
Fermentation Conditions Affect the Synthesis of Volatile Compounds, Dextran, and Organic Acids by Weissella confusa A16 in Faba Bean Protein Concentrate
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of Experiment (DoE)
2.2. Preparation of Faba Bean Sourdough
2.3. Volatile Analysis
2.4. Viscosity Measurement and Determination of Dextran
2.5. Acidity
2.5.1. pH and Total Titratable Acidity Measurements (TTA)
2.5.2. Determination of Organic Acids
2.6. Model Validity
2.6.1. Model Fit and Internal Validation
2.6.2. Optimal Set Points and External Validation
2.7. Statistical Analysis
3. Results and Discussion
3.1. Volatile Profiles of Fermented Samples
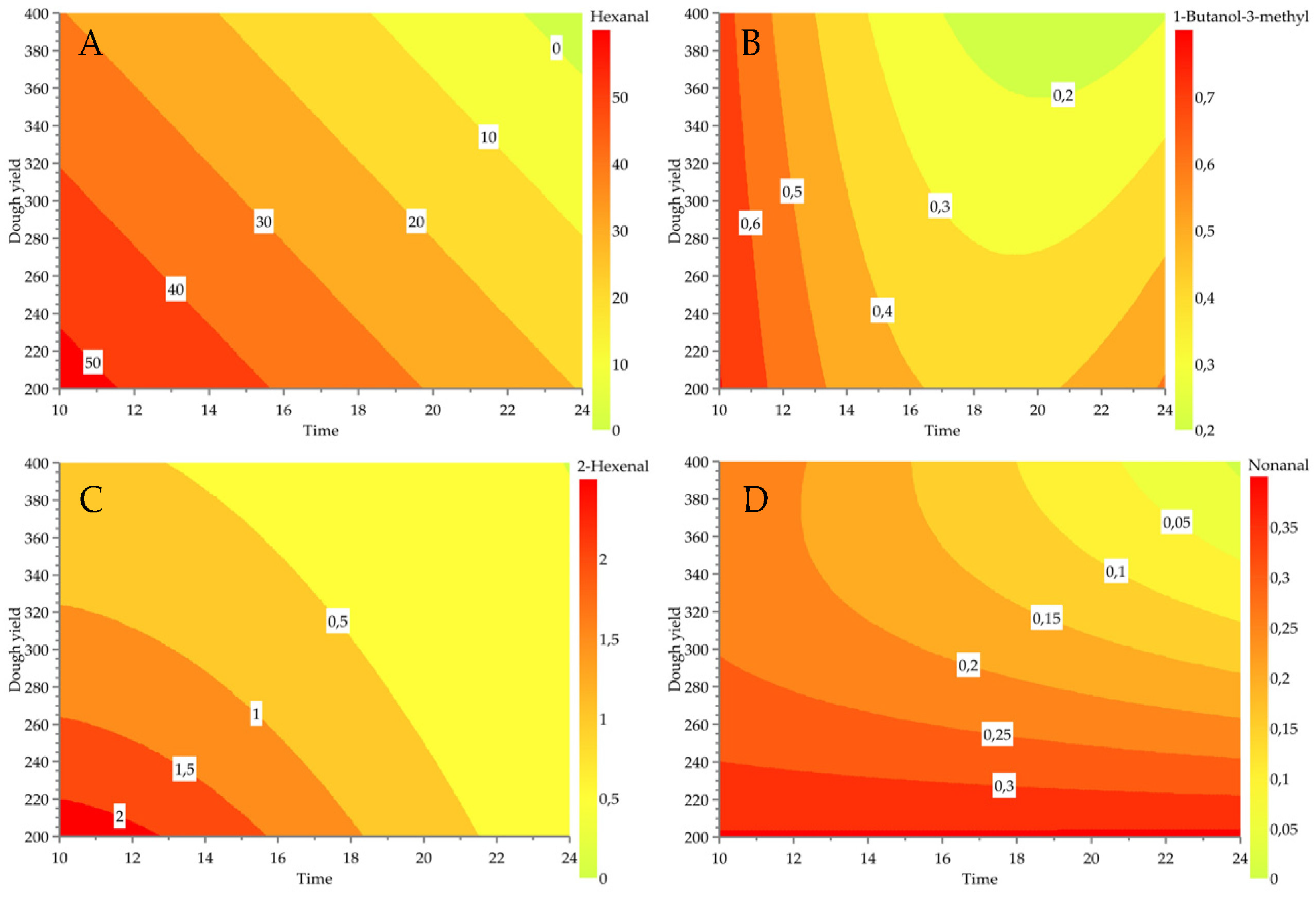
3.2. Effect of Fermentation Conditions on Modelable Volatile Compounds
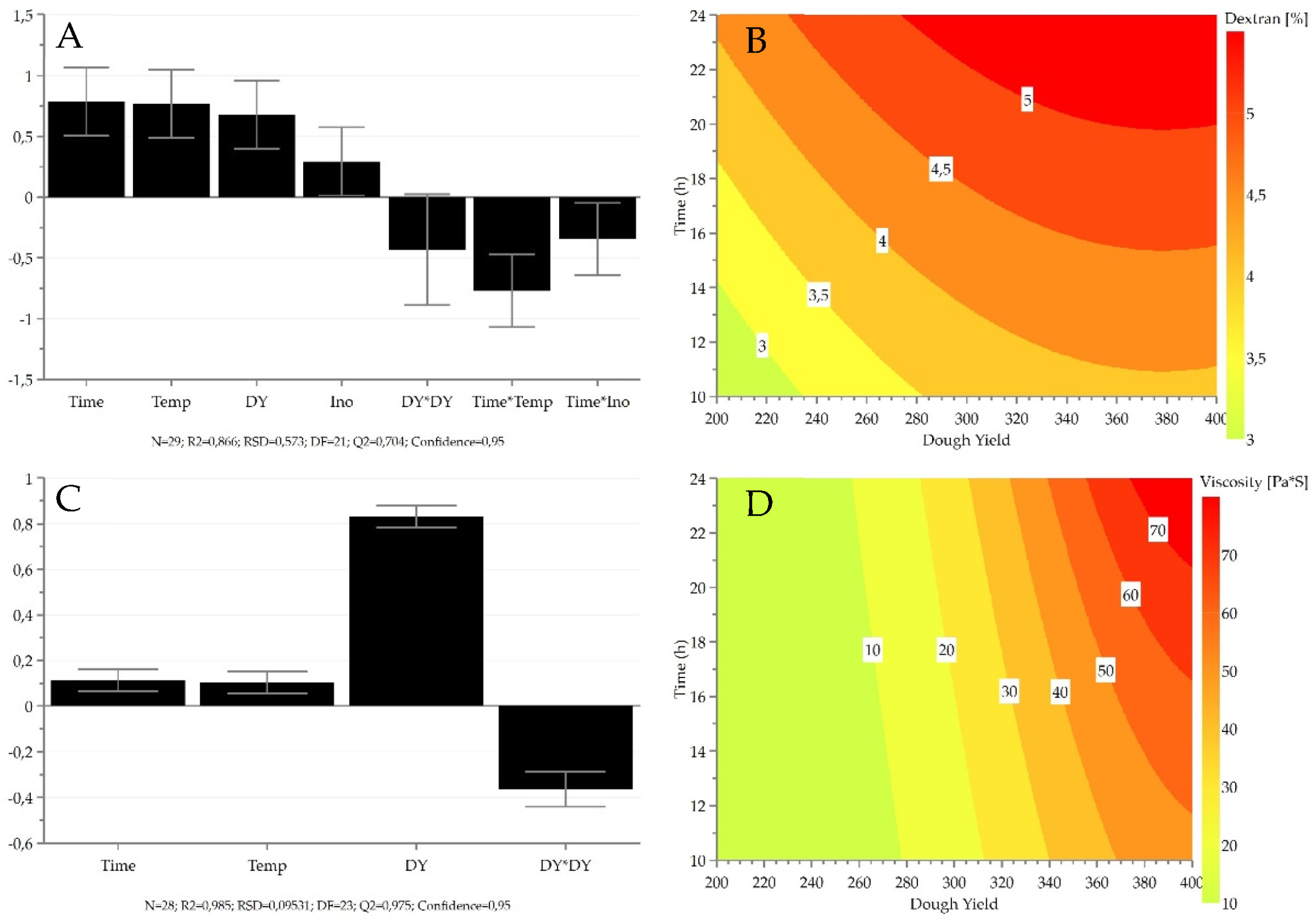
3.3. Effect of Fermentation Conditions on Viscosity and Dextran Production
3.4. Effect of Fermentation Conditions on Acidity
3.5. Optimized Fermentation Conditions for External Model Validation
3.6. Overall Effect of Fermentation Conditions on Metabolites
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graça, J.; Godinho, C.A.; Truninger, M. Reducing meat consumption and following plant-based diets: Current evidence and future directions to inform integrated transitions. Trends Food Sci. Tech. 2019, 91, 380–390. [Google Scholar] [CrossRef] [Green Version]
- Henning, B. Standing in livestock′s ‘‘long shadow’’: The ethics of eating meat on a small planet. Ethics Environ. 2011, 16, 63–93. [Google Scholar] [CrossRef]
- Curtain, F.; Grafenauer, S. Plant-based meat substitutes in the flexitarian age: An audit of products on supermarket shelves. Nutrients 2019, 11, 2603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharan, S.; Zanghelini, G.; Zotzel, J.; Bonerz, D.; Aschoff, J.; Saint-Eve, A.; Maillard, M.N. Fava bean (Vicia faba L.) for food applications: From seed to ingredient processing and its effect on functional properties, antinutritional factors, flavor, and color. Compr. Rev. Food Sci. Food Saf. 2021, 20, 401–428. [Google Scholar] [CrossRef]
- Multari, S.; Stewart, D.; Russell, W.R. Potential of fava bean as future protein supply to partially replace meat intake in the human diet. Compr. Rev. Food Sci. Food Saf. 2015, 14, 511–522. [Google Scholar] [CrossRef]
- Tuccillo, F.; Kantanen, K.; Wang, Y.; Ramos Diaz, J.M.; Pulkkinen, M.; Edelmann, M.; Knaapila, A.; Jouppila, K.; Piironen, V.; Lampi, A.M.; et al. The Flavor of Faba Bean Ingredients and Extrudates: Chemical and Sensory Properties. Food Res. Int. 2022, 162, 112036. [Google Scholar] [CrossRef]
- Schutyser, M.A.I.; Pelgrom, P.J.M.; Van der Goot, A.J.; Boom, R.M. Dry fractionation for sustainable production of functional legume protein concentrates. Trends Food Sci. Technol. 2015, 45, 327–335. [Google Scholar] [CrossRef]
- Kantanen, K.; Oksanen, A.; Edelmann, M.; Suhonen, H.; Sontag-Strohm, T.; Piironen, V.; Ramos Diaz, J.M.; Jouppila, K. Physical properties of extrudates with fibrous structures made of faba bean protein ingredients using high moisture extrusion. Foods 2022, 11, 1280. [Google Scholar] [CrossRef]
- do Carmo, C.S.; Knutsen, S.H.; Malizia, G.; Dessev, T.; Geny, A.; Zobel, H.; Myhrer, K.S.; Varela, P.; Sahlstrøm, S. Meat analogues from a faba bean concentrate can be generated by high moisture extrusion. Future Foods 2021, 3, 100014. [Google Scholar] [CrossRef]
- Wang, Y.; Tuccillo, F.; Lampi, A.M.; Knaapila, A.; Pulkkinen, M.; Kariluoto, S.; Coda, R.; Edelmann, M.; Jouppila, K.; Sandell, M.; et al. Flavor challenges in extruded plant-based meat alternatives: A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2898–2929. [Google Scholar] [CrossRef]
