In Vitro α-Glucosidase and α-Amylase Inhibitory Activities of Free and Bound Phenolic Extracts from the Bran and Kernel Fractions of Five Sorghum Grain Genotypes

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Sample and Preparation
2.3. Phenolic Extraction
2.4. Total Phenolic Content (TPC)
2.5. Total Flavonoid Content (TFC)
2.6. Total Tannin Content (TTC)
2.7. α-Glucosidase Inhibitory Activity of Sorghum Phenolic Extract
2.8. α-Amylase Inhibitory Activity of Sorghum Phenolic Extract
2.9. The Inhibition Efficiency of Sorghum Phenolic Extract
2.10. Half-Maximal Inhibitory Concentration (IC50) Determination
2.11. Statistical Analysis
3. Results and Discussion
3.1. Phenolic Contents
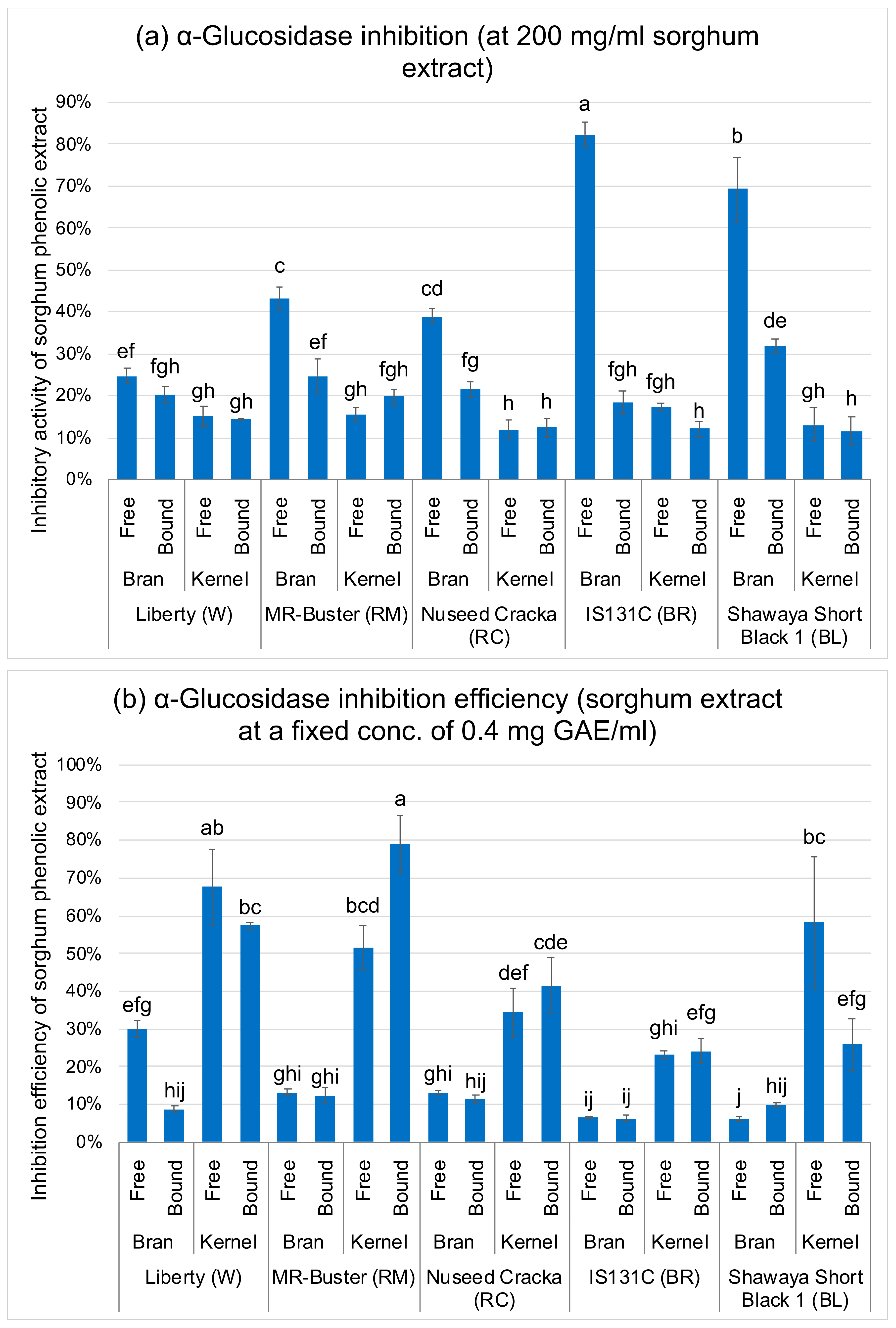
3.2. α-Glucosidase and α-Amylase Inhibition
3.3. Half-Maximal Inhibitory Concentration (IC50)
3.4. Inhibition Correlation with Sorghum Phenolic Compounds
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- American Diabetes Association. Diagnosis and classification of diabetes mellitus. Diabetes Care 2014, 37, S81–S90. [Google Scholar] [CrossRef] [Green Version]
- Sales, P.M.; Souza, P.M.; Simeoni, L.A.; Magalhães, P.O.; Silveira, D. α-Amylase inhibitors: A review of raw material and isolated compounds from plant source. J. Pharm. Pharm. Sci. 2012, 15, 141–183. [Google Scholar] [CrossRef] [Green Version]
