Bioavailability of Melatonin from Lentil Sprouts and Its Role in the Plasmatic Antioxidant Status in Rats

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Lentil Sprout Extracts
2.3. Extraction and Analysis of Phenolic Compounds in Raw Lentils and Lentil Sprouts
2.3.1. Extraction of Free and Bound Phenolic Compounds
2.3.2. HPLC–DAD–ESI/MSn qualitative and quantitative analyses of phenolic compounds
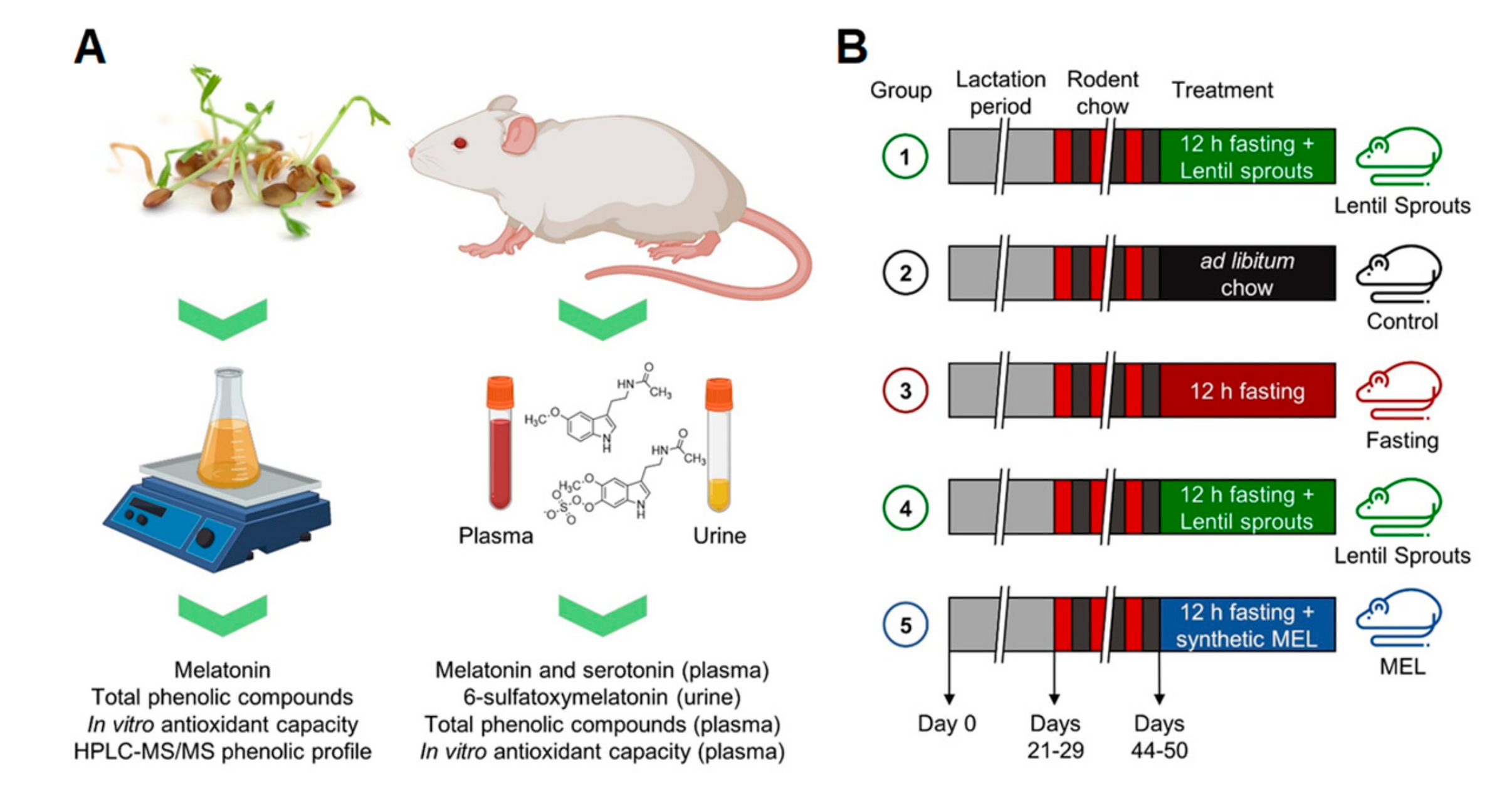
2.4. Animals Care and Diet
2.5. Pharmacokinetic Study
2.6. Bioavailability and Bioactivity Study
2.7. Melatonin Determination
2.8. 6-Sulfatoxymelatonin (aMT6s) Determination
2.9. Serotonin Quantification
2.10. Total Phenolic Compounds (TPC)
2.11. ORAC (Oxygen Radical Absorbance Capacity)
2.12. FRAP (Ferric Reducing Antioxidant Power)
2.13. Statistical Analysis
3. Results and Discussion
3.1. Lentil Sprouts are a Source of Melatonin and Antioxidant Phytochemicals
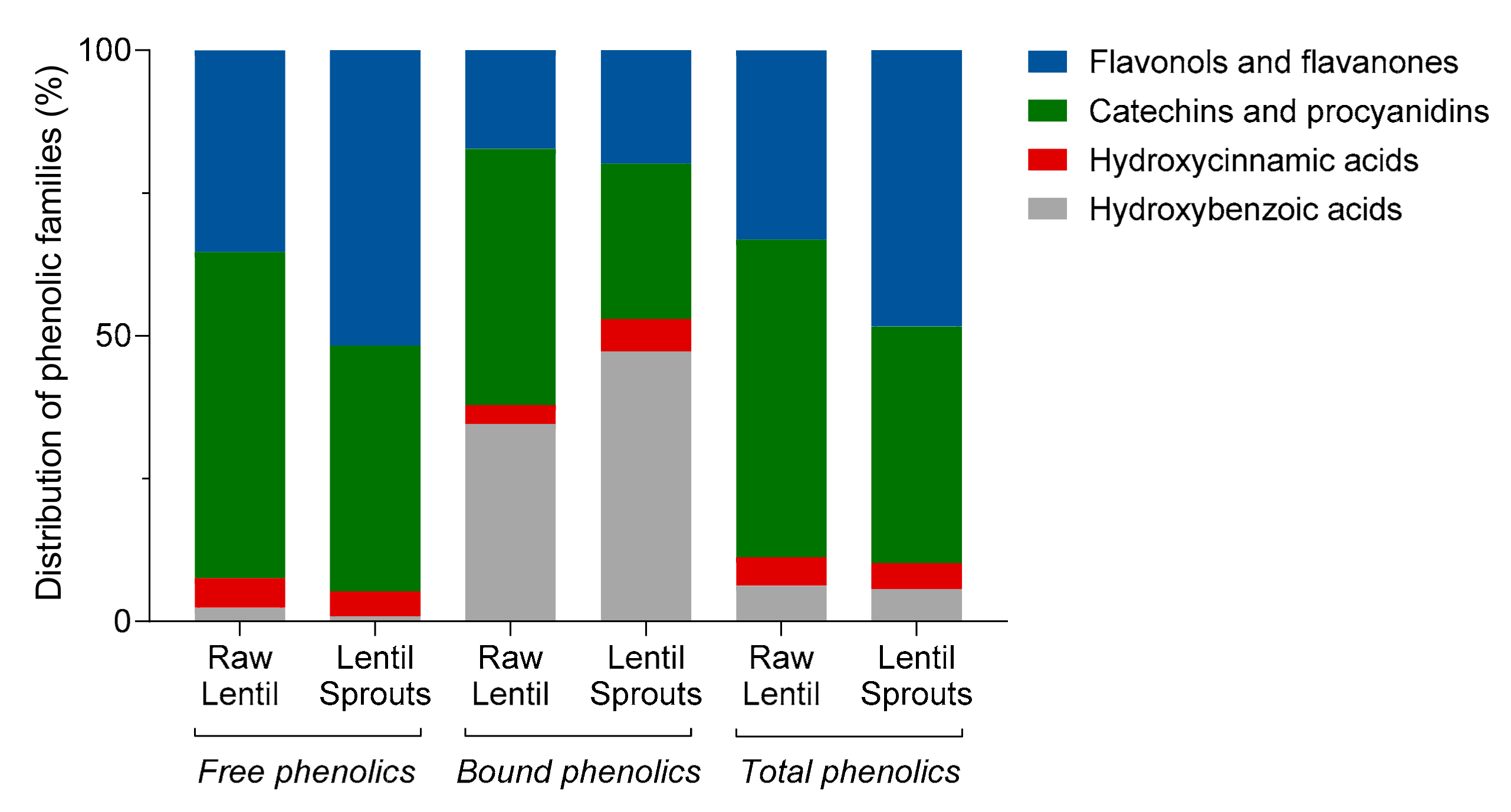
Germination Decreased the Concentration of Phenolic Acids and Flavan-3-ols, Preserving Flavonols
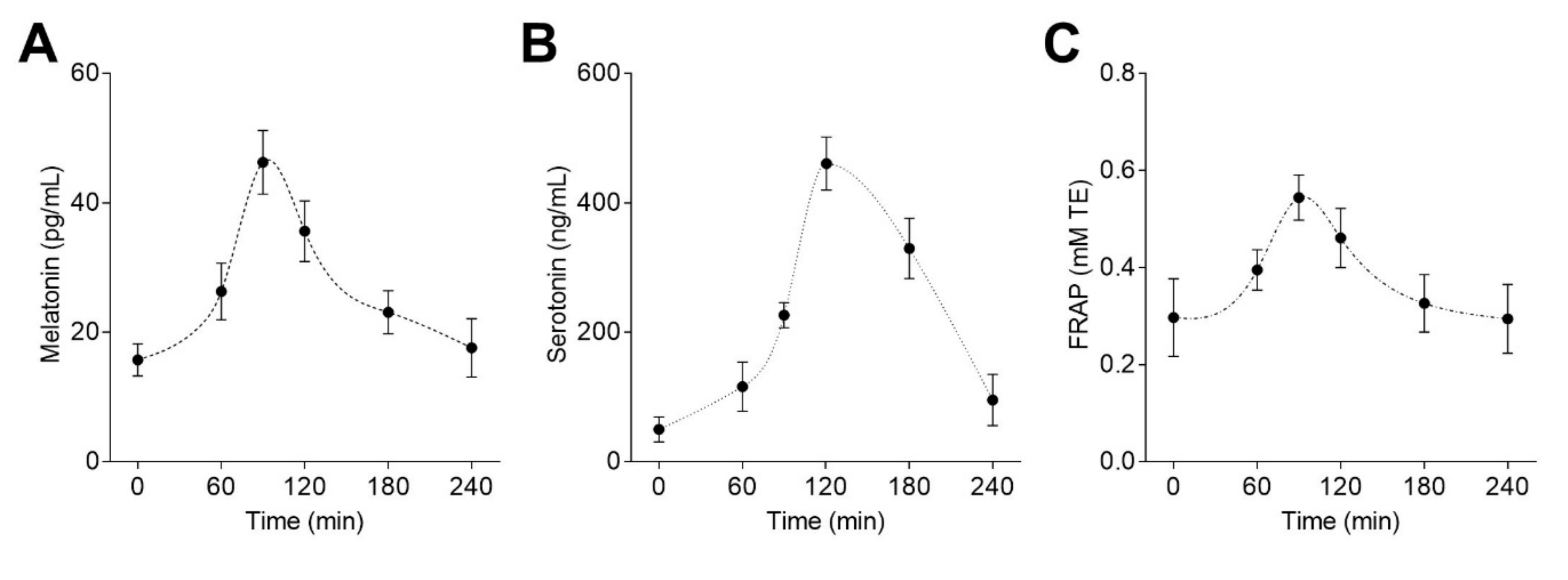
3.2. Bioavailability of Melatonin from Lentil Sprouts and its Correlation with Plasma Antioxidant Capacity
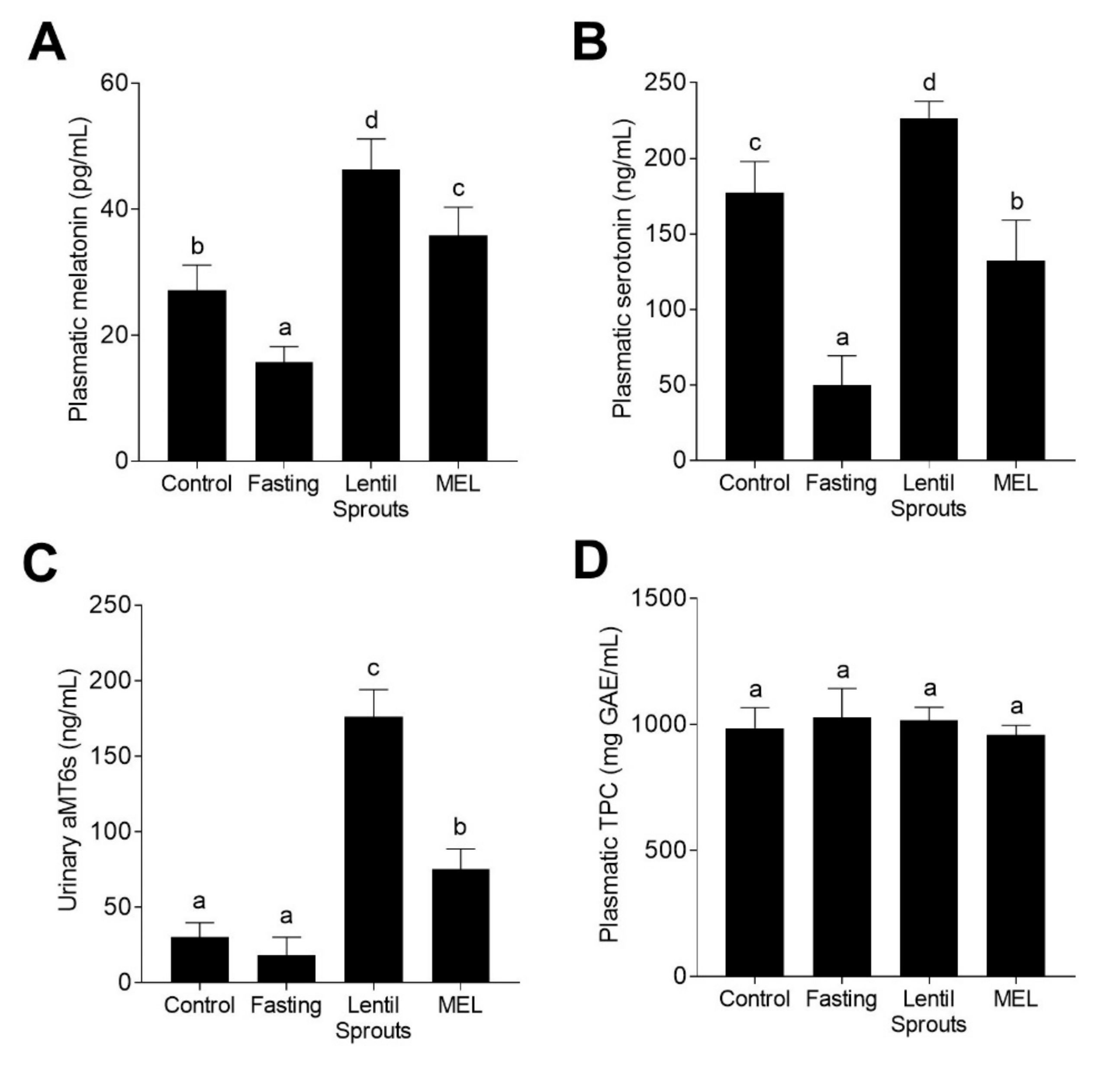
3.3. Lentil Sprouts Intake Greater Elevate Melatonin Biomarkers than the Synthetic Hormone
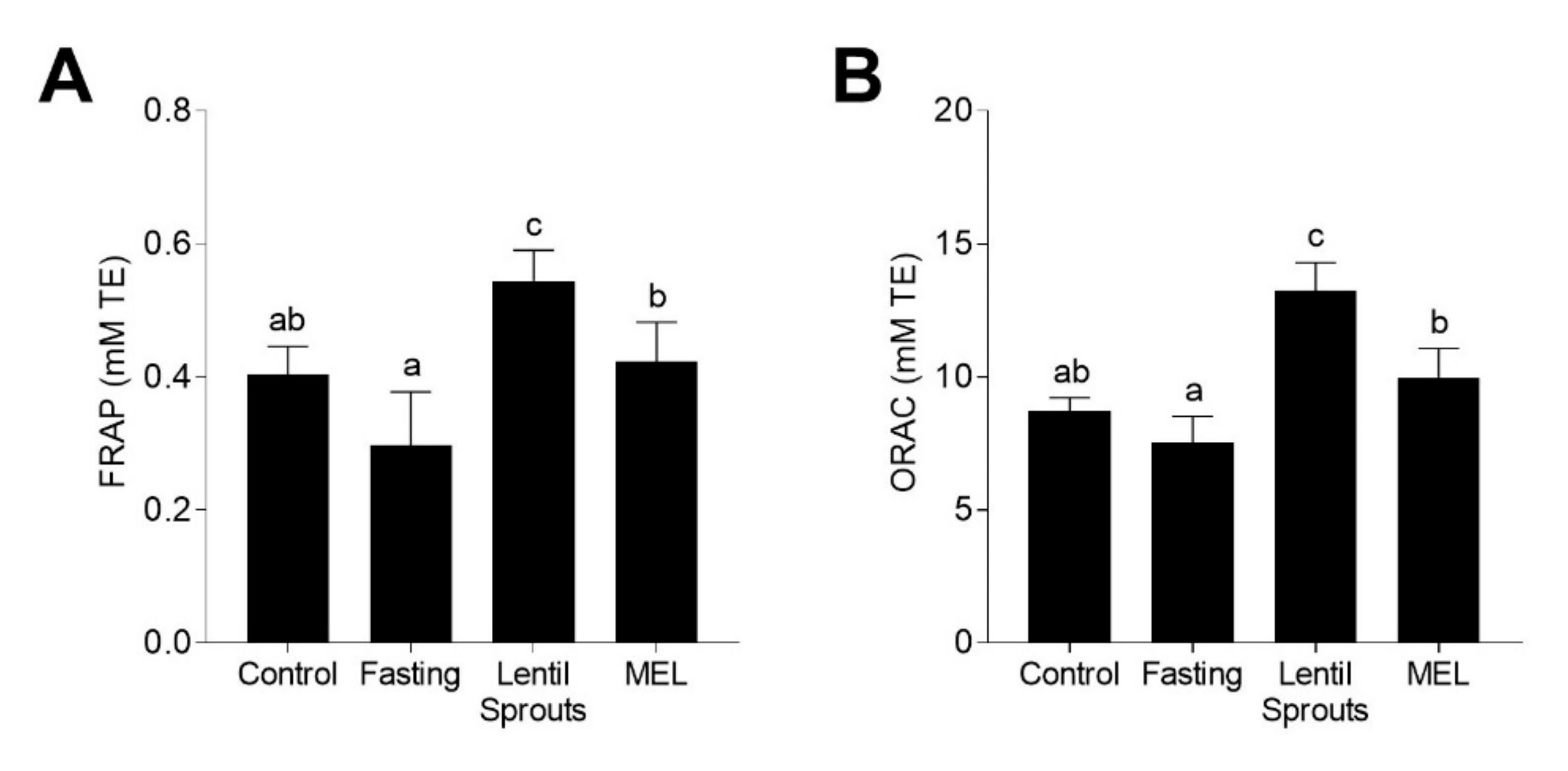
3.4. Administration of Lentil Sprouts Enhances Plasmatic Antioxidant Status
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnao, M.B.; Hernández-Ruiz, J. The Potential of Phytomelatonin as a Nutraceutical. Molecules 2018, 23, 238. [Google Scholar] [CrossRef] [Green Version]
- Loren, P.; Sánchez, R.; Arias, M.-E.; Felmer, R.; Risopatrón, J.; Cheuquemán, C. Melatonin Scavenger Properties against Oxidative and Nitrosative Stress: Impact on Gamete Handling and In Vitro Embryo Production in Humans and Other Mammals. Int. J. Mol. Sci. 2017, 18, 1119. [Google Scholar] [CrossRef] [Green Version]
- Tan, D.-X.; Manchester, L.C.; Qin, L.; Reiter, R.J. Melatonin: A Mitochondrial Targeting Molecule Involving Mitochondrial Protection and Dynamics. Int. J. Mol. Sci. 2016, 17, 2124. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin and the theories of aging: A critical appraisal of melatonin’s role in antiaging mechanisms. J. Pineal Res. 2013, 55, 325–356. [Google Scholar] [CrossRef] [PubMed]
- Zephy, D.; Ahmad, J. Type 2 diabetes mellitus: Role of melatonin and oxidative stress. Diabetes Metab. Syndr. Clin. Res. Rev. 2015, 9, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Cipolla-Neto, J.; Amaral, F.G.; Afeche, S.C.; Tan, D.X.; Reiter, R.J. Melatonin, energy metabolism, and obesity: A review. J. Pineal Res. 2014, 56, 371–381. [Google Scholar] [CrossRef] [Green Version]
- Favero, G.; Franceschetti, L.; Buffoli, B.; Moghadasian, M.H.; Reiter, R.J.; Rodella, L.F.; Rezzani, R. Melatonin: Protection against age-related cardiac pathology. Ageing Res. Rev. 2017, 35, 336–349. [Google Scholar] [CrossRef]
- Peuhkuri, K.; Sihvola, N.; Korpela, R. Dietary factors and fluctuating levels of melatonin. Food Nutr. Res. 2012, 56, 17252. [Google Scholar] [CrossRef]
- Iriti, M.; Varoni, E.M. Melatonin in Mediterranean diet, a new perspective. J. Sci. Food Agric. 2015, 95, 2355–2359. [Google Scholar] [CrossRef]