- Roland, W.S.; Pouvreau, L.; Curran, J.; van de Velde, F.; de Kok, P.M. Flavor aspects of pulse ingredients. Cereal. Chem. 2017, 94, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Frankel, E.N. Chemistry of Autoxidation: Mechanism, Products and Flavor Significance. In Flavor Chemistry of Fats and Oils; Min, D.B., Smouse, T.H., Eds.; AOCS: Champaign, France, 1985; pp. 1–37. [Google Scholar]
- Ritter, S.W.; Gastl, M.I.; Becker, T.M. The modification of volatile and nonvolatile compounds in lupines and faba beans by substrate modulation and lactic acid fermentation to facilitate their use for legume-based beverages—A review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4018–4055. [Google Scholar] [CrossRef] [PubMed]
- Bott, L.; Chambers, E., IV. Sensory characteristics of combinations of chemicals potentially associated with beany aroma in foods. J. Sens. Stud. 2006, 21, 308–321. [Google Scholar] [CrossRef]
- Li, X.; Li, J. The flavor of plant-based meat analogues. Cereal. Food World 2020, 65. [Google Scholar] [CrossRef]
- D’Souza, C.; Taghian, M.; Lamb, P.; Peretiatko, R. Green decisions: Demographics and consumer understanding of environmental labels. Int. J. Consum. Stud. 2007, 31, 371–376. [Google Scholar] [CrossRef]
- Borin, N.; Cerf, D.C.; Krishnan, R. Consumer effects of environmental impact in product labeling. J. Consum. Mark. 2011, 28, 76–86. [Google Scholar] [CrossRef]
- Rastogi, Y.R.; Thakur, R.; Thakur, P.; Mittal, A.; Chakrabarti, S.; Siwal, S.S.; Thakur, V.K.; Saini, A.K.; Saini, A.K. Food fermentation–Significance to public health and sustainability challenges of modern diet and food systems. Int. J. Food Microbiol. 2022, 371, 109666. [Google Scholar] [CrossRef]
- Verma, D.K.; Al-Sahlany, S.T.G.; Niamah, A.K.; Thakur, M.; Shah, N.; Singh, S.; Baranwalf, D.; Patel, A.R.; Utama, G.L.; Aguilar, C.N. Recent trends in microbial flavour compounds: A review on chemistry, synthesis mechanism and their application in food. Saudi J. Biol. Sci. 2021, 29, 1565–1576. [Google Scholar] [CrossRef]
- Arteaga, V.G.; Leffler, S.; Muranyi, I.; Eisner, P.; Schweiggert-Weisz, U. Sensory profile, functional properties and molecular weight distribution of fermented pea protein isolate. Curr. Res. Food Sci. 2021, 4, 1–10. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Tech. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- El Youssef, C.; Bonnarme, P.; Fraud, S.; Péron, A.C.; Helinck, S.; Landaud, S. Sensory improvement of a pea protein-based product using microbial co-cultures of lactic acid bacteria and yeasts. Foods 2020, 9, 349. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xie, C.; Pulkkinen, M.; Edelmann, M.; Chamlagain, B.; Coda, R.; Sandell, M.; Piironen, V.; Maina, N.H.; Katina, K. In situ production of vitamin B12 and dextran in soya flour and rice bran: A tool to improve flavour and texture of B12-fortified bread. LWT 2022, 161, 113407. [Google Scholar] [CrossRef]
- Wang, Y.; Sorvali, P.; Laitila, A.; Maina, N.H.; Coda, R.; Katina, K. Dextran produced in situ as a tool to improve the quality of wheat-faba bean composite bread. Food Hydrocoll. 2018, 84, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Trani, A.; Knaapila, A.; Hietala, S.; Coda, R.; Katina, K.; Maina, N.H. The effect of in situ produced dextran on flavour and texture perception of wholegrain sorghum bread. Food Hydrocoll. 2020, 106, 105913. [Google Scholar] [CrossRef]
