
Evaluation of the Availability and Antioxidant Capacity of Maillard Compounds Present in Bread Crust: Studies in Caco-2 Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Extraction of Bread Crust and Its Soluble Fractions with Different Molecular Weights
2.3. Characterization of Bread Crust and the LMW and HMW Fractions
2.4. Bioaccessibility of Amadori Compounds and HMF from Bread Crust
2.5. Cells
2.6. Amadori Compounds and HMF Absorption Experiments
2.7. Reactive Oxygen Species Generation
2.8. Statistical Treatment
3. Results and Discussion
3.1. Sample Characterization and Bioaccessibility of Amadori Compounds and HMF from Bread Crust
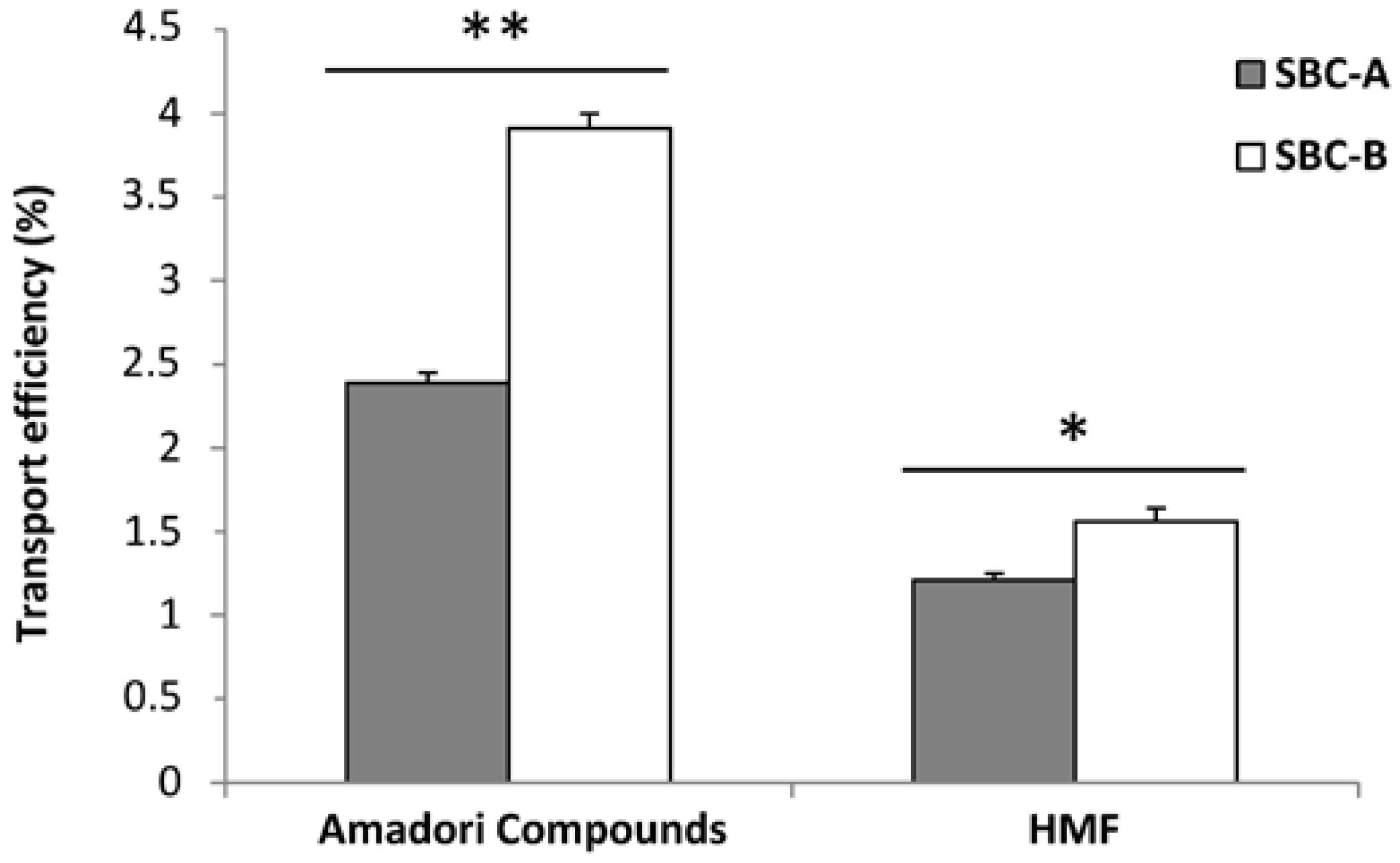
3.2. Amadori Compounds and HMF Transport in Cell Culture
3.3. ROS Generation
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kroh, L.W. Caramelisation in food and beverages. Food Chem. 1994, 51, 373–379. [Google Scholar] [CrossRef]
- Wagner, K.H.; Derkits, S.; Herr, M.; Schih, W.; Elmadfa, I. Antioxidative potential of melanoidins isolated from a roasted glucose-glycine model. Food Chem. 2002, 78, 375–382. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Morales, F.J. Effect of in vitro enzymatic digestion on antioxidant activity of coffee melanoidins and fractions. J. Agric. Food Chem. 2007, 55, 10016–10021. [Google Scholar] [CrossRef] [PubMed]
- Michalska, A.; Amigo-Benavent, M.; Zieliński, H.; del Castillo, M.D. Effect of baking on the formation of MRPs contributing to the overall antioxidant activity of rye bread. J. Cereal Sci. 2008, 48, 123–132. [Google Scholar] [CrossRef]
- Zielínski, H.; Michalska, A.; Ceglińska, A.; Lamparski, G. Antioxidant properties and sensory quality of traditional rye bread as affected by the incorporation of flour with different extraction rates in the formulation. Eur. Food Res. Technol. 2008, 226, 671–680. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Morales, F.J. Unraveling the contribution of melanoidins to the antioxidant activity of coffee brews. J. Agric. Food Chem. 2005, 53, 1403–1407. [Google Scholar] [CrossRef] [PubMed]
- Rufián-Henares, J.A.; Delgado-Andrade, C. Effect of digestive process on Maillard reaction indexes and antioxidant properties of breakfast cereals. Food Res. Int. 2009, 42, 394–400. [Google Scholar] [CrossRef]
- Andreasen, M.F.; Christensen, L.P.; Meyer, A.S.; Hansen, A. Ferulic acid dehydrodimers in rye (Secale cereale L.). J. Cereal Sci. 2000, 31, 303–307. [Google Scholar] [CrossRef]
- Bertais, S.; Luque, M.L.P.; Preziosi, P.; Fieux, B.; Torra de Flot, M.; Galan, P.; Hercherg, S. Contribution of ready-to-eat cereals to nutrition intakes in French adults and relations with corpulence. Ann. Nutr. Metab. 2000, 44, 249–255. [Google Scholar] [CrossRef]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, M.D.; Ames, J.M.; Gordon, M.H. Effect of roasting on the antioxidant activity of coffee brews. J. Agric. Food Chem. 2002, 50, 3698–3703. [Google Scholar] [CrossRef] [PubMed]
