
Estimation of Nuclear DNA Content in Some Aegilops Species: Best Analyzed Using Flow Cytometry
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
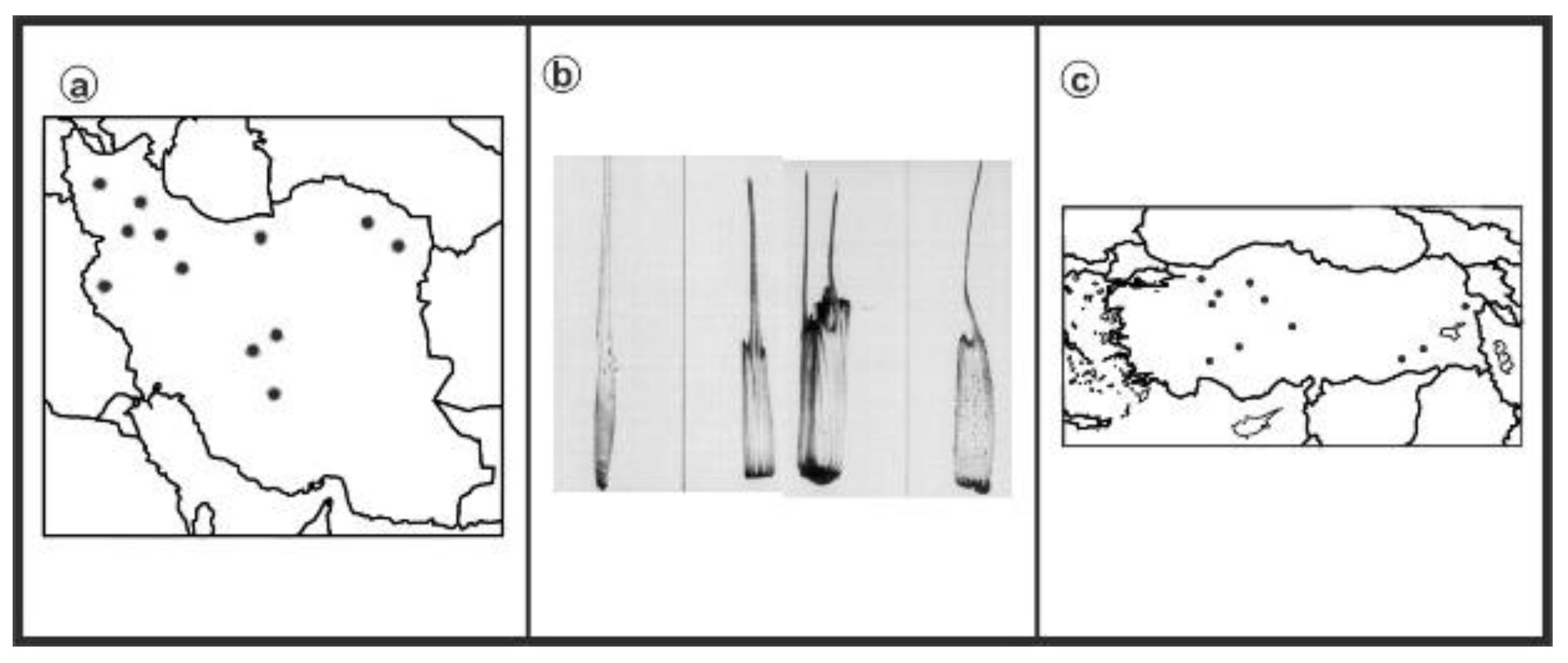
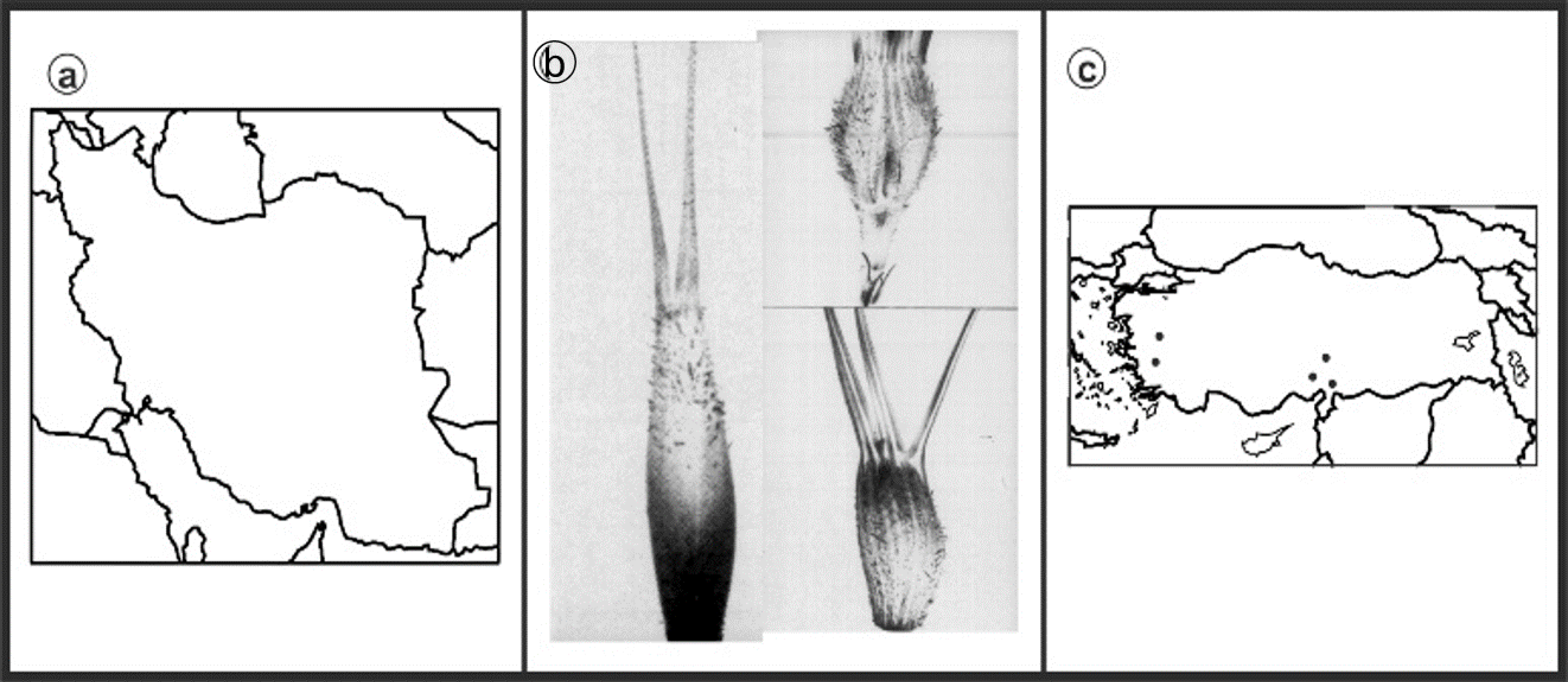
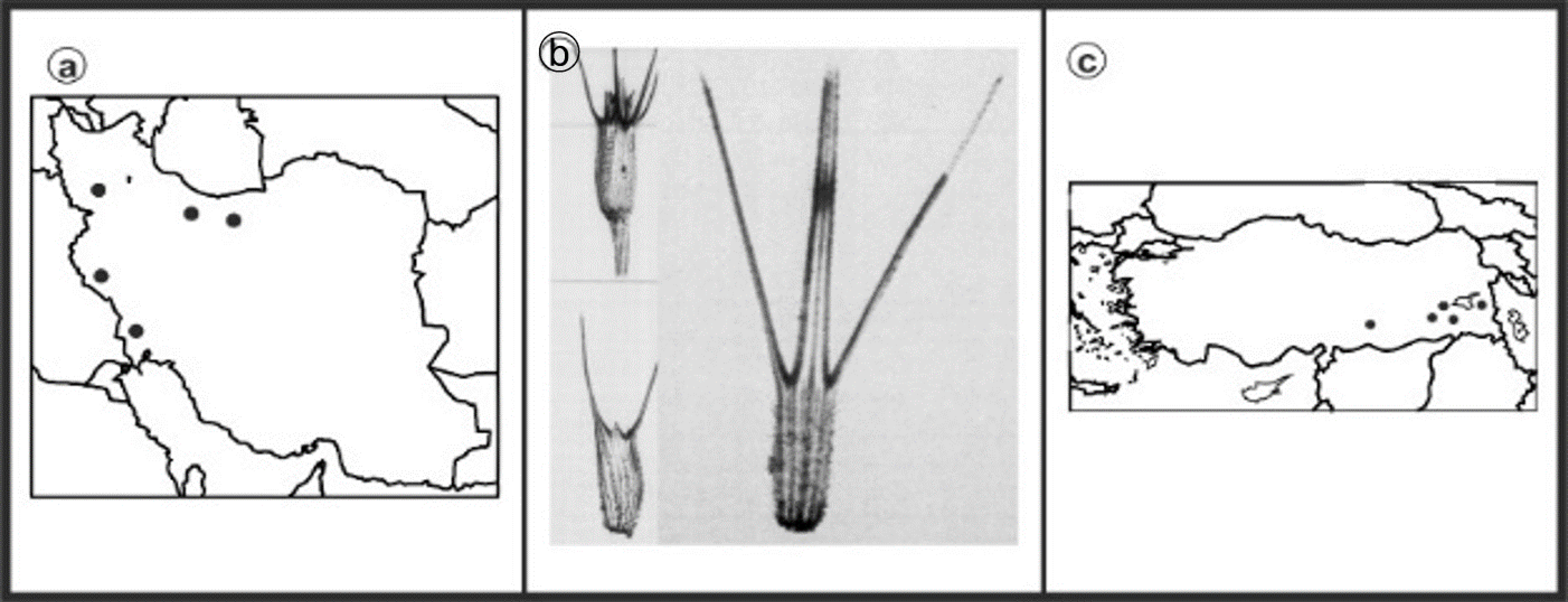
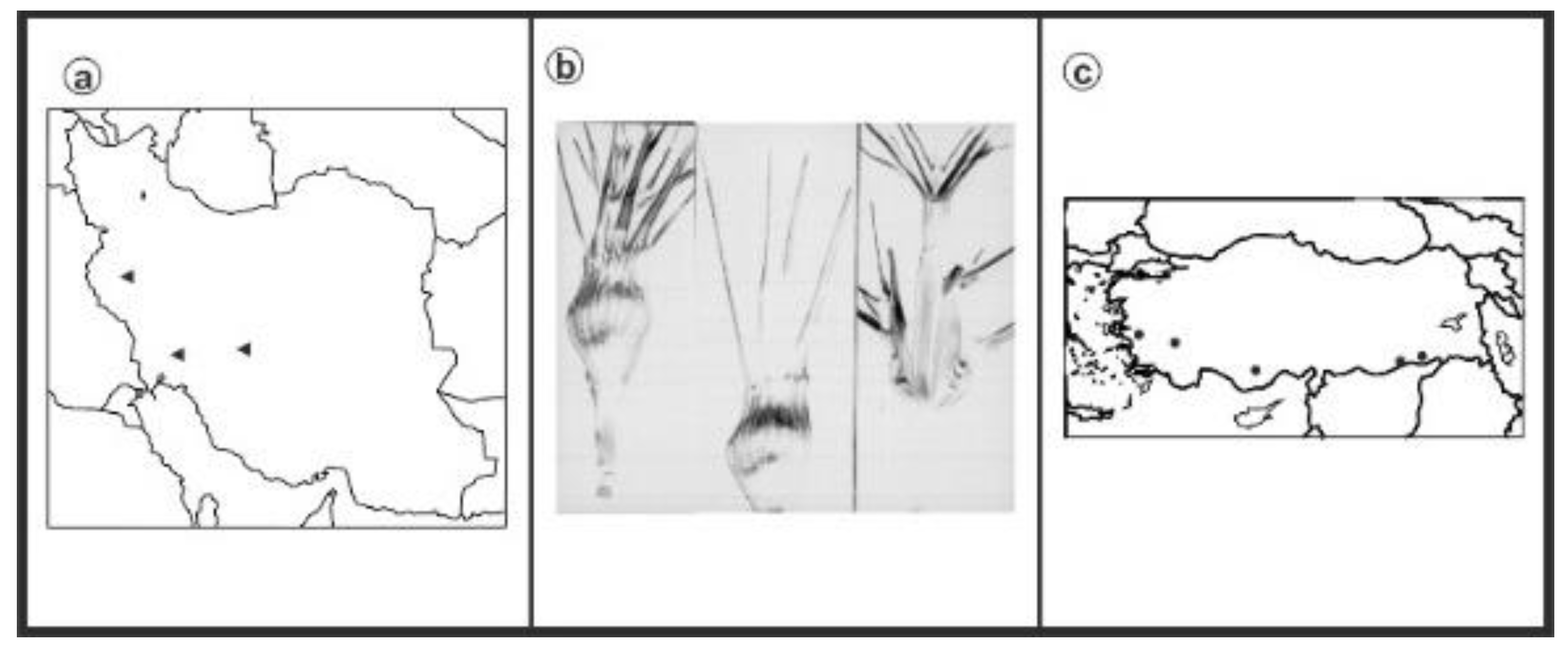
2.1. Experimental Materials
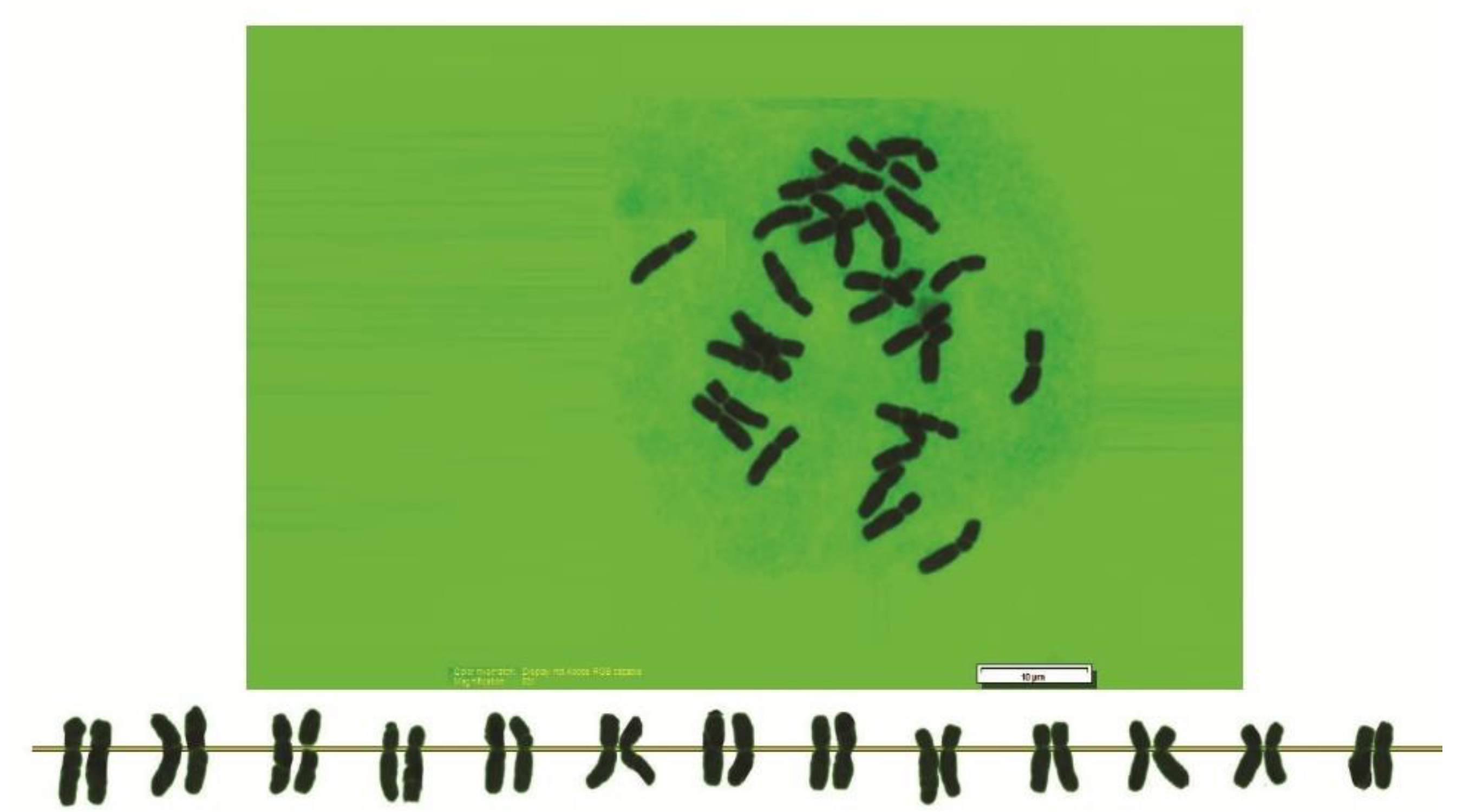
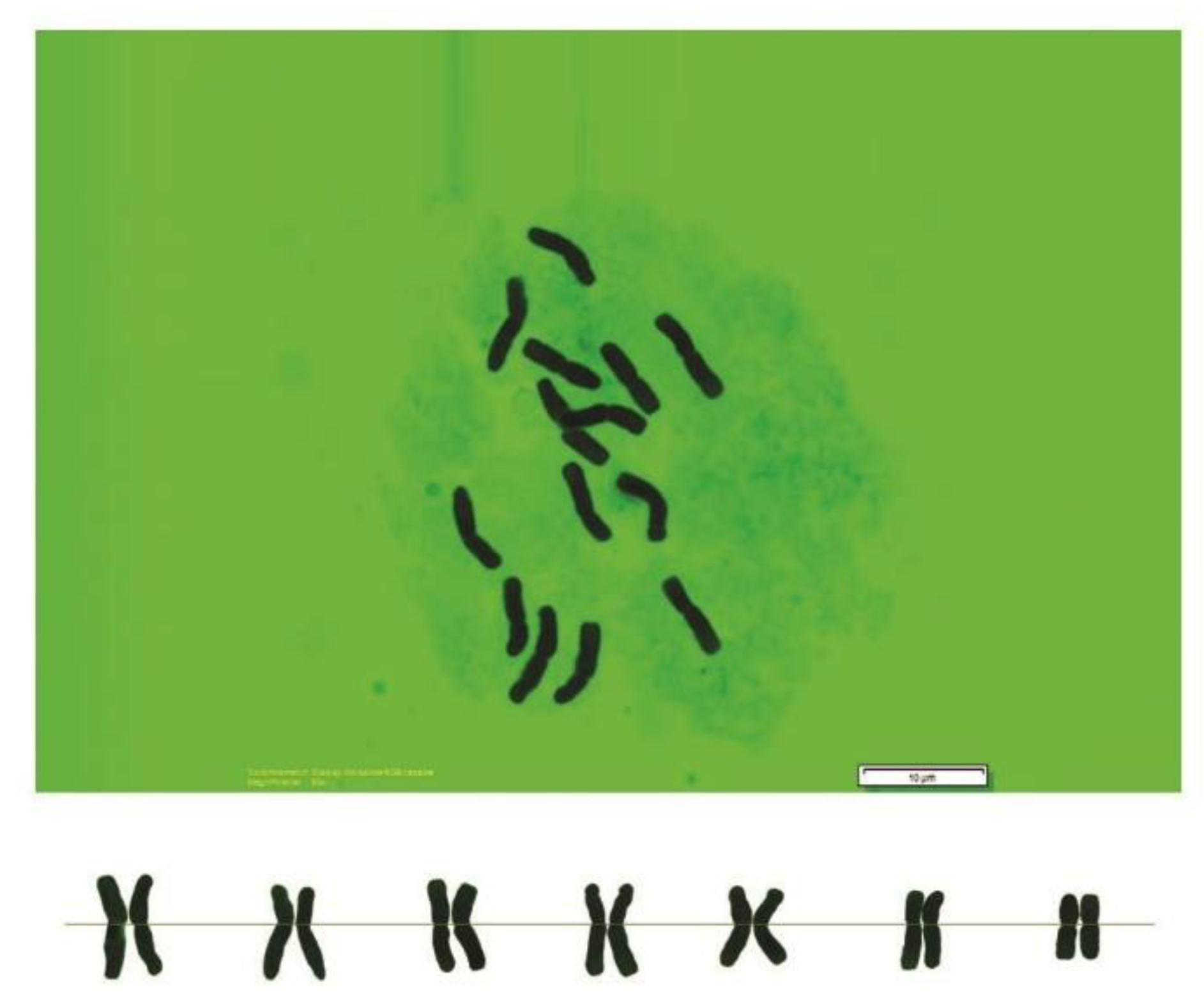
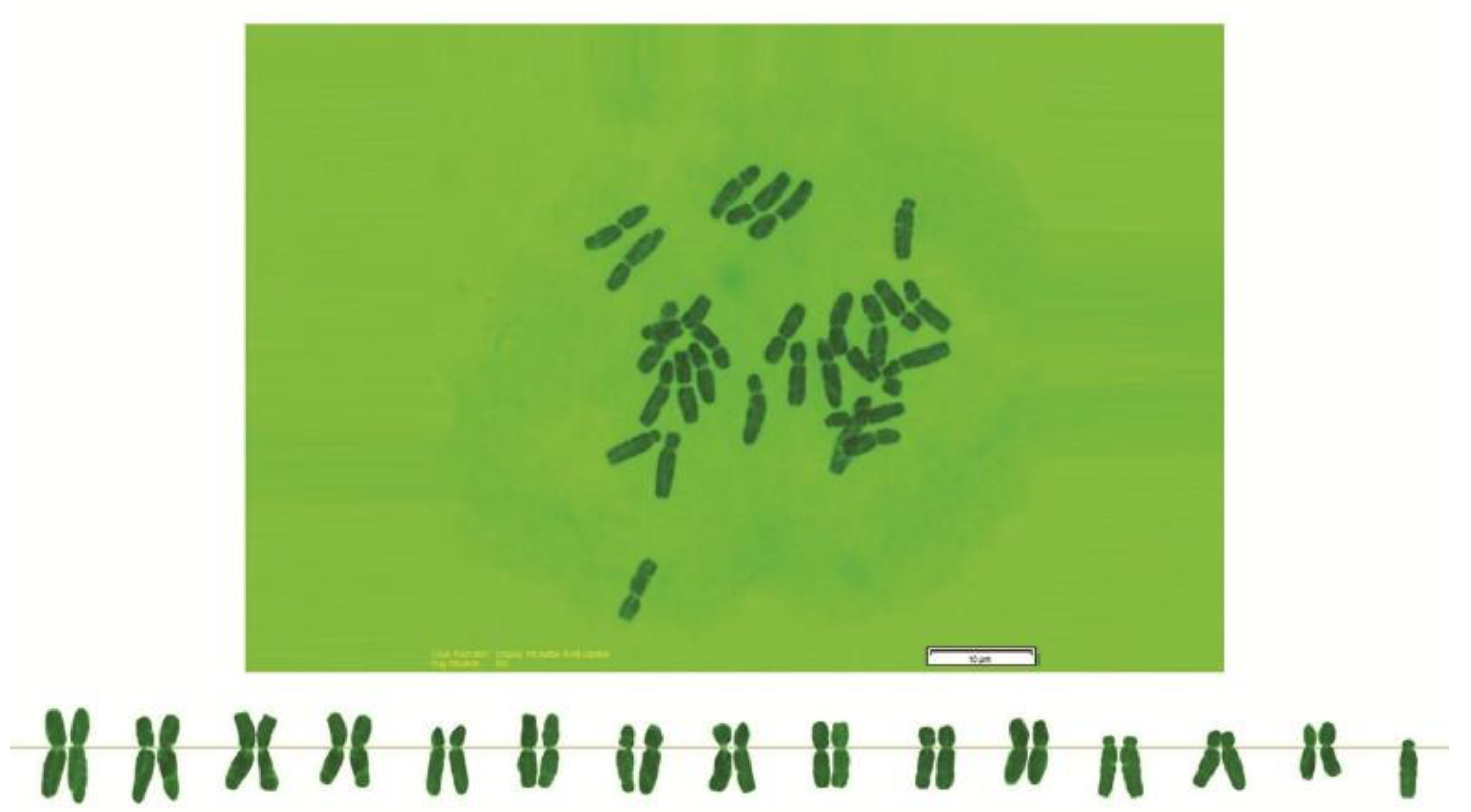
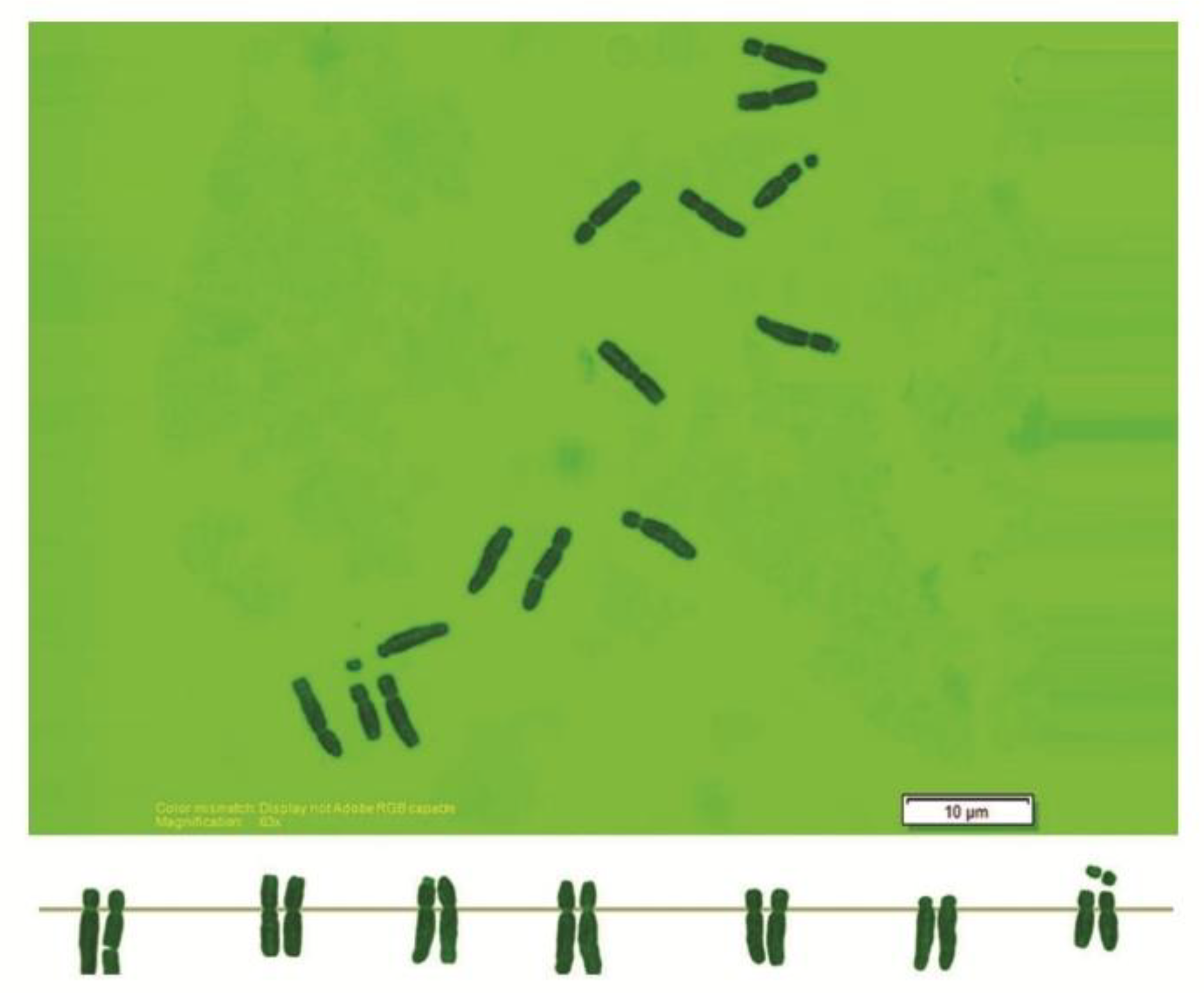
