Color Intensity of the Red-Fleshed Berry Phenotype of Vitis vinifera Teinturier Grapes Varies Due to a 408 bp Duplication in the Promoter of VvmybA1

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and SSR Marker Analysis
2.2. DNA Preparation, PCR Analysis and Sequencing of VvmybA1 Alleles
2.3. Extraction and Analysis of Anthocyanins
2.4. RNA Isolation, cDNA Synthesis and Quantitative Real-Time PCR (qRT-PCR)
2.5. Sequence Analysis and Statistics
3. Results
3.1. ‘Teinturier’ is a Periclinal Chimera and Related to ‘Pinot Noir’ and ‘Savagnin Blanc’
3.2. A Multiple Duplication of a 408 bp Promoter Fragment Is Associated with the Red-Fleshed Berry Phenotype
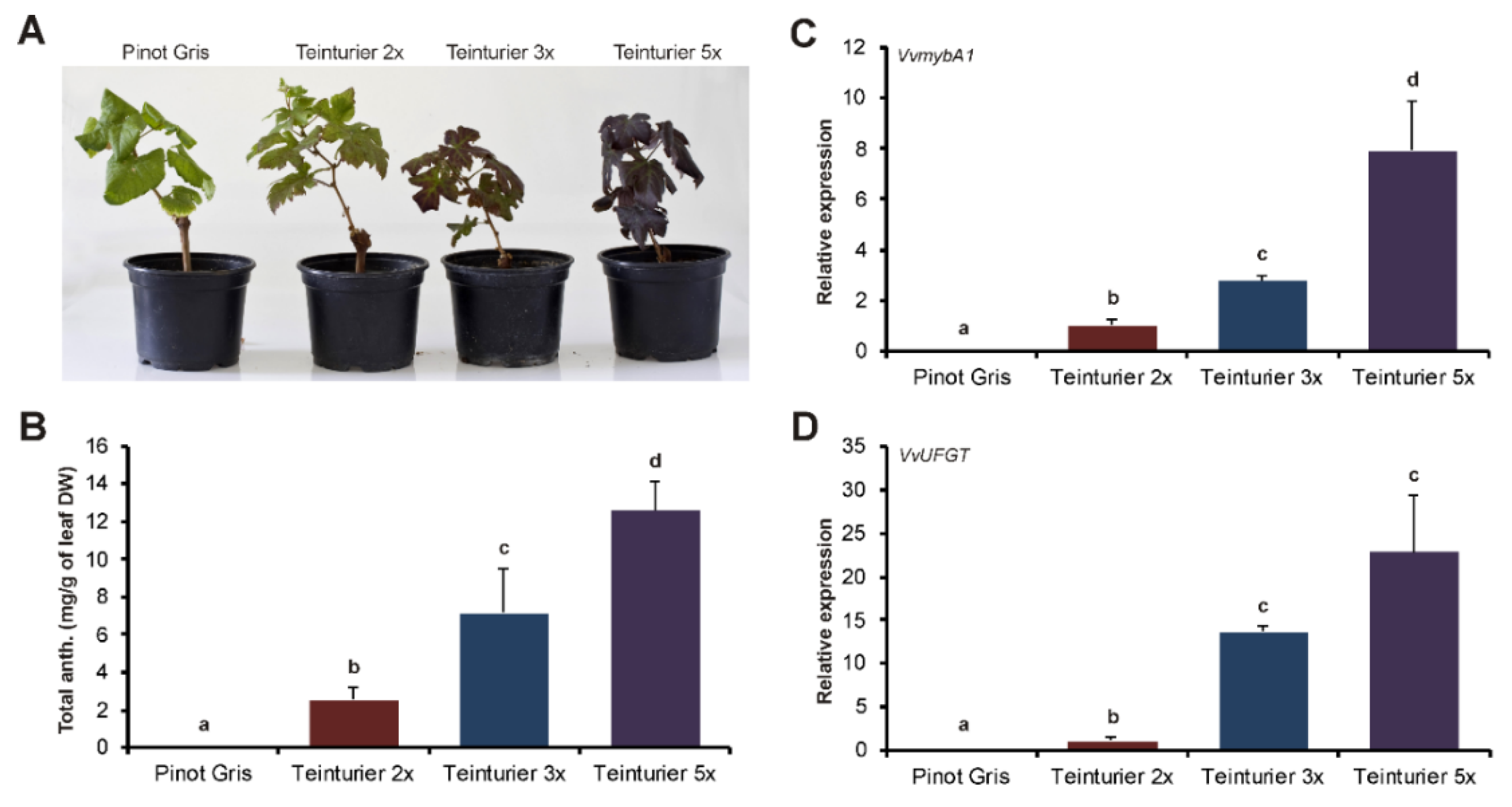
3.3. The Anthocyanin Concentration in Leaves Is Related to the GCE Number
3.4. Ratio of 3′- and 3′,5′-Substituted Anthocyanins in Leaves Differ Among ‘Teinturier’ Clones
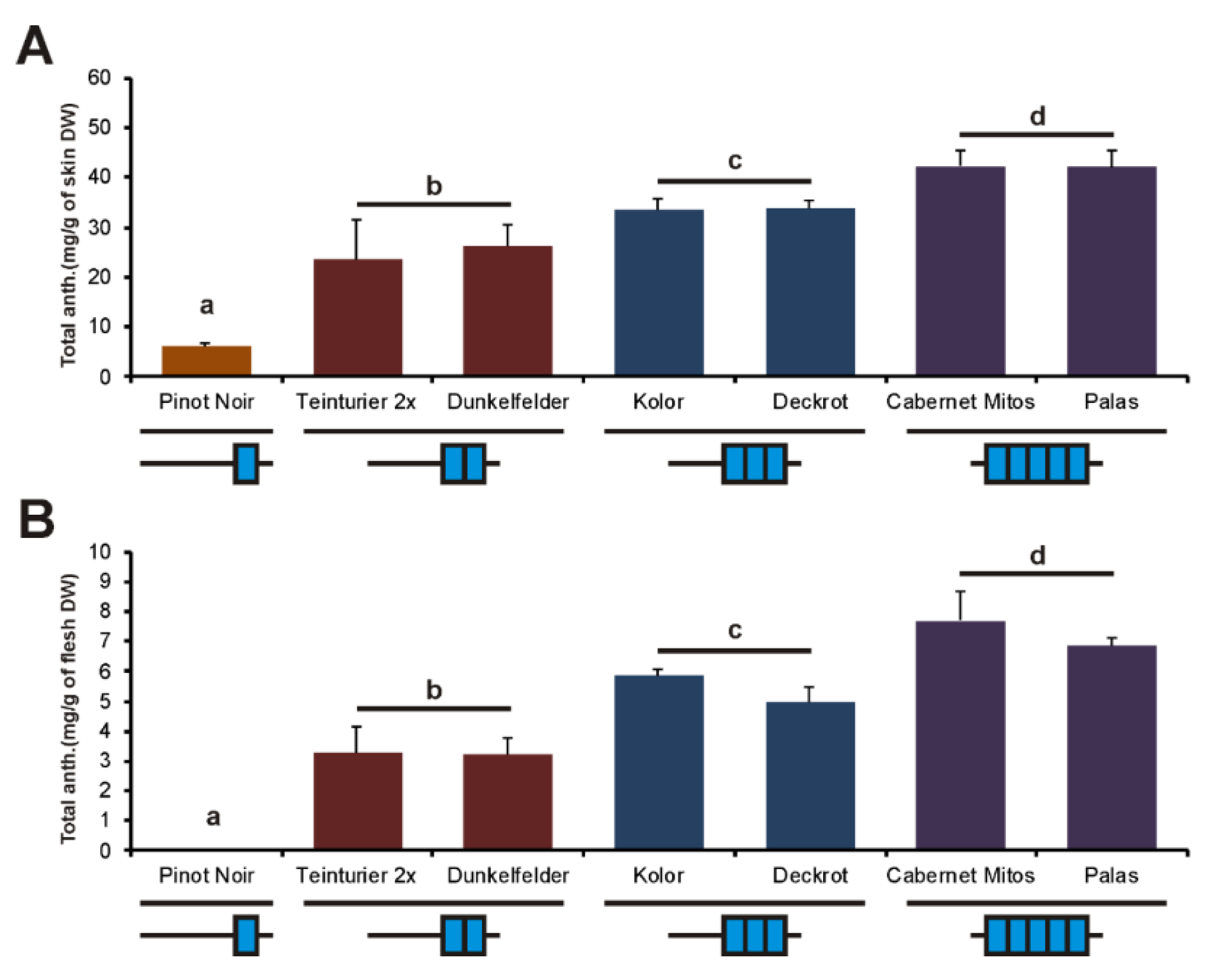
3.5. The Influence of the GCE Number on Berry Anthocyanins of the Progeny of ‘Teinturier’
4. Discussion
4.1. The Mutation of ‘Teinturier’ and Its Origin
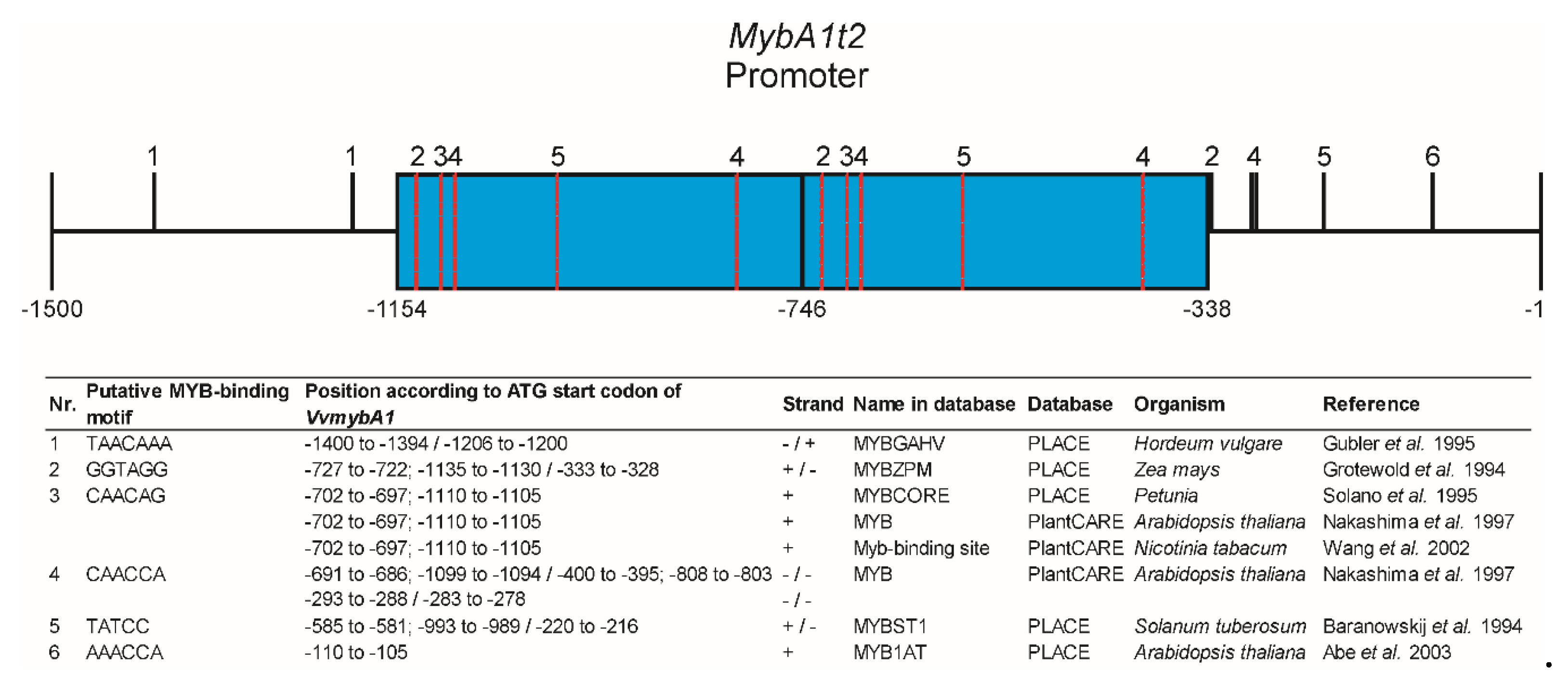
4.2. The Number of GCE Repeats in the Promoter of the MybA1t Alleles Affects the Spatiotemporal Anthocyanin Formation through Altered Regulation of MybA1-Dependent Genes
4.3. Somatic Mutations and Periclinal Chimeras Are Important Drivers for Increasing Grapevine Diversity
5. Conclusions
Supplementary Materials
Data Deposition
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kühnau, J. The flavonoids. A class of semi-essential food components: Their role in human nutrition. World Rev. Nutr. Diet. 1976, 24, 117–191. [Google Scholar]
- Macheix, J.