- Bourdichon, F.; Patrone, V.; Fontana, A.; Milani, G.; Morelli, L. Safety demonstration of a microbial species for use in the food chain: Weissella confusa. Int. J. Food Microbiol. 2021, 339, 109028. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, C.; Vogel, R.F.; Toelstede, S. Fermentation performance of lactic acid bacteria in different lupin substrates—Influence and degradation ability of antinutritives and secondary plant metabolites. J. Appl. Microbiol. 2015, 119, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Arbab Sakandar, H.; Chen, Y.; Peng, C.; Chen, X.; Imran, M.; Zhang, H. Impact of fermentation on antinutritional factors and protein degradation of legume seeds: A review. Food Rev. Int. 2021, in press. [CrossRef]
- Rizzello, C.G.; Coda, R.; Wang, Y.; Verni, M.; Kajala, I.; Katina, K.; Laitila, A. Characterization of indigenous Pediococcus pentosaceus, Leuconostoc kimchii, Weissella cibaria and Weissella confusa for faba bean bioprocessing. Int. J. Food Microbiol. 2019, 302, 24–34. [Google Scholar] [CrossRef]
- Coda, R.; Melama, L.; Rizzello, C.G.; Curiel, J.A.; Sibakov, J.; Holopainen, U.; Pulkkinen, M.; Sozer, N. Effect of air classification and fermentation by Lactobacillus plantarum VTT E-133328 on faba bean (Vicia faba L.) flour nutritional properties. Int. J. Food Microbiol. 2015, 193, 34–42. [Google Scholar] [CrossRef]
- Kaleda, A.; Talvistu, K.; Tamm, M.; Viirma, M.; Rosend, J.; Tanilas, K.; Kriisa, M.; Part, N.; Tammik, M.L. Impact of fermentation and phytase treatment of pea-oat protein blend on physicochemical, sensory, and nutritional properties of extruded meat analogs. Foods 2020, 9, 1059. [Google Scholar] [CrossRef]
- Laaksonen, O.; Kahala, M.; Marsol-Vall, A.; Blasco, L.; Järvenpää, E.; Rosenvald, S.; Virtanen, M.; Tarvainen, M.; Yang, B. Impact of lactic acid fermentation on sensory and chemical quality of dairy analogues prepared from lupine (Lupinus angustifolius L.) seeds. Food Chem. 2021, 346, 128852. [Google Scholar] [CrossRef] [PubMed]
- Keast, R.S.; Breslin, P.A. An overview of binary taste–taste interactions. Food Qual. Prefer. 2003, 14, 111–124. [Google Scholar] [CrossRef] [Green Version]
- Katina, K.; Maina, N.H.; Juvonen, R.; Flander, L.; Johansson, L.; Virkki, L.; Tenkanen, M.; Laitila, A. In situ production and analysis of Weissella confusa dextran in wheat sourdough. Food Microbiol. 2009, 26, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Immonen, M.; Maina, N.H.; Wang, Y.; Coda, R.; Katina, K. Waste bread recycling as a baking ingredient by tailored lactic acid fermentation. Int. J. Food Microbiol. 2020, 327, 108652. [Google Scholar] [CrossRef]
- Cosson, A.; Blumenthal, D.; Descamps, N.; Souchon, I.; Saint-Eve, A. Using a mixture design and fraction-based formulation to better understand perceptions of plant-protein-based solutions. Food Res. Int. 2021, 141, 110151. [Google Scholar] [CrossRef] [PubMed]
- Lampi, A.M.; Yang, Z.; Mustonen, O.; Piironen, V. Potential of faba bean lipase and lipoxygenase to promote formation of volatile lipid oxidation products in food models. Food Chem. 2020, 311, 125982. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Q.; Pulkkinen, M.; Wang, Y.J.; Lampi, A.M.; Stoddard, F.L.; Salovaara, H.; Piironen, V.; Sontag-Strohm, T. Faba bean flavour and technological property improvement by thermal pre-treatments. LWT 2016, 68, 295–305. [Google Scholar] [CrossRef]
- Akkad, R.; Buchko, A.; Johnston, S.P.; Han, J.; House, J.D.; Curtis, J.M. Sprouting improves the flavour quality of faba bean flours. Food Chem. 2021, 364, 130355. [Google Scholar] [CrossRef]
- Akkad, R.; Kharraz, E.; Han, J.; House, J.D.; Curtis, J.M. The effect of short-term storage temperature on the key headspace volatile compounds observed in Canadian faba bean flour. Food Sci. Technol. Int. 2022, 28, 135–143. [Google Scholar] [CrossRef]