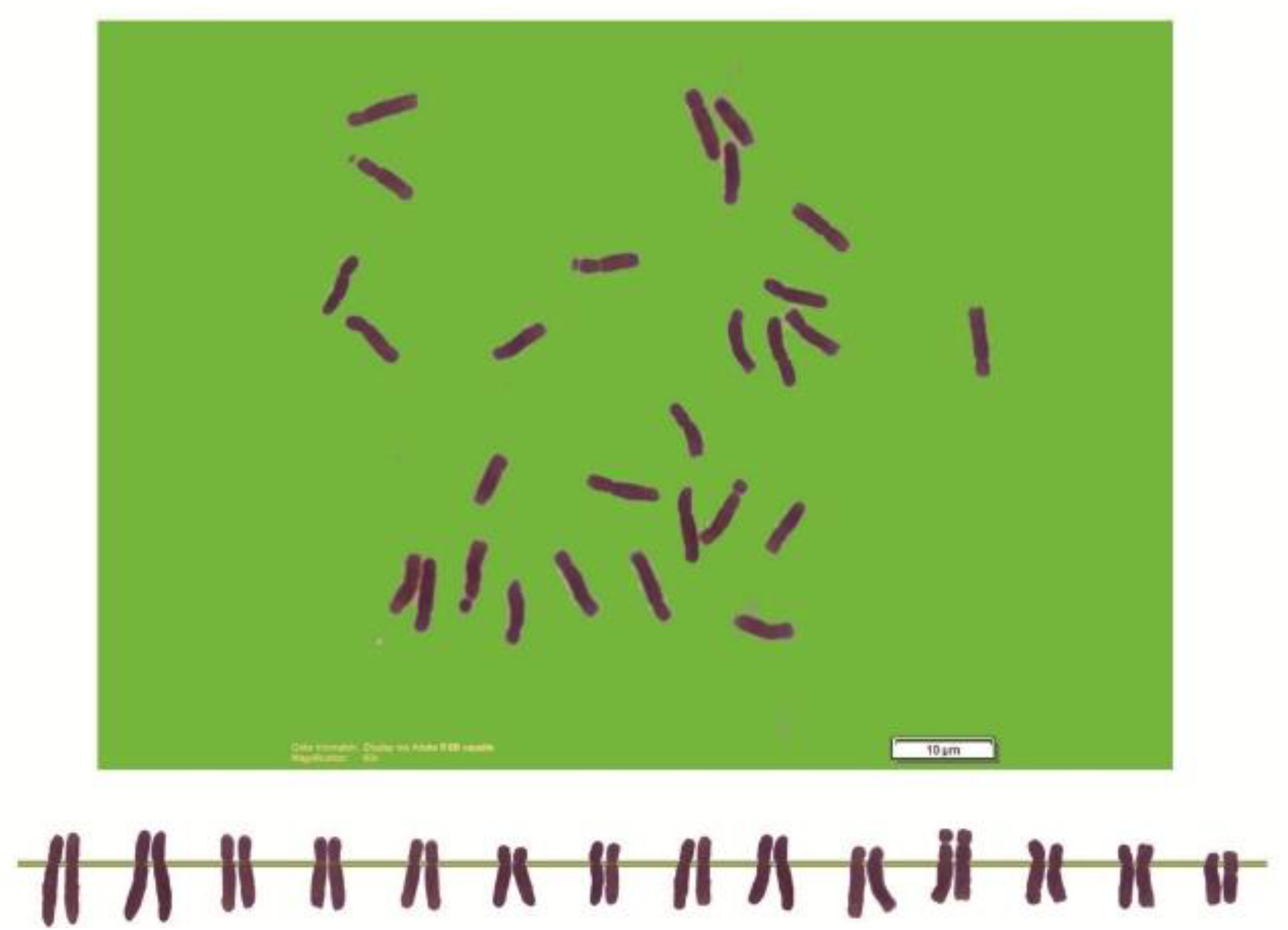
2.2. Chromosome Analysis
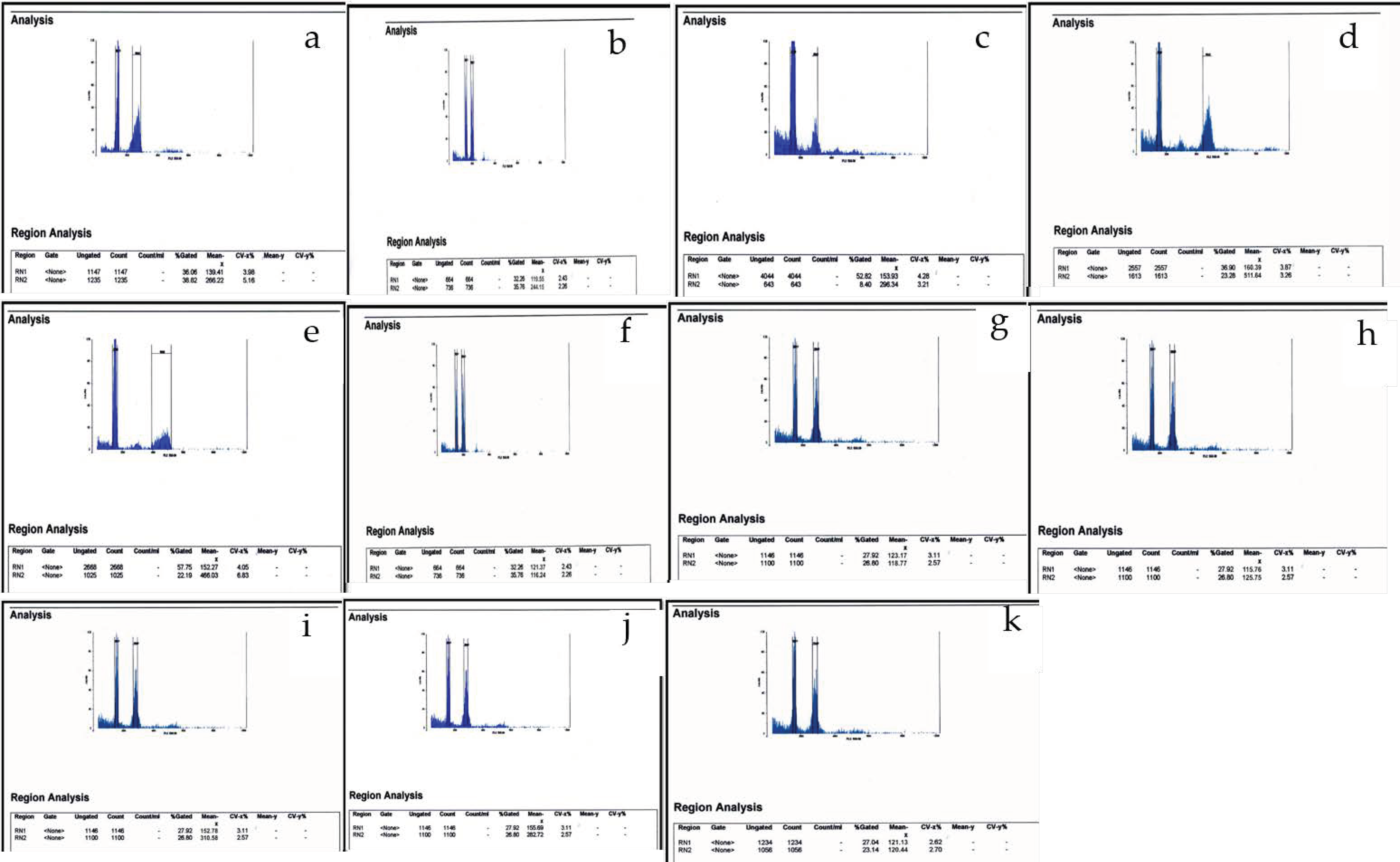
2.3. Determination of Nuclear DNA Content
3. Results























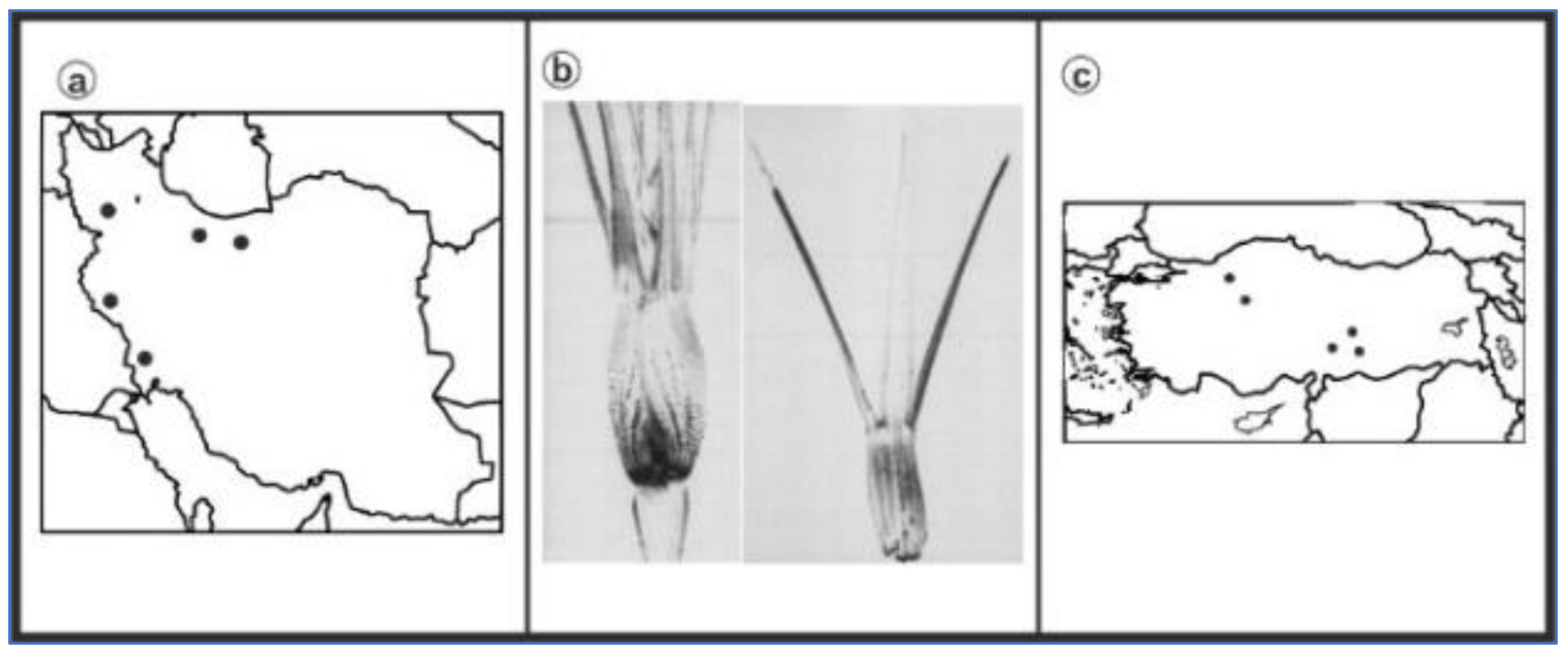{kind=link}
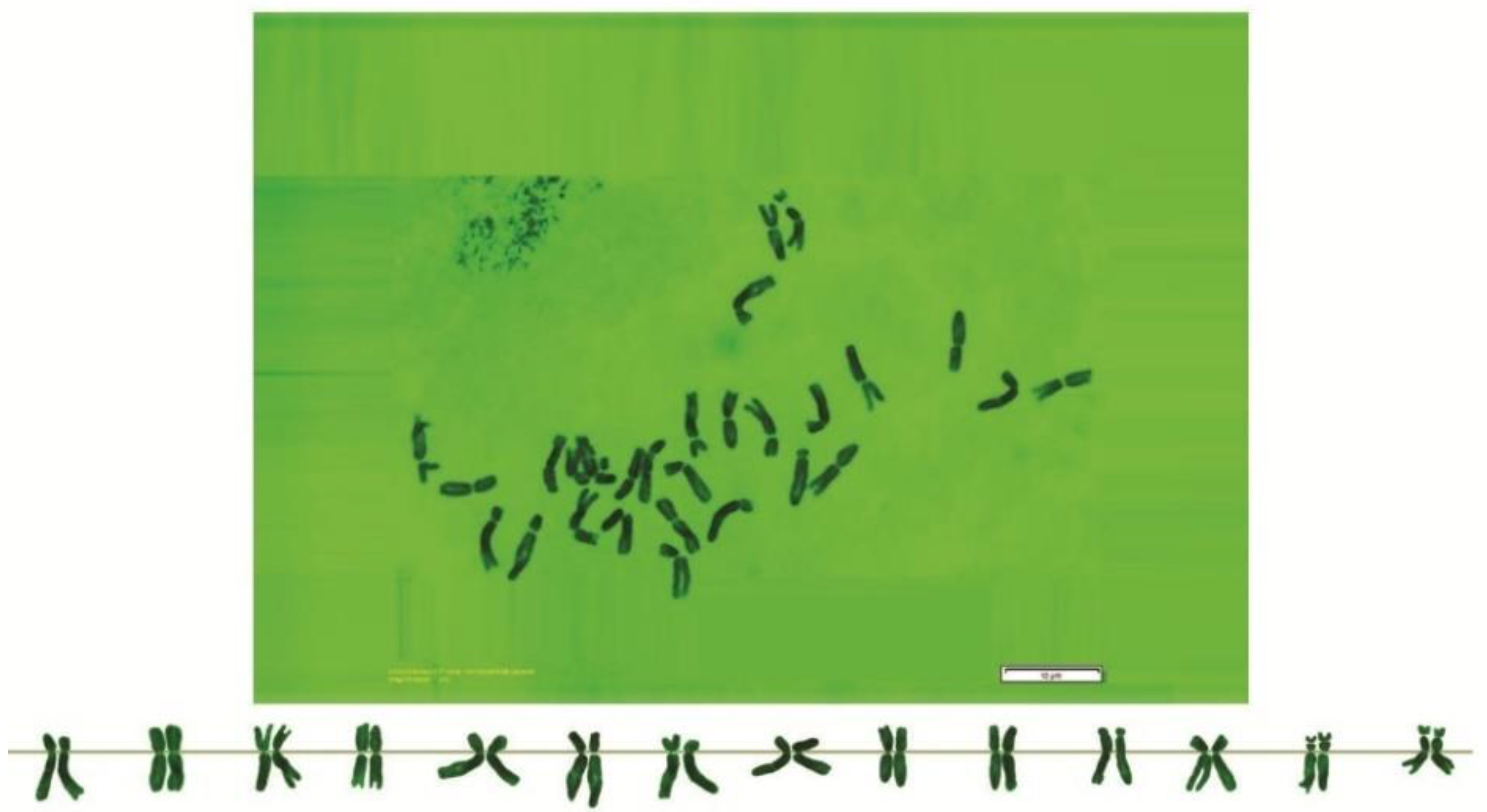{kind=link}
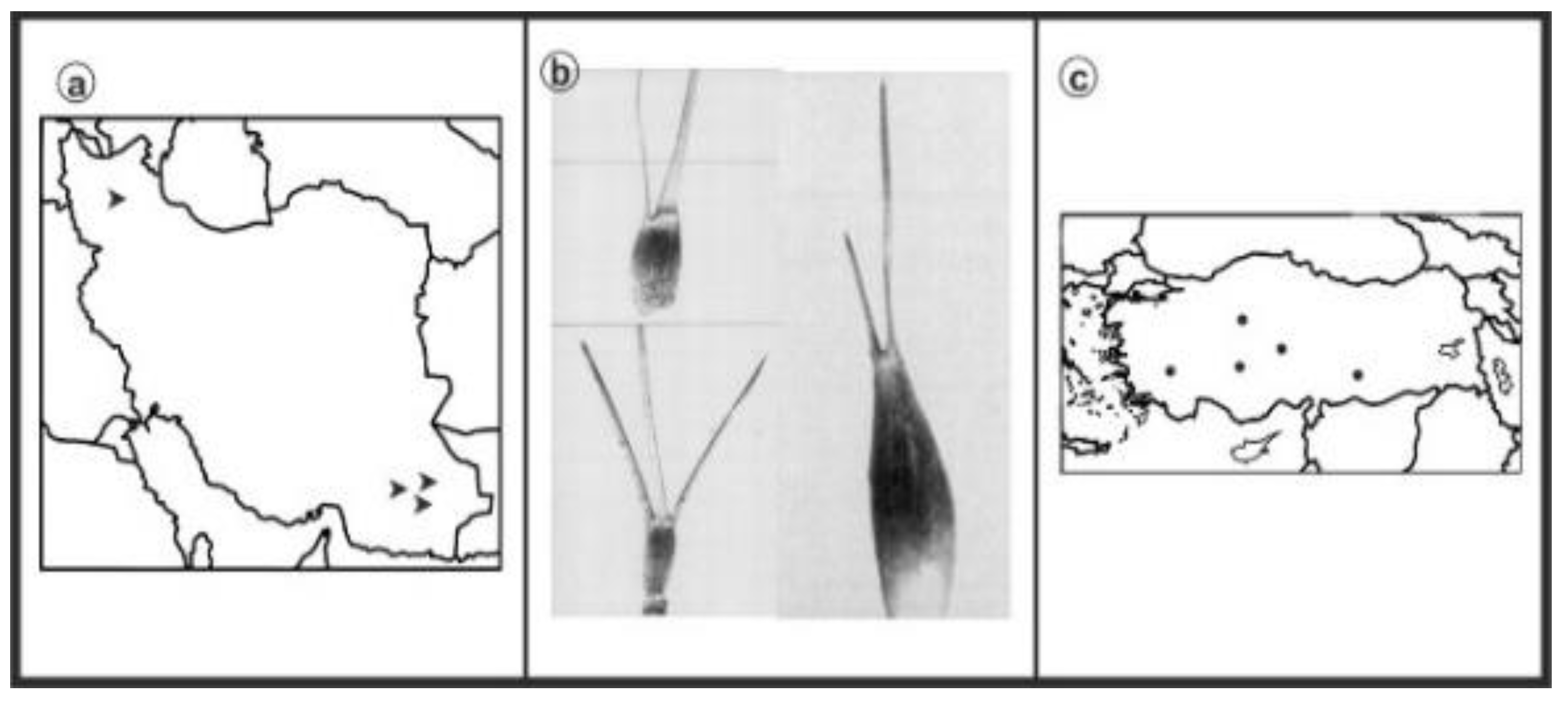{kind=link}
{kind=link}
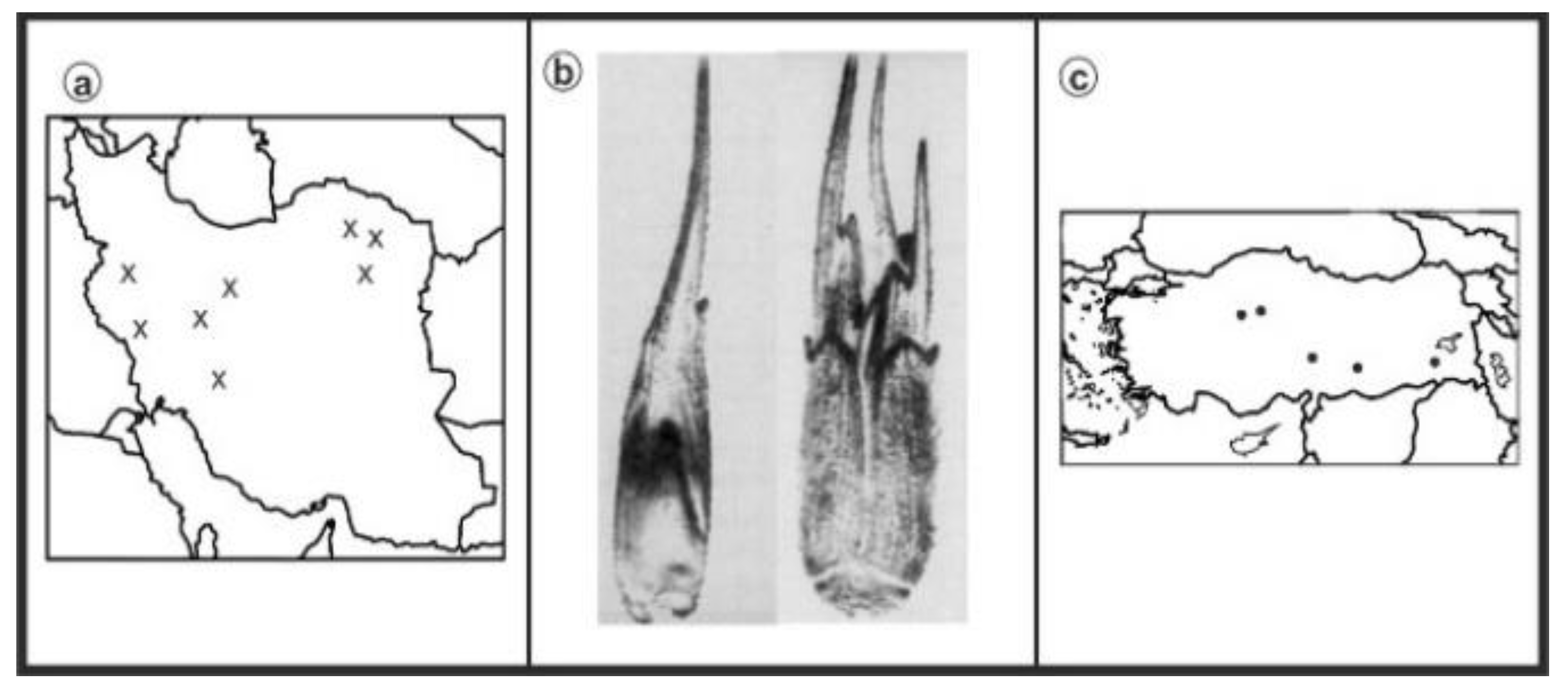{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | x | Genome 1 | DNA Content (pg/2C) * | DNA Content (pg/2C) ** | DNA