1. Introduction
Cardiorespiratory fitness (CRF) is positively associated with endurance exercise performance [
1] and is a strong prognostic factor of morbidity and mortality from all causes and, particularly, from cardiovascular disease (CVD) [
2,
3]. While both physical activity (PA) and exercise training can modify CRF and are inversely associated with morbidity and mortality rates [
4], CRF per se is a much stronger predictor of prognosis in CVD and metabolic disorders [
5,
6]. The measure of an individual’s peak capacity to perform dynamic aerobic exercise is dependent on the synergistic action of pulmonary, cardiovascular and muscle tissue via a suite of physiological actions that effectively transport and deliver oxygen from the atmosphere to mitochondria in working muscles [
7,
8]. Accordingly, CRF can be assessed by directly measuring the peak oxygen uptake (VO
2peak) reached during a graded dynamic exercise test until exhaustion, involving large muscle masses (e.g., running or bicycling), or by indirectly estimating this variable from the peak workload achieved. Nevertheless, direct evaluation of VO
2peak is considered the gold standard measure of CRF and, indeed, the American Heart Association recently advocated for the routine assessment of this measure as a clinical vital sign [
9].
VO
2peak is characterized by a high interindividual variability even in people of the same sex, age and with the same level of PA and exercise training. This variability is believed to be related, at least partly, to heredity. A seminal study by Claude Bouchard and colleagues found comparable VO
2peak values in brothers of the same sibship, and the similarities in VO
2peak were even greater in dizygotic and monozygotic twins [
10]. The authors suggested that the genetic effect on VO
2peak reached ~40%. In a similar study of 170 individuals and their offspring (n = 259), it was found that about 50% of the interindividual variance in VO
2peak corresponded to heritable factors after adjusting for age, sex, body mass, and body composition [
11]. These findings have been replicated in subsequent studies with siblings and twins [
12] and, to date, it is commonly accepted that VO
2peak is influenced by both genetic (~50–60%) and environmental factors. It has also been reported that twins with similar VO
2peak values present with comparable levels of a variety of PA indices [
13], suggesting that part of the heritability of VO
2peak in twins might be due to the similarity of their PA levels. In fact, in a recent analysis of 123,545 single nucleotide polymorphisms (SNPs), only nine were associated with VO
2peak [
14]. The authors of this study found that those individuals whose genotype was associated with a high VO
2peak value had a lower CVD risk (e.g., less visceral fat or lower total blood cholesterol), but they did not calculate the additive effect that the nine SNPs had on the interindividual variability of VO
2peak. There is therefore controversy on the influence of genetics on VO
2peak, which mostly likely stems from the discrepancies between studies conducted on siblings/twins vs those conducted on individuals with no familial connection. In this regard, determining the actual genetic contribution to the interindividual variability in VO
2peak would be of major importance to inform how environmental factors—including lifestyle—might contribute to heightened VO
2peak values. It is possible that if the influence of genetics on VO
2peak is low, exercise training might be a determining factor to enhance ‘innate’ VO
2peak even in those less genetically predisposed, with obvious subsequent benefits for cardiovascular health. Indeed, previous research has reported VO
2peak increases of up to 44% after strenuous training interventions, which would support a strong influence of environmental factors on CRF [
15].
Aerobic/endurance exercise-based training appears to be the most effective way to augment VO
2peak. Exercise training increases rather than decreases the individual differences seen at baseline VO
2peak because the response to training itself shows large variation [
16]. A genome-wide association study based on 324,611 SNPs found that only 21 SNPs could explain 48.6% of the change in VO
2peak induced by a 20-week exercise training program [
17]. Among them, rs6552828, located in the acyl-CoA synthase long-chain member 1 (
ACSL1) gene, accounted by itself for 6% of the training-induced enhancement in VO2
peak. In a recent meta-analysis of 35 articles on the genetic influence on VO
2peak trainability, a total of 97 genes were associated with this phenotype, although only 13 genetic variants were reproduced by more than two investigations [
18].