- Standl, E.; Schnell, O. Alpha-glucosidase inhibitors 2012–cardiovascular considerations and trial evaluation. Diab. Vasc. Dis. Res. 2012, 9, 163–169. [Google Scholar] [CrossRef]
- Chiasson, J.-L.; Josse, R.G.; Gomis, R.; Hanefeld, M.; Karasik, A.; Laakso, M.; Group, S.-N.T.R. Acarbose for prevention of type 2 diabetes mellitus: The STOP-NIDDM randomised trial. Lancet 2002, 359, 2072–2077. [Google Scholar] [CrossRef]
- Coman, C.; Rugina, O.D.; Socaciu, C. Plants and natural compounds with antidiabetic action. Not. Bot. Horticult. Agrobot. 2012, 40, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Venn, B.J.; Mann, J.I. Cereal grains, legumes and diabetes. Eur. J. Clin. Nutr. 2004, 58, 1443–1461. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.T.; Hu, F.B.; Pereira, M.A.; Liu, S.; Stampfer, M.J.; Colditz, G.A.; Willett, W.C. Whole-grain intake and the risk of type 2 diabetes: A prospective study in men. Am. J. Clin. Nutr. 2002, 76, 535–540. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-S.; Hyun, T.K.; Kim, M.-J. The inhibitory effects of ethanol extracts from sorghum, foxtail millet and proso millet on α-glucosidase and α-amylase activities. Food Chem. 2011, 124, 1647–1651. [Google Scholar] [CrossRef]
- FAO. FAOSTAT, Food and Agriculture Organization of the United Nation. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 12 September 2020).