- Bonomini, F.; Borsani, E.; Favero, G.; Rodella, L.; Rezzani, R. Dietary Melatonin Supplementation Could Be a Promising Preventing/Therapeutic Approach for a Variety of Liver Diseases. Nutr. 2018, 10, 1135. [Google Scholar] [CrossRef] [Green Version]
- Cardinali, D.P. Melatonin as a chronobiotic/cytoprotector: Its role in healthy aging. Biol. Rhythm Res. 2019, 50, 28–45. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the substantiation of a health claim related to melatonin and reduction of sleep onset latency (ID 1698, 1780, 4080) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2241. [Google Scholar] [CrossRef] [Green Version]
- Jiki, Z.; Lecour, S.; Nduhirabandi, F. Cardiovascular Benefits of Dietary Melatonin: A Myth or a Reality? Front. Physiol. 2018, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- Herrera, T.; Aguilera, Y.; Rebollo-Hernanz, M.; Bravo, E.; Benítez, V.; Martínez-Sáez, N.; Arribas, S.M.; del Castillo, M.D.; Martín-Cabrejas, M.A. Teas and herbal infusions as sources of melatonin and other bioactive non-nutrient components. LWT 2018, 89, 65–73. [Google Scholar] [CrossRef]
- Meng, J.-F.; Shi, T.-C.; Song, S.; Zhang, Z.-W.; Fang, Y.-L. Melatonin in grapes and grape-related foodstuffs: A review. Food Chem. 2017, 231, 185–191. [Google Scholar] [CrossRef]
- Tan, D.-X.; Zanghi, B.M.; Manchester, L.C.; Reiter, R.J. Melatonin identified in meats and other food stuffs: Potentially nutritional impact. J. Pineal Res. 2014, 57, 213–218. [Google Scholar] [CrossRef]
- Aguilera, Y.; Rebollo-Hernanz, M.; Martín-Cabrejas, M.A. Melatonin. In Legumes: Nutritional Quality, Processing and Potential Health Benefits; Martín-Cabrejas, M.A., Ed.; CRC Press: Boca Raton, FL, USA, 2019; pp. 129–151. ISBN 9781482204056. [Google Scholar]
- Martín-Cabrejas, M.A.; Aguilera, Y.; Benítez, V.; Reiter, R.J. Chapter 6—Melatonin Synthesis in Fermented Foods. In Fermented Foods in Health and Disease Prevention; Elsevier: New York, USA, 2017; pp. 105–129. ISBN 9780128023099. [Google Scholar]
- Vaz Patto, M.C.; Amarowicz, R.; Aryee, A.N.A.; Boye, J.I.; Chung, H.-J.; Martín-Cabrejas, M.A.; Domoney, C. Achievements and Challenges in Improving the Nutritional Quality of Food Legumes. CRC. Crit. Rev. Plant Sci. 2015, 34, 105–143. [Google Scholar] [CrossRef]
- Díaz, M.F.; Martínez, M.; Savón, L.L.; Torres, V.; Coto, G. CHAPTER 8. Obtaining, Chemically Characterizing and Nutritionally Evaluating Seasonal Legume Sprouts as a Feed Alternative. In Legumes; Royal Society of Chemistry: London, UK, 2019; pp. 177–195. [Google Scholar] [CrossRef]
- Aguilera, Y.; Liébana, R.; Herrera, T.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Benítez, V.; Martín-Cabrejas, M.A. Effect of illumination on the content of melatonin, phenolic compounds, and antioxidant activity during germination of lentils (Lens culinaris L.) and kidney beans (Phaseolus vulgaris L.). J. Agric. Food Chem. 2014, 62, 10736–10743. [Google Scholar] [CrossRef]
- Kennaway, D.J. Are the proposed benefits of melatonin-rich foods too hard to swallow? Crit. Rev. Food Sci. Nutr. 2017, 57, 958–962. [Google Scholar] [CrossRef]