Content (pg/2C) *** | DNA Content (pg/2C) **** | DNA Content (pg/2C) ***** |
|---|---|---|---|---|---|---|---|
| Ae. speltoides Var. aucheri | 2n = 2x = 14 | SS | - | - | - | 10.15 ± 0.04 | 10.22 ± 0.02 |
| Ae. speltoides Var. ligustica | 2n = 2x = 14 | SS | - | - | - | 10.95 ± 0.12 | 11.56 ± 0.01 |
| Ae. speltoides Var. Taush | 2n = 2x = 14 | SS | 10.20 ± 0.09 | 10.2 | 10.34 ± 0.08 | 10.25 ± 0.27 | 10.36 ± 0.36 |
| Ae. umbellulata | 2n = 2x = 14 | UU | 10.59 ± 0.11 | 10.1 | 10.76 ± 0.07 | 10.09 ± 0.04 | 10.58 ± 0.56 |
| Ae. biuncialis | 2n = 4x = 28 | UUMM | 20.61 ± 0.22 | 22.6 | 20.74 ± 0.04 | 19.89 ± 0.29 | 20.23 ± 0.35 |
| Ae. columnaris | 2n = 4x = 28 | UUMM | 21.75 ± 0.20 | 21.0 | 21.72 | 20.17 ± 0.16 | 21.75 ± 0.36 |
| Ae. crassa (4x) | 2n = 4x = 28 | DDMM | 21.29 ± 0.24 | 20.9 | 21.72 | 20.08 ± 0.01 | 20.44 ± 0.29 |
| Ae.cylindrica (4x) | 2n = 4x = 28 | CCDD | 18.79 ± 0.09 | 9.3 | 19.18 | 18.09 ± 0.05 | 18.61 ± 0.07 |
| Ae. triaristata | 2n = 4x = 28 | UUMM | 21.87 ± 0.25 | 31.0 | 21.28 ± 0.40 | 21.65 ± 0.05 | - |