-J.; Fleuriet, A. Fruit Phenolics; CRC Press: Boca Raton, FL, USA, 1990. [Google Scholar]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Fournier-Level, A.; Le Cunff, L.; Gomez, C.; Doligez, A.; Ageorges, A.; Roux, C.; Bertrand, Y.; Souquet, J.M.; Cheynier, V.; This, P. Quantitative Genetic Bases of Anthocyanin Variation in Grape (Vitis vinifera L. ssp sativa) Berry: A Quantitative Trait Locus to Quantitative Trait Nucleotide Integrated Study. Genetics 2009, 183, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Mu, L.; Yan, G.L.; Liang, N.N.; Pan, Q.H.; Wang, J.; Reeves, M.J.; Duan, C.Q. Biosynthesis of anthocyanins and their regulation in colored grapes. Molecules 2010, 15, 9057–9091. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Race, E.J.; Shrikhande, A.J. Characterization of anthocyanins in grape juices by ion trap liquid chromatography—Mass spectrometry. J. Agric. Food Chem. 2003, 51, 1839–1844. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C.; Robinson, S.P. Anthocyanin composition and anthocyanin pathway gene expression in grapevine sports differing in berry skin colour. Aust. J. Grape Wine Res. 1996, 2, 163–170. [Google Scholar] [CrossRef]
- Czemmel, S.; Heppel, S.C.; Bogs, J. R2R3 MYB transcription factors: Key regulators of the flavonoid biosynthetic pathway in grapevine. Protoplasma 2012, 249, 109–118. [Google Scholar] [CrossRef]
- Mu, L.; He, J.J.; Pan, Q.H.; He, F.; Duan, C.Q. Tissue-specific Accumulation of Flavonoids in Grape Berries is Related to Transcriptional Expression of VvF3’H and VvF3’5’H. S. Afr. J. Enol. Vitic. 2014, 35, 68–81. [Google Scholar] [CrossRef] [Green Version]
- Azuma, A.; Ban, Y.; Sato, A.; Kono, A.; Shiraishi, M.; Yakushiji, H.; Kobayashi, S. MYB diplotypes at the color locus affect the ratios of tri/di-hydroxylated and methylated/non-methylated anthocyanins in grape berry skin. Tree Genet. Genomes 2015, 11, 31. [Google Scholar] [CrossRef]
- Boss, P.K.; Davies, C.; Robinson, S.P. Expression of anthocyanin biosynthesis pathway genes in red and white grapes. Plant Mol. Biol. 1996, 32, 565–569. [Google Scholar] [CrossRef]