- Bhattacharjee, P.; Probir, K.G.; Vernekar, M.; Singhal, R.S. Role of Dietary Serotonin and Melatonin in Human Nutrition. Serotonin and Melatonin; Taylor & Francis Group: Didcot, UK, 2016; pp. 317–332. [Google Scholar]
- Abeysuriya, R.G.; Lockley, S.W.; Robinson, P.A.; Postnova, S. A unified model of melatonin, 6-sulfatoxymelatonin, and sleep dynamics. J. Pineal Res. 2018, 64, e12474. [Google Scholar] [CrossRef]
- González-Flores, D.; Gamero, E.; Garrido, M.; Ramírez, R.; Moreno, D.; Delgado, J.; Valdés, E.; Barriga, C.; Rodríguez, A.B.; Paredes, S.D. Urinary 6-sulfatoxymelatonin and total antioxidant capacity increase after the intake of a grape juice cv. Tempranillo stabilized with HHP. Food Funct. 2012, 3, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Sae-Teaw, M.; Johns, J.; Johns, N.P.; Subongkot, S. Serum melatonin levels and antioxidant capacities after consumption of pineapple, orange, or banana by healthy male volunteers. J. Pineal Res. 2013, 55, 58–64. [Google Scholar] [CrossRef] [PubMed]
- López-Martínez, L.X.; Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Heredia, J.B. Effect of cooking and germination on bioactive compounds in pulses and their health benefits. J. Funct. Foods 2017, 38, 624–634. [Google Scholar] [CrossRef]
- Ganesan, K.; Xu, B. Polyphenol-rich lentils and their health promoting effects. Int. J. Mol. Sci. 2017, 11, 2390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilera, Y.; Herrera, T.; Liébana, R.; Rebollo-Hernanz, M.; Sanchez-Puelles, C.; Martín-Cabrejas, M.A. Impact of Melatonin Enrichment during Germination of Legumes on Bioactive Compounds and Antioxidant Activity. J. Agric. Food Chem. 2015, 63, 7967–7974. [Google Scholar] [CrossRef]
- Zhang, Y.; Huo, M.; Zhou, J.; Xie, S. PKSolver: An add-in program for pharmacokinetic and pharmacodynamic data analysis in Microsoft Excel. Comput. Methods Programs Biomed. 2010, 99, 306–314. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1998, 299, 152–178. [Google Scholar]
- Arnao, M.B.; Hernández-Ruiz, J. Phytomelatonin, natural melatonin from plants as a novel dietary supplement: Sources, activities and world market. J. Funct. Foods 2018, 48, 37–42. [Google Scholar] [CrossRef]
- Meng, X.; Li, Y.; Li, S.; Zhou, Y.; Gan, R.Y.; Xu, D.P.; Li, H.-B. Dietary sources and bioactivities of melatonin. Nutr. 2017, 9, 367. [Google Scholar] [CrossRef] [Green Version]
- Rebollo-Hernanz, M.; Fernández-Gómez, B.; Herrero, M.; Aguilera, Y.; Martín-Cabrejas, M.A.; Uribarri, J.; del Castillo, M.D. Inhibition of the Maillard Reaction by Phytochemicals Composing an Aqueous Coffee Silverskin Extract via a Mixed Mechanism of Action. Foods 2019, 8, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Jin, Z.; Peckrul, A.; Chen, B. Pulse seed germination improves antioxidative activity of phenolic compounds in stripped soybean oil-in-water emulsions. Food Chem. 2018, 250, 140–147. [Google Scholar] [CrossRef]
- Di Stefano, E.; Tsopmo, A.; Oliviero, T.; Fogliano, V.; Udenigwe, C.C. Bioprocessing of common pulses changed seed microstructures, and improved dipeptidyl peptidase-IV and α-glucosidase inhibitory activities. Sci. Rep. 2019, 9, 15308. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Rao, J.; Chen, B. Phenolic compounds in germinated cereal and pulse seeds: Classification, transformation, and metabolic process. Crit. Rev. Food Sci. Nutr. 2019, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Haytowitz, D.B.; Bhagwat, S. USDA Database for the Oxygen Radical Absorbance Capacity (ORAC) of Selected Foods, Release 2. US Dep. Agric. 2010, 25, 10–48. [Google Scholar]