Knowledge on the genetic influence on baseline VO2peak (i.e., in isolation from training) is mainly based on studies conducted on siblings/twins or in individuals of Caucasian descent, and it remains to be determined whether the genetic variants that might be associated with baseline VO2peak are similar or different in individuals of other ethnicities. Thus, the aim of the present study was to assess the influence of several candidate genetic variants in the interindividual variation of baseline CRF measured as VO2peak, in Han Chinese individuals.
3. Results
Of the initial 1047 individuals recruited, valid VO
2peak measurements were obtained for 1009 individuals (566 women), and thus only these participants were included in the analyses. The main characteristics of the participants are shown in
Table 2.
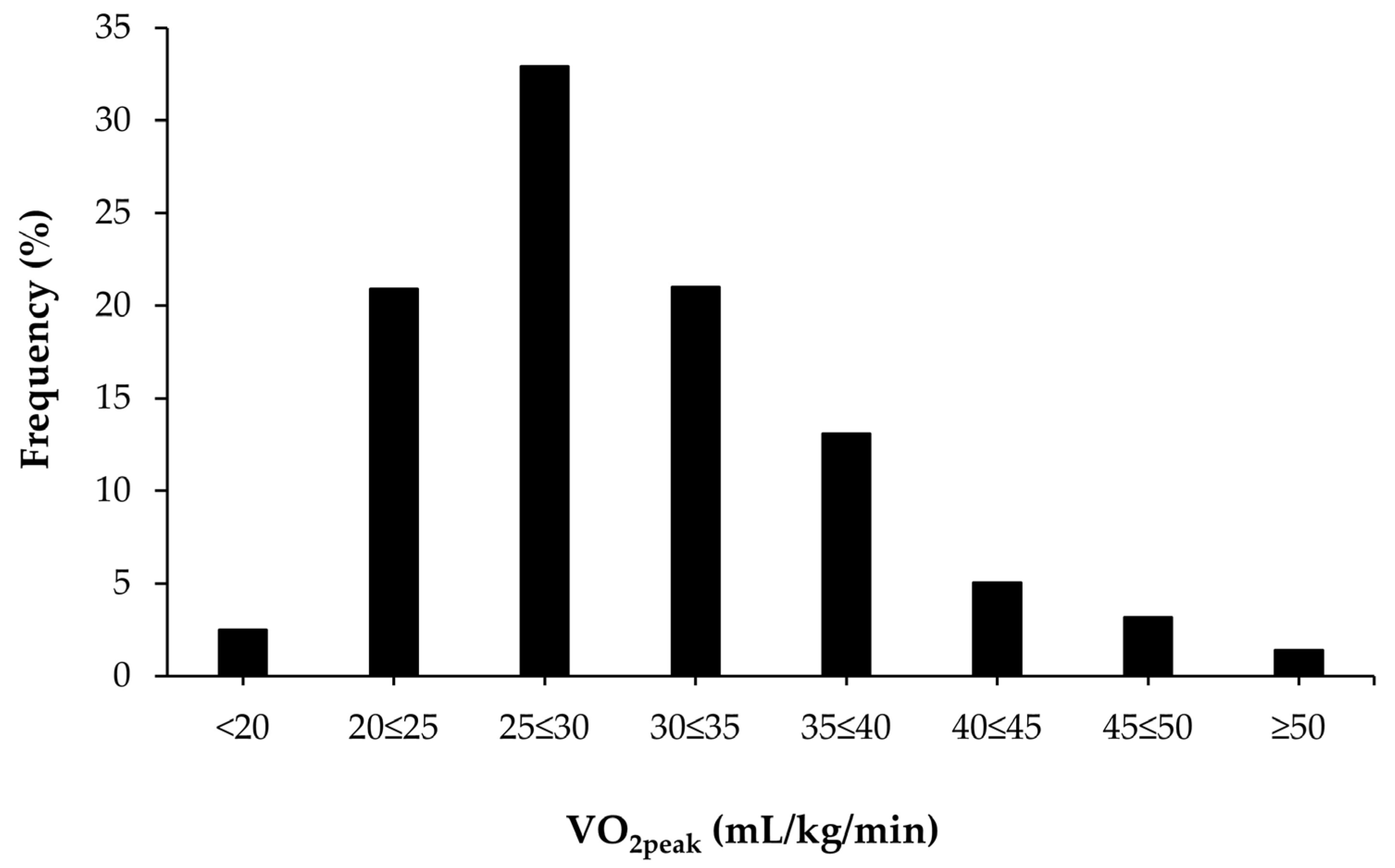
Figure 1 shows the distribution of VO
2peak values in the study sample. Approximately 2.5% of all participants had a VO
2peak <20 mL/kg/min and 1.4% had a VO
2peak level >50 mL/kg/min.