- Schittenhelm, S.; Schroetter, S. Comparison of drought tolerance of maize, sweet sorghum and sorghum-sudangrass hybrids. J. Agron. Crop. Sci. 2014, 200, 46–53. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. Sorghum Grain: From Genotype, Nutrition, and Phenolic Profile to Its Health Benefits and Food Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 2025–2046. [Google Scholar] [CrossRef] [Green Version]
- Ragaee, S.; Abdel-Aal, E.-S.M.; Noaman, M. Antioxidant activity and nutrient composition of selected cereals for food use. Food Chem. 2006, 98, 32–38. [Google Scholar] [CrossRef]
- Chung, I.-M.; Kim, E.-H.; Yeo, M.-A.; Kim, S.-J.; Seo, M.C.; Moon, H.-I. Antidiabetic effects of three Korean sorghum phenolic extracts in normal and streptozotocin-induced diabetic rats. Food Res. Int. 2011, 44, 127–132. [Google Scholar] [CrossRef]
- Kim, J.; Park, Y. Anti-diabetic effect of sorghum extract on hepatic gluconeogenesis of streptozotocin-induced diabetic rats. Nutr. Metab. 2012, 9, 106. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, I.M.; Li-Chan, E.C. Overview of food products and dietary constituents with antidiabetic properties and their putative mechanisms of action: A natural approach to complement pharmacotherapy in the management of diabetes. Mol. Nutr. Food Res. 2014, 58, 61–78. [Google Scholar] [CrossRef]
- Hargrove, J.L.; Greenspan, P.; Hartle, D.K.; Dowd, C. Inhibition of aromatase and α-amylase by flavonoids and proanthocyanidins from Sorghum bicolor bran extracts. J. Med. Food 2011, 14, 799–807. [Google Scholar] [CrossRef]
- Mkandawire, N.L.; Kaufman, R.C.; Bean, S.R.; Weller, C.L.; Jackson, D.S.; Rose, D.J. Effects of sorghum (Sorghum bicolor (L.) Moench) tannins on alpha-amylase activity and in vitro digestibility of starch in raw and processed flours. J. Agric. Food Chem. 2013, 61, 4448–4454. [Google Scholar] [CrossRef] [PubMed]
- Links, M.R.; Taylor, J.; Kruger, M.C.; Taylor, J.R.N. Sorghum condensed tannins encapsulated in kafirin microparticles as a nutraceutical for inhibition of amylases during digestion to attenuate hyperglycaemia. J. Funct. Foods 2015, 12, 55–63. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Shen, S.; Johnson, S.; Fang, Z. Comprehensive profiling of phenolic compounds by HPLC-DAD-ESI-QTOF-MS/MS to reveal their location and form of presence in different sorghum grain genotypes. Food Res. Int. 2020. (Accepted). [Google Scholar]
- Xiong, Y.; Zhang, P.; Luo, J.; Johnson, S.; Fang, Z. Effect of processing on the phenolic contents, antioxidant activity and volatile compounds of sorghum grain tea. J. Cereal Sci. 2019, 85, 6–14. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Price, M.L.; van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agricult. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Ng, K.; Gu, C.; Zhang, H.; Putri, C.Y. Evaluation of α-amylase and α-glucosidase inhibitory activity of flavonoids. Int. J. Food Nutr. Sci. 2015, 2, 174–179. [Google Scholar] [CrossRef]
- Sirisena, S.; Ng, K.; Ajlouni, S. Antioxidant activities and inhibitory effects of free and bound polyphenols from date (Phoenix dactylifera L.) seeds on starch digestive enzymes. Int. J. Food Stud. 2016, 5. [Google Scholar] [CrossRef]
- Phan, M.A.T.; Wang, J.; Tang, J.; Lee, Y.Z.; Ng, K. Evaluation of α-glucosidase inhibition potential of some flavonoids from Epimedium brevicornum. LWT-Food Sci. Technol. 2013, 53, 492–498. [Google Scholar] [CrossRef]
- Abdi, H.; Williams, L.J. Partial Least Squares Methods: Partial Least Squares Correlation and Partial Least Square Regression. In Computational Toxicology: Volume II; Reisfeld, B., Mayeno, A.N., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 549–579. [Google Scholar] [CrossRef]
- Bhagwat, S.; Haytowitz, D. USDA Database for the Proanthocyanidin Content of Selected Foods, Release 2; US Department of Agriculture: Beltsville, MD, USA, 2015; pp. 1–45.