- Fournier-Level, A.; Hugueney, P.; Verries, C.; This, P.; Ageorges, A. Genetic mechanisms underlying the methylation level of anthocyanins in grape (Vitis vinifera L.). BMC Plant Biol. 2011, 11, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinaldo, A.R.; Cavallini, E.; Jia, Y.; Moss, S.M.A.; McDavid, D.A.J.; Hooper, L.C.; Robinson, S.P.; Tornielli, G.B.; Zenoni, S.; Ford, C.M.; et al. A Grapevine Anthocyanin Acyltransferase, Transcriptionally Regulated by VvMYBA, Can Produce Most Acylated Anthocyanins Present in Grape Skins. Plant Physiol. 2015, 169, 1897–1916. [Google Scholar] [CrossRef] [PubMed]
- Springob, K.; Nakajima, J.-I.; Yamazaki, M.; Saito, K. Recent advances in the biosynthesis and accumulation of anthocyanins. Nat. Prod. Rep. 2003, 20, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Goto-Yamamoto, N.; Kitayama, M.; Hashizume, K. Loss of anthocyanins in red-wine grape under high temperature. J. Exp. Bot. 2007, 58, 1935–1945. [Google Scholar] [CrossRef]
- Jeong, S.T.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, A. Effects of plant hormones and shading on the accumulation of anthocyanins and the expression of anthocyanin biosynthetic genes in grape berry skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- Castellarin, S.D.; Di Gaspero, G. Transcriptional control of anthocyanin biosynthetic genes in extreme phenotypes for berry pigmentation of naturally occurring grapevines. BMC Plant Biol. 2007, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Peppi, M.C.; Walker, M.A.; Fidelibus, M.W. Application of abscisic acid rapidly upregulated UFGT gene expression and improved color of grape berries. Vitis 2008, 47, 11–14. [Google Scholar]
- Pelsy, F.; Dumas, V.; Bevilacqua, L.; Hocquigny, S.; Merdinoglu, D. Chromosome Replacement and Deletion Lead to Clonal Polymorphism of Berry Color in Grapevine. PLoS Genet. 2015, 11, e1005081. [Google Scholar] [CrossRef]