- Chiba, H.; Takahashi, N.; Sasaki, R. Enzymatic improvement of food flavor II. Removal of beany flavor from soybean products by aldehyde dehydrogenase. Agric. Biol. Chem. 1979, 43, 1883–1889. [Google Scholar]
- Sethi, S.; Tyagi, S.K.; Anurag, R.K. Plant-based milk alternatives an emerging segment of functional beverages: A review. J. Food Sci. Technol. 2016, 53, 3408–3423. [Google Scholar] [CrossRef]
- Monasterio, R.P. Transformations of food flavor due to industrially processing of elaboration. Adv. Food Nutr. Res. 2013, 2, 279–319. [Google Scholar]
- Akkad, R.; Kharraz, E.; Han, J.; House, J.D.; Curtis, J.M. Characterisation of the volatile flavour compounds in low and high tannin faba beans (Vicia faba var. minor) grown in Alberta, Canada. Food Res. Int. 2019, 120, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Vara-Ubol, S.; Chambers, E.; Chambers, D.H. Sensory characteristics of chemical compounds potentially associated with beany aroma in foods. J. Sens. Stud. 2004, 19, 15–26. [Google Scholar] [CrossRef]
- Thomas-Danguin, T.; Barba, C.; Salles, C.; Guichard, E. Perception of mixtures of odorants and tastants: Sensory and analytical points of view. In Flavour: From Food to Perception, 1st ed.; Guichard, E., Salles, C., Morzel, M., Le Bon, A.M., Eds.; Wiley Blackwell: Chichester, UK, 2017; pp. 319–340. [Google Scholar]
- Oomah, B.D.; Razafindrainibe, M.; Drover, J.C. Headspace volatile components of Canadian grown low-tannin faba bean (Vicia faba L.) genotypes. J. Sci. Food Agric. 2014, 94, 473–481. [Google Scholar] [CrossRef]
- Nardi, M.; Fiez-Vandal, C.; Tailliez, P.; Monnet, V. The EstA esterase is responsible for the main capacity of Lactococcus lactis to synthesize short chain fatty acid esters in vitro. J. Appl. Microbiol. 2002, 93, 994–1002. [Google Scholar] [CrossRef]
- Teixeira, C.G.; Fusieger, A.; Milião, G.L.; Martins, E.; Drider, D.; Nero, L.A.; de Carvalho, A.F. Weissella: An emerging bacterium with promising health benefits. Probiotics Antimicrob. 2021, 13, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Gänzle, M.G. Effect of temperature on production of oligosaccharides and dextran by Weissella cibaria 10 M. Int. J. Food Microbiol. 2018, 280, 27–34. [Google Scholar] [CrossRef]
- Amari, M.; Arango, L.F.G.; Gabriel, V.; Robert, H.; Morel, S.; Moulis, C.; Gabriel, B.; Remaud-Siméon, M.; Fontagné-Faucher, C. Characterization of a novel dextransucrase from Weissella confusa isolated from sourdough. Appl. Microbiol. Biotechnol. 2013, 97, 5413–5422. [Google Scholar] [CrossRef]
- Kajala, I.; Shi, Q.; Nyyssölä, A.; Maina, N.H.; Hou, Y.; Katina, K.; Tenkanen, M.; Juvonen, R. Cloning and characterization of a Weissella confusa dextransucrase and its application in high fibre baking. PLoS ONE 2015, 10, e0116418. [Google Scholar] [CrossRef] [Green Version]
- Besrour-Aouam, N.; Fhoula, I.; Hernández-Alcántara, A.M.; Mohedano, M.L.; Najjari, A.; Prieto, A.; López, P.; Ouzari, H.I. The role of dextran production in the metabolic context of Leuconostoc and Weissella Tunisian strains. Carbohydr. Polym. 2021, 253, 117254. [Google Scholar] [CrossRef] [PubMed]
- Peyer, L.C.; Zannini, E.; Arendt, E.K. Lactic acid bacteria as sensory biomodulators for fermented cereal-based beverages. Trends Food Sci. Tech. 2016, 54, 17–25. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Galle, S.; Schwab, C.; Arendt, E.; Gänzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef]
- Wang, Y.; Tuccillo, F.; Niklander, K.; Livi, G.; Siitonen, A.; Pöri, P.; Edelmann, M.; Jouppila, K.; Lampi, A.M.; Sandell, M.; et al. Masking off-flavors of faba bean protein concentrate and extrudate: The role of in-situ and in-vitro produced dextran. Food Hydrocoll. 2022, in press.
