The LDLR, APOB, and PCSK9 Variants of Index Patients with Familial Hypercholesterolemia in Russia

 , , , ,
, , , ,  , , add
Show full author list
, , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Description of the Patients
2.2. Systematic Review
2.3. Molecular Genetic Analysis
2.3.1. Target Sequencing
2.3.2. Whole Genome Sequencing and Bioinformatic Analysis
2.3.3. Clinical Interpretation
2.3.4. Sanger Sequencing
3. Results
3.1. Systematic Literature Review
3.2. Genetic Test Results
3.3. Description of All Variants in Russia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

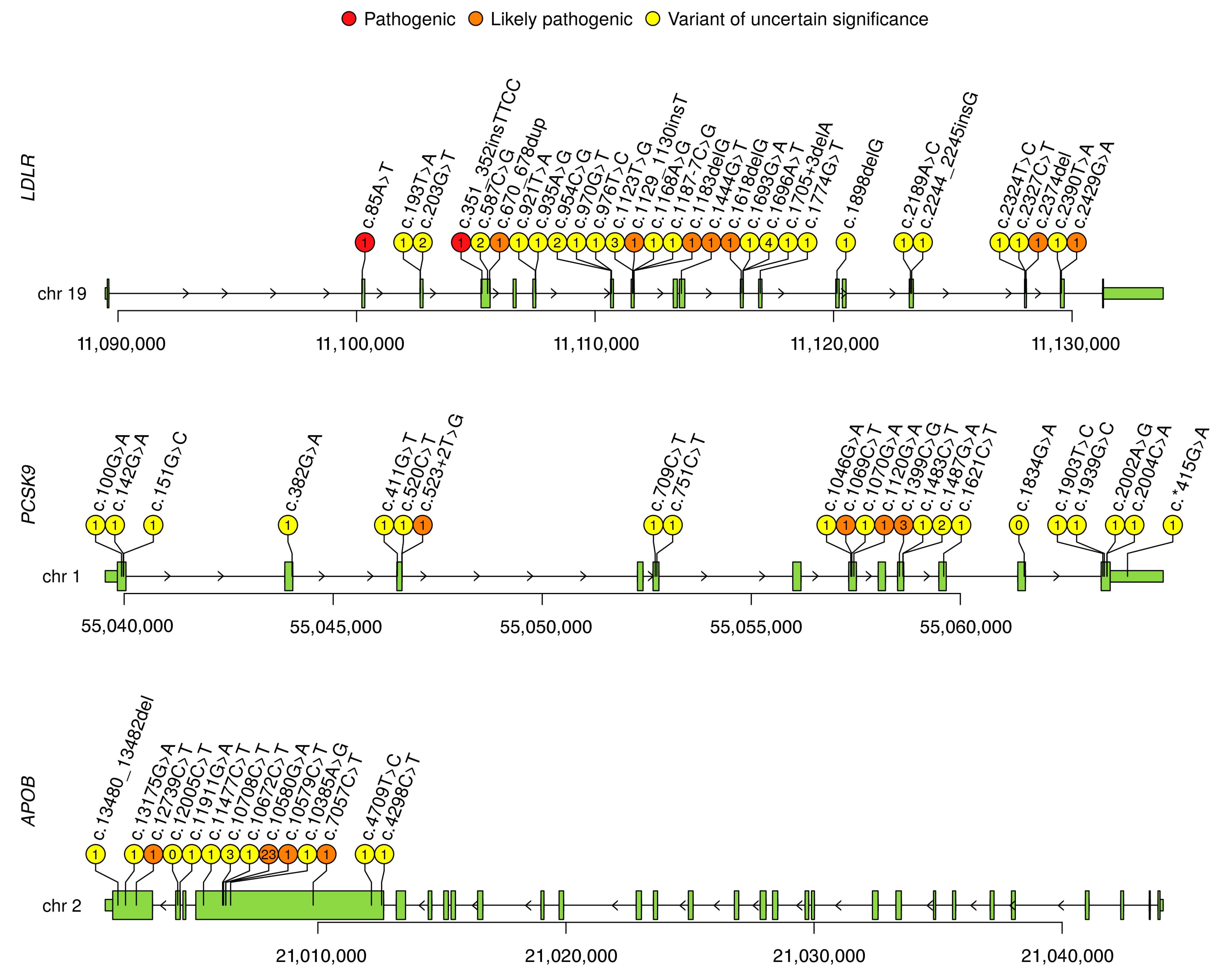{kind=link}
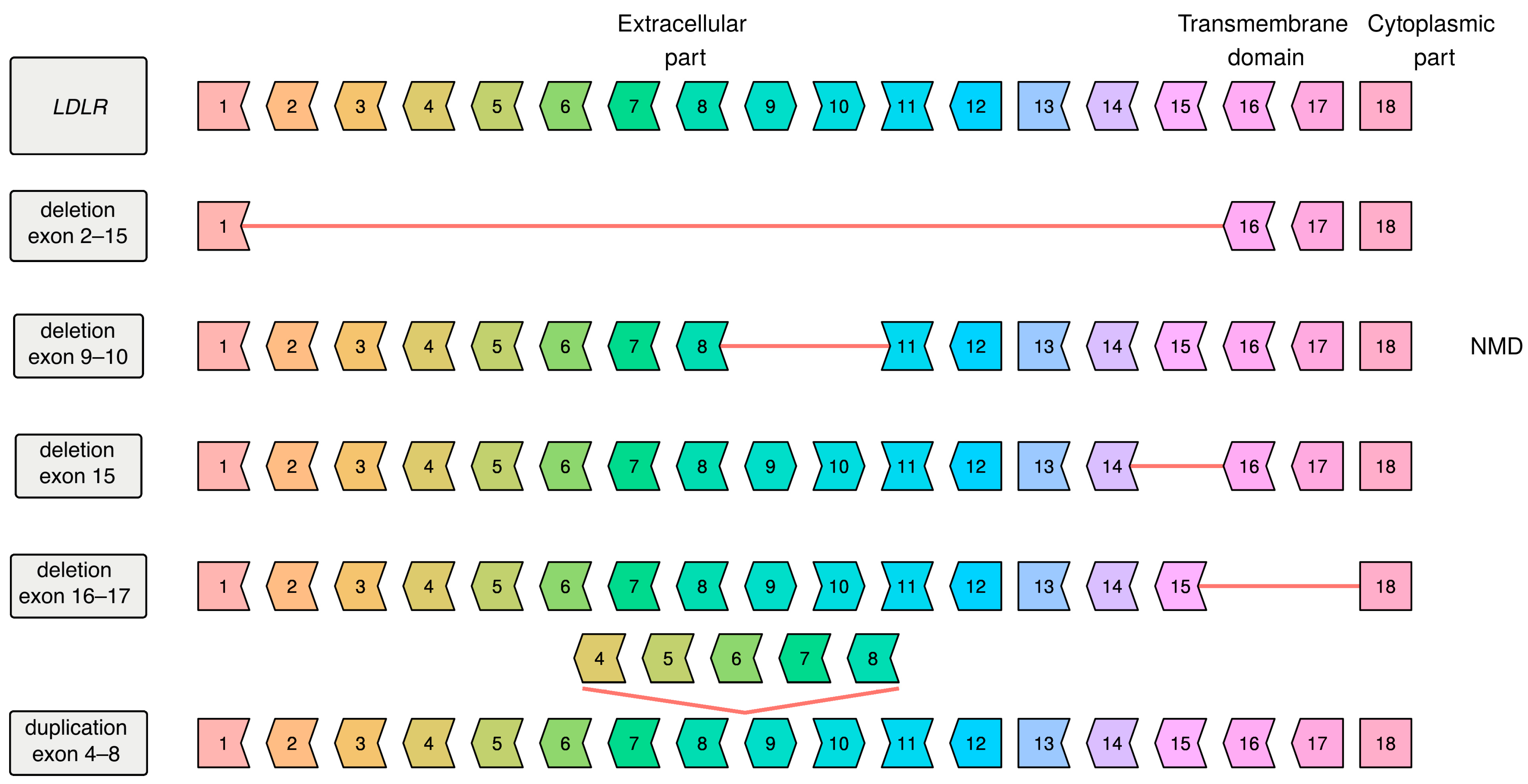{kind=link}
| Number of Index Patients 1 | Variant Data 2 | Exon | DNA Change | Protein Change | dbSNP ID | gnomAD MAF (v. 2.1.1) | ACMG Interpretation | ClinVar Interpretation | ClinVar ID | References |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 1i–15i | c.68-366_2312-791del | P | ||||||
| 0 | 6 | 2 | p.Cys27Trp | rs2228671 | VUS | P/LP | 226304 | [30] | ||
| 1 | 1 | 2 | c.85A > T | p.Arg29Ter | rs879254401 | P | P | 251011 | ||
| 0 | 6 | 2 | c.97C > T | p.Gln33Ter | rs121908024 | 0.000007963 | P | P | 3683 | [6,20,30] |
| 0 | 6 | 3 | c.191_313del | p.Leu64_Pro105delinsSer | P | [9] | ||||
| 0 | 5 | 3 | c.193_202delTCTGTCACCTinsGGACTTCA | p.Ser65Glyfs * 64 | LP | [8,10,25,27,29] | ||||
| 1 | 1 | 3 | c.193T > A | p.Ser65Thr | VUS | |||||
| 0 | 5 | 3 | c.195dupT | p.Val66Cysfs * 64 | rs879254435 | P | P | 251075 | [8,10,25,29] | |
| 2 | 4 | 3 | c.200C > T | p.Thr67Ile | rs1337448484 | 0.00001060 | VUS | VUS | 629411 | |
| 2 | 1 | 3 | c.203G > T | p.Cys68Phe | VUS | |||||
| 2 | 2 | 3 | c.230dup | p.Arg78ProfsTer55 | rs879254440 | P | P | 251083 | [30,37,39] | |
| 1 | 4 | 3 | c.241C > T | p.Arg81Cys | rs730882078 | 0.000007953 | VUS | P/LP/VUS | 183083 | |
| 0 | 6 | 3 | c.245G > C | p.Cys82Ser | VUS | VUS | 431509 | [10] | ||
| 1 | 4 | 3 | c.246C > A | p.Cys82Ter | rs875989891 | P | P | 226309 | ||
| 0 | 6 | 3 | c.285C > A | p.Cys95Ter | rs139400379 | P | P | 251115 | [20,21,30] | |
| 0 | 6 | 3i | c.313 + 1G > A | rs112029328 | 0.00002784 | P | P/LP | 3736 | [6,20] | |
| 0 | 5 | 3i | c.313 + 2T > G | LP | [10] | |||||
| 1 | 3 | 4–8 | c.317-1185dup | p.Pro106_Val395dup | LP | [9] | ||||
| 2 | 4 | 4 | c.326G > A | p.Cys109Tyr | rs121908042 | 0.000003996 | LP | P/LP | 226319 | |
| 2 | 4 | 4 | c.343C > T | p.Arg115Cys | rs774723292 | 0.00002792 | LP | P/LP/VUS | 251162 | |
| 1 | 4 | 4 | c.347G > A | p.Cys116Tyr | LP | |||||
| 0 | 6 | 4 | c.347_349delGCC | p.Cys116_His117delinsTyr | rs879254483 | LP | LP | 251164 | [20,26,30] | |
| 1 | 1 | 4 | c.351_352insTTCC | p.Asp118PhefsTer13 | P | [7] | ||||
| 1 | 5 | 4 | c.355_356insTTCC | p.Gly119ValfsTer12 | P | [9] | ||||
| 1 | 4 | 4 | c.420G > C | p.Glu140Asp | rs879254520 | LP | P/LP | 251216 | ||
| 0 | 5 | 4 | c.444T > G | p.Cys148Trp | rs879254528 | LP | LP | 251228 | [20,24,30] | |
| 0 | 6 | 4 | c.451G > C | p.Ala151Pro | rs763233960 | 0.00001195 | LP | VUS | 251234 | [20,28,30] |
| 6 | 2 | 4 | c.478T > G | p.Cys160Gly | rs879254540 | LP | LP | 251248 | [20,24,29,30] | |
| 0 | 6 | 4 | c.499T > C | p.Cys167Arg | rs879254547 | LP | P/LP | 251255 | [20,28] | |
| 1 | 4 | 4 | c.502G > C | p.Asp168His | rs200727689 | LP | P/LP | 251258 | ||
| 1 | 4 | 4 | c.519C > G | p.Cys173Trp | rs769318035 | 0.000007958 | LP | P/LP | 251277 | |
| 0 | 6 | 4 | c.530C > T | p.Ser177Leu | rs121908026 | 0.00001592 | LP | P/LP | 3686 | [9] |
| 1 | 3 | 4 | c.534T > G | p.Asp178Glu | rs879254566 | LP | P/LP | 251287 | [36] | |
| 0 | 6 | 4 | c.542C > T | p.Pro181Leu | rs557344672 | 0.000007958 | LP | LP | 431512 | [9] |
| 2 | 4 | 4 | c.551G > A | p.Cys184Tyr | rs121908039 | 0.00009554 | LP | P/LP | 3739 | |
| 2 | 1 | 4 | c.587C > G | p.Pro196Arg | VUS | |||||
| 0 | 6 | 4 | c.618T > G | p.Ser206Arg | rs879254595 | LP | P/LP | 251325 | [8,10,25,29] | |
| 5 | 3 | 4 | c.622G > A | p.Glu208Lys | rs879254597 | LP | P/LP | 251328 | [32] | |
| 0 | 6 | 4 | c.626G > A | p.Cys209Tyr | rs879254600 | LP | P/LP | 251332 | [20,28,30] | |