- Azuma, A.; Kobayashi, S.; Mitani, N.; Shiraishi, M.; Yamada, M.; Ueno, T.; Kono, A.; Yakushiji, H.; Koshita, Y. Genomic and genetic analysis of Myb-related genes that regulate anthocyanin biosynthesis in grape berry skin. Theor. Appl. Genet. 2008, 117, 1009–1019. [Google Scholar] [CrossRef]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.A.; Thomas, M.R.; Robinson, S.P. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. Cell Mol. Biol. 2007, 49, 772–785. [Google Scholar] [CrossRef]
- Azuma, A.; Kobayashi, S.; Goto-Yamamoto, N.; Shiraishi, M.; Mitani, N.; Yakushiji, H.; Koshita, Y. Color recovery in berries of grape (Vitis vinifera L.) ‘Benitaka’, a bud sport of ‘Italia’, is caused by a novel allele at the VvmybA1 locus. Plant Sci. 2009, 176, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Azuma, A.; Kobayashi, S.; Yakushiji, H.; Yamada, M.; Mitani, N.; Sato, A. VvmybA1 genotype determines grape skin color. Vitis 2007, 46, 154–155. [Google Scholar]
- Falginella, L.; Di Gaspero, G.; Castellarin, S.D. Expression of flavonoid genes in the red grape berry of ‘Alicante Bouschet’ varies with the histological distribution of anthocyanins and their chemical composition. Planta 2012, 236, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- He, J.J.; Liu, Y.X.; Pan, Q.H.; Cui, X.Y.; Duan, C.Q. Different Anthocyanin Profiles of the Skin and the Pulp of Yan73 (Muscat Hamburg x Alicante Bouschet) Grape Berries. Molecules 2010, 15, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Roy-Chevrier, J. Ampélographie Rétrospective; Coulet et Fils: Paris, France; Masson et Cie: Montpellier, France, 1900; p. 562. [Google Scholar]
- Galet, P. Encyplopedic Dictionary of Grapevine Cultivars; Hachette Livre: Paris, France, 2000. [Google Scholar]
- Anderson, K.; Aryal, N.R. Which Winegrape Varieties are Grown where? A Global Empirical Picture; University of Adelaide Press: Adelaide, Australia, 2015. [Google Scholar]
- California Department of Food and Agriculture. California Grape Crush Report Final 2019; California Department of Food and Agriculture: Sacramento, CA, USA, 10 March 2020; p. 159.