- Wu, G.; Johnson, S.K.; Bornman, J.F.; Bennett, S.J.; Clarke, M.W.; Singh, V.; Fang, Z. Growth temperature and genotype both play important roles in sorghum grain phenolic composition. Sci. Rep. 2016, 6, 21835. [Google Scholar] [CrossRef]
- Wu, G.; Bennett, S.J.; Bornman, J.F.; Clarke, M.W.; Fang, Z.; Johnson, S.K. Phenolic profile and content of sorghum grains under different irrigation managements. Food Res. Int. 2017, 97, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, P.; Warner, R.D.; Fang, Z. 3-Deoxyanthocyanidin colorant: Nature, health, synthesis and food applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1533–1549. [Google Scholar] [CrossRef] [Green Version]
- Etxeberria, U.; de la Garza, A.L.; Campión, J.; Martinez, J.A.; Milagro, F.I. Antidiabetic effects of natural plant extracts via inhibition of carbohydrate hydrolysis enzymes with emphasis on pancreatic alpha amylase. Expert Opin. Ther. 2012, 16, 269–297. [Google Scholar] [CrossRef] [Green Version]
- Shinde, J.; Taldone, T.; Barletta, M.; Kunaparaju, N.; Hu, B.; Kumar, S.; Placido, J.; Zito, S.W. α-Glucosidase inhibitory activity of Syzygium cumini (Linn.) Skeels seed kernel in vitro and in Goto–Kakizaki (GK) rats. Carbohydr. Res. 2008, 343, 1278–1281. [Google Scholar] [CrossRef]
- Kim, K.Y.; Nguyen, T.H.; Kurihara, H.; Kim, S.M. α-glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J. Food Sci. 2010, 75, H145–H150. [Google Scholar] [CrossRef]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Ali Asgar, M.D. Anti-diabetic potential of phenolic compounds: A review. Int. J. Food Prop. 2013, 16, 91–103. [Google Scholar] [CrossRef]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of dietary polyphenols on carbohydrate metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [Green Version]
- Funke, I.; Melzig, M.F. Effect of different phenolic compounds on alpha-amylase activity: Screening by microplate-reader based kinetic assay. Pharmazie 2005, 60, 796–797. [Google Scholar]
- Xiao, J.; Ni, X.; Kai, G.; Chen, X. A review on structure–activity relationship of dietary polyphenols inhibiting α-amylase. Crit. Rev. Food Sci. Nutr. 2013, 53, 497–506. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| α-Glucosidase | α-Amylase | |||||
|---|---|---|---|---|---|---|
| IC50 (mg sorghum/mL) * | IC50 (mg GAE/mL) ** | IC50 (mg sorghum/mL) * | IC50 (mg GAE/mL) ** | |||
| Liberty (W) | Bran | Free | 301 ± 30 | 0.50 ± 0.05 | >350 | >0.58 |
| Bound | >300 | >1.38 | >350 | >1.61 | ||
| Kernel | Free | >300 | >0.14 | >350 | >0.16 | |
| Bound | >300 | >0.15 | >350 | >0.18 | ||
| MR-Buster (RM) | Bran | Free | 222 ± 10 | 1.45 ± 0.06 | 260 ± 17 | 1.70 ± 0.11 |