| 5 | 3 | 4 | c.654_656delTGG | p.Gly219del | rs121908027 | P | P/LP | 226329 | [6,20,23,29,30,31] | |
| 1 | 3 | 4 | c.658_663delCCCGAC | p.Pro220_Asp221del | rs1555803409 | LP | P | 440589 | [9] | |
| 0 | 5 | 4–6 | Del 5 kb incl. ex. 4–6 | LP | [38] | |||||
| 3 | 4 | 4 | c.666C > A | p.Cys222Ter | rs756613387 | 0.000004005 | P | P | 251364 | |
| 1 | 4 | 4 | c.672_686delCAAATCTGACGAGGA | p.Asp224_Glu228del | rs1555803439 | LP | LP | 441189 | [7] | |
| 0 | 5 | 4 | c.670_671insG | p.Asp224GlyfsTer4 | rs879254629 | P | P | 251372 | [6,20,30] | |
| 1 | 1 | 4 | c.670_678dup | p.Asp224_Ser226dup | LP | [7] | ||||
| 3 | 4 | 4 | c.682G > A | p.Glu228Lys | rs121908029 | 0.00001614 | LP | P/LP | 3691 | [30,37,39] |
| 0 | 6 | 4 | c.682G > T | p.Glu228Ter | rs121908029 | 0.00001074 | P | P/LP | 226333 | [6,20,30] |
| 1 | 4 | 4 | c.693C > A | p.Cys231Ter | rs121908035 | P | P/LP | 3730 | ||
| 0 | 5 | 5 | p.Glu240Ter * | LP | [30] | |||||
| 0 | 6 | 5 | p.Glu240Lys * | VUS | P/LP/VUS | 200920 | [30] | |||
| 1 | 4 | 5 | c.768C > A | p.Asp256Glu | rs879254671 | VUS | LP | 438322 | ||
| 0 | 6 | 5 | p.Cys261Phe * | VUS | LP | 3740 | [30] | |||
| 0 | 6 | 5 | c.796G > A | p.Asp266Asn | rs875989907 | 0.00001193 | LP | P/LP | 226334 | [34,36] |
| 3 | 4 | 5 | c.798T > A | p.Asp266Glu | rs139043155 | 0.00003535 | VUS | P/LP/VUS | 161287 | |
| 2 | 3 | 5 | c.810C > A | p.Cys270Ter | rs773328511 | P | P | 251465 | [6,20,30] | |
| 1 | 4 | 6 | c.825_826delCT | p.Cys276ArgfsTer24 | rs879254691 | P | P | 251478 | ||
| 2 | 4 | 6 | c.829G > A | p.Glu277Lys | rs148698650 | 0.0005056 | VUS/LB | P/VUS/LB/B | 183097 | |
| 0 | 6 | 6 | p.Glu288Lys | VUS | P/LP/VUS | 161268 | [30] | |||
| 2 | 4 | 6 | c.905G > T | p.Cys302Phe | rs879254715 | LP | P | 430768 | ||
| 0 | 6 | 6 | c.922G > A | p.Glu308Lys | rs879254721 | VUS | LP | 251528 | [9] | |
| 0 | 6 | 6 | c.925_931delCCCATCA | p.Pro309LysfsTer59 | rs387906304 | P | P | 3729 | [6,8,10,20,22,25,29,30] | |
| 1 | 1 | 6 | c.921T > A | p.Asp307Glu | VUS | |||||
| 1 | 1 | 6 | c.935A > G | p.Glu312Gly | rs1380197577 | 0.000003984 | VUS | |||
| 0 | 5 | 6 | c.939_940 + 3delCGGTG | p.Cys313AspfsTer17 | rs879254727 | P | P | 251536 | [6,20,30] | |
| 3 | 3 | 6i | c.940 + 3_940 + 6del | VUS | P/VUS | 869390 | [9] | |||
| 0 | 5 | 5i_6i | c.817 + 303_940 + 943del | p.Val273_Cys313del | VUS | [32] | ||||
| 1 | 4 | 6i | c.941-3C > G | VUS | ||||||
| 1 | 4 | 6i | c.941-2A > G | rs112366278 | P | P/LP | 251554 | |||
| 1 | 4 | 7 | c.949G > A | p.Glu317Lys | rs746834464 | 0.00005311 | VUS | P/LP | 251567 | |
| 2 | 1 | 7 | c.954C > G | p.Cys318Trp | VUS | |||||
| 1 | 1 | 7 | c.970G > T | p.Gly324Cys | VUS | |||||
| 1 | 4 | 7 | c.974G > A | p.Cys325Tyr | rs879254746 | VUS | P/LP | 251580 | ||
| 1 | 1 | 7 | c.976T > C | p.Ser326Pro | VUS | |||||
| 8 | 3 | 7 | c.986G > A | p.Cys329Tyr | rs761954844 | 0.00002479 | VUS | P/LP/LB | 226344 | [6,9,20,30,36] |
| 0 | 6 | 7 | c.1009G > A | p.Glu337Lys | rs539080792 | 0.0000935 | VUS | VUS | 523729 | [34,36] |
| 0 | 6 | 7 | rs755757866 | NA (G > A)\0.000007967 (G > T) | LP (G > A)\NA (G > T) | 251600 (G > A)\NA (G > T) | [34,36] | |||
| 1 | 4 | 7 | c.1027G > A | p.Gly343Ser | rs730882096 | 0.00002832 | VUS | P/LP/VUS | 183106 | |
| 1 | 4 | 7 | c.1048C > T | p.Arg350Ter | rs769737896 | 0.000007977 | P | P | 226342 | |
| 1 | 3 | 7 | c.1054T > C | p.Cys352Arg | rs879254769 | VUS | LP | 251618 | [9,34,36] | |
| 3 | 4 | 7i | c.1061-8T > C | rs72658861 | 0.005498 | VUS/LB | VUS/PB/B | 36451 | ||
| 3 | 1 | 8 | c.1123T > G | p.Tyr375Asp | VUS | |||||
| 1 | 1 | 8 | c.1129_1130insT | p.Cys377LeufsTer1 | LP | |||||
| 1 | 4 | 8 | c.1162del | p.His388ThrfsTer25 | P | P | 226348 | |||
| 1 | 1 | 8 | c.1168A > G | p.Lys390Glu | VUS | |||||
| 1 | 1 | 8 | c.1183delG | p.V395fs | LP | |||||
| 1 | 4 | 8i | c.1186 + 1G > T | rs730880131 | P | P | 180403 | |||
| 1 | 1 | 8i-10i | c.1186 + 568_1586 + 1067del | LP | ||||||
| 3 | 4 | 8i | c.1187-10G > A | rs765696008 | 0.00002798 | VUS | P/LP | 226349 | ||
| 1 | 1 | 8i | c.1187-7C > G | VUS | ||||||
