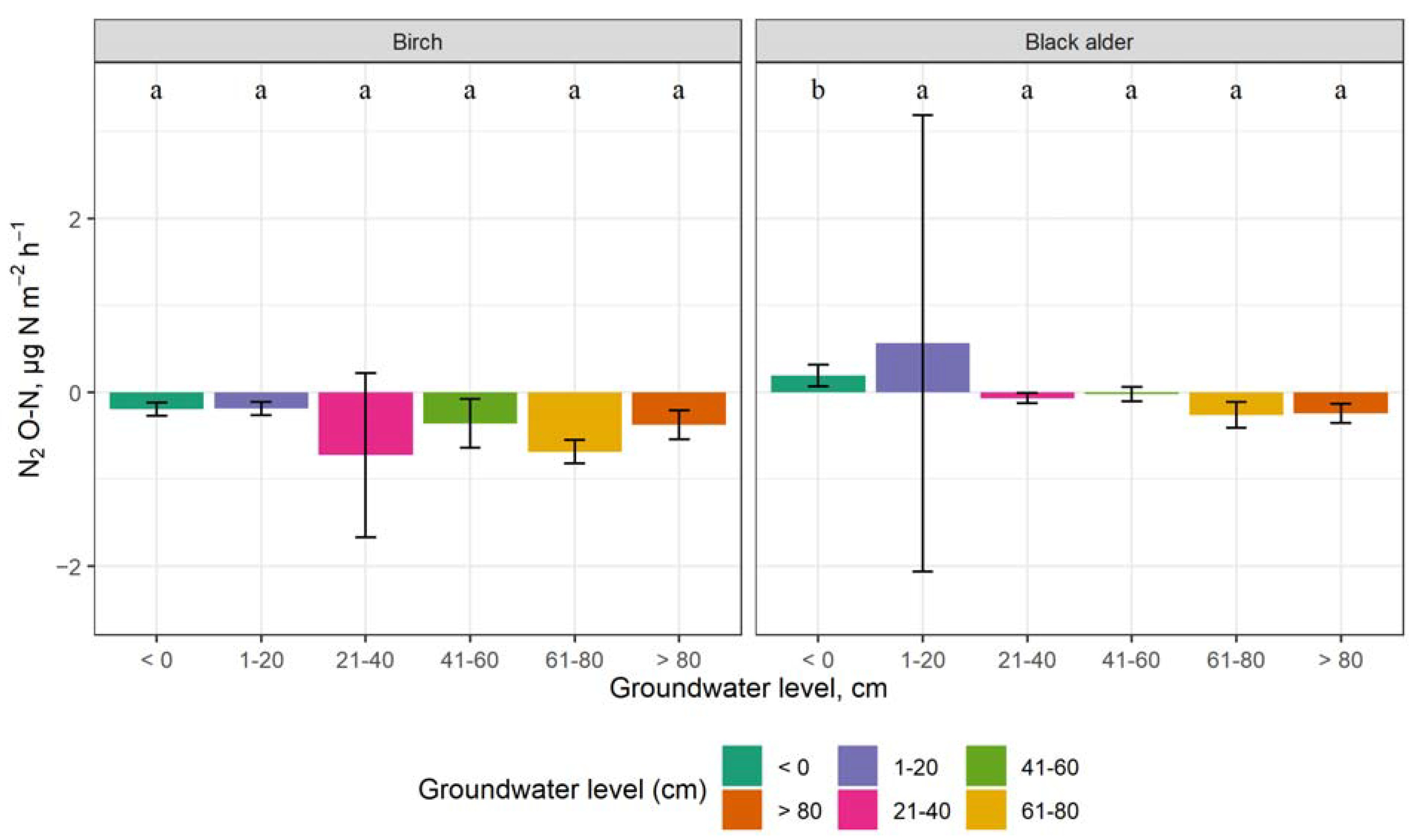
3.3. The Impact of Groundwater Level on GHG Emissions
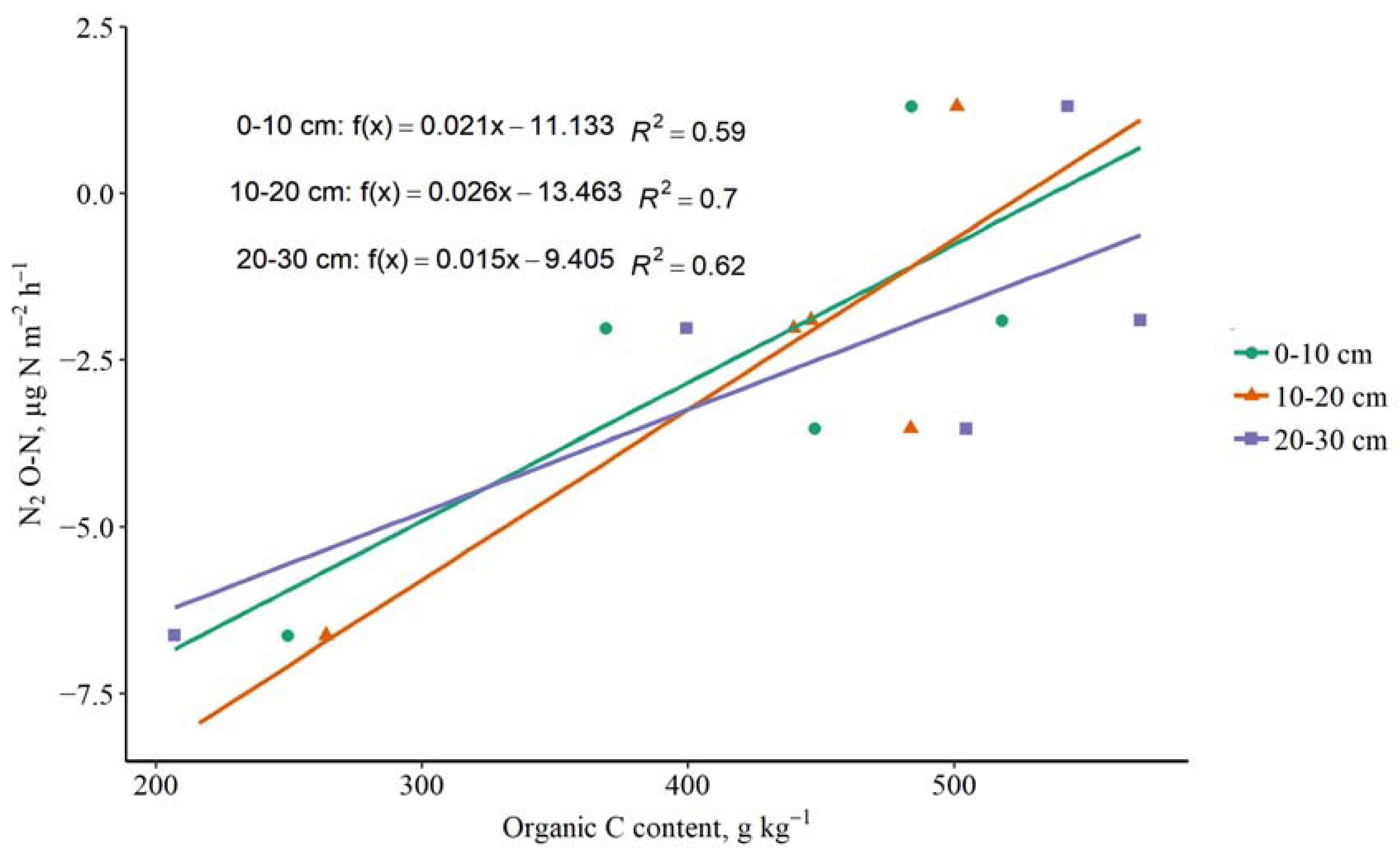
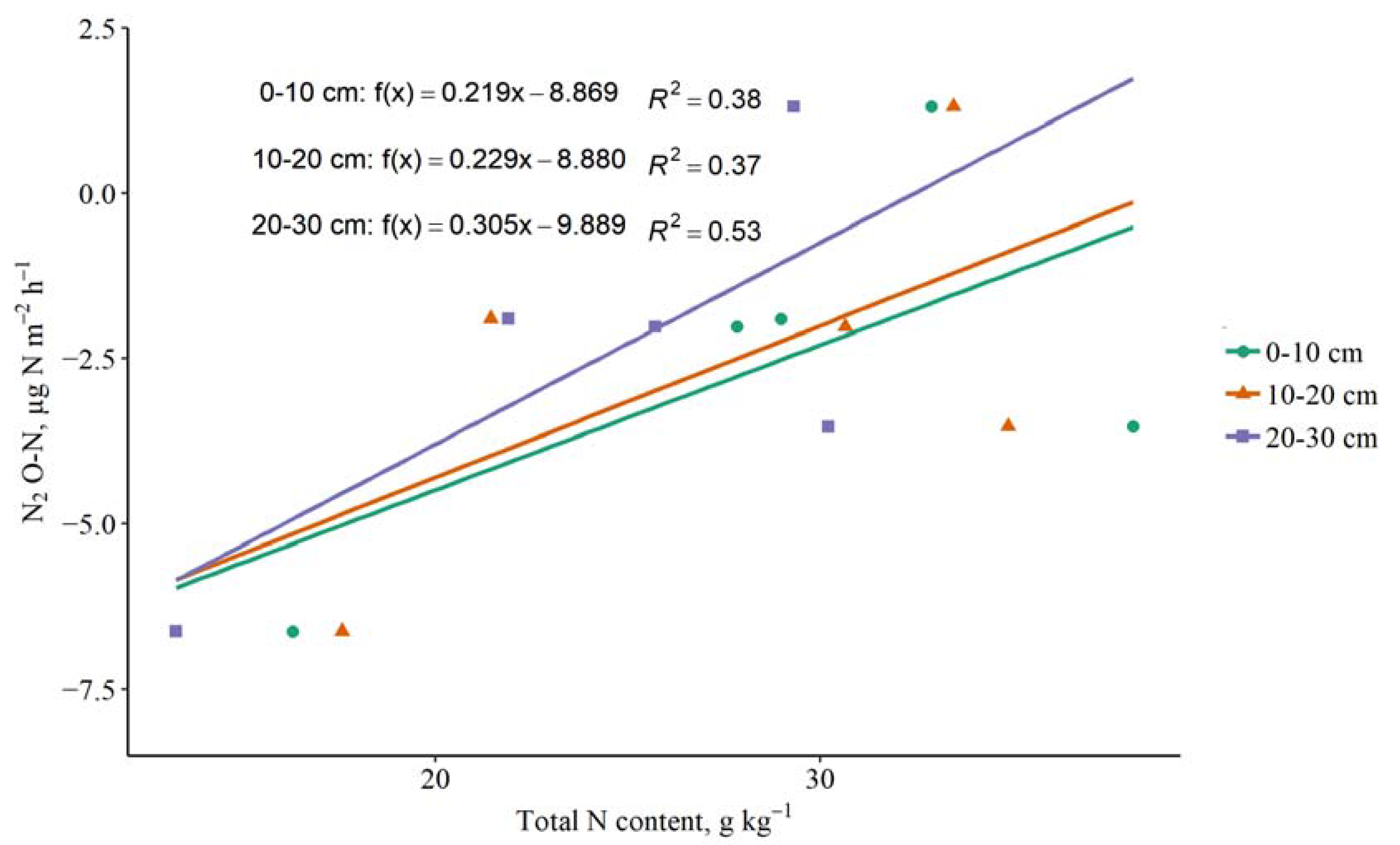
3.4. The Impact of Soil C and N Contents on GHG Emissions
4. Discussion
4.1. Seasonality of GHG Fluxes and the Impact of Tree Species
4.2. The Impact of Soil and Air Temperature
4.3. The Impact of Groundwater Level
4.4. The Impact of Soil C and N Contents
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Covey, K.R.; Megonigal, J.P. Methane production and emissions in trees and forests. New Phytol. 2019, 222, 35–51. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.; Phillips, O.; Jackson, R. The Structure, distribution and biomass of the World’s forests. Annu. Rev. Ecol. Syst. 2013, 44, 593–622. [Google Scholar] [CrossRef] [Green Version]
- Latvijas Valsts Mežzinātnes Institūts “Silava”. Available online: www.silava.lv (accessed on 23 January 2023).
- Valsts Meža Dienests. Available online: www.vmd.gov.lv (accessed on 23 January 2023).
- Latvia’s National Inventory Report Submission under UNFCCC and the Kyoto Protocol Common Reporting Formats (CRF) 1990–2017. Available online: https://unfccc.int/documents/194812 (accessed on 23 January 2023).