- Aguilera, Y.; Herrera, T.; Benítez, V.; Arribas, S.M.; López de Pablo, A.L.; Esteban, R.M.; Martín-Cabrejas, M.A. Estimation of scavenging capacity of melatonin and other antioxidants: Contribution and evaluation in germinated seeds. Food Chem. 2015, 170, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Yoshiara, L.; Madeira, T.; de Camargo, A.; Shahidi, F.; Ida, E. Multistep Optimization of β-Glucosidase Extraction from Germinated Soybeans (Glycine max L. Merril) and Recovery of Isoflavone Aglycones. Foods 2018, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- López, A.; El-Naggar, T.; Dueñas, M.; Ortega, T.; Estrella, I.; Hernández, T.; Gómez-Serranillos, M.P.; Palomino, O.M.; Carretero, M.E. Influence of Processing in the Phenolic Composition and Health-Promoting Properties of Lentils (Lens culinaris L.). J. Food Process. Preserv. 2017, 41, e13113. [Google Scholar] [CrossRef]
- Aguilera, Y.; Rebollo-Hernanz, M.; Cañas, S.; Taladrid, D.; Martín-Cabrejas, M.A. Response surface methodology to optimise the heat-assisted aqueous extraction of phenolic compounds from coffee parchment and their comprehensive analysis. Food Funct. 2019, 10, 4739–4750. [Google Scholar] [CrossRef]
- Harpsøe, N.G.; Andersen, L.P.H.; Gögenur, I.; Rosenberg, J. Clinical pharmacokinetics of melatonin: A systematic review. Eur. J. Clin. Pharmacol. 2015, 71, 901–909. [Google Scholar] [CrossRef]
- Jana, S.; Rastogi, H. Effects of Caffeic Acid and Quercetin on In Vitro Permeability, Metabolism and In Vivo Pharmacokinetics of Melatonin in Rats: Potential for Herb-Drug Interaction. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 781–791. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Taxon- and site-specific Melatonin catabolism. Molecules 2017, 22, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghazadeh-Sanai, N.; Downs, J.L.; Mattison, J.A.; Ingram, D.K.; Kohama, S.G.; Urbanski, H.F. Effect of caloric restriction on plasma melatonin levels in aged rhesus macaques. Neurobiol. Aging 2017, 55, 213–216. [Google Scholar] [CrossRef] [PubMed]
- Almeneessier, A.S.; Bahammam, A.S. How does diurnal intermittent fasting impact sleep, daytime sleepiness, and markers of the biological clock? Current insights. Nat. Sci. Sleep 2018, 10, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Delgado, J.; Pilar Terrón, M.; Garrido, M.; Pariente, J.A.; Barriga, C.; Rodríguez, A.B.; Paredes, S.D. A cherry nutraceutical modulates melatonin, serotonin, corticosterone, and total antioxidant capacity levels: Effect on ageing and chronotype. J. Appl. Biomed. 2012, 10, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Bravo, R.; Matito, S.; Cubero, J.; Paredes, S.D.; Franco, L.; Rivero, M.; Rodríguez, A.B.; Barriga, C. Tryptophan-enriched cereal intake improves nocturnal sleep, melatonin, serotonin, and total antioxidant capacity levels and mood in elderly humans. Age 2013, 35, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Khazaei, H.; Subedi, M.; Nickerson, M.; Martínez-Villaluenga, C.; Frias, J.; Vandenberg, A. Seed Protein of Lentils: Current Status, Progress, and Food Applications. Foods 2019, 8, 391. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Manchester, L.C.; Tan, D.X. Melatonin in walnuts: Influence on levels of melatonin and total antioxidant capacity of blood. Nutrition 2005, 21, 920–924. [Google Scholar] [CrossRef]