| 28 | 3 | 9 | c.1202T > A | p.Leu401His | rs121908038 | VUS | P/LP | 3735 | [6,20,34,36] | |
| 2 | 4 | 9 | c.1217G > A | p.Arg406Gln | rs552422789 | 0.00001593 | VUS | P/LP/VUS | 228798 | |
| 5 | 3 | 9 | c.1222G > A | p.Glu408Lys | rs137943601 | 0.000007965 | LP | P/LP | 36453 | [10] |
| 0 | 5 | 9 | p.Arg410Gly * | VUS | [30] | |||||
| 0 | 5 | 9 | p.Met412Val * | VUS | [30] | |||||
| 4 | 3 | 9 | c.1246C > T | p.Arg416Trp | rs570942190 | 0.00002389 | LP | P/LP | 183110 | [9,30,34,36] |
| 1 | 3 | 9 | c.1252G > T | p.Glu418Ter | rs869320651 | P | P | 251755 | [20,26,30] | |
| 0 | 5 | 9 | p.Glu418Gly * | VUS | [30] | |||||
| 0 | 6 | 9 | c.1277T > C | p.Leu426Pro | rs879254851 | VUS | P/LB | 251763 | [25] | |
| 1 | 4 | 9 | c.1285G > A | p.Val429Met | rs28942078 | 0.00001194 | LP | P/LP | 3694 | |
| 0 | 5 | 9 | c.1291_1331del41 | p.Ala431Ter | rs879254854 | LP | [6,20,30] | |||
| 1 | 4 | 9 | c.1292C > T | p.Ala431Val | VUS | |||||
| 0 | 6 | 9 | p.Leu432Arg * | VUS | [30] | |||||
| 0 | 5 | 9 | p.Asp433Glu * | rs778309692 | 0.000003980 | VUS | [30] | |||
| 0 | 6 | 9 | p.Asp433His * | VUS | [30] | |||||
| 0 | 6 | 9 | p.Asp433Tyr * | VUS | [30] | |||||
| 0 | 6 | 9 | c.1302delG | p.Glu435MetfsTer15 | P | [6,20,30] | ||||
| 1 | 4 | 9 | c.1322T > A | p.Ile441Asn | rs879254862 | VUS | LP | 251782 | ||
| 2 | 2 | 9 | c.1327T > C | p.Trp443Arg | rs773566855 | 0.000003980 | LP | [9,10,30,33] | ||
| 0 | 6 | 9 | c.1328G > A | p.Trp443Ter | rs879254866 | P | P | 251789 | [6,20] | |
| 0 | 6 | 9 | c.1340C > G | p.Ser447Cys | rs879254870 | VUS | LP | 251797 | [8,10,25] | |
| 0 | 6 | 9i | c.1358 + 1G > A | rs775924858 | P | P/LP | 251802 | [6,20] | ||
| 1 | 1 | 10 | c.1444G > T | p.Asp482Tyr | rs139624145 | LP | LP | 251845 | [30] | |
| 1 | 2 | 10 | c.1465T > A | p.Tyr489Asn | VUS | [9] | ||||
| 1 | 4 | 10 | c.1471A > G | p.Thr491Ala | VUS | |||||
| 1 | 4 | 10 | c.1474G > A | p.Asp492Asn | rs373646964 | 0.00002386 | VUS | P/LP/VUS | 161285 | |
| 1 | 4 | 10 | c.1502C > T | p.Ala501Val | rs755667663 | 0.000007954 | LP | P/LP | 251874 | |
| 0 | 6 | 10 | c.1532T > C | p.Leu511Ser | rs879254932 | VUS | LP | 251887 | [8,25,33] | |
| 1 | 4 | 10 | c.1561G > A | p.Ala521Thr | rs879254940 | VUS | VUS | 251898 | ||
| 1 | 4 | 10 | c.1577C > A | p.Pro526His | rs879254944 | VUS | VUS | 496019 | ||
| 1 | 4 | 10i | c.1586 + 5G > A | rs781362878 | 0.00003189 | VUS | LP/VUS | 251909 | ||
| 1 | 1 | 11 | c.1618delG | p.Ala540ProfsTer8 | LP | |||||
| 1 | 3 | 11 | c.1633G > A | p.Gly545Arg | rs879254965 | LP | P/LP | 251942 | [9] | |
| 0 | 6 | 11 | p.Gly549Asp * | rs28941776 | 0.00002386 | VUS | P/LP | 3698 | [30] | |
| 0 | 5 | 11 | c.1655_1672del | LP | [34,36] | |||||
| 1 | 4 | 11 | c.1672G > T | p.Glu558Ter | rs879254980 | P | P | 251964 | ||
| 0 | 5 | 11 | p.Glu558Lys * | VUS | [30] | |||||
| 0 | 5 | 11 | c.1686_1693delGCCCAATGinsT | p.Trp562CysfsTer5 | rs879254984 | P | P | 251968 | [8,25,33] | |
| 1 | 1 | 11 | c.1693G > A | p.Gly565Ser | rs1344561983 | 0.000003978 | VUS | |||
| 4 | 1 | 11 | c.1696A > T | p.Ile566Phe | VUS | |||||
| 1 | 1 | 11 | c.1705 + 3delA | VUS | ||||||
| 0 | 6 | 11 | p.Leu568Va l * | VUS | [30] | |||||
| 1 | 4 | 12 | c.1706-10G > A | rs17248882 | 0.002220 | VUS/LB | VUS/LB/B | 226368 | ||
| 1 | 4 | 12 | c.1708_1710delCTC | p.Leu571del | rs772492150 | 0.000007953 | VUS | |||
| 1 | 4 | 12 | c.1729T > C | p.Trp577Arg | rs879255000 | LP | P/LP | 252001 | ||
| 0 | 5 | 12 | c.1741A > C | p.Lys581Gln | VUS | [9] | ||||
| 2 | 4 | 12 | c.1747C > T | p.His583Tyr | rs730882109 | 0.0001025 | VUS | P/LP | 200921 | |
| 1 | 4 | 12 | c.1750T > C | p.Ser584Pro | rs879255010 | VUS | LP/VUS | 252015 | ||
| 2 | 2 | 12 | c.1756T > C | p.Ser586Pro | VUS | [9] | ||||
| 1 | 1 | 12 | c.1774G > T | p.Gly592Trp | VUS | |||||
| 29 | 3 | 12 | c.1775G > A | p.Gly592Glu | rs137929307 | 0.00005656 | LP | P/LP | 161271 | [9,20,21,30] |
| 2 | 4 | 12 | c.1784G > A | p.Arg595Gln | rs201102492 | 0.00003889 | VUS | P/LP/VUS | 183126 | |
| 0 | 5 | 12 | p.Leu605Val * | VUS | [30] | |||||
| 0 | 5 | 12 | p.Leu605Arg * | VUS | [30] | |||||
| 0 | 5 | 12 | p.Ala612Gly * | VUS | [30] | |||||