| Ae. triuncialis | 2n = 4x = 28 | UUCC | 19.40 ± 0.17 | 18.9 | 19.86 ± 0.04 | 19.05 ± 0.03 | 19.27 ± 0.04 |
| Ae. crassa (6x) | 2n = 6x = 42 | DDDDMM | 33.63 ± 0.33 | 31.4 | 31.59 ± 0.03 | 33.97 ± 0.06 | |
| Ae.cylindrica (6x) | 2n = 6x = 42 | CCDDDD | - | - | - | 31. 81 ± 0.08 | 32.58 ± 0.25 |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arzani, A.; Ashraf, M. Cultivated ancient wheats (Triticum spp.): A potential source of health-beneficial food products. Compr. Rev. Food Sci. Food Saf. 2016, 16, 477–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzani, A.; Ashraf, M. Smart engineering of genetic resources for enhanced salinity tolerance in crop plants. Crit. Rev. Plant Sci. 2016, 35, 146–189. [Google Scholar] [CrossRef]
- Song, Z.P.; Zuo, Y.Y.; Xiang, Q.; Li, W.J.; Jian, L.I.; Gang, L.I.U.; Dai, S.F.; Yan, Z.H. Investigation of Aegilops umbellulata for stripe rust resistance, heading date, and iron, zinc, and gluten protein content. J. Integ. Agric. 2022, in press.