- Meinlschmidt, P.; Ueberham, E.; Lehmann, J.; Schweiggert-Weisz, U.; Eisner, P. Immunoreactivity, sensory and physicochemical properties of fermented soy protein isolate. Food Chem. 2016, 205, 229–238. [Google Scholar] [CrossRef]
- Kaczmarska, K.T.; Chandra-Hioe, M.V.; Frank, D.; Arcot, J. Aroma characteristics of lupin and soybean after germination and effect of fermentation on lupin aroma. LWT 2018, 87, 225–233. [Google Scholar] [CrossRef]
- Paradiso, V.M.; Summo, C.; Pasqualone, A.; Caponio, F. Evaluation of different natural antioxidants as affecting volatile lipid oxidation products related to off-flavours in corn flakes. Food Chem. 2009, 113, 543–549. [Google Scholar] [CrossRef]
- Flavornet.org. Available online: http://www.flavornet.org/flavornet.html (accessed on 30 October 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment Number | Fermentation Parameters | Sourdough Recipes | ||||||
|---|---|---|---|---|---|---|---|---|
| Time (h) | Temperature (°C) | Dough Yield | Inoculum Ratio (CFU/g) | FPC (g) | Water (mL) | Sucrose (g) | W. confusa A16 ** (µL) | |
| N1 | 10 | 20 | 200 | 105 | 90 | 100 | 10 | 30 |
| N2 | 24 | 20 | 200 | 105 | 90 | 100 | 10 | 30 |
| N3 | 10 | 30 | 200 | 105 | 90 | 100 | 10 | 30 |
| N4 | 24 | 30 | 200 | 105 | 90 | 100 | 10 | 30 |
| N5 | 10 | 20 | 400 | 105 | 90 | 300 | 10 | 50 |
| N6 | 24 | 20 | 400 | 105 | 90 | 300 | 10 | 50 |
| N7 | 10 | 30 | 400 | 105 | 90 | 300 | 10 | 50 |
| N8 | 24 | 30 | 400 | 105 | 90 | 300 | 10 | 50 |
| N9 | 10 | 20 | 200 | 107 | 90 | 100 | 10 | 2000 |
| N10 | 24 | 20 | 200 | 107 | 90 | 100 | 10 | 2000 |
| N11 | 10 | 30 | 200 | 107 | 90 | 100 | 10 | 2000 |
| N12 | 24 | 30 | 200 | 107 | 90 | 100 | 10 | 2000 |
| N13 | 10 | 20 | 400 | 107 | 90 | 300 | 10 | 4000 |
| N14 | 24 | 20 | 400 | 107 | 90 | 300 | 10 | 4000 |
| N15 | 10 | 30 | 400 | 107 | 90 | 300 | 10 | 4000 |
| N16 | 24 | 30 | 400 | 107 | 90 | 300 | 10 | 4000 |
| N17 | 10 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N18 | 24 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N19 | 17 | 20 | 300 | 106 | 90 | 200 | 10 | 300 |
| N20 | 17 | 30 | 300 | 106 | 90 | 200 | 10 | 300 |
| N21 | 17 | 25 | 200 | 106 | 90 | 100 | 10 | 200 |
| N22 | 17 | 25 | 400 | 106 | 90 | 300 | 10 | 400 |
| N23 | 17 | 25 | 300 | 105 | 90 | 200 | 10 | 40 |
| N24 | 17 | 25 | 300 | 107 | 90 | 200 | 10 | 3000 |
| N25 * | 17 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N26 * | 17 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N27 * | 17 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N28 * | 17 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| N29 * | 17 | 25 | 300 | 106 | 90 | 200 | 10 | 300 |
| Volatile Compounds | LRI | Relative Peak Area (%) | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N1 | N2 | N3 | N4 | N5 | N6 | N7 | N8 | N9 | N10 | N11 | N12 | N13 | N14 | N15 | N16 | N17 | N18 | N19 | N20 | N21 | N22 | N23 | N24 | N25-29 | ||
| Organic Acids | ||||||||||||||||||||||||||
| Acetic acid | 752 | - | 12.3 | 11.8 | 10.9 | - | 5.0 | 4.5 | 7.4 | 0.9 | 8.6 | 11.9 | 11.5 | 4.0 | 7.4 | 5.7 | 8.6 | 10.3 | 10.1 | 7.6 | 11.9 | 8.5 | 5.2 | 6.9 | 9.2 | 7.6 |
| Isovaleric acid | 959 | - | 6.8 | 5.2 | 5.4 | - | 2.6 | 3.1 | 3.0 | 1.5 | 4.6 | 4.3 | 4.7 | 2.8 | 3.9 | 3.5 | 3.3 | 4.9 | 3.7 | 3.5 | 3.8 | 4.1 | 2.5 | 3.0 | 2.9 | 2.9 |