| 1 | 2 | 12i | c.1846-3T > G | VUS | [9] | |||||
| 0 | 5 | 13 | c.1855–1856insA | p.Phe619TyrfsTer26 | rs879255053 | P | P | 252082 | [6,20,30] | |
| 0 | 6 | 13 | c.1859G > C | p.Trp620Ser | VUS | [33] | ||||
| 1 | 3 | 13 | c.1864G > A | p.Asp622Asn | rs879255059 | LP | LP | 252092 | [6,20,30] | |
| 1 | 1 | 13 | c.1898delG | p.Arg633fs | VUS | |||||
| 2 | 4 | 13 | c.1898G > A | p.Arg633His | rs754536745 | 0.00002121 | VUS | P/LP/VUS | 226380 | |
| 0 | 5 | 13 | c.1936C > A | p.Leu646Ile | rs779940524 | 0.000003977 | VUS | LP | 252118 | [8,10,25] |
| 1 | 4 | 13 | c.1945C > T | p.Pro649Ser | rs879255080 | VUS | LP | 252121 | ||
| 3 | 4 | 13 | c.1955T > C | p.Met652Thr | rs875989936 | 0.000003977 | VUS | LP/VUS | 226382 | |
| 1 | 4 | 13 | c.1966C > A | p.His656Asn | rs762815611 | LP | B/LP/VUS | 252136 | ||
| 1 | 4 | 14 | c.1998G > A | p.Trp666Ter | rs752935814 | P | P | 252161 | ||
| 0 | 5 | 14 | c.1999T > A | p.Cys667Ser | rs150021927 | VUS | LP | 252162 | [6,20,30] | |
| 1 | 4 | 14 | c.2001_2002delTG | p.Cys667_Glu668delinsTer | rs1600743301 | LP | P | 630543 | ||
| 1 | 4 | 14 | c.2043C > A | p.Cys681Ter | rs121908031 | 0.000007959 | P | P/LP | 3699 | |
| 3 | 4 | 14 | c.2089G > C | p.Ala697Pro | rs776217028 | VUS | LP | 252213 | ||
| 0 | 6 | 14i–16i | c.2141-966_2390-330del | p.Glu714_Ile796del | LP | [9] | ||||
| 1 | 1 | 14i-15i | c.2141-799_2311 + 689del | LP | ||||||
| 1 | 1 | 15 | c.2189A > C | p.Lys730Thr | VUS | |||||
| 0 | 5 | 15 | c.2191delG | p.Val731SerfsTer6 | rs879255161 | P | P | 252253 | [8,25,29,33] | |
| 0 | 6 | 15 | c.2215C > T | p.Gln739Ter | rs370018159 | P | P/LP | 252258 | [9] | |
| 1 | 4 | 15 | c.2230C > T | p.Arg744Ter | rs200793488 | 0.000003979 | LP | P | 430795 | |
| 0 | 6 | 15 | c.2231G > A | p.Arg744Gln | rs137853963 | 0.0008030 | VUS | LP/VUS/LB/B | 68104 | [20,21] |
| 1 | 1 | 15 | c.2244_2245insG | p.Thr749AspfsTer33 | VUS | |||||
| 1 | 1 | 15i-17i | c.2312-2107_2547 + 620del | LP | ||||||
| 1 | 1 | 16 | c.2324T > C | p.Val775Ala | rs780300776 | 0.00002121 | VUS | LB/VUS | 440691 | |
| 0 | 5 | 16 | c.2326G > T | p.Ala776Ser | VUS | [32] | ||||
| 1 | 1 | 16 | c.2327C > T | p.Ala776Val | VUS | |||||
| 1 | 4 | 16 | c.2347A > C | p.Lys783Gln | rs769318961 | 0.000007954 | VUS | |||
| 1 | 1 | 16 | c.2374del | p.Ile792LeufsTer137 | LP | |||||
| 3 | 3 | 16 | c.2389G > A | p.Val797Met | rs750518671 | 0.000007957 | VUS | P/LP/VUS | 226393 | [6,20,30] |
| 1 | 4 | 16 | c.2389G > C | p.Val797Leu | rs750518671 | VUS | VUS | 565983 | ||
| 1 | 4 | 16 | c.2389 + 2T > G | rs879255188 | P | LP | 252302 | |||
| 4 | 3 | 16i | c.2389 + 5G > C | rs879255191 | VUS | VUS | 661713 | [9] | ||
| 2 | 4 | 16 | c.2389 + 5G > A | rs879255191 | VUS | P/LB | 252306 | |||
| 1 | 1 | 17 | c.2390T > A | p.Val797Glu | VUS | |||||
| 2 | 4 | 17 | c.2416_2417insG | p.Val806fs | P | P/LP | 252330 | |||
| 0 | 6 | 17 | c.2416dupG | p.Val806GlyfsTer11 | rs773618064 | P | P/LP | 252330 | [9] | |
| 1 | 1 | 17 | c.2429G > A | p.Trp810Ter | LP | |||||
| 1 | 4 | 17 | c.2448G > C | p.Lys816Asn | rs1399689294 | 0.00003186 | VUS | LP/VUS | 440698 | |
| 1 | 4 | 17 | c.2473A > G | p.Asn825Asp | rs879255215 | VUS | LP | 252340 | ||
| 1 | 3 | 17 | c.2479G > A | p.Val827Ile | rs137853964 | 0.0009193 | VUS/LB | LP/VUS/LB/B | 36462 | [6,20,34,36] |
| 1 | 4 | 17 | c.2531G > A | p.Gly844Asp | rs121908037 | VUS | LP | 3734 |
| Number of Index Patients 1 | Exon | DNA Change | Protein Change | dbSNP ID | gnomAD MAF (v. 2.1.1) | ACMG Interpretation | ClinVar Interpretation | ClinVar ID | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | c.100G > A | p.Glu34Lys | rs371030381 | 0.00001626 | VUCS | VUS | 536202 | |
| 1 | 1 | c.142G > A | p.Glu48Lys | rs1278890129 | 0.00002190 | VUCS | P/VUS | 440707 | |
| 1 | 1 | c.151G > C | p.Gly51Arg | VUCS | |||||
| 1 | 2 | c.382G > A | p.Gly128Ser | rs766314770 | 0.00003978 | VUCS | |||
| 1 | 3 | c.411G > T | p.Leu137Phe | VUCS | |||||
| 1 | 3 | c.520C > T | p.Pro174Ser | rs533273863 | 0.00007089 | VUCS | VUS | 496561 | |
| 1 | 3 | c.523 + 2T > G | LP | ||||||
| 1 | 5 | c.709C > T | p.Arg237Trp | rs148195424 | 0.0006952 | VUCS | VUS/LB | 265933 | |