- Pelsy, F. Molecular and cellular mechanisms of diversity within grapevine varieties. Heredity 2010, 104, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Liu, Z.; Guan, L.; Zheng, T.; Jiu, S.; Zhu, X.; Jia, H.; Fang, J. Changes of Anthocyanin Component Biosynthesis in ‘Summer Black’ Grape Berries after the Red Flesh Mutation Occurred. J. Agric. Food Chem. 2018, 66, 9209–9218. [Google Scholar] [CrossRef]
- Guan, L.; Dai, Z.W.; Wu, B.H.; Wu, J.; Merlin, I.; Hilbert, G.; Renaud, C.; Gomes, E.; Edwards, E.; Li, S.H.; et al. Anthocyanin biosynthesis is differentially regulated by light in the skin and flesh of white-fleshed and teinturier grape berries. Planta 2016, 243, 23–41. [Google Scholar] [CrossRef]
- Vezzulli, S.; Leonardelli, L.; Malossini, U.; Stefanini, M.; Velasco, R.; Moser, C. Pinot blanc and Pinot gris arose as independent somatic mutations of Pinot noir. J. Exp. Bot. 2012, 63, 6359–6369. [Google Scholar] [CrossRef] [Green Version]
- Walker, A.R.; Lee, E.; Robinson, S.P. Two new grape cultivars, bud sports of Cabernet Sauvignon bearing pale-coloured berries, are the result of deletion of two regulatory genes of the berry colour locus. Plant Mol. Biol. 2006, 62, 623–635. [Google Scholar] [CrossRef]
- Hocquigny, S.; Pelsy, F.; Dumas, V.; Kindt, S.; Heloir, M.C.; Merdinoglu, D. Diversification within grapevine cultivars goes through chimeric states. Genome 2004, 47, 579–589. [Google Scholar] [CrossRef]
- Maul, E.; Sudharma, K.N.; Kecke, S.; Marx, G.; Muller, C.; Audegun, L.; Boselli, M.; Boursiquot, J.M.; Bucchetti, B.; Cabello, F.; et al. The European Vitis Database (www.eu-vitis.de)—A technical innovation through an online uploading and interactive modification system. Vitis 2012, 51, 145. [Google Scholar]
- Huber, F.; Rockel, F.; Schwander, F.; Maul, E.; Eibach, R.; Cousins, P.; Topfer, R. A view into American grapevine history: Vitis vinifera cv. ‘Semillon’ is an ancestor of ‘Catawba’ and ‘Concord’. Vitis 2016, 55, 53–56. [Google Scholar] [CrossRef]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- Rustioni, L.; Maghradze, D.; Popescu, C.F.; Cola, G.; Abashidze, E.; Aroutiounian, R.; Brazao, J.; Coletti, S.; Cornea, V.; Dejeu, L.; et al. First results of the European grapevine collections’ collaborative network: Validation of a standard eno-carpological phenotyping method. Vitis 2014, 53, 219–226. [Google Scholar]
- OIV. HPLC-Determination of nine major anthocyanins in red and rosé wine (Resolution Oeno 22/2003 modified by Oeno 12/2007). In Compendium of International Methods of Analysis-OIV Anthocyanins; OIV-MA-AS315-11; OIV: Paris, France, 2007; pp. 1–13. [Google Scholar]
- Chandra, A.; Rana, J.; Li, Y.Q. Separation, identification, quantification, and method validation of anthocyanins in botanical supplement raw materials by HPLC and HPLC-MS. J. Agric. Food Chem. 2001, 49, 3515–3521. [Google Scholar] [CrossRef]
- De Rosso, M.; Tonidandel, L.; Larcher, R.; Nicolini, G.; Ruggeri, V.; Dalla Vedova, A.; De Marchi, F.; Gardiman, M.; Flamini, R. Study of anthocyanic profiles of twenty-one hybrid grape varieties by liquid chromatography and precursor-ion mass spectrometry. Anal. Chim. Acta 2012, 732, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Kammerer, D.; Kljusuric, J.G.; Carle, R.; Schieber, A. Recovery of anthocyanins from grape pomace extracts (Vitis vinifera L. cv. Cabernet Mitos) using a polymeric adsorber resin. Eur. Food Res. Technol. 2005, 220, 431–437. [Google Scholar] [CrossRef]
- Iandolino, A.B.; Da Silva, F.G.; Lim, H.; Choi, H.; Williams, L.E.; Cook, D.R. High-quality RNA, cDNA, and derived EST libraries from grapevine (Vitis vinifera L.). Plant Mol. Biol. Rep. 2004, 22, 269–278. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Reid, K.E.; Olsson, N.; Schlosser, J.; Peng, F.; Lund, S.T. An optimized grapevine RNA isolation procedure and statistical determination of reference genes for real-time RT-PCR during berry development. BMC Plant Biol. 2006, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Howard, S.; Chen, S.; Wang, Y.; Yu, O.; Kovacs, L.G.; Qiu, W. Berry skin development in Norton grape: Distinct patterns of transcriptional regulation and flavonoid biosynthesis. BMC Plant Biol. 2011, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canaguier, A.; Grimplet, J.; Di Gaspero, G.; Scalabrin, S.; Duchêne, E.; Choisne, N.; Mohellibi, N.; Guichard, C.; Rombauts, S.; Le Clainche, I.; et al. A new version of the grapevine reference genome assembly (12X.v2) and of its annotation (VCost.v3). Genom. Data 2017, 14, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–465. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van De Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. ML-RELATE: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes 2006, 6, 576–579. [Google Scholar] [CrossRef]
- Lacombe, T.; Boursiquot, J.M.; Laucou, V.; Di Vecchi-Staraz, M.; Peros, J.P.; This, P. Large-scale parentage analysis in an extended set of grapevine cultivars (Vitis vinifera L.). Theor. Appl. Genet. 2013, 126, 401–414. [Google Scholar] [CrossRef]
- Venables, W.N.; Smith, D.M.; The R Core Team. An Introduction to R. Notes on R: A Programming Environment for Data Analysis and Graphics. Available online: https://onlinebooks.library.upenn.edu/webbin/book/lookupid?key=olbp44950 (accessed on 29 November 2018).