| Alcohols | ||||||||||||||||||||||||||
| Ethanol | 3.56 ** | 0.7 | 6.7 | 2.0 | 16.7 | 0.3 | 11.6 | 7.3 | 16.6 | 1.0 | 10.5 | 4.5 | 19.2 | 1.3 | 11.4 | 8.7 | 17.6 | 4.6 | 13.3 | 6.7 | 15.6 | 7.4 | 10.5 | 12.6 | 8.3 | 12.6 |
| 3-Methyl-1-butanol | 813 | 0.7 | 0.7 | 0.7 | 0.3 | 0.7 | 0.4 | 0.6 | 0.1 | 1.0 | 0.6 | 0.4 | 0.5 | 0.8 | 0.3 | 0.5 | 0.1 | 0.8 | 0.1 | 0.4 | 0.1 | 0.5 | 0.2 | 0.3 | 0.2 | 0.3 |
| 1-Pentanol | 845 | 1.7 | 1.5 | 1.1 | 0.7 | 1.6 | 0.7 | 1.3 | 0.2 | 1.4 | 1.0 | 1.3 | 0.6 | 1.8 | 0.5 | 1.1 | 0.1 | 0.7 | 0.4 | 0.8 | 0.4 | 1.1 | 0.5 | 0.6 | 0.5 | 0.4 |
| 1-Penten-3-ol | 756 | 0.3 | 0.3 | 0.3 | 0.2 | 0.1 | 0.1 | 0.1 | 0.01 | 0.3 | 0.2 | 0.2 | 0.1 | 0.1 | 0.03 | 0.1 | 0.01 | 0.1 | 0.03 | 0.1 | 0.04 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 |
| 3-Hexen-1-ol | 939 | - | 0.5 | - | 0.6 | - | 0.3 | 0.6 | 0.1 | - | 0.5 | - | 0.2 | 0.9 | 0.3 | 0.4 | 0.1 | 0.7 | 0.1 | 0.7 | 0.2 | 0.6 | 0.2 | 0.4 | 0.5 | 0.4 |
| 1-Hexanol | 947 | 11.7 | 45.2 | 19.8 | 15.8 | 23.4 | 46.4 | 52.0 | 11.7 | 14.7 | 25.1 | 31.5 | 28.0 | 37.4 | 38.8 | 49.6 | 11.9 | 32.3 | 29.0 | 37.1 | 24.7 | 26.1 | 44.1 | 38.7 | 40.1 | 22.5 |
| 1-Octanol | 816 | 0.4 | 0.3 | 0.4 | 0.2 | 0.6 | 0.3 | 0.3 | 0.1 | 0.6 | 0.3 | 0.4 | 0.4 | 0.5 | 0.2 | 0.3 | 0.1 | 0.4 | 0.1 | 0.3 | 0.1 | 0.4 | 0.2 | 0.3 | 0.3 | 0.3 |
| Aldehydes | ||||||||||||||||||||||||||
| 3-Methylbutanal * | 717 | 0.2 | 0.1 | 0.4 | 0.1 | 0.4 | 0.1 | 0.05 | 0.02 | 0.3 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.03 | 0.02 | 0.1 | 0.04 | 0.03 | 0.1 | 0.1 | 0.01 | 0.05 | - | 0.03 |
| Pentanal | 759 | 0.6 | 0.1 | 0.5 | 0.2 | 0.9 | 0.1 | 0.2 | 0.1 | 0.5 | 0.3 | 0.3 | 0.1 | 0.4 | 0.1 | 0.1 | 0.05 | 0.2 | 0.1 | 0.1 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 |
| Hexanal | 865 | 77.3 | 16.7 | 48.3 | 18.3 | 59.5 | 1.9 | 13.9 | 0.1 | 70.7 | 29.8 | 34.4 | 11.9 | 41.5 | 0.7 | 7.3 | - | 37.1 | 2.2 | 30.0 | 10.2 | 35.2 | 6.9 | 15.5 | 20.1 | 27.1 |
| 2-Hexenal | 936 | 2.6 | 0.6 | 1.8 | - | 1.7 | - | 0.5 | - | 2.8 | 0.6 | 1.7 | 0.2 | 1.0 | - | 0.2 | - | 1.1 | 0.03 | 0.7 | 0.2 | 1.0 | 0.1 | 0.6 | 0.6 | 0.9 |
| Heptanal | 971 | 0.3 | 0.1 | 0.2 | 0.1 | 0.3 | 0.2 | 0.04 | - | 0.3 | 0.2 | 0.1 | - | - | - | 0.1 | - | 0.1 | - | - | - | 0.1 | - | - | - | - |
| 2-Heptenal | 1045 | 0.5 | 0.3 | 0.4 | 0.6 | 0.1 | - | 0.2 | 0.02 | 0.3 | 0.5 | 0.5 | 0.3 | 0.3 | 0.1 | 0.2 | 0.04 | 0.1 | 0.1 | 0.3 | 0.3 | 0.7 | 0.1 | 0.2 | 0.3 | 0.4 |
| 2-Octenal | 1152 | - | 0.3 | - | 0.6 | - | - | - | - | - | 0.6 | 0.4 | 0.4 | - | 0.02 | - | - | 0.2 | 0.1 | 0.3 | 0.3 | 0.7 | - | - | - | - |
| Nonanal | 1182 | 0.4 | 0.4 | 0.3 | 0.4 | 0.4 | - | 0.2 | - | 0.4 | 0.4 | 0.3 | 0.2 | 0.2 | - | 0.1 | - | 0.2 | 0.1 | 0.3 | 0.2 | 0.4 | 0.1 | 0.1 | 0.2 | 0.2 |
| Alkanes | ||||||||||||||||||||||||||
| Hexane | 620 | 0.1 | 0.1 | 0.1 | - | 0.7 | 0.1 | 0.2 | - | 0.05 | 0.03 | 0.1 | - | 0.3 | 0.1 | 0.1 | - | 0.2 | - | - | - | - | - | - | - | - |
| Octane | 823 | 0.3 | 0.3 | 0.3 | 0.1 | 2.6 | 0.4 | 0.7 | 0.1 | 0.2 | 0.1 | 0.2 | 0.1 | 0.9 | 0.3 | 0.6 | 0.03 | 0.4 | 0.03 | 0.2 | 0.1 | 0.1 | 0.2 | 0.2 | 0.2 | 0.1 |
| Esters | ||||||||||||||||||||||||||
| Ethyl acetate | 664 | - | 1.6 | 0.6 | 19.3 | - | 21.9 | 9.1 | 43.4 | - | 7.8 | 1.6 | 13.9 | 1.4 | 26.4 | 16.1 | 38.1 | 1.5 | 23.3 | 5.6 | 20.7 | 6.4 | 21.9 | 15.0 | 11.3 | 18.6 |