| Bound | 326 ± 74 | 1.30 ± 0.29 | 327 ± 74 | 1.31 ± 0.29 | ||
| Kernel | Free | >300 | >0.18 | 249 ± 27 | 0.15 ± 0.02 | |
| Bound | >300 | >0.15 | 160 ± 12 | 0.08 ± 0.01 | ||
| Nuseed Cracka (RC) | Bran | Free | 259 ± 21 | 1.53 ± 0.12 | 320 ± 60 | 1.89 ± 0.36 |
| Bound | >300 | >1.14 | 350 ± 27 | 1.33 ± 0.10 | ||
| Kernel | Free | >300 | >0.21 | >350 | >0.25 | |
| Bound | >300 | >0.18 | >350 | >0.21 | ||
| IS131C (BR) | Bran | Free | 18 ± 11 | 0.47 ± 0.28 | 236 ± 7 | 5.98 ± 0.17 |
| Bound | >300 | >1.73 | 237 ± 9 | 1.37 ± 0.05 | ||
| Kernel | Free | >300 | >0.45 | 305 ± 22 | 0.46 ± 0.03 | |
| Bound | >300 | >0.30 | 216 ± 8 | 0.22 ± 0.01 | ||
| Shawaya Short Black 1 (BL) | Bran | Free | 105 ± 27 | 2.36 ± 0.60 | 256 ± 45 | 5.75 ± 1.00 |
| Bound | 297 ± 27 | 1.92 ± 0.18 | 318 ± 67 | 2.05 ± 0.43 | ||
| Kernel | Free | >300 | >0.14 | 370 ± 99 | 0.17 ± 0.04 | |
| Bound | >300 | >0.27 | 335 ± 11 | 0.30 ± 0.01 | ||
| Acarbose | 1.39 ± 0.23 mg acarbose/mL | 0.50 ± 0.03 mg acarbose/mL | ||||
| Variables | Glu | Amy | TPC | TFC | TTC | W | RM | RC | BR | BL | Bran | Kernel | Free | Bound |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Glu 1 | 1.000 | 0.305 | 0.961 ** | 0.954 ** | 0.955 ** | −0.194 | −0.004 | −0.124 | 0.175 | 0.148 | 0.617 ** | −0.617 ** | 0.384 | −0.384 |
| Amy 1 | 0.305 | 1.000 | 0.288 | 0.310 | 0.289 | −0.453 * | 0.447 * | −0.362 | 0.369 | −0.001 | 0.012 | −0.012 | −0.011 | 0.011 |
| TPC | 0.961 ** | 0.288 | 1.000 | 0.990 ** | 0.960 ** | −0.213 | −0.131 | −0.143 | 0.274 | 0.212 | 0.584 ** | −0.584 ** | 0.276 | −0.276 |
| TFC | 0.954 ** | 0.310 | 0.990 ** | 1.000 | 0.941 ** | −0.222 | −0.121 | −0.151 | 0.280 | 0.213 | 0.478 * | −0.478 * | 0.323 | −0.323 |
| TTC | 0.955 ** | 0.289 | 0.960 ** | 0.941 ** | 1.000 | −0.321 | −0.134 | −0.040 | 0.228 | 0.267 | 0.610 ** | −0.610 ** | 0.311 | −0.311 |
| W 2 | −0.194 | −0.453 * | −0.213 | −0.222 | −0.321 | 1.000 | −0.250 | −0.250 | −0.250 | −0.250 | 0.000 | 0.000 | 0.000 | 0.000 |
| RM 2 | −0.004 | 0.447 * | −0.131 | −0.121 | −0.134 | −0.250 | 1.000 | −0.250 | −0.250 | −0.250 | 0.000 | 0.000 | 0.000 | 0.000 |
| RC 2 | −0.124 | −0.362 | −0.143 | −0.151 | −0.040 | −0.250 | −0.250 | 1.000 | −0.250 | −0.250 | 0.000 | 0.000 | 0.000 | 0.000 |
| BR 2 | 0.175 | 0.369 | 0.274 | 0.280 | 0.228 | −0.250 | −0.250 | −0.250 | 1.000 | −0.250 | 0.000 | 0.000 | 0.000 | 0.000 |
| BL 2 | 0.148 | −0.001 | 0.212 | 0.213 | 0.267 | −0.250 | −0.250 | −0.250 | −0.250 | 1.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Bran 3 | 0.617 ** | 0.012 | 0.584 ** | 0.478 * | 0.610 ** | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1.000 | −1.000 ** | 0.000 | 0.000 |