Genotyping was successful (i.e., successful determinations for all SNPs) in 1006 of 1009 participants (99.7%). From the 125 SNPs analyzed, 10 were discarded because they deviated from HWE (
Table A2), 10 because they had a MAF <5% (
Table A3), and two because only one genotype was detected across the group of participants (
Table A4). From the remaining pool of 103 SNPs, only rs4295, located in the angiotensin-converting enzyme (
ACE) gene, was significantly associated with VO
2peak (
p < 2.9 × 10
−4, β = 0.87; minor allele (G) frequency, 38.1%, heterozygosity frequency, 47.4%).
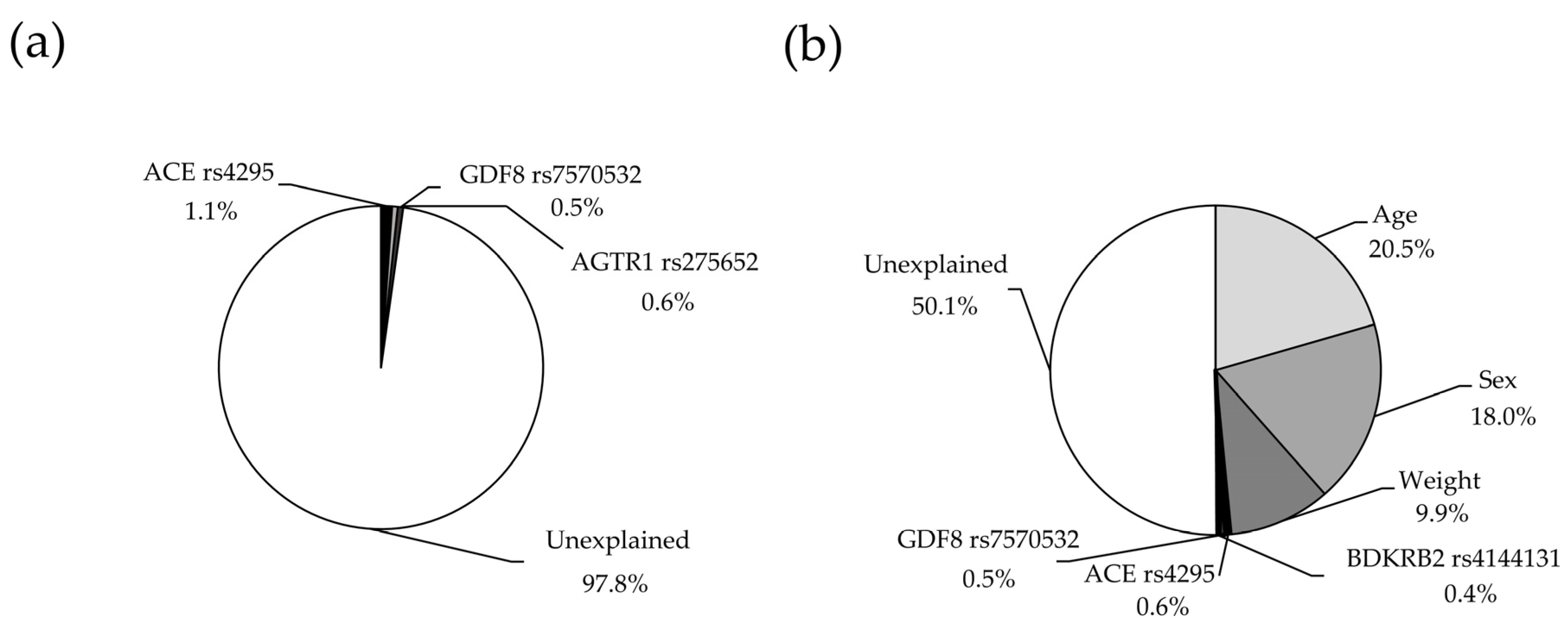
In multiple regression analysis, and after excluding those SNPs with collinearity, only three were retained in the final model (
ACE rs4295,
AGTR1 rs275652,
GDF8 rs7570532), which explained together 2.2% (
p = 0.0007) of the variance in VO
2peak (
Figure 2a, statistical power = 0.987). The partial contribution of each SNP to the variance in VO
2peak is shown in
Table 3. The explained variance of VO
2peak increased to 50.1% (
p < 0.0001) when including covariates such as age, sex and weight in the model (
Figure 2b, statistical power = 1.00).