| Isoamyl acetate | 931 | - | 0.1 | 0.2 | 0.3 | - | 0.4 | 0.1 | 0.2 | - | 0.3 | 0.1 | 0.2 | 0.1 | 0.4 | 0.2 | 0.2 | 0.04 | 0.3 | 0.1 | 0.3 | 0.2 | 0.3 | 0.2 | 0.2 | 0.3 |
| Acetic acid methyl ester | 4.49 ** | 0.04 | 0.1 | 0.3 | 0.3 | 0.05 | 0.5 | 0.4 | 0.3 | 0.1 | 0.3 | 0.3 | 0.3 | 0.7 | 0.5 | 0.5 | 0.3 | 0.3 | 0.3 | 0.4 | 0.3 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| Acetic acid hexyl ester | 1070 | - | 0.6 | 0.3 | 1.1 | - | 2.2 | 0.8 | 1.2 | - | 1.0 | 0.5 | 0.8 | 0.5 | 2.5 | 1.3 | 1.5 | 0.3 | 3.4 | 1.3 | 1.9 | 1.0 | 2.4 | 1.5 | 1.5 | 1.5 |
| Hexanoic acid ethyl ester | 1054 | - | - | - | 2.2 | - | 1.0 | - | 6.3 | - | 0.4 | - | 1.0 | - | 1.6 | - | 5.9 | - | 5.1 | 0.9 | 2.5 | - | 1.2 | 0.6 | 0.3 | 0.9 |
| Heptanoic acid ethyl ester | 1156 | - | - | - | 0.1 | - | - | - | 0.2 | - | - | - | - | - | - | - | 0.3 | - | 0.2 | - | 0.1 | - | - | - | - | - |
| Octanoic acid ethyl ester | 1257 | - | - | - | 0.7 | - | 0.2 | - | 1.0 | - | 0.2 | - | 0.1 | - | 0.4 | - | 1.5 | - | 1.2 | 0.1 | 0.5 | 0.1 | 0.2 | - | - | - |
| Isoamyl valerate * | 1163 | 0.4 | 0.6 | 0.6 | 0.3 | 0.4 | 0.3 | 0.3 | 0.1 | 0.5 | 0.8 | 0.6 | 0.3 | 0.3 | 0.3 | 0.2 | 0.1 | 0.5 | 0.2 | 0.5 | 0.2 | 0.6 | 0.2 | 0.4 | 0.5 | 0.4 |
| Methyl isovalerate | 827 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | - | 0.02 | 0.1 | 0.2 | 0.1 | 0.1 | 0.1 | 0.1 | - | - | 0.05 | 0.04 | 0.04 | - | 0.2 | 0.04 | 0.05 | 0.03 | 0.1 |
| Ethyl isovalerate | 903 | - | 0.8 | 0.1 | 0.8 | - | 0.4 | 0.1 | 0.2 | - | 1.8 | 0.4 | 0.6 | - | 0.4 | 0.2 | 0.2 | 0.1 | 0.3 | 0.3 | 0.3 | 1.2 | 0.3 | 0.3 | 0.2 | 0.4 |
| Propyl isovalerate | 1202 | - | 0.1 | 0.4 | 0.1 | 0.1 | 0.03 | 0.01 | - | 0.03 | 0.1 | - | - | 0.02 | 0.04 | 0.04 | - | 0.04 | - | 0.1 | - | 0.05 | 0.01 | - | - | - |
| Ethyl lactate | 889 | - | 0.03 | - | 0.8 | - | 0.4 | 0.1 | 5.8 | - | 0.1 | - | 0.3 | - | 0.8 | 0.2 | 6.1 | - | 1.8 | 0.1 | 1.1 | 0.1 | 0.3 | 0.1 | 0.1 | 0.3 |
| Ketones | ||||||||||||||||||||||||||
| Acetoin | 805 | - | 0.04 | 0.3 | 0.5 | - | 0.03 | - | 0.2 | 0.02 | 0.04 | 0.1 | 2.5 | - | 0.2 | 0.04 | 0.1 | 0.02 | 0.6 | - | 0.4 | 0.02 | - | - | 0.1 | 0.03 |
| 2-Hexanone * | 859 | 0.1 | 0.1 | 0.1 | - | 0.1 | - | - | - | 0.1 | - | - | - | 0.1 | - | 0.1 | - | - | - | - | - | - | - | - | - | - |
| 3-Hexanone * | 851 | 0.1 | 0.04 | 0.1 | 0.05 | 0.1 | - | - | 0.1 | 0.1 | - | 0.1 | - | 0.04 | 0.03 | 0.04 | 0.1 | - | - | - | - | - | - | - | - | - |
| Aromatic Compounds | ||||||||||||||||||||||||||
| 2-Methylfuran * | 655 | 0.5 | 0.8 | 1.0 | 0.5 | 1.1 | 0.3 | 0.8 | 2.4 | 0.5 | 0.5 | 1.0 | 0.3 | 0.6 | 0.5 | 0.5 | 3.4 | 1.0 | 3.1 | 0.4 | 1.8 | 0.3 | 0.2 | 0.2 | 0.3 | 0.2 |
| 2-Pentylfuran | 1039 | 0.4 | 0.3 | 0.3 | 0.6 | 0.9 | 0.6 | 0.6 | 0.2 | 0.3 | 0.5 | 0.5 | 0.3 | 0.5 | 0.5 | 0.6 | 0.4 | 0.3 | 0.5 | 0.4 | 0.5 | 0.3 | 0.5 | 0.4 | 0.4 | 0.4 |
| Terpenes | ||||||||||||||||||||||||||
| Alpha-pinene | 975 | - | 0.1 | 0.2 | 0.05 | 0.5 | 0.2 | 0.1 | - | - | 0.1 | 0.1 | - | - | 0.1 | 0.2 | - | 0.2 | - | - | - | 0.1 | - | - | - | - |
| Delta-3-carene * | 1056 | 0.5 | 1.4 | 1.9 | 1.0 | 2.6 | 1.3 | 1.4 | - | 1.3 | 1.5 | 1.9 | 0.9 | 1.3 | 1.1 | 1.1 | - | 1.2 | - | 1.0 | 1.0 | 1.2 | 1.1 | 1.1 | 1.2 | 0.6 |