| 1 | 5 | c.751C > T | p.Arg251Cys | rs778900671 | 0.00002009 | VUCS | |||
| 1 | 7 | c.1046G > A | p.Gly349Glu | VUCS | |||||
| 1 | 7 | c.1069C > T | p.Arg357Cys | rs148562777 | 0.0001450 | LP | VUS | 575758 | |
| 1 | 7 | c.1070G > A | p.Arg357His | rs370507566 | 0.00003978 | VUCS | VUS | 403288 | |
| 1 | 7 | c.1120G > A | p.Asp374Asn | rs137852912 | 0.00007079 | LP | |||
| 3 | 9 | c.1399C > G | p.Pro467Ala | rs772677312 | 0.00002829 | LP | LP/VUS | 265944 | |
| 1 | 9 | c.1483C > T | p.Arg495Trp | rs758999339 | 0.00001607 | VUCS | |||
| 2 | 9 | c.1487G > A | p.Arg496Gln | rs139669564 | 0.0002363 | VUCS | VUS/LB | 438338 | |
| 1 | 10 | c.1621C > T | p.Pro541Ser | rs369996097 | 0.00001056 | VUCS | |||
| 0 | 11 | c.1834G > A | p.Glu612Lys | VUCS | [32] | ||||
| 1 | 12 | c.1903T > C | p.Cys635Arg | VUCS | |||||
| 1 | 12 | c.1939G > C | p.Ala647Pro | VUCS | |||||
| 1 | 12 | c.2002A > G | p.Ser668Gly | rs775077080 | 0.00004790 | VUCS | VUS | 297707 | |
| 1 | 12 | c.2004C > A | p.Ser668Arg | rs762298323 | 0.00002397 | VUCS | VUS | 403291 | |
| 1 | c.*415G > A | VUCS | [32] |
| Number of Index Patients 1 | Exon | DNA Change | Protein Change | dbSNP ID | gnomAD MAF (v. 2.1.1) | ACMG Interpretation | ClinVar Interpretation | Clinvar ID | References |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 26 | c.4298C > T | p.Ser1433Leu | rs200708197 | 7.583 × 105 | VUCS | VUS | 630306 | |
| 1 | 26 | c.4709T > C | p.Leu1570Ser | VUCS | |||||
| 1 | 26 | c.7057C > T | p.Gln2353Ter | LP | |||||
| 1 | 26 | c.10385A > G | p.Tyr3462Cys | VUCS | |||||
| 1 | 26 | c.10579C > T | p.Arg3527Trp | rs144467873 | 0.0001595 | LP | P/LP | 40223 | |
| 23 | 26 | c.10580G > A | p.Arg3527Gln | rs5742904 | 0.0002942 | LP | P/LP | 17890 | [9,35,36] |
| 1 | 26 | c.10672C > T | p.Arg3558Cys | VUCS | |||||
| 3 | 26 | c.10708C > T | p.His3570Tyr | rs201736972 | VUCS | ||||
| 1 | 26 | c.11477C > T | p.Thr3826Met | rs61744153 | 0.001592 | VUCS | LP/VUS/LB/B | 237735 | |
| 1 | 28 | c.11911G > A | p.Glu3971Lys | VUCS | |||||
| 0 | 28 | c.12005C > T | p.Ala4002Val | rs369364335 | 1.195 × 105 | VUCS | VUS | 898076 | [32] |
| 1 | 29 | c.12739C > T | p.Gln4247Ter | rs907126709 | LP | VUS | 544074 | ||
| 1 | 29 | c.13175G > A | p.Ser4392Asn | VUCS | |||||
| 1 | 29 | c.13480_13482del | p.Gln4494del | rs756545438 | VUCS | LP/VUS | 265896 |
| Patient Numbers | Phenotype | Variant 1 (Gene/Variant/Zygosity) | Variant 2 (Gene/Variant/Zygosity) | Cis/Trans Position (Evidence) |
|---|---|---|---|---|
| 423 | HoFH | LDLR:p.Gly592Glu (het) | LDLR:p.Glu418Ter (het) | Trans (genetic test of relatives) |
| 474 | Severe HetFH | LDLR:p.Gly592Glu (het) | LDLR:p.Ala771Glufs*9 (het) | Trans (long read sequencing) |
| 166 | HoFH | LDLR:c.941-3C > G (het) | LDLR:p.Cys329Tyr (het) | Trans (genetic test of relatives) |
| 722 | HoFH | LDLR:c.940 + 3_940 + 6del (het) | LDLR:p.Arg416Trp (het) | Unknown |
| 668 | HoFH | LDLR:p.Cys329Tyr (het) | LDLR:p.Gly592Glu (het) | Trans (genetic test of relatives) |
| 675 | HoFH | LDLR:p.Trp577Arg (hom) | ||
| 355 | HoFH | LDLR:p.Ile441Asn (het) | LDLR:p.Ile792LeufsTer137 (het) | Unknown |
| 687 | HetFH | LDLR:p.Leu401His (het) | PCSK9:p.Arg357Cys (het) | |
| 211 | Severe HetFH | LDLR:p.Cys329Tyr (het) | LDLR:p.Gly592Glu (het) | Cis (genetic test of relatives) |
| 336 | HetFH | LDLR:p.Lys390Glu (het) | PCSK9:p.Glu34Lys (het) | |
| R-6 | HetFH | LDLR:p.Val429Met (het) | APOB:p.Glu3971Lys (het) | |
| R-35 | HetFH | LDLR:p.Gly592Glu (het) | APOB:p.Arg3527Gln (het) | |
| R-83 | HetFH | LDLR:p.Gly592Glu (het) | APOB:p.Ser4392Asn (het) + PCSK9:p.Gly51Arg (het) | |
| R-115 | HetFH | APOB:p.Arg3527Gln (het) | PCSK9:p.Pro174Ser (het) | |
| 969 | HetFH | APOB:p.Arg3527Gln (het) | APOB:p.Gln4494del (het) | Unknown |
References
- Berberich, A.J.; Hegele, R.A. The complex molecular genetics of familial hypercholesterolaemia. Nat. Rev. Cardiol. 2019, 16, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Dharmayat, K.I.; Stevens, C.A.T.; Sharabiani, M.T.A.; Jones, R.S.; Watts, G.F.; Genest, J.; Ray, K.K.; Vallejo-Vaz, A.J. Prevalence of familial hypercholesterolemia among the general population and patients with atherosclerotic cardiovascular disease. Circulation 2020, 141, 1742–1759. [Google Scholar] [CrossRef] [PubMed]