- Saikawa, E.; Prinn, R.G.; Dlugokencky, E.; Ishijima, K.; Dutton, G.S.; Hall, B.D.; Langenfelds, R.; Tohjima, Y.; Machida, T.; Manizza, M.; et al. Global and regional emissions estimates for N2O. Atmos. Chem. Phys. 2014, 14, 4617–4641. [Google Scholar] [CrossRef] [Green Version]
- Crowther, T.W.; Glick, H.B.; Covey, K.R.; Bettigole, C.; Maynard, D.S.; Thomas, S.M.; Smith, J.R.; Hintler, G.; Duguid, M.C.; Amatulli, G.; et al. Mapping tree density at a global scale. Nature 2015, 525, 201–205. [Google Scholar] [CrossRef]
- Wang, Z.-P.; Han, S.-J.; Li, H.-L.; Deng, F.-D.; Zheng, Y.-H.; Liu, H.-F.; Han, X.-G. Methane production explained largely by water content in the heartwood of living trees in upland forests. J. Geophys. Res. Biogeosci. 2017, 122, 2479–2489. [Google Scholar] [CrossRef]
- Pitz, S.L.; Megonigal, J.P.; Chang, C.-H.; Szlavecz, K. Methane fluxes from tree stems and soils along a habitat gradient. Biogeochemistry 2018, 137, 307–320. [Google Scholar] [CrossRef]
- Yip, D.Z.; Veach, A.M.; Yang, Z.K.; Cregger, M.A.; Schadt, C.W. Methanogenic Archaea dominate mature heartwood habitats of Eastern Cottonwood (Populus deltoides). New Phytol. 2019, 222, 115–121. [Google Scholar] [CrossRef]
- Maier, M.; Machacova, K.; Lang, F.; Svobodova, K.; Urban, O. Combining soil and tree-stem flux measurements and soil gas profiles to understand CH4 pathways in Fagus sylvatica forests. J. Plant. Nutr. Soil Sci. 2018, 181, 31–35. [Google Scholar] [CrossRef] [Green Version]
- Ward, N.D.; Indivero, J.; Gunn, C.; Wang, W.; Bailey, V.; McDovell, N.G. Longitudinal Gradients in Tree Stem Greenhouse Gas Concentrations Across Six Pacific Northwest Coastal Forests. J. Geophys. Res. 2019, 124, 1401–1412. [Google Scholar] [CrossRef] [Green Version]
- Sjögersten, S.; Siegenthaler, A.; Lopez, O.R.; Aplin, P.; Turner, B.; Gauci, V. Methane emissions from tree stems in neotropical peatlands. New Phytol. 2020, 225, 769–781. [Google Scholar] [CrossRef] [Green Version]
- Pangala, S.R.; Hornibrook, E.R.C.; Gowing, D.J.; Gauci, V. The contribution of trees to ecosystem methane emissions in a temperate forested wetland. Glob. Chang. Biol. 2015, 21, 2642–2654. [Google Scholar] [CrossRef]
- Schindler, T.; Mander, Ü.; Machachova, K.; Espenberg, M.; Krasnov, D.; Escuer-Gatius, J.; Verb, G.; Pärn, J.; Sooasaar, K. Short-term flooding increases CH4 and N2O emissions from trees in a riparian forest soil-stem continuum. Sci. Rep. 2020, 10, 3204. [Google Scholar] [CrossRef] [Green Version]
- Mander, Ü.; Krasnova, A.; Schindler, T.; Megonigal, J.P.; Escuer-Gatius, J.; Espenberg, M.; Machaccova, K.; Maddison, M.; Pärn, J.; Ranniku, R.; et al. Long-term Dynamics of soil, tree stem and ecosystem methane fluxes in a riparian forest. Sci. Total Environ. 2022, 809, 151723. [Google Scholar] [CrossRef]
- Bužková, R.; Acosta, M.; Dařenová, E.; Pokorný, R.; Pavelka, M. Environmental factors influencing the relationship between stem CO2 efflux and sap flow. Trees 2015, 29, 333–343. [Google Scholar] [CrossRef]
- Teskey, R.O.; McGuire, M.A. Carbon dioxide transport in xylem causes errors in estimation of rates of respiration in stems and branches of trees. Plant Cell Environ. 2002, 25, 1571–1577. [Google Scholar] [CrossRef]
- Pfanz, H.; Aschan, G. The existence of bark and stem photosynthesis and its significance for the overall carbon gain: An eco-physiological and ecological approach. Prog. Bot. 2000, 62, 477–510. [Google Scholar]