- Johns, N.P.; Johns, J.; Porasuphatana, S.; Plaimee, P.; Sae-Teaw, M. Dietary Intake of Melatonin from Tropical Fruit Altered Urinary Excretion of 6-Sulfatoxymelatonin in Healthy Volunteers. J. Agric. Food Chem. 2013, 61, 913–919. [Google Scholar] [CrossRef]
- Maldonado, M.D.; Moreno, H.; Calvo, J.R. Melatonin present in beer contributes to increase the levels of melatonin and antioxidant capacity of the human serum. Clin. Nutr. 2009, 28, 188–191. [Google Scholar] [CrossRef]
- McClements, D.J. Enhancing nutraceutical bioavailability through food matrix design. Curr. Opin. Food Sci. 2015, 4, 1–6. [Google Scholar] [CrossRef]
- Hansen, M.B.; Witte, A.-B. The role of serotonin in intestinal luminal sensing and secretion. Acta Physiol. 2008, 193, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Martin, A.M.; Lumsden, A.L.; Young, R.L.; Jessup, C.F.; Spencer, N.J.; Keating, D.J. Regional differences in nutrient-induced secretion of gut serotonin. Physiol. Rep. 2017, 5, e13199. [Google Scholar] [CrossRef] [PubMed]
- Lund, M.L.; Egerod, K.L.; Engelstoft, M.S.; Dmytriyeva, O.; Theodorsson, E.; Patel, B.A.; Schwartz, T.W. Enterochromaffin 5-HT cells – A major target for GLP-1 and gut microbial metabolites. Mol. Metab. 2018, 11, 70–83. [Google Scholar] [CrossRef]
- Sumara, G.; Sumara, O.; Kim, J.K.; Karsenty, G. Gut-derived serotonin is a multifunctional determinant to fasting adaptation. Cell Metab. 2012, 16, 588–600. [Google Scholar] [CrossRef] [Green Version]
- Zagajewski, J.; Drozdowicz, D.; Brzozowska, I.; Hubalewska-Mazgaj, M.; Stelmaszynska, T.; Laidler, P.M.; Brzozowski, T. Conversion L-tryptophan to melatonin in the gastrointestinal tract: The new high performance liquid chromatography method enabling simultaneous determination of six metabolites of L-tryptophan by native fluorescence and UV-VIS detection. J. Physiol. Pharmacol. 2012, 63, 613–621. [Google Scholar]
- Fernstrom, J.D. A Perspective on the Safety of Supplemental Tryptophan Based on Its Metabolic Fates. J. Nutr. 2016, 146, 2601S–2608S. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, Y.; Yuan, B.; Liu, S.; Man, S.; Xu, H.; Lu, X. A novel LC-MS/MS assay for the simultaneous determination of melatonin and its two major metabolites, 6-hydroxymelatonin and 6-sulfatoxymelatonin in dog plasma: Application to a pharmacokinetic study. J. Pharm. Biomed. Anal. 2016, 117, 390–397. [Google Scholar] [CrossRef]
- Tian, X.; Huo, X.; Dong, P.; Wu, B.; Wang, X.; Wang, C.; Liu, K.; Ma, X. Sulfation of melatonin: Enzymatic characterization, differences of organs, species and genders, and bioactivity variation. Biochem. Pharmacol. 2015, 94, 282–296. [Google Scholar] [CrossRef]
- Oba, S.; Nakamura, K.; Sahashi, Y.; Hattori, A.; Nagata, C. Consumption of vegetables alters morning urinary 6-sulfatoxymelatonin concentration. J. Pineal Res. 2008, 45, 17–23. [Google Scholar] [CrossRef]