- Van Barneveld, J.R.; Batterham, E.S.; Skingle, D.C.; Norton, B.W. The effect of heat on amino acids for growing pigs. 4. Nitrogen balance and urine, serum and plasma composition of growing pigs fed on raw or heated-treated field peas (Pisum sativum). Br. J. Nutr. 1995, 73, 259–273. [Google Scholar]
- Delgado-Andrade, C. Carboxymethyl-lysine: Thirty years of investigation in the field of AGE formation. Food Funct. 2016, 7, 46–57. [Google Scholar] [CrossRef] [PubMed]
- Leuner, B.; Ruhs, S.; Broemme, H.J.; Bierhaus, A.; Sel, S.; Silber, R.E.; Somoza, V.; Simma, A.; Nass, N. RAGE-dependent activation of gene expression of superoxide dismutase and vanins by AGE-rich extracts in mice cardiac tissue and murine cardiac fibroblasts. Food Funct. 2012, 3, 1091–1098. [Google Scholar] [CrossRef] [PubMed]
- Nursten, H.E. Recent developments in studies of the Maillard reaction. Food Chem. 1980, 6, 263–277. [Google Scholar] [CrossRef]
- Ramírez-Jiménez, A.; Guerra-Hernández, E.; García-Villanova, B. Browning indicators in bread. J. Agric. Food Chem. 2000, 48, 4176–4181. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Rufián-Henares, J.A.; Morales, F.J. Lysine availability is diminished in commercial fibre-enriched breakfast cereals. Food Chem. 2007, 100, 725–731. [Google Scholar] [CrossRef]
- Surh, Y.J.; Tannenbaum, S.R. Activation of the Maillard reaction product 5-hydroxymethylfurfural to strong mutagens via allylic sulfonation and chlorination. Chem. Res. Toxicol. 1994, 7, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Pastoriza, S.; Álvarez, J.; Végvári, A.; Montilla-Gómez, J.; Cruz-López, O.; Delgado-Andrade, C.; Rufián-Henares, J.A. Relationship between HMF intake and SMF formation in vivo: An animal and human study. Mol. Nutr. Food Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.J.; Liem, A.; Miller, J.A.; Tannenbaum, S.R. 5-Sulfooximethylfurfural as a possible ultimate mutagenic and carcinogenic metabolite of the Maillard reaction product, 5-hydroxymethylfurfural. Carcinogenesis 1994, 15, 2375–2377. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Tessier, F.J.; Niquet-Leridon, C.; Seiquer, I.; Navarro, M.P. Study of the urinary and faecal excretion of Nε-carboxymethyllysine in young human volunteers. Amino Acids 2012, 43, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Mesías, M.; Seiquer, I.; Delgado-Andrade, C.; Galdó, G.; Navarro, M.P. Intake of Maillard reaction products reduces iron bioavailability in male adolescents. Mol. Nutr. Food Res. 2009, 53, 1551–1560. [Google Scholar]
- Llana-Ruiz-Cabello, M.; Gutiérrez-Praena, D.; Puerto, M.; Pichardo, S.; Moreno, F.J.; Baños, A.; Nuñez, C.; Guillamón, E.; Cameán, A.M. Acute toxicological studies of the main organosulfur compound derived from Allium sp. intended to be used in active food packaging. Food Chem. Toxicol. 2015, 82, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Seiquer, I.; Navarro, M.P.; Morales, F.J. Estimation of hydroxymethylfurfural availability in breakfast cereals. Studies in Caco-2 cells. Food Chem Toxicol. 2008, 46, 1600–1607. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Roca, B.; Delgado-Andrade, C.; Navarro, M.P.; Seiquer, I. Effects of Maillard reaction products from glucose-lysine model systems on oxidative stress markers and against oxidative induction by hydrogen peroxide in Caco-2 cells. J. Food Nutr. Res. 2011, 50, 237–248. [Google Scholar]