- Pfanz, H.; Aschan, G.; Langenfeld-Heyser, R.; Wittmann, C.; Loose, M. Ecology and ecophysiology of tree stems: Corticular andwood photosynthesis. Naturwissenschaften 2002, 89, 147–162. [Google Scholar]
- Barba, J.; Bradford, M.A.; Brewer, P.E.; Bruhn, D.; Covey, K.; Van Haren, J.; Megonigal, J.P.; Mikkelsen, T.N.; Pangala, S.R.; Pihlatie, M.; et al. Methane emissions from tree stems: A new frontier in the global carbon cycle. New Phytol. 2019, 222, 18–28. [Google Scholar] [CrossRef]
- Vargas, R.; Barba, J. Greenhouse Gas Fluxes From Tree Stems. Trends Plant Sci. 2019, 24, 296–299. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Buchholz, J.; Rennenberg, H. Emission of Methane and Nitrous Oxide by Australian Mangrove Ecosystems. Plant Biol. 2023, 5, 423–431. [Google Scholar] [CrossRef]
- Rusch, H.; Rennenberg, H. Black alder (Alnus glutinosa (L.) Gaertn.) trees mediate methane and nitrous oxide emission from the soil to the atmosphere. Plant Soil 1998, 201, 1–7. [Google Scholar] [CrossRef]
- Welch, B.; Gauci, V.; Sayer, E.J. Tree stem bases are sources of CH4 and N2O in a tropical forest on upland soil during the dry to wet season transition. Glob. Change Biol. 2019, 25, 361–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Pinés, E.; Heras, P.; Gasche, R.; Rubio, A.; Rennenberg, H.; Butterbach-Bahl, K.; Kiese, R. Nitrous oxide emissions from stems of ash (Fraxinus angustifolia Vahl) and European beech (Fagus sylvatica L.). Plant Soil 2016, 398, 35–45. [Google Scholar] [CrossRef]
- Wen, Y.; Corre, M.D.; Rachow, C.; Chen, L.; Veldkamp, E. Nitrous oxide emissions from stems of alder, beech and spruce in a temperate forest. Plant Soil 2017, 420, 423–434. [Google Scholar] [CrossRef]
- Oliveira, R.S.; Castro, P.M.L.; Dodd, J.C.; Vosátka, M. Synergistic effect of Glomus intraradices and Frankia spp. on the growth and stress recovery of Alnus glutinosa in an alkaline anthropogenic sediment. Chemosphere 2005, 60, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Rochet, J.; Manzi, S.; Jargeat, P.; Gryta, H.; Moreau, P.; Gardes, M. What determines Alnus-associated ectomycorrhizal community diversity and specificity? A comparison of host and habitat effects at a regional scale. New Phytol. 2013, 198, 1228–1238. [Google Scholar]
- Huth, V.; Hoffmann, M.; Bereswill, S.; Popova, Y.; Zak, D.; Augustin, J. The climate warming effect of a fen peat meadow with fluctuating water table is reduced by young alder trees. Mires Peat 2018, 21, 1–18. [Google Scholar]
- Minkkinen, K.; Ojanen, P.; Koskinen, M.; Penttilä, T. Nitrous oxide emissions of undrained, forestry-drained, and rewetted boreal peatlands. For. Ecol. Manag. 2020, 478, 118494. [Google Scholar] [CrossRef]
- Smart, D.R.; Bloom, A.J. Wheat leaves emit nitrous oxide during nitrate assimilation. Proc. Natl. Acad. Sci. USA 2021, 98, 7875–7878. [Google Scholar] [CrossRef] [Green Version]
- Machacova, K.; Vaino, E.; Urban, O.; Pihlatie, M. Seasonal dynamics of stem N2O exchange follow the physiological activity of boreal trees. Nat. Commun. 2019, 10, 4989. [Google Scholar] [CrossRef] [Green Version]
- Latvijas Vides, Ģeoloģijas un Meteoroloģijas Centrs. Available online: https://videscentrs.lvgmc.lv/ (accessed on 26 January 2023).
- Nacionālā Enciklopēdija. Available online: www.enciklopedija.lv (accessed on 26 January 2023).