- Howatson, G.; Bell, P.G.; Tallent, J.; Middleton, B.; McHugh, M.P.; Ellis, J. Effect of tart cherry juice (Prunus cerasus) on melatonin levels and enhanced sleep quality. Eur. J. Nutr. 2012, 51, 909–916. [Google Scholar] [CrossRef] [PubMed]
- Cubero, J.; Valero, V.; Sánchez, J.; Rivero, M.; Parvez, H.; Rodríquez, A.B.; Barriga, C. The circadian rhythm of tryptophan in breast milk affects the rhythms of 6-sulfatoxymelatonin and sleep in newborn. Neuroendocrinol. Lett. 2005, 26, 657–661. [Google Scholar] [PubMed]
- Huether, G.; Poeggeler, B.; Reimer, A.; George, A. Effect of tryptophan administration on circulating melatonin levels in chicks and rats: Evidence for stimulation of melatonin synthesis and release in the gastrointestinal tract. Life Sci. 1992, 51, 945–953. [Google Scholar] [CrossRef]
- Huether, G. Melatonin Synthesis in the Gastrointestinal Tract and the Impact of Nutritional Factors on Circulating Melatonin. Ann. N. Y. Acad. Sci. 1994, 719, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, S.; Sánchez, C.L.; Paredes, S.D.; Rodriguez, A.B.; Barriga, C. The effect of tryptophan administration on the circadian rhythms of melatonin in plasma and the pineal gland of rats. J. Appl. Biomed. 2008, 6, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Tang, Y.; Chen, P.X.; Liu, R.; Dan Ramdath, D.; Liu, Q.; Hernandez, M.; Tsao, R. Bioaccessibility, in vitro antioxidant and anti-inflammatory activities of phenolics in cooked green lentil (Lens culinaris). J. Funct. Foods 2017, 32, 248–255. [Google Scholar] [CrossRef]
- Velderrain-Rodríguez, G.R.; Palafox-Carlos, H.; Wall-Medrano, A.; Ayala-Zavala, J.F.; Chen, C.Y.O.; Robles-Sánchez, M.; Astiazaran-García, H.; Alvarez-Parrilla, E.; González-Aguilar, G.A. Phenolic compounds: Their journey after intake. Food Funct. 2014, 5, 189–197. [Google Scholar] [CrossRef]
- Domínguez-Avila, J.A.; Wall-Medrano, A.; Velderrain-Rodríguez, G.R.; Chen, C.Y.O.; Salazar-López, N.J.; Robles-Sánchez, M.; González-Aguilar, G.A. Gastrointestinal interactions, absorption, splanchnic metabolism and pharmacokinetics of orally ingested phenolic compounds. Food Funct. 2017, 8, 15–38. [Google Scholar] [CrossRef]
- Hasslauer, I.; Oehme, A.; Locher, S.; Valotis, A.; Van’t Slot, G.; Humpf, H.-U.; Schreier, P. Flavan-3-ol C-glycosides—Preparation and model experiments mimicking their human intestinal transit. Mol. Nutr. Food Res. 2010, 54, 1546–1555. [Google Scholar] [CrossRef]
- Garrido, M.; Paredes, S.D.; Cubero, J.; Lozano, M.; Toribio-Delgado, A.F.; Munoz, J.L.; Reiter, R.J.; Barriga, C.; Rodriguez, A.B. Jerte Valley Cherry-Enriched Diets Improve Nocturnal Rest and Increase 6-Sulfatoxymelatonin and Total Antioxidant Capacity in the Urine of Middle-Aged and Elderly Humans. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 909–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selby-Pham, S.N.B.; Cottrell, J.J.; Dunshea, F.R.; Ng, K.; Bennett, L.E.; Howell, K.S. Dietary phytochemicals promote health by enhancing antioxidant defence in a pig model. Nutr. 2017, 9, 758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharopov, F.; Fokou, P.V.T.; Kobylinska, A.; Jonge, L.; Tadio, K.; Sharifi-Rad, J.; Posmyk, M.M.; Martorell, M.; Martins, N.; et al. Melatonin in Medicinal and Food Plants: Occurrence, Bioavailability, and Health Potential for Humans. Cells 2019, 8, 681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczyk-Golec, K.; Rajewski, P.; Gackowski, M.; Mila-Kierzenkowska, C.; Wesołowski, R.; Sutkowy, P.; Pawłowska, M.; Woźniak, A. Melatonin Supplementation Lowers Oxidative Stress and Regulates Adipokines in Obese Patients on a Calorie-Restricted Diet. Oxid. Med. Cell. Longev. 2017, 2017, 8494107. [Google Scholar] [CrossRef]
- Parkar, S.G.; Kalsbeek, A.; Cheeseman, J.F. Potential role for the gut microbiota in modulating host circadian rhythms and metabolic health. Microorganisms 2019, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pharmacokinetic Study | Bioavailability and Bioactivity Study | ||||
|---|---|---|---|---|---|
| Control | Fasting | Lentil Sprout | MEL | ||
| Group | 1 | 2 | 3 | 4 | 5 |
| Intervention | 12 h-fast Lentil sprout | None | 12 h-fast | 12 h-fast Lentil sprout | 12 h-fast Melatonin |
| Rat population (N) | 8 ♂ | 10 ♂ | 10 ♂ | 10 ♂ | 10 ♂ |
| Entrance into inverse photocycle (day) | 23 | 21 | 24 | 29 | 29 |
| Time in inverse photocycle (days) | 22 | 25 | 20 | 20 | 21 |
| Sampling day | 45 | 46 | 44 | 49 | 50 |
| Weight (g) | 195 ± 11 | 203 ± 11 | 197 ± 9 | 198 ± 10 | 202 ± 8 |
| Sampling time | 10:00 to 14:00 | 11:00 | 11:00 | 11:00 | 11:00 |
| Melatonin (µg) | FPC (mg GAE) | BPC (mg GAE) | TPC (mg GAE) | FPC: BPC (ratio) | ORAC (µmol TE) | |
|---|---|---|---|---|---|---|
| Raw lentils (per g) | (0.46 ± 0.06) · 10‒3 | 4.50 ± 0.17 | 0.38 ± 0.02 | 4.88 ± 0.19 | (12 ± 1):1 | 20.24 ± 1.54 |
| Lentil sprouts (per g) | 1.01 ± 0.09 *** | 3.15 ± 0.21 ** | 0.32 ± 0.03 * | 3.47 ± 0.24 ** | (10 ± 2):1 | 77.23 ± 5.78 *** |
| Dose (per kg) | 50.0 | 155.9 | 51.0 | 171.8 | - | 52.0 |
| Compounds | Rt (min) | λmax (nm) | Molecular ion [M-H]−(m/z) | MS2 (m/z) | Concentration (μg/g) | |||
| Free phenolics | Bound phenolics | |||||||
| Raw lentil | Lentil sprout | Raw lentil | Lentil sprout | |||||
| Hydroxybenzoic acids | ||||||||
| p-Hydroxybenzoic-hexoside | 5.48 | 255 | 299 | – | 8.14 ± 4.16 | 3.34 ± 0.20 | 34.32 ± 6.84 | 27.06 ± 1.58 |
| Protocatechuic acid | 6.70 | 257, 294 | 153 | – | 23.22 ± 2.52 | 4.36 ± 0.60*** | 26.34 ± 3.00 | 21.48 ± 10.92 |
| Total | 31.36 ± 6.68 | 7.70 ± 0.80** | 60.66 ± 9.84 | 48.54 ± 12.50 | ||||
| Hydroxycinnamic acids | ||||||||
| trans-p-Coumaric derivative | 10.04 | 309 | 417 | – | 18.94 ± 2.08 | 5.16 ± 1.08*** | nd | nd |
| p-Coumaroyl malic acid | 12.46 | 308 | 278 | 163 | 9.50 ± 1.06 | 11.10 ± 2.88 | nd | nd |
| p-Coumaroyl glycolic acid | 12.80 | 314 | 220 | 163 | 12.48 ± 2.32 | 13.68 ± 2.16 | nd | nd |
| p-Coumaric acid derivative | 13.10 | 312 | – | 163 | 13.08 ± 0.78 | nd*** | nd | nd |
| trans-p-Coumaric acid | 18.20 | 314 | 163 | – | 12.72 ± 0.00 | 8.78 ± 1.46** | 2.58 ± 0.00 | 2.64 ± 0.62 |
| trans-Ferulic acid | 20.4 | 322 | 193 | – | nd | nd | 3.18 ± 0.34 | 3.16 ± 1.04 |
| Total | 66.72 ± 6.24 | 38.72 ± 7.58** | 5.76 ± 0.34 | 5.80 ± 1.66 | ||||
| Catechins and procyanidins | ||||||||
| (+)-Catechin | 10.90 | 280 | 289 | 245, 203, 161 | 262.98 ± 37.20 | 150.06 ± 14.08** | 78.74 ± 14.54 | 27.94 ± 5.22*** |