- Jeong, S.T.; Goto-Yamamoto, N.; Hashizume, K.; Kobayashi, S.; Esaka, M. Expression of VvmybA1 gene and anthocyanin accumulation in various grape organs. Am. J. Enol. Vitic. 2006, 57, 507–510. [Google Scholar]
- Yakushiji, H.; Kobayashi, S.; Goto-Yamamoto, N.; Tae Jeong, S.; Sueta, T.; Mitani, N.; Azuma, A. A skin color mutation of grapevine, from black-skinned Pinot Noir to white-skinned Pinot Blanc, is caused by deletion of the functional VvmybA1 allele. Biosci. Biotechnol. Biochem. 2006, 70, 1506–1508. [Google Scholar] [CrossRef] [Green Version]
- Franks, T.; Botta, R.; Thomas, M.R. Chimerism in grapevines: Implications for cultivar identity, ancestry and genetic improvement. Theor. Appl. Genet. 2002, 104, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Migliaro, D.; Crespan, M.; Munoz-Organero, G.; Velasco, R.; Moser, C.; Vezzulli, S. Structural dynamics at the berry colour locus in Vitis vinifera L. somatic variants. Aust. J. Grape Wine Res. 2014, 20, 485–495. [Google Scholar] [CrossRef]
- Porret, N.A. Realtime PCR Systems to Monitor Yeasts in Grape Must and Wine. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, 2006. [Google Scholar]
- Guan, L.; Li, J.H.; Fan, P.G.; Li, S.H.; Fang, J.B.; Dai, Z.W.; Delrot, S.; Wang, L.J.; Wu, B.H. Regulation of Anthocyanin Biosynthesis in Tissues of a Teinturier Grape Cultivar under Sunlight Exclusion. Am. J. Enol. Vitic. 2014, 65, 363–374. [Google Scholar] [CrossRef]
- Fournier-Level, A.; Lacombe, T.; Le Cunff, L.; Boursiquot, J.M.; This, P. Evolution of the VvMybA gene family, the major determinant of berry colour in cultivated grapevine (Vitis vinifera L.). Heredity 2010, 104, 351–362. [Google Scholar] [CrossRef] [Green Version]
- Padeken, J.; Zeller, P.; Gasser, S.M. Repeat DNA in genome organization and stability. Curr. Opin. Genet. Dev. 2015, 31, 12–19. [Google Scholar] [CrossRef]
- Puchta, H. The repair of double-strand breaks in plants: Mechanisms and consequences for genome evolution. J. Exp. Bot. 2005, 56, 1–14. [Google Scholar] [CrossRef] [Green Version]
- KalDZXia, R.K.; Rai, M.K.; Kalia, S.; Singh, R.; Dhawan, A.K. Microsatellite markers: An overview of the recent progress in plants. Euphytica 2011, 177, 309–334. [Google Scholar] [CrossRef]
- Vaughn, J.N.; Bennetzen, J.L. Natural insertions in rice commonly form tandem duplications indicative of patch-mediated double-strand break induction and repair. Proc. Natl. Acad. Sci. USA 2014, 111, 6684–6689. [Google Scholar] [CrossRef] [Green Version]
- Schiml, S.; Fauser, F.; Puchta, H. Repair of adjacent single-strand breaks is often accompanied by the formation of tandem sequence duplications in plant genomes. Proc. Natl. Acad. Sci. USA 2016, 113, 7266–7271. [Google Scholar] [CrossRef] [Green Version]
- Chagne, D.; Kui, L.W.; Espley, R.V.; Volz, R.K.; How, N.M.; Rouse, S.; Brendolise, C.; Carlisle, C.M.; Kumar, S.; De Silva, N.; et al. An Ancient Duplication of Apple MYB Transcription Factors Is Responsible for Novel Red Fruit-Flesh Phenotypes. Plant Physiol. 2013, 161, 225–239. [Google Scholar] [CrossRef] [Green Version]