- Beheshti, S.O.; Madsen, C.M.; Varbo, A.; Nordestgaard, B.G. Worldwide prevalence of familial hypercholesterolemia: Meta-analyses of 11 million subjects. J. Am. Coll. Cardiol. 2020, 75, 2553–2566. [Google Scholar] [CrossRef] [PubMed]
- Cuchel, M.; Bruckert, E.; Ginsberg, H.N.; Raal, F.J.; Santos, R.D.; Hegele, R.A.; Kuivenhoven, J.A.; Nordestgaard, B.G.; Descamps, O.S.; Steinhagen-Thiessen, E. Homozygous familial hypercholesterolaemia: New insights and guidance for clinicians to improve detection and clinical management. A position paper from the Consensus Panel on Familial Hypercholesterolaemia of the European Atherosclerosis Society. Eur. Heart J. 2014, 35, 2146–2157. [Google Scholar] [CrossRef] [PubMed]
- Iacocca, M.A.; Chora, J.R.; Carrié, A.; Freiberger, T.; Leigh, S.E.; Defesche, J.C.; Kurtz, C.L.; DiStefano, M.T.; Santos, R.D.; Humphries, S.E.; et al. ClinVar database of global familial hypercholesterolemia-associated DNA variants. Hum. Mutat. 2018, 39, 1631–1640. [Google Scholar] [CrossRef]
- Zakharova, F.M.; Damgaard, D.; Mandelshtam, M.Y.; Golubkov, V.I.; Nissen, P.H.; Nilsen, G.G.; Stenderup, A.; Lipovetsky, B.M.; Konstantinov, V.O.; Denisenko, A.D.; et al. Familial hypercholesterolemia in St.-Petersburg: The known and novel mutations found in the low density lipoprotein receptor gene in Russia. BMC Med. Genet. 2005, 6, 6. [Google Scholar] [CrossRef] [Green Version]
- Meshkov, A.N.; Malyshev, P.P.; Kukharchuk, V.V. Familial hypercholesterolemia in Russia: Genetic and phenotypic characteristics. Ter. Arkhiv 2009, 81, 23–28. (In Russian) [Google Scholar]
- Komarova, T.Y.; Korneva, V.A.; Kuznetsova, T.Y.; Golovina, A.S.; Vasilyev, V.B.; Mandelshtam, M.Y. Familial hypercholesterolemia mutations in Petrozavodsk: No similarity to St. Petersburg mutation spectrum. BMC Med. Genet. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Semenova, A.E.; Sergienko, I.V.; García-Giustiniani, D.; Monserrat, L.; Popova, A.B.; Nozadze, D.N.; Ezhov, M.V. Verification of underlying genetic cause in a cohort of Russian patients with familial hypercholesterolemia using targeted next generation sequencing. J. Cardiovasc. Dev. Dis. 2020, 7, 16. [Google Scholar] [CrossRef]
- Korneva, V.A.; Kuznetsova, T.Y.; Bogoslovskaya, T.Y.; Polyakov, D.S.; Vasilyev, V.B.; Orlov, A.V.; Mandelshtam, M.Y. Cholesterol levels in genetically determined familial hypercholesterolaemia in Russian Karelia. Cholesterol 2017, 2017, 9375818. [Google Scholar] [CrossRef]
- Civeira, F. Guidelines for the diagnosis and management of heterozygous familial hypercholesterolemia. Atherosclerosis 2004, 173, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.F.; Rodeh, O.; Penn, J.; Bai, X.; Reid, J.G.; Krasheninina, O.; Salerno, W.J. GLnexus: Joint variant calling for large cohort sequencing. bioRxiv 2018, 343970. [Google Scholar] [CrossRef]
- Smoove. Available online: https://github.com/brentp/smoove (accessed on 10 December 2020).
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl variant effect predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Gardner, E.J.; Lam, V.K.; Harris, D.N.; Chuang, N.T.; Scott, E.C.; Pittard, W.S.; Mills, R.E.; 1000 Genomes Project Consortium; Devine, S.E. The Mobile Element Locator Tool (MELT): Population-scale mobile element discovery and biology. Genome Res. 2017, 27, 1916–1929. [Google Scholar] [CrossRef] [Green Version]
- Ou, J.; Zhu, L.J. trackViewer: A Bioconductor package for interactive and integrative visualization of multi-omics data. Nat. Methods 2019, 16, 453–454. [Google Scholar] [CrossRef]
- Chora, J.R.; Medeiros, A.M.; Alves, A.C.; Bourbon, M. Analysis of publicly available LDLR, APOB, and PCSK9 variants associated with familial hypercholesterolemia: Application of ACMG guidelines and implications for familial hypercholesterolemia diagnosis. Genet. Med. 2018, 20, 591–598. [Google Scholar] [CrossRef] [Green Version]