- Hajjar, R.; Hodgking, T. The use of wild relatives in crop improvement: A survey of developments over the last 20 years. Euphytica 2007, 156, 1–13. [Google Scholar] [CrossRef]
- Zaveri, E.B.; Lobell, D. The role of irrigation in changing wheat yields and heat sensitivity in India. Nat. Commun. 2019, 10, 4144. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Liu, G.; Wei, J.; Chen, Z.; Ding, Y.; Zheng, J. The climatic effects of irrigation over the middle and lower reaches of the Yangtze River, China. Agric. For. Meteorol. 2021, 308–309, 108550. [Google Scholar] [CrossRef]
- Jiang, X.; Hua, M.; Yang, X.; Hu, N.; Qiu, R.; Yang, S. Impacts of mist spray on rice field micrometeorology and rice yield under heat stress condition. Sci. Rep. 2020, 10, 1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issaka, Z.; Li, H.; Yue, J.; Tang, P.; Darko, R.O. Water-smart sprinkler irrigation, prerequisite to climate change adaptation: A review. J. Water Clim. Chang. 2018, 9, 383–398. [Google Scholar] [CrossRef] [Green Version]
- Uygan, D.; Cetin, O.; Alveroglu, V.; Sofuoglu, A. Improvement of water saving and economic productivity based on quotation with sugar content of sugar beet using linear move sprinkler irrigation. Agric. Water Manag. 2021, 255, 106989. [Google Scholar] [CrossRef]
- International Commission on Irrigation and Drainage. Sprinkler and Micro Irrigated Area in the World. Available online: http://www.icid.org/sprinklerandmircro (accessed on 13 October 2021).
- Pour-Aboughadareh, A.; Ahmadi, J.; Mehrabi, A.A.; Moghaddam, M.; Etminan, E.; Siddique, K.H.M. Physiological responses to drought stress in wild relatives of wheat: Implications for wheat improvement. Acta Physiol. Plant. 2017, 39, 49. [Google Scholar] [CrossRef]
- Davis, P.H. Gramineae. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1965; Volume 1. [Google Scholar]
- Slageren, M.W. Wild Wheats: A Monograph of Aegilops L. and Amblyopyrum (Jaub. and Spach) Eig (Poaceae); Wageningen Agricultural University: Wageningen, The Netherlands, 1994; pp. 94–107. [Google Scholar]
- Bor, N.L. Gramineae. In Flora Iranica; Rechinger, K.H., Ed.; Akademische Druk-Und Verlagsanstalt: Graz, Austria, 1970; Volume 70. [Google Scholar]
- Eig, A.V. Monographisch-Kristische Ubersicht der Gattung Aegilops; Verlag des Repertoriums: Berlin, Germany, 1929. [Google Scholar]
- McFadden, E.S.; Sears, E.R. The origin of Triticum spelta and its free testing hexaploid relatives. J. Heredity 1994, 37, 107–116. [Google Scholar] [CrossRef]
- Saini, P.; Kaur, H.; Tyagi, V.; Saini, P.; Ahmed, N.; Dhaliwal, H.S.; Sheikh, I. Nutritional value and end-use quality of durum wheat. Cereal Res. Commun. 2022, 1–12. [Google Scholar] [CrossRef]
- Schneider, A.; Molnar, I.; Molnar-Lang, M. Utilization of Aegilops (goat grass) species to widen the genetic diversity of cultivated wheat. Euphitica 2008, 163, 1–19. [Google Scholar] [CrossRef]
- Weide, A.; Rieh, S.; Zeidi, M.; Conard, N.J. Using new morphological criteria to identify domesticated emmer wheat at the aceramic Neolithic site of Chogha Golan (Iran). J. Archaeol. Sci. 2013, 57, 109–118. [Google Scholar] [CrossRef]
- Kilian, B.; Ozkan, H.; Deusch, O.; Effgen, S.; Brandolini, A.; Kohl, J.; Martin, W.; Salamini, F. Independent wheat B and G genome origins in outcrossing Aegilops progenitor haplotypes. Mol. Bio. Evol. 2007, 24, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Kilian, B.; Mammen, K.; Millet, E.; Sharma, R.; Graner, A.; Salamini, F.; Hammer, K.; Ozkan, H. Wild Crop Relatives: Genomic and Breeding Resources: Cereals; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar]
- Swift, H. The constancy of deoxyribose nucleic acid in plant nuclei. Proc. Natl. Acad. Sci. USA 1950, 360, 643–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuta, Y.; Nishikawa, K.; Kimizuka, T. Quantitative comparison of nuclear DNA in section Sitopsis of genus Aegilops. Jap. J. Gen. 1977, 52, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Furuta, Y. DNA content per nucleus in Aegilops species. Wheat Inf. Ser. 1970, 30, 20–22. [Google Scholar]
- Maranon, T.; Grubb, P.J. Physiological basis and ecological significance of the seed size and relative growth rate relationship in Mediterranean annuals. Func. Ecol. 1993, 7, 591–599. [Google Scholar] [CrossRef]
- Eilam, T.; Anikster, Y.; Millet, E.; Manisterski, J.; Sagi-Assif, O.; Feldman, M. Genome size and genome evolution in diploid Triticeae species. Genome 2007, 50, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Rees, H.; Walters, M.R. Nuclear DNA and the evolution of wheat. Heredity 1965, 20, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, K.; Furuta, Y. DNA content of nucleus and individual chromosomes and its evolutionary significance. In Proceedings of the 5th International Wheat Genetics Symposium, New Delhi, India, 23–28 February 1978; Society of Genetics and Plant Breeding, Indian Agricultural Research Institute: New Delhi, Indian, 1978; pp. 133–138. [Google Scholar]
- Hulgenhof, E.; Weidhase, R.A.; Schlegel, R.; Tewes, A. Flow cytometric determination of DNA content in isolated nuclei of cereals. Genome 1988, 30, 565–569. [Google Scholar] [CrossRef]
- Bennett, M.D.; Smith, J.B. Nuclear DNA amounts in angiosperms. Philos. Trans. R. Soc. Lond. B 1976, 274, 227–274. [Google Scholar] [CrossRef]
- Eilam, T.; Anikster, Y.; Millet, E.; Manisterski, J.; Feldman, M. Nuclear DNA amount and genome downsizing in natural and synthetic allopolyploids of the genera Aegilops and Triticum. Genome 2008, 51, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Brummer, E.C.; Cezcarro, P.M.; Luth, D. Ploidy determination of alfalfa germplasm accessions using flow cytometry. Crop Sci. 1999, 39, 1202–1207. [Google Scholar] [CrossRef]
- Rayburn, A.L.; Auger, J.A.; Benzinger, E.A.; Hepburn, A.G. Detection of intraspecific DNA variation in Zea mays L. by flow cytometry. J. Exper. Bot. 1989, 40, 1179–1185. [Google Scholar] [CrossRef]
- Heslop-Harrison, J.S. Flow cytometry and chromosome sorting. In Plant Chromosomes: Laboratory Methods; Heslop-Harrison, J.S., Schwarzacher, T., Eds.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Arumuganathan, K.; Earle, E.D. Estimation of nuclear DNA content of plants by flow cytometry. Plant Mol. Biol. Rep. 1991, 9, 229–233. [Google Scholar] [CrossRef]
- Vogel, K.P.K.; Arumuganathan, K.; Jensen, B. Nuclear DNA content of perennial grasses of the tribe Triticea. Crop Sci. 1999, 39, 661–667. [Google Scholar] [CrossRef] [Green Version]
- Hultquist, S.J.; Vogel, K.P.; Lee, D.J.; Arumuganathan, K.; Kaeppeler, S. DNA content and chloroplast DNA polymorphisms among Switch grasses from remnant midwestern prairies. Crop Sci. 1997, 37, 594–598. [Google Scholar] [CrossRef]
- Lu, K.; Kaepler, S.M.; Vogel, K.P.; Arumuganathan, K.; Lee, D.J. Nuclear DNA content and chromosome numbers in switchgrass. Great Plains Res. 1998, 8, 269–280. [Google Scholar]
- Dpooležel, J.; Binarová, P.; Lcretti, S. Analysis of nuclear DNA content in plant cells by flow cytometry. Biol. Planta 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82 (Suppl. 1), 17–26. [Google Scholar] [CrossRef] [Green Version]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
- Tuna, M.; Teykin, E.; Büyükbaşar, A.; Arslanoglu, F.; Ozkan, H.; Şehirali, S.; Akkaya, M. Nuclear DNA Amounts of Diploid and Polyploid Aegilops and Triticum Species, The 3rd Asian Chromosome Colloquium, 2008.