| D-Limonene | 1077 | 0.4 | 0.2 | 0.2 | 0.2 | 0.7 | 0.2 | 0.2 | - | 0.2 | 0.3 | 0.3 | - | 0.2 | 0.2 | 0.2 | - | 0.2 | 0.1 | 0.5 | 0.1 | 0.2 | 0.1 | 0.1 | 0.2 | - |
| Modellable Volatile Compounds | Model Statistics | Model Terms Coefficients | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DF | RSD | R2 | Q2 | Repr | Tim | Temp | DY | Ino | Tim × Tim | DY × DY | Ino × Ino | Tim × Temp | Tim × DY | Temp × DY | DY × Ino | |
| Ethanol | 24 | 1.85 | 0.90 | 0.85 | 0.98 | 1.0 | 0.6 | 0.2 | −0.5 | |||||||
| 3-Methyl-1-butanol | 23 | 0.11 | 0.82 | 0.72 | 0.76 | −0.7 | −0.5 | −0.4 | 0.9 | −0.3 | ||||||
| 1-Pentanol | 23 | 0.16 | 0.91 | 0.85 | 0.98 | −0.7 | −0.5 | −0.3 | 1.1 | −0.3 | ||||||
| 1-Penten-3-ol | 19 | 0.02 | 0.97 | 0.92 | 0.99 | −0.4 | −0.2 | −0.9 | −0.2 | 0.3 | 0.8 | −0.2 | −0.1 | 0.1 | ||
| Pentanal | 24 | 0.13 | 0.86 | 0.77 | 0.98 | −0.9 | −0.4 | −0.5 | 0.7 | |||||||
| Hexanal | 24 | 6.19 | 0.92 | 0.87 | 0.98 | −0.8 | −0.5 | −0.6 | 0.4 | |||||||
| 2-Hexenal | 24 | 0.18 | 0.88 | 0.82 | 0.96 | −0.9 | −0.4 | −0.6 | −0.4 | |||||||
| Nonanal | 23 | 0.05 | 0.87 | 0.77 | 0.96 | −0.5 | −0.3 | −0.9 | 0.4 | −0.5 | ||||||
| Hexane | 21 | 0.09 | 0.89 | 0.74 | 1.00 | −0.7 | −0.3 | 0.3 | −0.2 | 1.2 | −0.3 | −0.2 | ||||
| Octane | 20 | 0.15 | 0.90 | 0.77 | 0.99 | −0.8 | −0.5 | 0.4 | −0.2 | 0.8 | −0.2 | −0.3 | −0.3 | |||
| Ethyl acetate | 22 | 3.50 | 0.93 | 0.87 | 0.99 | 0.8 | 0.5 | 0.6 | 0.2 | 0.3 | 0.2 | |||||
| Hexanoic acid ethyl ester | 22 | 0.22 | 0.90 | 0.84 | 0.99 | 0.9 | 0.3 | 0.3 | −0.6 | 0.3 | 0.3 | |||||
| Ethyl isovalerate | 21 | 0.11 | 0.91 | 0.80 | 0.94 | 0.9 | 0.2 | −0.4 | −0.9 | 0.5 | −0.5 | −0.3 | ||||
| Ethyl lactate | 22 | 0.15 | 0.93 | 0.88 | 0.98 | 0.8 | 0.5 | 0.5 | 0.3 | 0.3 | 0.2 | |||||
| Fermentation Conditions | OSP 1 | OSP 2 | OSP 3 |
|---|---|---|---|
| Time (h) | 11.4 | 15.1 | 10.3 |
| Temperature (°C) | 23 | 23 | 25 |
| Dough Yield | 380 | 400 | 350.5 |
| Inoculum ration (log CFU/g) | 6.8 | 6.7 | 6 |
| Predicted and Measured Responses | |||
| Dextran (%) | 4.0; 4.7 | 4.3; 4.9 | 3.9; 4.5 |
| Viscosity (Pa·s) | 41.5; 47.7 | 51.7; 71.2 | 34.3; 20.4 * |
| Acetic acid (mg/g) | 1.8; 2.5 | 2.1; 2.7 | 1.7; 2.1 |
| Lactic acid (mg/g) | 2.9; 5.7 | 7.2; 8.8 | 3.1; 4.2 |
| pH | 6.0; 5.8 | 5.8; 5.5 * | 6.0; 5.9 |
| TTA | 9.1; 10.2 | 9.9; 12.6 * | 10.0; 10.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tuccillo, F.; Wang, Y.; Edelmann, M.; Lampi, A.-M.; Coda, R.; Katina, K. Fermentation Conditions Affect the Synthesis of Volatile Compounds, Dextran, and Organic Acids by Weissella confusa A16 in Faba Bean Protein Concentrate. Foods 2022, 11, 3579. https://doi.org/10.3390/foods11223579
Tuccillo F, Wang Y, Edelmann M, Lampi A-M, Coda R, Katina K. Fermentation Conditions Affect the Synthesis of Volatile Compounds, Dextran, and Organic Acids by Weissella confusa A16 in Faba Bean Protein Concentrate. Foods. 2022; 11(22):3579. https://doi.org/10.3390/foods11223579
Chicago/Turabian StyleTuccillo, Fabio, Yaqin Wang, Minnamari Edelmann, Anna-Maija Lampi, Rossana Coda, and Kati Katina. 2022. "Fermentation Conditions Affect the Synthesis of Volatile Compounds, Dextran, and Organic Acids by Weissella confusa A16 in Faba Bean Protein Concentrate" Foods 11, no. 22: 3579. https://doi.org/10.3390/foods11223579