- Roncero-Ramos, I.; Delgado-Andrade, C.; Tessier, F.J.; Niquet-Léridon, C.; Strauch, C.; Monnier, V.M.; Navarro, M.P. Metabolic transit of Nε-carboxymethyl-lysine after consumption of AGEs from bread crust. Food Funct. 2013, 4, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Rufián-Henares, J.A.; Morales, F.J. Procedure to analyse Maillard reaction-associated fluorescence in cereal-based products. Czech J. Food Sci. 2008, 26, 339–346. [Google Scholar]
- Pérez-Jiménez, J.; Saura-Calixto, F. Literature data may underestimate the actual antioxidant capacity of cereals. J. Agric. Food Chem. 2005, 53, 5036–5040. [Google Scholar] [CrossRef] [PubMed]
- Rufián-Henares, J.A.; Morales, F.J. Functional properties of melanoidins: In vitro antioxidant, antimicrobial and antihypertensive activities. Food Res. Int. 2007, 40, 995–1002. [Google Scholar] [CrossRef]
- Resmini, P.; Pellegrino, L. Analysis of food heat damage by direct HPLC of furosine. Int. Chromatogr. Lab. 1991, 6, 7–11. [Google Scholar]
- Delgado-Andrade, C.; Rufián-Henares, J.A.; Morales, F.J. Fast method to determine furosine in breakfast cereals by capillary zone electrophoresis. Eur. Food Res. Technol. 2005, 221, 707–711. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; García-Villanova, B.; Guerra-Hernández, E. Determination of furfural compounds in enteral formula. J. Liq. Chromatogr. Relat. Technol. 2001, 24, 3049–3062. [Google Scholar] [CrossRef]
- Rufián-Henares, J.A.; Delgado-Andrade, C.; Morales, F.J. Application of a fast high-performance liquid chromatography method for simultaneous determination of furanic compounds and glucosylisomaltol in breakfast cereals. J. AOAC Int. 2006, 89, 161–165. [Google Scholar] [PubMed]
- Navarro, P.; Aspe, T.; Seiquer, I. Zinc transport in caco-2 cells and zinc transport in rats: Influence of the heat treatment of casein–glucose–fructose mixture. J. Agric. Food Chem. 2000, 48, 3585–3596. [Google Scholar] [CrossRef]
- Ruiz-Roca, B.; Navarro, M.P.; Seiquer, I. Antioxidant properties and metal chelating activity of glucose-lysine heated mixtures. Relationships with mineral absorption across Caco-2 cell monolayers. J. Agric. Food Chem. 2008, 56, 9056–9063. [Google Scholar] [CrossRef] [PubMed]