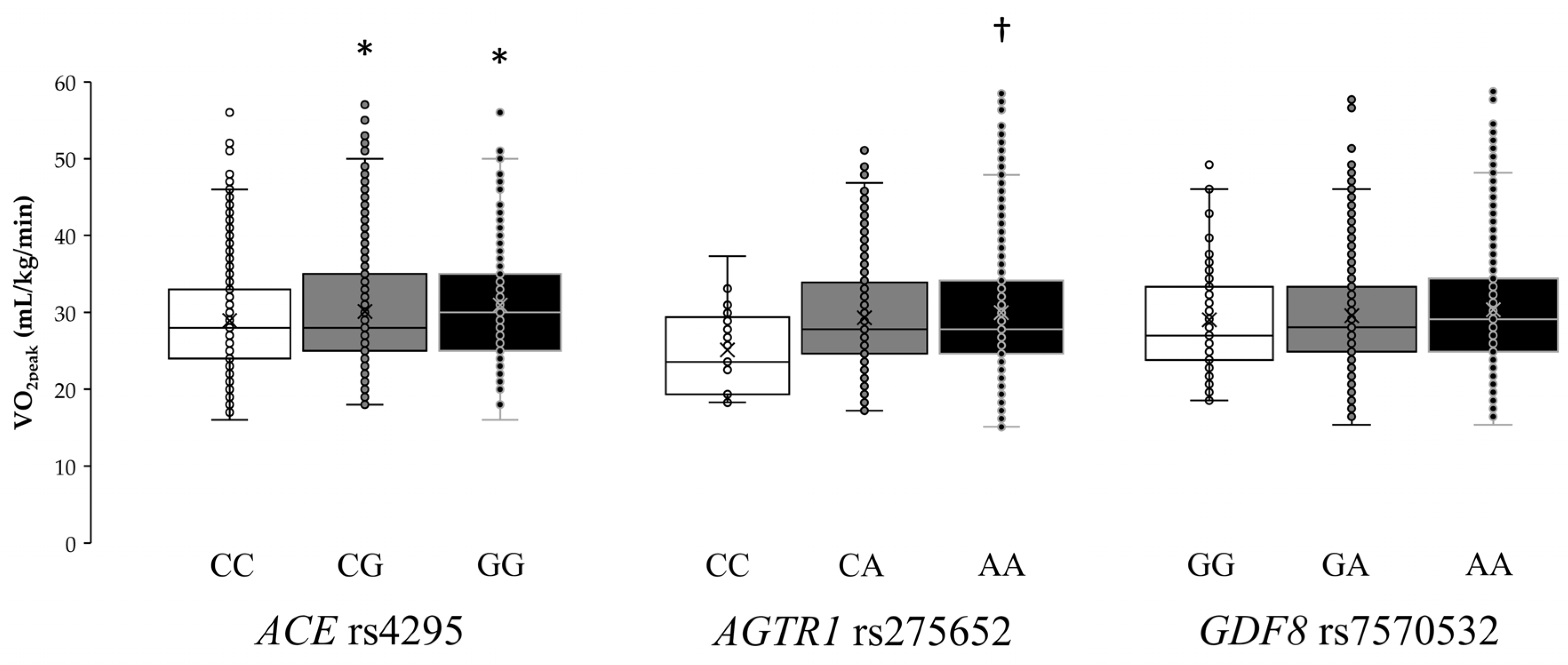
Individual VO
2peak values for each genotype of the
ACE rs4295,
AGTR1 rs275652, and
GDF8 rs7570532 polymorphisms are shown in
Figure 3. The one-way ANOVA revealed statistically significant differences in
ACE rs4295 (F = 4.95,
p = 0.007) and
AGTR1 rs275652 (F = 3.90,
p = 0.021) polymorphisms, while the ANOVA did not show differences for
GDF8 rs7570532 (F = 1.64,
p = 0.194) polymorphism. Specifically, GG homozygotes in
ACE rs4295 had a mean VO
2peak of 31.1 ± 7.9 mL/kg/min, which was higher than that found in heterozygotes (GC, 29.8±6.9 mL/kg/min;
p = 0.049) or in homozygotes for the common allele (CC, 28.9 ± 6.7 mL/kg/min;
p = 0.013). In addition, AA homozygotes in
AGTR1 rs275652 had a mean VO
2peak of 30.0 ± 7.3 mL/kg/min, which was higher than that found in homozygotes for the minor allele (CC, 25.5 ± 5.3 mL/kg/min;
p = 0.024).