- Leigh, S.; Futema, M.; Whittall, R.; Taylor-Beadling, A.; Williams, M.; den Dunnen, J.T.; Humphries, S.E. The UCL low-density lipoprotein receptor gene variant database: Pathogenicity update. J. Med. Genet. 2017, 54, 217–223. [Google Scholar] [CrossRef] [Green Version]
- Zakharova, F.M.; Tatishcheva, Y.A.; Golubkov, V.I.; Lipovetsky, B.M.; Konstantinov, V.O.; Denisenko, A.D.; Faergeman, O.; Vasilyev, V.B.; Mandelshtam, M.Y. Familial hypercholesterolemia in St. Petersburg: Diversity of mutations argues against a strong founder effect. Russ. J. Genet. 2007, 43, 1046–1052. [Google Scholar] [CrossRef]
- Zakharova, F.M.; Golubkov, V.I.; Mandelshtam, M.J.; Lipovetskii, B.M.; Gaitskhoki, V.S. Identification of novel missense mutation G571E, novel silent mutation H229H, nonsense mutation C74X, and four single nucleotide polymorphisms in the Low-Density Lipoprotein Receptor gene in patients with familial hypercholesterolemia from St. Petersburg. Russ. J. Bioorganic Chem. 2001, 27, 349–351. [Google Scholar] [CrossRef]
- Komarova, T.Y.; Golovina, A.S.; Grudinina, N.A.; Zakharova, F.M.; Korneva, V.A.; Lipovetskii, B.M.; Serebrenitskaya, M.P.; Konstantinov, V.O.; Vasilyev, V.B.; Mandelshtam, M.Y. “Finnish” mutations in LDL Receptor gene: A rare cause of familial hypercholesterolemia in St. Petersburg and Petrozavodsk. Bull. Exp. Biol. Med. 2013, 155, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Durst, R.; Colombo, R.; Shpitzen, S.; Avi, L.B.; Friedlander, Y.; Wexler, R.; Raal, F.J.; Marais, D.A.; Defesche, J.C.; Mandelshtam, M.Y.; et al. Recent origin and spread of a common Lithuanian mutation, G197del LDLR, causing familial hypercholesterolemia: Positive selection is not always necessary to account for disease incidence among Ashkenazi Jews. Am. J. Hum. Genet. 2001, 68, 1172–1188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakir, K.; Skobeleva, N.A.; Shevtsov, S.P.; Konstantinov, V.O.; Denisenko, A.D.; Schwartz, E.I. Two Novel Slavic Point Mutations in the Low-Density Lipoprotein Receptor Gene in Patients with Familial Hypercholesterolemia from St. Petersburg, Russia. Mol. Genet. Metab. 1998, 63, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Korneva, V.A.; Bogoslovskaia, T.I.; Kuznetsova, T.I.; Mandel’shtam, M.I.; Vasil’ev, V.B. Familial hypercholesterolemia due to a new mutation in the low density lipoprotein receptor gene. Klin. Med. 2014, 92, 49–53. (In Russian) [Google Scholar]
- Chakir, K.; Ju, M.M.; Shevtsov, S.P.; Golubkov, V.I.; Skobeleva, N.A.; Shur, Y.A.; Zakharova, F.M.; Lipovetskyi, B.M.; Konstantinov, V.O.; Denisenko, A.D.; et al. Two Novel Low-Density Lipoprotein Receptor Gene Mutations (E397X and 347delGCC) in St. Petersburg Familial Hypercholesterolemia. Mol. Genet. Metab. 1998, 65, 311–314. [Google Scholar] [CrossRef]
- Korneva, V.A.; Kuznetsova, T.I.; Komarova, T.I.; Golovina, A.S.; Mandel’shtam, M.I.; Konstantinov, V.O.; Vasil’ev, V.B. A case of familial hypercholesterolemia caused by a novel mutation p. FsS65:D129X of human low density lipoprotein receptor gene. Kardiologiia 2013, 53, 50–54. (In Russian) [Google Scholar]
- Tatishcheva, Y.A.; Mandelshtam, M.Y.; Golubkov, V.I.; Lipovetsky, B.M.; Gaitskhoki, V.S. Four New Mutations and Two Polymorphic Variants of the Low-Density Lipoprotein Receptor Gene in Familial Hypercholesterolemia Patients from St. Petersburg. Russ. J. Genet. 2001, 37, 1082–1087. [Google Scholar] [CrossRef]
- Komarova, T.Y.; Golovina, A.S.; Grudinina, N.A.; Zakharova, F.M.; Korneva, V.A.; Lipovetsky, B.M.; Serebrenitskaya, M.P.; Konstantinov, V.O.; Vasilyev, V.B.; Mandelshtam, M.Y. New mutations in low-density lipoprotein receptor gene in familial hypercholesterolemia patients from Petrozavodsk. Russ. J. Genet. 2013, 49, 673–676. [Google Scholar] [CrossRef]
- Voevoda, M.I.; Kulikov, I.V.; Shakhtshneider, E.V.; Maksimov, V.N.; Pilipenko, I.V.; Tereschenko, I.P.; Kobzev, V.F.; Romaschenko, A.G.; Nikitin, Y.P. The spectrum of mutations in the low-density lipoprotein receptor gene in the Russian population. Russ. J. Genet. 2008, 44, 1191–1194. [Google Scholar] [CrossRef]