- Goya, L.; Mateos, R.; Bravo, L. Effect of the olive oil phenol hydroxytyrosol on human hepatoma HepG2 cells. Protection against oxidative stress induced by tert-butylhydroperoxide. Eur. J. Nutr. 2007, 46, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Fogliano, V.; Morales, F.J. Estimation of dietary intake of melanoidins from coffee and bread crust. Food Funct. 2011, 2, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helou, C.; Jacolot, P.; Niquet-Léridon, C.; Gadonna-Widehem, P.; Tessier, F.J. Maillard reaction products in bread: A novel semi-quantitative method for evaluating melanoidins in bread. Food Chem. 2016, 190, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Andrade, C.; Conde-Aguilera, J.A.; Haro, A.; de la Cueva, S.P.; Rufián-Henares, J.A. A combined procedure to evaluate the global antioxidant response of bread. J. Cereal Sci. 2010, 52, 239–246. [Google Scholar] [CrossRef]
- Hidalgo, A.; Brandolini, A. Evaluation of heat damage, sugars, amylases and colour in breads from einkorn, durum and bread wheat flours. J. Cereal Sci. 2011, 54, 90–97. [Google Scholar] [CrossRef]
- Seidowski, A.; Lunow, D.; Henle, T. Amadori products—Substrates and inhibitors for intestinal brush border glycosidases. Czech J. Food Sci. 2009, 27, S146–S148. [Google Scholar]
- Stadler, N.C.; Somoza, V.; Schwab, W. Absorption of 3(2H)-Furanones by human intestinal epithelial Caco-2 cells. J. Agric. Food Chem. 2009, 57, 3949–3954. [Google Scholar] [CrossRef] [PubMed]
- Ganong, W.F. Digestion and absorption. In Review of Medical Physiology, 14th ed.; Ganong, W.F., Ed.; Appleton and Lange: San Francisco, CA, USA, 1993; pp. 505–516. [Google Scholar]
- Borges, T.; Cabrera-Vique, C.; Seiquer, I. Antioxidant properties of chemical extracts and bioaccessible fractions obtained from six Spanish monovarietal extra virgin olive oils: Assays in Caco-2 cells. Food Funct. 2015, 6, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Lindenmeier, M.; Somoza, V.; Hofmann, T. Structural and functional characterization of pronyl-lysine, a novel protein modification in bread crust melanoidins showing in vitro antioxidative and phase I/II enzyme modulating activity. J. Agric. Food Chem. 2002, 50, 6997–7006. [Google Scholar] [CrossRef] [PubMed]
- Pötzsch, S.; Dalgalarrondo, M.; Bakan, B.; Marion, D.; Somoza, V.; Stangl, G.; Silber, R.E.; Simm, A.; Navarrete Santos, A. The antioxidative effect of bread crust in a mouse macrophage reporter cell line. Free Radic. Biol. Med. 2014, 75, S19. [Google Scholar] [CrossRef] [PubMed]
- Pastoriza, S.; Roncero-Ramos, I.; Rufián-Henares, J.A.; Delgado-Andrade, C. Antioxidant balance after long-term consumption of standard diets including bread crust glycated compounds by adult rats. Food Res. Int. 2014, 64, 106–113. [Google Scholar] [CrossRef]
- Pastoriza, S.; Rufián-Henares, J.A.; Delgado-Andrade, C. Effects of long-term consumption of standard diets including glucose-lysine model glycated compounds on the antioxidant status of adult rats. Food Chem. 2015, 183, 283–290. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Samples | N (%) | C (%) | DPPH (mmol·Trolox/kg) | FRAP (mmol·Trolox/kg Powder Sample) | Amadori Compounds (as Furosine) (mg/100 g Protein in Powder Sample) | Amadori Compounds (as Furosine ) in Bioaccessible Fraction (mg/kg Powder BC) | Bioaccessible Amadori Compounds (as Furosine) (%) | HMF (mg/kg Powder Sample) | HMF in Bioaccessible Fraction (mg/kg Powder BC) | Bioaccessible HMF (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| BC | 2.32 ± 0.03 b | 44.60 ± 0.01 c | 10.88 ± 0.34 c | 98.5 ± 3.4 a | 377.9 ± 12.3 a | 31.7 ± 3.9 | 6.41 ± 0.52 | 27 ± 0.3 b | 1.1 ± 0.2 | 4.03 ± 0.46 |