A weighted genotype score was constructed using the three SNPs shown in model 1 of genetic-only influence. Participants were categorized with a genotype score from 0 a.u., indicating the presence of homozygosity for all the alleles associated with a lower VO
2peak in
ACE (rs4295),
AGTR1 (rs275652) and
GDF8 (rs7570532), to 6 a.u., indicating the presence of homozygosity for all the alleles associated with a higher VO
2peak in the aforementioned SNPs. A linear effect was found for genotype score on VO
2peak (
Figure 4). Specifically, the individuals with 6 a.u. had a higher VO
2peak than those with scores up to 4.0 a.u. (
p < 0.05). In addition, participants with scores >2 a.u. had a higher VO
2peak than those with scores <1.0 a.u. (
p < 0.05). ROC analysis showed significant discriminatory accuracy of the weighted genotype score in the identification of individuals with low/intermediate CRF (AUC = 0.542) with a sensitivity of 0.733 and a specificity of 0.305.
4. Discussion
CRF, particularly when objectively determined as VO
2peak, is strongly associated with endurance performance and health outcomes. Indeed, VO
2peak reflects the peak integrative ability of the organism to deliver oxygen from the atmosphere to the mitochondria of working muscles. The VO
2peak is thus determined, among other factors, by peak cardiac output and pulmonary ventilation, lung diffusion capacity, blood and plasma volume, hemoglobin mass, and muscle capillary density and oxidative capacity [
53]. Importantly, the mean values of VO
2peak of our participants (29.9 ± 7.1 mL/kg/min or 8.5 metabolic equivalents, i.e., METs) were barely above the minimum healthy threshold for all-cause and CVD mortality in middle-aged men/women (i.e., 8 METs [
52]). It is thus of medical importance to determine whether genetic factors (including specific gene variants) are associated with variability of CRF around (i.e., above vs below) the 8-MET cutoff. Previous research in siblings/twins suggests that 50%–60% of the variance of VO
2peak is associated with heredity [
10,
12]. These values seem surprisingly high given the variety of physiological processes and body tissues involved in the uptake and utilization of oxygen in muscle mitochondria. Indeed, there is open debate about the limits of the evidence that support the relative influence of genetics on the variability and trainability of CRF [
54,
55].