- LVS EN ISO/IEC 17025:2018; General Requirements for the Competence of Testing and Calibration Laboratories. 3rd ed. ISO: Geneva, Switzerland, 2017; p. 30.
- LVS ISO 11464:2006; Soil Quality—Pretreatment of Samples for Physico-Chemical Analysis. 2nd ed. ISO: Geneva, Switzerland, 2006; p. 11.
- LVS ISO 10390:2021; Soil, Treated Biowaste and Sludge—Determination of pH. 3rd ed. ISO: Geneva, Switzerland, 2021; p. 8.
- LVS ISO 10694:2006 A/L; Soil Quality—Determination of Organic and Total Carbon after Dry Combustion (Elementary Analysis). ISO: Geneva, Switzerland, 2006.
- LVS ISO 13878:1998; Soil Quality—Determination of Total Nitrogen Content by Dry Combustion (“Elemental Analysis”). 1st ed. ISO: Geneva, Switzerland, 1998; p. 5.
- Barba, J.; Poyatos, R.; Vargas, R. Automated measurements of greenhouse gases fluxes from tree stems and soils: Magnitudes, patterns and drivers. Sci. Rep. 2019, 9, 4005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindler, T.; Machacova, K.; Mander, Ü.; Escuer-Gatius, J.; Soosaar, K. Diurnal tree stem CH4 and N2O flux dynamics from a riparian alder forest. Forests 2021, 12, 863. [Google Scholar] [CrossRef]
- Ranniku, R.; Schindler, T.; Escuer-Gatius, J.; Mander, Ü.; Machacova, K.; Soosaar, K. Greenhouse Gas Dynamics in a Drained Peatland Forest: Annual CH4 and N2O Fluxes from Tree Stems and Soil. In Proceedings of the EGU General Assembly, Vienna, Austria, 23–27 May 2022. [Google Scholar]
- Vainio, E.; Haikarainen, I.P.; Machacova, K.; Putkinen, A.; Santalahti, M.; Koskinen, M.; Fritze, M.; Tuomivirta, T.; Pihlatie, M. Soil-tree-atmosphere CH4 flux dynamics of boreal birch and spruce trees during spring leaf-out. Plant Soil 2022, 478, 391–407. [Google Scholar] [CrossRef]
- Ojanen, P.; Minkkinen, K.; Alm, J.; Penttilä, T. Soil–atmosphere CO2, CH4 and N2O fluxes in boreal forestry-drained peatlands. For. Ecol. Manag. 2010, 260, 411–421. [Google Scholar] [CrossRef]
- Köhn, D.; Günther, A.; Schwabe, I.; Jurasinski, G. Short-lived peaks of stem methane emissions from mature black alder (Alnus glutinosa (L.) Gaertn.)—Irrelevant for ecosystem methane budgets? Plant-Environ. Interact. 2021, 2, 16–27. [Google Scholar] [CrossRef]
- Vanags-Duka, M.; Bārdule, A.; Butlers, A.; Upenieks, E.M.; Lazdiņš, A.; Purviņa, D.; Līcīte, I. GHG Emissions from Drainage Ditches in Peat Extraction Sites and Peatland Forests in Hemiboreal Latvia. Land 2022, 11, 2233. [Google Scholar] [CrossRef]
- Machacova, K.; Borak, L.; Agyei, T.; Schindler, T.; Soosaar, K.; Mander, Ü.; Ah-Peng, C. Trees as net sinks for methane (CH4) and nitrous oxide (N2O) in the lowland tropical rain forest on volcanic Réunion Island. New Phytol. 2021, 229, 1983–1994. [Google Scholar] [CrossRef]
- Hölttä, T.; Kolari, P. Interpretation of stem CO2 efflux measurements. Tree Physiol. 2009, 29, 1447–1456. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Calcerrada, J.; López, R.; Salomón, R.; Gordaliza, G.G.; Valbuena-Carabaña, M.; Oleksyn, J.; Luis, G. Stem CO2 efflux in six co-occurring tree species: Underlying factors and ecological implications. Plant Cell Environ. 2015, 38, 1104–1115. [Google Scholar] [CrossRef]