| LMW | 4.36 ± 0.04 c | 35.95 ± 0.07 a | 9.01 ± 0.42 b | 206.5 ± 6.8 c | 906.0 ± 41.7 b | - | - | 40 ± 0.6 c | - | - |
| HMW | 2.20 ± 0.01 a | 38.95 ± 0.07 b | 0.92 ± 0.01 a | 167.7 ± 4.3 b | 1376.1 ± 38.2 c | - | - | 20 ± 0.3 a | - | - |
| Solutions | Amadori Compounds Transport (as Furosine 1) | HMF Transport | ||
|---|---|---|---|---|
| ng/well | % | ng/well | % | |
| SBC-A | 303 ± 8 a | 40.1 ± 1.1 a | 8.6 ± 0.3 a | 35.5 ± 1.2 d |
| SBC-B | 211 ± 4 b | 56.0 ± 1.2 b | 4.8 ± 0.2 b | 35.4 ± 1.6 d |
| LMW-B | 118 ± 1 c | 99.2 ± 1.2 c | 3.2 ± 0.2 c | 96.0 ± 6.7 a |
| HMW-B | 109 ± 3 c | 55.6 ± 1.5 b | 1.1 ± 0.1 d | 55.6 ± 4.3 c |
| HMW-C | 73 ± 1 d | 55.6 ± 1.1 b | 0.8 ± 0.0 d | 69.4 ± 6.9 b |
| Solutions | Apical Chamber | Basolateral Chamber | |
|---|---|---|---|
| Initial Abs (AU) | Final Abs (AU) | Recovery (%) | |
| 280 nm | |||
| SBC-A | 73.06 ± 0.10 a | 1.140 ± 0.029 a | 1.56 ± 0.04 d |
| SBC-B | 3.768 ± 0.02 b | 0.384 ± 0.010 b | 10.19 ± 0.26 c |
| LMW-B | 2.620 ± 0.01 c | 0.391 ± 0.005 b | 14.94 ± 0.21 a |
| HMW-B | 2.384 ± 0.01 d | 0.306 ± 0.007 c | 12.82 ± 0.28 b |
| HMW-C | 1.787 ± 0.01 e | 0.219 ± 0.013 d | 12.24 ± 0.74 b |
| 420 nm | |||
| SBC-A | 2.346 ± 0.01 a | 0.070 ± 0.005 a | 3.00 ± 0.20 d |
| SBC-B | 1.126 ± 0.01 b | 0.046 ± 0.006 b | 4.11 ± 0.57 d |
| LMW-B | 0.283 ± 0.01 c | 0.031 ± 0.001 c | 10.95 ± 0.20 c |
| HMW-B | 0.154 ± 0.01 d | 0.032 ± 0.004 c | 21.00 ± 2.50 b |
| HMW-C | 0.109 ± 0.01 e | 0.032 ± 0.001 c | 29.66 ± 1.22 a |
| Samples | ROS (Fluorescent Units × 104) | |
|---|---|---|
| Basal Effect | Protective Effect | |
| Control 1 | 18.13 ± 2.79 a | 36.25 ± 2.02 a |
| SBC-A | 2.86 ± 0.44 c | 10.61 ± 0.59 c |
| SBC-B | 7.82 ± 1.20 b | 18.35 ± 1.02 b |
| LMW-B | 18.04 ± 2.78 a | 35.57 ± 1.98 a |
| HMW-B | 19.91 ± 3.07 a | 30.99 ± 1.73 a |
| HMW-C | 14.21 ± 2.19 a | 29.60 ± 1.65 a |
© 2017 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Cueva, S.P.; Seiquer, I.; Mesías, M.; Rufián-Henares, J.Á.; Delgado-Andrade, C. Evaluation of the Availability and Antioxidant Capacity of Maillard Compounds Present in Bread Crust: Studies in Caco-2 Cells. Foods 2017, 6, 5. https://doi.org/10.3390/foods6010005
De la Cueva SP, Seiquer I, Mesías M, Rufián-Henares JÁ, Delgado-Andrade C. Evaluation of the Availability and Antioxidant Capacity of Maillard Compounds Present in Bread Crust: Studies in Caco-2 Cells. Foods. 2017; 6(1):5. https://doi.org/10.3390/foods6010005
Chicago/Turabian StyleDe la Cueva, Silvia Pastoriza, Isabel Seiquer, Marta Mesías, José Ángel Rufián-Henares, and Cristina Delgado-Andrade. 2017. "Evaluation of the Availability and Antioxidant Capacity of Maillard Compounds Present in Bread Crust: Studies in Caco-2 Cells" Foods 6, no. 1: 5. https://doi.org/10.3390/foods6010005