Our findings question the high heritability of VO
2peak, at least in Chinese individuals with no familial connection. From the 125 SNPs selected for our study, only one (
ACE rs4295) was associated with VO
2peak. Also, the best model obtained through multiple regression analyses could only explain ~2.2% of the interindividual variance in VO
2peak. As in the study by Bye et al. [
14], we created a polygenic score to determine whether those individuals with a higher number of alleles associated with VO
2peak did indeed present with higher values of this parameter. The only differences found between our findings and those of the Bye et al. study were the number of SNPs included in the polygenic score (7 vs 3, respectively) and the use of an intermediate genotype score for heterozygotes, which was not included by Bye et al. Interestingly, in both studies, participants with the theoretically lowest (or ‘less favorable’) genotype scores had the lowest VO
2peak (22–24 mL/kg/min), which was significantly lower than for those with the theoretically highest (or ‘most favorable’) genotype score (~32 mL/kg/min). These findings suggest that only a small number of SNPs are associated with the odds of having high VO
2peak values in untrained individuals. The variance of the interindividual variability in VO
2peak explained with these genotypes is low and the addition of favorable alleles might produce a change of 8–10 mL/kg/min. This genetic influence might be considerable in clinical terms because each 1-MET (or 3.5 mL/kg/min) increase in CRF has been shown to confer a 12% improvement in survival in Caucasian (North-American) men [
6]. Moreover, as mentioned above, it is of clinical importance to surpass the 8-MET threshold, and in fact, adults with a CRF clearly above this level (>10 METs) have a remarkably reduced CVD risk [
56]. In this regard, the probability of surpassing the 8-MET threshold (equivalent to 28 mL/kg/min) was doubled in those participants that carried the six ‘favorable’ alleles (
Figure 4).
Only three SNPs were included in the final multiple regression model.
ACE rs4295 has not been previously associated with endurance performance, but it is located in the same linkage disequilibrium block as the widely studied
ACE insert(I)/deletion(D) polymorphism (rs4340) [
57]. The
ACE gene encodes angiotensin-converting enzyme and the I allele might be associated with lower circulating levels of enzyme, and the II genotype potentially associated with performance in endurance athletes (odds ratio 1.35; 95% confidence interval, 1.17 to 1.55 [
58]). However, several studies have found no association between the
ACE I/D genotype and VO
2peak values in trained [
59] and untrained [
60,
61] individuals. With regard to the
ACE rs4295 variation found in the present study, although its influence on CRF needs to be replicated in other cohorts, our findings bolster the role of angiotensin-converting enzyme and its coding gene as predictors of CRF-related phenotypes. We also found that carriage of the C allele in the
AGTR1 rs275652 polymorphism was negatively associated with VO
2peak values. This gene encodes the angiotensin II receptor 1 (AT
1R), and polymorphisms in
AGTR1 have been suggested to be involved in the physiological response to hypoxia [
62]. AT
1R is broadly expressed in different tissues and mediates most of the classical actions of angiotensin II, including vasoconstriction and vascular smooth muscle cell proliferation [
63]. Thus, under hypoxic conditions, angiotensin II engages AT
1R to modulate the pulmonary vasoconstrictive response [
64]. Although speculative, it is possible that the C allele in
AGTR1 rs275652 might exacerbate pulmonary vasoconstriction during exercise owing to a higher activation of AT
1R for a given concentration of angiotensin II [
65]. The last SNP included in the model explaining VO
2peak was rs7570532, a genetic variation in
GDF8 encoding myostatin. This and other SNPs in
GDF8 have been indirectly associated with a major cardiometabolic condition, obesity [
39], but other authors have reported no association of rs7570532 with endurance performance [
40]. Myostatin controls the differentiation and proliferation of skeletal muscle throughout embryonic development and regulates muscle growth during adulthood. Mutations in
GDF8 that produce non-functional myostatin result in the increased growth of skeletal muscle, demonstrating the existence of a powerful mechanism to control muscle size in normal individuals through this protein [
66]. Based on these findings and given the positive association between muscle mass and VO
2peak [
66,
67,
68], it is possible that
GDF8 rs7570532 confers a small but significant predisposition to higher VO
2peak values. Further research on these three SNPs is clearly warranted.
We acknowledge that the current investigation has some limitations. First, our study sample was heterogeneous in terms of age, sex, and anthropometric characteristics (
Table 1). Although we used these variables as covariates in linear regression analyses, the high variability of these variables might have partially influenced our results. In fact, when they were included in multiple regression analyses (
Figure 2b), the explained variance of VO
2peak increased up to 50.1%. Second, our study only included participants of Han Chinese descent and the results might therefore not be applicable to other ethnicities. Of note, the Han Chinese constitute the world’s largest ethnic group (constituting ~18% of the global population), but further studies in other large ethnic groups will be needed to confirm/discard the generalizability of these results. Lastly, we only analyzed 125 SNPs and thus it is plausible that other candidate genes might have an influence on VO
2peak.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}