- El-Sharkawy, I.; Liang, D.; Xu, K.N. Transcriptome analysis of an apple (Malus x domestica) yellow fruit somatic mutation identifies a gene network module highly associated with anthocyanin and epigenetic regulation. J. Exp. Bot. 2015, 66, 7359–7376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butelli, E.; Licciardello, C.; Zhang, Y.; Liu, J.J.; Mackay, S.; Bailey, P.; Reforgiato-Recupero, G.; Martin, C. Retrotransposons Control Fruit-Specific, Cold-Dependent Accumulation of Anthocyanins in Blood Oranges. Plant Cell 2012, 24, 1242–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, X.; Song, W.; Bao, Y.; Jin, Y.; Jiang, C.; Wang, C.; Li, B.; Zhang, H. PdMYB118, isolated from a red leaf mutant of Populus deltoids, is a new transcription factor regulating anthocyanin biosynthesis in poplar. Plant Cell Rep. 2019, 38, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Zhu, L.; Zhao, C.; Li, K.; Shang, C.; Hou, L.; Wang, M.; Shi, J.; Fan, S.; Wang, X. Comparative transcriptome analysis of anthocyanin synthesis in black and pink peanut. Plant Signal. Behav. 2020, 15, 1721044. [Google Scholar] [CrossRef]
- Chiu, L.W.; Zhou, X.; Burke, S.; Wu, X.; Prior, R.L.; Li, L. The purple cauliflower arises from activation of a MYB transcription factor. Plant Physiol. 2010, 154, 1470–1480. [Google Scholar] [CrossRef] [Green Version]
- Espley, R.V.; Brendolise, C.; Chagne, D.; Kutty-Amma, S.; Green, S.; Volz, R.; Putterill, J.; Schouten, H.J.; Gardiner, S.E.; Hellens, R.P.; et al. Multiple Repeats of a Promoter Segment Causes Transcription Factor Autoregulation in Red Apples. Plant Cell 2009, 21, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Grotewold, E.; Drummond, B.J.; Bowen, B.; Peterson, T. The Myb-Homologous P-Gene Controls Phlobaphene Pigmentation in Maize Floral Organs by Directly Activating a Flavonoid Biosynthetic Gene Subset. Cell 1994, 76, 543–553. [Google Scholar] [CrossRef]
- Solano, R.; Nieto, C.; Avila, J.; Canas, L.; Diaz, I.; Pazares, J. Dual DNA-Binding Specificity of a Petal Epidermis-Specific Myb Transcription Factor (Myb.Ph3) from Petunia-Hybrida. EMBO J. 1995, 14, 1773–1784. [Google Scholar] [CrossRef]
- Baranowskij, N.; Frohberg, C.; Prat, S.; Willmitzer, L. A Novel DNA-Binding Protein with Homology to Myb Oncoproteins Containing Only One Repeat Can Function as a Transcriptional Activator. EMBO J. 1994, 13, 5383–5392. [Google Scholar] [CrossRef]
- Nakashima, K.; Kiyosue, T.; YamaguchiShinozaki, K.; Shinozaki, K. A nuclear gene, erd1 encoding a chloroplast-targeted Clp protease regulatory subunit homolog is not only induced by water stress but also developmentally up-regulated during senescence in Arabidopsis thaliana. Plant J. 1997, 12, 851–861. [Google Scholar] [CrossRef]
- Xie, S.; Qao, X.L.; Chen, H.W.; Nan, H.; Zhang, Z.W. Coordinated Regulation of Grape Berry Flesh Color by Transcriptional Activators and Repressors. J. Agric. Food Chem. 2019, 67, 11815–11824. [Google Scholar] [CrossRef] [PubMed]
- Carbonell-Bejerano, P.; Royo, C.; Mauri, N.; Ibáñez, J.; Martínez Zapater, J.M. Somatic Variation and Cultivar Innovation in Grapevine. In Advances in Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2019; p. 22. [Google Scholar] [CrossRef] [Green Version]
- Marcotrigiano, M.; Bernatzky, R. Arrangement of Cell-Layers in the Shoot Apical Meristems of Periclinal Chimeras Influences Cell Fate. Plant J. 1995, 7, 193–202. [Google Scholar] [CrossRef]
- Ramos-Madrigal, J.; Runge, A.K.W.; Bouby, L.; Lacombe, T.; Castruita, J.A.S.; Adam-Blondon, A.F.; Figueiral, I.; Hallavant, C.; Martinez-Zapater, J.M.; Schaal, C.; et al. Palaeogenomic insights into the origins of French grapevine diversity. Nat. Plants 2019, 5, 595–603. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prime Name (Official Variety Name) | Accession Name (Name in Repository) | Accession No./Origin | MybA1a White | MybA1c Red | MybA1t Repeat Number |
|---|---|---|---|---|---|
| Pinot Noir | Spaetburgunder M242 | DEU098-1980-006 | x | x | - |