- Mandelshtam, M.; Chakir, K.; Shevtsov, S.; Golubkov, V.; Skobeleva, N.; Lipovetsky, B.; Konstantinov, V.; Denisenko, A.; Gaitskhoki, V.; Schwartz, E. Prevalence of Lithuanian mutation among St. Petersburg Jews with familial hypercholesterolemia. Hum. Mutat. 1998, 12, 255–258. [Google Scholar] [CrossRef]
- Averkova, A.O.; Brazhnik, V.A.; Speshilov, G.I.; Rogozhina, A.A.; Koroleva, O.S.; Zubova, E.A.; Galyavich, A.S.; Tereshenko, S.N.; Boyeva, O.I.; Zateyshchikov, D.A. Targeted sequencing in patients with clinically diagnosed hereditary lipid metabolism disorder and acute coronary syndrome. Bull. RSMU 2018, 5, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Korneva, V.A.; Kuznetsova, T.Y.; Murtazina, R.Z.; Didio, A.V.; Bogoslovskaya, T.Y.; Mandelshtam, M.Y.; Vasilyev, V.B. The Familial Hypercholesterolemia caused by a novel human Low Density Lipoprotein Receptor Gene mutation c.1327 T>C (p.W433R). Kardiologiia 2017, 57, 12–16. (In Russian) [Google Scholar] [PubMed]
- Shakhtshneider, E.; Orlov, P.; Ivanoshchuk, D.; Makarenkova, K.; Ragino, Y.; Voevoda, M. Analysis of the LDLR gene variability in patients with Familial Hypercholesterolemia in Russia using targeted high throughput resequencing. Atherosclerosis 2017, 263, e227. [Google Scholar] [CrossRef]
- Pogoda, T.; Metelskaya, V.; Perova, N.; Limborska, S. Detection of the apoB-3500 mutation in a Russian family with coronary heart disease. Hum. Hered. 1998, 48, 291–292. [Google Scholar] [CrossRef]
- Shakhtshneider, E.; Ivanoshchuk, D.; Orlov, P.; Timoshchenko, O.; Voevoda, M. Analysis of the LDLR, APOB, PCSK9 and LDLRAP1 genes variability in patients with Familial Hypercholesterolemia in West Siberia using targeted high throughput resequencing. Atherosclerosis 2019, 287, e285. [Google Scholar] [CrossRef]
- Meshkov, A.N.; Stambol’skiĭ, D.V.; Krapivner, S.R.; Bochkov, V.N.; Kukharchuk, V.V.; Malyshev, P.P. Low density lipoprotein receptor gene mutations in patients with clinical diagnosis of familial hypercholesterolemia. Kardiologiia 2004, 44, 58–61. (In Russian) [Google Scholar]
- Mandelshtam, M.J.; Lipovetskyi, B.M.; Schwartzman, A.L.; Gaitskhoki, V.S. A novel deletion in the low-density lipoprotein receptor gene in a patient with familial hypercholesterolemia from Petersburg. Hum. Mutat. 1993, 2, 256–260. [Google Scholar] [CrossRef]
- Meshkov, A.N.; Stambol’skiĭ, D.V.; Nikitina, L.A.; Abdullaev, S.M.; Bochkov, V.N.; Tkachuk, V.A.; Kukharchuk, V.V.; Malyshev, P.P. Genetic factors of risk of ischemic heart disease development in patients with familial hypercholesterolemia. Kardiologiia 2005, 45, 10–14. (In Russian) [Google Scholar]


| Gene | Total (P/LP/VUS) | Possibly Unique including Novel for the Russian Population and Described Earlier (P/LP/VUS) | Novel (P/LP/VUS) | Described in Other World Populations |
|---|---|---|---|---|
| LDLR | 187 (38/53/95) * | 67 (11/19/37) | 34 (3/10/21) | 120 (27/34/58) |
| APOB | 14 (0/4/10) | 6 (0/1/5) | 6(0/1/5) | 8 (0/3/5) |
| PCSK9 | 23 (0/4/19) | 8(0/1/7) | 6 (0/1/5) | 5 (0/3/12) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meshkov, A.; Ershova, A.; Kiseleva, A.; Zotova, E.; Sotnikova, E.; Petukhova, A.; Zharikova, A.; Malyshev, P.; Rozhkova, T.; Blokhina, A.; et al. The LDLR, APOB, and PCSK9 Variants of Index Patients with Familial Hypercholesterolemia in Russia. Genes 2021, 12, 66. https://doi.org/10.3390/genes12010066
Meshkov A, Ershova A, Kiseleva A, Zotova E, Sotnikova E, Petukhova A, Zharikova A, Malyshev P, Rozhkova T, Blokhina A, et al. The LDLR, APOB, and PCSK9 Variants of Index Patients with Familial Hypercholesterolemia in Russia. Genes. 2021; 12(1):66. https://doi.org/10.3390/genes12010066
Chicago/Turabian StyleMeshkov, Alexey, Alexandra Ershova, Anna Kiseleva, Evgenia Zotova, Evgeniia Sotnikova, Anna Petukhova, Anastasia Zharikova, Pavel Malyshev, Tatyana Rozhkova, Anastasia Blokhina, and et al. 2021. "The LDLR, APOB, and PCSK9 Variants of Index Patients with Familial Hypercholesterolemia in Russia" Genes 12, no. 1: 66. https://doi.org/10.3390/genes12010066