- Obidziński, A. Black alder (Alnus glutinosa Gaertn.) as pioneer species in regeneration of fresh oak-linden-hornbeam forest (Tilio-Carpientum typicum Traczyk 1962) in Białowieża Forest (east Poland). Pol. J. Ecol. 2004, 52, 533–551. [Google Scholar]
- Sakalli, A. Simulation of potential distribution and migration of Alnus spp. under climate change. Appl. Ecol. Environ. Res. 2017, 15, 1039–1070. [Google Scholar] [CrossRef]
- Chanton, J.P.; Whiting, G.J.; Happell, J.D.; Gerard, G. Contrasting rates and diurnal patterns of methane emission from emergent aquatic macrophytes. Aquat. Bot. 1993, 46, 111–128. [Google Scholar] [CrossRef]
- Kim, J.; Verma, S.B.; Billesbach, D.P. Seasonal variation in methane emission from a temperate Phragmites-dominated marsh: Effect of growth stage and plant-mediated transport. Glob. Chang. Biol. 1999, 5, 433–440. [Google Scholar] [CrossRef]
- Pangala, S.R.; Gowing, D.J.; Hornibrook, E.R.C.; Gauci, V. Controls on methane emissions from Alnus glutinosa saplings. New Phytol. 2014, 201, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Sevanto, S.; Suni, T.M.; Pumpanen, J.; Grönholm, T.; Kolari, P.; Nikinmaa, E.; Hari, P.; Vesala, T. Wintertime photosynthesis and water uptake in a boreal forest. Tree Physiol. 2006, 26, 749–757. [Google Scholar] [CrossRef] [Green Version]
- Kolari, P.; Kulmala, L.; Pumpanen, J.; Launiainen, S.; Ilvesniemi, H.; Hari, P.; Nikinmaa, E. CO2 exchange and component CO2 fluxes of a boreal Scots pine forest. Boreal Environ. Res. 2009, 14, 761–783. [Google Scholar]
- Acosta, M.; Pavelka, M.; Pokorony, R.; Janouš, D.; Marek, M.V. Seasonal variation in CO2 efflux of stems and branches of Norway spruce trees. Ann. Bot. 2008, 101, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Wei, X.; Fan, H.; Wu, J.; Hao, J.; Pei, Q. Disentangling critical drivers of stem CO2 efflux from Pinus elliottii trees in Subtropical China. Agric. For. Meteorol. 2017, 237–238, 296–302. [Google Scholar] [CrossRef]
- Guidolotti, G.; Rey, A.; D’Andrea, E.; Matteucci, G.; De Angelis, P. Effect of environmental variables and stand structure on ecosystem respiration components in a Mediterranean beech forest. Tree Physiol. 2013, 33, 960–972. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.P.; Zhang, W.J.; Hu, C.S.; Tang, X.G. Soil greenhouse gas fluxes from different tree species on Taihang Mountain, North China. Biogeosciences 2014, 11, 1649–1666. [Google Scholar] [CrossRef] [Green Version]
- Zhao, K.; Fahey, T.J.; Wanh, X.; Warg, J.; He, F.; Fan, C.; Jia, Z.; Li, X. Effect of thinning intensity on the stem CO2 efflux of Larix principis-rupprechtii Mayr. For. Ecosyst. 2021, 8, 63. [Google Scholar] [CrossRef]
- Gauci, V.; Hornibrook, E.; Davis, J.; Dise, N. Woody stem methane emission in mature wetland alder trees. Atmos. Environ. 2010, 44, 2157–2160. [Google Scholar] [CrossRef] [Green Version]
- Terazawa, K.; Ishizuka, S.; Sakata, T.; Yamada, K.; Takahashi, M. Methane emissions from stems of Fraxinus mandshurica var. japonica trees in a floodplain forest. Soil. Biol. Biochem. 2007, 39, 2689–2692. [Google Scholar] [CrossRef]