| Pinot Gris | Pinot Gris Cl. Dunkelgrau | DEU098-1980-058 | x | x | - |
| Pinot Teinturier | Pinot Teinturier | DEU454-L-28-4-2 | x | x | - |
| Gamay Teinturier de Bouze | Gamay Teinturier de Bouze | DEU098-1980-196 | x | x | - |
| Teinturier | Teinturier | DEU098-1993-213 | x | x | 2 |
| Teinturier | Farbtraube | DEU098-2002-003 | x | x | 2 |
| Teinturier | Farbtraube | DEU098-2011-063 | x | x | 2 |
| Teinturier | Farbtraube | DEU363-282 | x | x | 2 |
| Teinturier | Färbertraube | DEU454-L-9-4-2 | x | x | 2 |
| Teinturier | Teinturier mâle | DEU456-1068 | x | x | 2 |
| Teinturier | Färbertraube | DEU616 | x | x | 2 |
| Teinturier | Farbtraube (Fröhlich) | DEU616 | x | x | 2 |
| Teinturier | Plant rouge femelle | FRA139-303Mtp2 | x | x | 2 |
| Teinturier | Serzial | FRA139-303Mtp6 | x | x | 2 |
| Teinturier | Teinturier | DEU616 | x | x | 3 |
| Teinturier | Plant rouge mâle | FRA139-303Mtp1 | x | x | 3 |
| Teinturier | Teinturier du Cher PVM | FRA139-0Mtp1749 | x | x | 3 |
| Teinturier | Rubintraube | DEU456-880 | x | x | 5 |
| Teinturier | Teinturier du Cher | FRA139-303Mtp7 | x | x | 5 |
| Dunkelfelder | Dunkelfelder | DEU098-1980-014 | x | - | 2 |
| Rubired | Rubired | DEU098-1991-123 | x | - | 2 |
| Titan | Titan | DEU098-1980-459 | x | - | 2 |
| Teinturier Luebeck | Teinturier male | DEU098-1995-052 | x | - | 2 |
| Seibel 5437 | Seibel 5437 | DEU098-2003-093 | x | - | 2 |
| Golubok | Golubok | DEU098-1988-077 | x | - | 2 |
| Farbfraenkisch | Farbfraenkisch | DEU454-L-27-5-2 | x | - | 2 |
| Alicante Henri Bouschet | Alicante Henri Bouschet | DEU098-1990-007 | - | x | 2 |
| Bouschet Petit | Bouschet Petit | DEU098-2001-016 | - | x | 2 |
| Karmin | Karmin | DEU098-1991-143 | - | x | 2 |
| Biborkadarka | Biborkadarka | DEU098-1991-136 | - | x | 2 |
| Kurucver | Kurucver | DEU098-1991-149 | - | x | 2 |
| Royalty | Royalty | DEU098-1991-119 | - | x | 2 |
| Grand Noir | Grand Noir | DEU098-1980-125 | - | x | 2 |
| Karmrahyut | Karmrahyut | DEU098-1988-079 | - | x | 2 |
| Morrastel Bouschet | Morrastel Bouschet | DEU098-1980-268 | - | x | 2 |
| Kolor | Kolor | DEU098-1980-033 | x | - | 3 |
| Deckrot | Deckrot | DEU098-1980-010 | x | - | 3 |
| Freiburg 54–64 | Freiburg 54–64 | DEU098-1980-671 | x | - | 3 |
| Teinturier femelle (n. i. 1) | Teinturier femelle | DEU456-1127 | x | - | 3 |
| Accent | Accent | DEU098-2013-031 | x | - | 3 |
| Dakapo | Dakapo | DEU098-2011-046 | - | x | 3 |
| Cabernet Mitos | Cabernet Mitos | DEU098-2010-046 | x | - | 5 |
| Palas | Palas | DEU098-2010-049 | x | - | 5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Röckel, F.; Moock, C.; Braun, U.; Schwander, F.; Cousins, P.; Maul, E.; Töpfer, R.; Hausmann, L. Color Intensity of the Red-Fleshed Berry Phenotype of Vitis vinifera Teinturier Grapes Varies Due to a 408 bp Duplication in the Promoter of VvmybA1. Genes 2020, 11, 891. https://doi.org/10.3390/genes11080891
Röckel F, Moock C, Braun U, Schwander F, Cousins P, Maul E, Töpfer R, Hausmann L. Color Intensity of the Red-Fleshed Berry Phenotype of Vitis vinifera Teinturier Grapes Varies Due to a 408 bp Duplication in the Promoter of VvmybA1. Genes. 2020; 11(8):891. https://doi.org/10.3390/genes11080891
Chicago/Turabian StyleRöckel, Franco, Carina Moock, Ulrike Braun, Florian Schwander, Peter Cousins, Erika Maul, Reinhard Töpfer, and Ludger Hausmann. 2020. "Color Intensity of the Red-Fleshed Berry Phenotype of Vitis vinifera Teinturier Grapes Varies Due to a 408 bp Duplication in the Promoter of VvmybA1" Genes 11, no. 8: 891. https://doi.org/10.3390/genes11080891