| (+)-Catechin 3-O-hexoside | 9.02 | 280 | 451 | 289 | 298.58 ± 15.76 | 206.32 ± 5.26*** | nd | nd |
| Dimer prodelphinidin | 7.76 | 276 | 593 | 447, 441, 423, 305, 289 | 120.20 ± 2.54 | nd*** | nd | nd |
| Dimer procyanidin | 10.40 | 279 | 577 | 289 | 52.68 ± 0.02 | 23.86 ± 2.00*** | nd | nd |
| Total | 734.44 ± 55.52 | 380.24 ± 19.34*** | 78.74 ± 14.54 | 27.94 ± 5.22*** | ||||
| Flavonols and flavanone | ||||||||
| Kaempferol glucuronide dihexoside | 10.12 | 346 | 785 | 285 | nd | 2.56 ± 0.60** | nd | nd |
| Kaempferol dirutinoside | 14.05 | 346 | 901 | 755, 593, 285 | 408.60 ± 3.72 | 305.74 ± 10.04*** | 22.70 ± 4.72 | 8.44 ± 0.46** |
| Kaempferol rutinoside hexoside (I) | 15.00 | 345 | 755 | 593, 285 | 15.02 ± 0.26 | 8.92 ± 1.26** | 1.82 ± 0.12 | 1.90 ± 0.36 |
| Kaempferol rutinoside hexoside (II) | 17.04 | 346 | 755 | 593, 285 | nd | 7.10 ± 0.24*** | nd | nd |
| Kaempferol rutinoside rhamnoside (I) | 18.30 | 346 | 739 | 593, 285 | 12.68 ± 0.04 | 16.10 ± 1.00** | 0.82 ± 0.06 | 1.24 ± 0.26 |
| Kaempferol rutinoside rhamnoside (II) | 19.14 | 346 | 739 | 593, 285 | nd | 11.32 ± 3.08** | nd | nd |
| Quercetin 3-glucoside | 20.1 | 356 | 463 | 301 | 3.30 ± 0.26 | 3.02 ± 0.32 | nd | nd |
| Kaempferol glucuronide | 25.40 | 346 | 461 | 285 | 11.22 ± 2.48 | 14.10 ± 1.24 | nd | nd |
| Kaempferol rhamnoside | 34.10 | 346 | 431 | 285 | 10.52 ± 2.06 | 69.00 ± 8.54*** | 4.84 ± 0.46 | 8.76 ± 1.36*** |
| Eriodyctyol hexoside | 16.30 | 288, 338 (sh) | 449 | 287 | 8.92 ± 1.26 | 18.78 ± 0.96*** | nd | nd |
| Total | 470.26 ± 10.08 | 456.64 ± 27.28 | 30.18 ± 5.36 | 20.34 ± 2.44* | ||||
| Total phenolic compounds | 1302.78 ± 78.52 | 883.30 ± 55.00** | 175.34 ± 30.08 | 102.62 ± 21.82* | ||||
| Parameter | Oral lentil sprouts (50 µg melatonin/kg) |
|---|---|
| Tmax (min) | 90.0 ± 0.0 |
| Cmax (pg/mL) | 45.4 ± 3.5 |
| Ke (1/h) | 0.39 ± 0.05 |
| t1/2 (min) | 108.9 ± 13.0 |
| AUC 0-t (pg/mL min) | 6560 ± 963 |
| AUC 0-t /dose (pg/mL min)/(μg/kg) | 131.2 ± 19.3 |
| AUC 0-∞ (pg/mL min) | 9381 ± 2008 |
| AUC 0-t /AUC 0-∞ | 0.71 ± 0.05 |
| AUMC 0-∞ (pg/mL min2) | 1897356 ± 589030 |
| MRT 0-∞ (min) | 199.4 ± 19.8 |
| VD/F (μg/kg)/(pg/mL) | 0.85 ± 0.08 |
| Cl/F (μg/kg)/(pg/mL)/h | 0.33 ± 0.07 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rebollo-Hernanz, M.; Aguilera, Y.; Herrera, T.; Cayuelas, L.T.; Dueñas, M.; Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Arribas, S.M.; Martín-Cabrejas, M.A. Bioavailability of Melatonin from Lentil Sprouts and Its Role in the Plasmatic Antioxidant Status in Rats. Foods 2020, 9, 330. https://doi.org/10.3390/foods9030330
Rebollo-Hernanz M, Aguilera Y, Herrera T, Cayuelas LT, Dueñas M, Rodríguez-Rodríguez P, Ramiro-Cortijo D, Arribas SM, Martín-Cabrejas MA. Bioavailability of Melatonin from Lentil Sprouts and Its Role in the Plasmatic Antioxidant Status in Rats. Foods. 2020; 9(3):330. https://doi.org/10.3390/foods9030330
Chicago/Turabian StyleRebollo-Hernanz, Miguel, Yolanda Aguilera, Teresa Herrera, L. Tábata Cayuelas, Montserrat Dueñas, Pilar Rodríguez-Rodríguez, David Ramiro-Cortijo, Silvia M. Arribas, and María A. Martín-Cabrejas. 2020. "Bioavailability of Melatonin from Lentil Sprouts and Its Role in the Plasmatic Antioxidant Status in Rats" Foods 9, no. 3: 330. https://doi.org/10.3390/foods9030330