- Torga, R.; Mander, Ü.; Soosaar, K.; Kupper, P.; Tullus, A.; Rosenvald, K.; Ostonen, I.; Kutti, S.; Jaagus, J.; Sober, J.; et al. Weather extremes and tree species shape soil greenhouse gas fluxes in an experimental fast-growing deciduous forest of air humidity manipulation. Ecol. Eng. 2017, 106, 369–377. [Google Scholar] [CrossRef]
- Leppelt, T.; Dechow, R.; Gebbert, S.; Freibauer, A.; Lohila, A.; Augustin, J.; Drösler, M.; Fiedler, S.; Glatzel, S.; Höper, H.; et al. Nitrous oxide emission hotspots from organic soils in Europe. Biogeosci. Discuss 2014, 11, 9135–9182. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Sample Plot | Tree Species | Soil Moisture Condition | Location, WGS84 | Stand Age, Years | Diameter at Breast (1.3 m) Height, cm | The Average Tree Height, m | |
|---|---|---|---|---|---|---|---|
| X | Y | ||||||
| 1 | Silver birch | Drained | 57.3218 | 26.0641 | 20 | 15 | 18 |
| 2 | Silver birch | Drained | 56.6873 | 25.0482 | 30 | 12 | 17 |
| 3 | Silver birch | Drained | 56.6942 | 24.5836 | 59 | 27 | 21 |
| 4 | Silver birch | Wet | 57.2906 | 25.9987 | 60 | 19 | 18 |
| 5 | Silver birch | Wet | 56.9289 | 24.9666 | 70 | 22 | 22 |
| 6 | Black alder | Wet | 56.6596 | 24.1421 | 23 | 9 | 11 |
| 7 | Black alder | Wet | 56.9280 | 56.9280 | 53 | 22 | 24 |
| 8 | Black alder | Wet | 56.5737 | 56.5737 | 72 | 29 | 29 |
| Tree Species | Diameter of Trees at Breast (1.3 m) Height, cm | Tree Height, m | ||||
|---|---|---|---|---|---|---|
| Tree 1 | Tree 2 | Tree 3 | Tree 1 | Tree 2 | Tree 3 | |
| Silver birch | 20.5 | 16.4 | 11.5 | 18.5 | 17.8 | 16.0 |
| Silver birch | 20.3 | 14.7 | 12.7 | 21.4 | 18.2 | 19.4 |
| Silver birch | 28.6 | 25.9 | 19.5 | 22.7 | 22.4 | 18.2 |
| Silver birch | 14.5 | 10.6 | 9.2 | 12.6 | 12.2 | 12.0 |
| Silver birch | 20.5 | 18.0 | 12.9 | 22.4 | 21.3 | 20.2 |
| Silver birch | 29.9 | 21.0 | 14.0 | 25.4 | 23.6 | 20.8 |
| Black alder | 11.3 | 11.1 | 8.7 | 11.4 | 11.4 | 9.7 |
| Black alder | 24.3 | 19.3 | 14.2 | 23.7 | 21.3 | 19.8 |
| Black alder | 36.9 | 23.9 | 21.2 | 30.2 | 27.1 | 26.7 |
| Chamber ID | Height, cm | Width, cm | Thickness, cm | Volume, m3 | Surface Area, m2 |
|---|---|---|---|---|---|
| 1 | 20.1 | 25.0 | 2.2 | 0.0011 | 0.0503 |
| 2 | 20.5 | 42.0 | 2.3 | 0.0020 | 0.0861 |
| 3 | 20.0 | 56.5 | 2.5 | 0.0028 | 0.1130 |
| 4 | 19.0 | 73.0 | 2.8 | 0.0039 | 0.1387 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petaja, G.; Ancāns, R.; Bārdule, A.; Spalva, G.; Meļņiks, R.N.; Purviņa, D.; Lazdiņš, A. Carbon Dioxide, Methane and Nitrous Oxide Fluxes from Tree Stems in Silver Birch and Black Alder Stands with Drained and Naturally Wet Peat Soils. Forests 2023, 14, 521. https://doi.org/10.3390/f14030521
Petaja G, Ancāns R, Bārdule A, Spalva G, Meļņiks RN, Purviņa D, Lazdiņš A. Carbon Dioxide, Methane and Nitrous Oxide Fluxes from Tree Stems in Silver Birch and Black Alder Stands with Drained and Naturally Wet Peat Soils. Forests. 2023; 14(3):521. https://doi.org/10.3390/f14030521
Chicago/Turabian StylePetaja, Guna, Ritvars Ancāns, Arta Bārdule, Gints Spalva, Raitis Normunds Meļņiks, Dana Purviņa, and Andis Lazdiņš. 2023. "Carbon Dioxide, Methane and Nitrous Oxide Fluxes from Tree Stems in Silver Birch and Black Alder Stands with Drained and Naturally Wet Peat Soils" Forests 14, no. 3: 521. https://doi.org/10.3390/f14030521