| Kernel 3 | −0.617 ** | −0.012 | −0.584 ** | −0.478 * | −0.610 ** | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | −1.000 ** | 1.000 | 0.000 | 0.000 |
| Free 4 | 0.384 | −0.011 | 0.276 | 0.323 | 0.311 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1.000 | −1.000 ** |
| Bound 4 | −0.384 | 0.011 | −0.276 | −0.323 | −0.311 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | −1.000 ** | 1.000 |
| Proanthocyanidin dimer5 | 0.842 ** | 0.295 | 0.880 ** | 0.907 ** | 0.784 ** | −0.156 | −0.156 | −0.156 | 0.415 | 0.052 | 0.292 | −0.292 | 0.311 | −0.311 |
| Flavonol 5 | 0.722 ** | 0.267 | 0.739 ** | 0.755** | 0.650 ** | −0.143 | −0.132 | −0.141 | 0.526 * | −0.110 | 0.273 | −0.273 | 0.263 | −0.263 |
| Flavone 5 | 0.269 | −0.077 | 0.284 | 0.171 | 0.323 | −0.126 | 0.040 | 0.097 | −0.135 | 0.124 | 0.745 ** | −0.745 ** | −0.364 | 0.364 |
| Flavanone 5 | 0.892 ** | 0.260 | 0.879 ** | 0.851 ** | 0.875 ** | −0.306 | 0.025 | −0.112 | 0.253 | 0.140 | 0.639 ** | −0.639 ** | 0.167 | −0.167 |
| Flavan-3-ol 5 | 0.581 ** | 0.160 | 0.609 ** | 0.584 ** | 0.529 * | −0.285 | 0.134 | 0.020 | 0.339 | −0.208 | 0.529 * | −0.529 * | −0.017 | 0.017 |
| Anthocyanidin 5 | −0.033 | 0.148 | 0.047 | −0.035 | 0.161 | −0.187 | −0.187 | −0.187 | 0.292 | 0.268 | 0.287 | −0.287 | −0.373 | 0.373 |
| 3-Deoxyanthocyanidin 5 | 0.895 ** | 0.208 | 0.789 ** | 0.778 ** | 0.859 ** | −0.305 | 0.117 | 0.068 | 0.073 | 0.047 | 0.591 ** | −0.591 ** | 0.453 * | −0.453 * |
| Hydroxybenzoic acid 5 | 0.484 * | −0.070 | 0.412 | 0.330 | 0.454 * | 0.244 | −0.141 | −0.026 | −0.171 | 0.094 | 0.839 ** | −0.839 ** | 0.174 | −0.174 |
| Hydroxycinnamic acid 5 | 0.318 | −0.117 | 0.342 | 0.231 | 0.311 | 0.001 | 0.148 | 0.008 | −0.171 | 0.014 | 0.807 ** | −0.807 ** | −0.229 | 0.229 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Y.; Ng, K.; Zhang, P.; Warner, R.D.; Shen, S.; Tang, H.-Y.; Liang, Z.; Fang, Z. In Vitro α-Glucosidase and α-Amylase Inhibitory Activities of Free and Bound Phenolic Extracts from the Bran and Kernel Fractions of Five Sorghum Grain Genotypes. Foods 2020, 9, 1301. https://doi.org/10.3390/foods9091301
Xiong Y, Ng K, Zhang P, Warner RD, Shen S, Tang H-Y, Liang Z, Fang Z. In Vitro α-Glucosidase and α-Amylase Inhibitory Activities of Free and Bound Phenolic Extracts from the Bran and Kernel Fractions of Five Sorghum Grain Genotypes. Foods. 2020; 9(9):1301. https://doi.org/10.3390/foods9091301
Chicago/Turabian StyleXiong, Yun, Ken Ng, Pangzhen Zhang, Robyn Dorothy Warner, Shuibao Shen, Hsi-Yang Tang, Zijian Liang, and Zhongxiang Fang. 2020. "In Vitro α-Glucosidase and α-Amylase Inhibitory Activities of Free and Bound Phenolic Extracts from the Bran and Kernel Fractions of Five Sorghum Grain Genotypes" Foods 9, no. 9: 1301. https://doi.org/10.3390/foods9091301