- Kimber, G.; Tsunewaki, K. Genome symbols and plasma types in the wheat group. In Proceedings of the 7th International Wheat Genetics Symposium, Cambridge, UK, 13–19 July 1988; pp. 1209–1211. [Google Scholar]
- Temsch, E.M.; Koutecký, P.; Urfus, T.; Šmarda, P.; Doležel, J. Reference standards for flow cytometric estimation of absolute nuclear DNA content in plants. Cytom. Part A 2022, 101, 710–724. [Google Scholar] [CrossRef] [PubMed]
- Vrána, J.; Cápal, P.; Bednářová, M.; Doležel, J. Flow cytometry in plant research: A success story. In Applied Plant Cell Biology; Nick, P., Opatrný, Z., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Greilhuber, J.; Doležel, J. 2C or not 2C: A closer look at cell nuclei and their DNA content. Chromosoma 2009, 118, 391–400. [Google Scholar] [CrossRef]
- Greilhuber, J.; Doležel, J.; Lysák, M.A.; Bennett, M.D. The origin, evolution and proposed stabilisation of the terms ‘genome size’ and ‘C-value’ to describe nuclear DNA contents. Ann. Bot. 2005, 94, 255–260. [Google Scholar] [CrossRef]
- Bakhshi, B.; Aghaei, M.; Bihamta, M.; Darvish, F.; Zarifi, E. Ploidy determination of Aegilops cylindrica host accessions of Iran by using flow cytometry and chromosome counting. Iran. J. Bot. 2010, 16, 258–266. [Google Scholar]
- Olanj, N.; Sonboli, A. Variation of DNA amount in 22 populations of Tanacetum L. (Asteraceae, Anthemideae) in Iran: Palynology, Morphology and Ecological implications. J. Mol. Cell. Res. 2021, 33, 299–309. [Google Scholar]
- Greilhuber, J. Intraspecific variation in genome size in angiosperms: Identifying its existence. Ann. Bot. 2005, 95, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.G. When does intraspecific variation become taxonomically significant? Ann. Bot. 2005, 95, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.; Khurshid, Z.; Sabreena; Bali, B.S.; Ganai, B.A.; Sayyed, R.Z.; Poczai, P.; Zaman, M. Congruency between environmental DNA and palaeoecology for biodiversity monitoring and palaeo-environmental reconstruction: A critical Appraisal. Int. J. Env. Res. Pub. Health 2022, 19, 9445. [Google Scholar] [CrossRef]
- Hassan, S.; Sabreena; Poczai, P.; Ganai, B.A.; Almalki, W.H.; Gafur, A.; Sayyed, R.Z. Environmental DNA Metabarcoding: A Novel contrivance for documenting terrestrial biodiversity. Biology 2022, 11, 1297. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Youzhi, M.; Toshiyuki, W.; Lian Kee, C.; Young, K.; Seong-Whan, P.; Susumu, U.; Kiichi, F. Flow karyotypes and chromosomal DNA contents of genus Triticum species and rye (Secale cereale). Chromosome Res. 2004, 12, 93–102. [Google Scholar] [CrossRef] [PubMed]
| Turkey | Iran | ||
|---|---|---|---|
| Species | Ecotypes | Species | Ecotypes |
| Ae. biuncialis | Adiyaman | Ae. biuncialis | Urmia |
| Ae. biuncialis | Ankara | Ae. biuncialis | Rasht |
| Ae. biuncialis | Kahraman Marash | Ae. biuncialis | Saari |
| Ae. biuncialis | Malatya | Ae. biuncialis | Kerman |
| Ae. biuncialis | Gerede | Ae. biuncialis | Shiraz |
| Ae. columnaris | Adiyaman | Ae. columnaris | Neyriz |
| Ae. columnaris | Ankara | Ae. columnaris | Khoramabad |
| Ae. columnaris | Nevsehir | Ae. columnaris | Saghez |
| Ae. columnaris | Van | Ae. columnaris | Zahedan |
| Ae. columnaris | Denizli | Ae. columnaris | Zanjan |
| Ae. crassa | Adiyaman | Ae. crassa | Gazvin |
| Ae. crassa | Ankara | Ae. crassa | Aligudarz |
| Ae. crassa | Kirikkale | Ae. crassa | Shiraz |
| Ae. crassa | Tufanbeyli | Ae. crassa | Ilam |
| Ae. crassa | Konya | Ae. crassa | Marvdasht |
| Ae. cylindrica | Ankara | Ae. cylindrica | Semnan |
| Ae. cylindrica | Konya | Ae. cylindrica | Tabriz |
| Ae. cylindrica | Kirikkale | Ae. cylindrica | Mashhad |
| Ae. cylindrica | Haymana | Ae. cylindrica | Shiraz |
| Ae. cylindrica | Igdir | Ae. cylindrica | Ghorghan |
| Ae. speltoides var. aucheri | Ankara | Ae. speltoides var. aucheri | Saari |
| Ae. speltoides var. aucheri | Konya | Ae. speltoides var. aucheri | Ilam |
| Ae. speltoides var. aucheri | Sanliurfa | Ae. speltoides var. aucheri | Zahedan |
| Ae. speltoides var. aucheri | Corum | Ae. speltoides var. aucheri | Arak |
| Ae. speltoides var. aucheri | Mus | Ae. speltoides var. aucheri | Sanandaj |
| Ae. speltoides var. ligustica | Konya | Ae. speltoides var. ligustica | Kermanshah |
| Ae. speltoides var. ligustica | Erzurum | Ae. speltoides var. ligustica | Ghuchan |
| Ae. speltoides var. ligustica | Igdir | Ae. speltoides var. ligustica | Zanjan |
| Ae. speltoides var. ligustica | Corum | Ae. speltoides var. ligustica | Shiraz |
| Ae. speltoides var. ligustica | Karaman | Ae. speltoides var. ligustica | Saari |
| Ae. tauschii | Kirsehir | Ae. tauschii | Shiraz |
| Ae. tauschii | Konya | Ae. tauschii | Yasuj |
| Ae. tauschii | Sanliurfa | Ae. tauschii | Khorasan |
| Ae. tauschii | Van | Ae. tauschii | Urmia |
| Ae. tauschii | Kahramanmaras | Ae. tauschii | Zahedan |
| Ae. triaristata | Adana | Ae. triuncialis | Hamedan |
| Ae. triaristata | Osmaniye | Ae. triuncialis | Ahvaz |
| Ae. triaristata | Tufanbeyli | Ae. triuncialis | Lahijan |
| Ae. triaristata | Aydin | Ae. triuncialis | Kermanshah |
| Ae. triaristata | Manisa | Ae. triuncialis | Shiraz |
| Ae. triuncialis | Adiyaman | Ae. umbellulata | Tabriz |
| Ae. triuncialis | Batman | Ae. umbellulata | Esfahan |
| Ae. triuncialis | Siirt | Ae. umbellulata | Ghonbad |
| Ae. triuncialis | Van | Ae. umbellulata | Ardebil |
| Ae. triuncialis | Bitlis | Ae. umbellulata | Kerman |
| Ae. umbellulata | Mardin | ||
| Ae. umbellulata | Nuseybin | ||
| Ae. umbellulata | Denizli | ||
| Ae. umbellulata | Selcuk | ||
| Ae. umbellulata | Mus | ||
| Species | 2n | MTCL ± Se (µm) | AR + Se | KF |
|---|---|---|---|---|
| Ae. biuncialis | 28 | 108.32 ± 0.63 | 2.54 ± 0.92 | 10sm + 4st + 3sat |
| Ae. columnaris | 28 | 103 ± 0.21 | 2.12 ± 0.29 | 10sm + 3m + 1st + 2sat |
| Ae. crassa (4x) | 28 | 156 ± 0.21 | 1.46 ± 0.03 | 11m + 3sm + 2sat |
| Ae. crassa (6x) | 42 | 217.39 ± 0.14 | 1.36 ± 0.03 | 20m + 1sm + 3sat |
| Ae. cylindrica (4x) | 28 | 105.97 ± 0.19 | 2.37 ± 0.53 | 8sm + 5st + 1m + 1sat |
| Ae. cylindrica (6x) | 42 | 237 ± 0.12 | 2.29 ± 0.19 | 12sm + 8st + 2m + 2sat |
| Ae. speltoides Var. Aucheri | 14 | 56.22 ± 0.37 | 1.37 ± 0.04 | 6m + 1sm + 1sat |
| Ae. speltoides Var. Ligustica | 14 | 82.12 ± 0.17 | 1.52 ± 0.22 | 5sm + 2m + 5sat |
| Ae. speltoides Var. Taush. | 14 | 63.18 ± 0.33 | 1.38 ± 0.18 | 7m |
| Ae. triuncialis | 28 | 108.52 ± 0.16 | 2.45 ± 0.91 | 14st |
| Ae. umbellulata | 28 | 25.95 ± 0.09 | 2.84 ± 0.17 | 5st + 1sm + 1t + 1sat |
| Species | 2n | MTCL | AR | KF |
|---|---|---|---|---|
| Ae. biuncialis | 28 | 98.69 ± 0.61 | 2.15 ± 0.69 | 13sm + 1m + 3sat |
| Ae. columnaris | 28 | 103.32 ± 0.79 | 2.12 ± 1.17 | 10sm + 3m + 1st + 2sat |
| Ae. crassa (4x) | 28 | 153.35 ± 0.49 | 1.61 ± 0.05 | 11m + 3sm + 2sat |
| Ae. crassa (6x) | 42 | 206.11 ± 0.03 | 1.13 ± 0.04 | 10m + 1sm + 3sat |
| Ae. cylindrica (4x) | 28 | 81.57 ± 0.10 | 2.39 ± 0.21 | 8sm + 5st + 1m + 1sat |
| Ae. cylindrica (6x) | 42 | 245 ± 0.08 | 2.30 ± 0.12 | 12sm + 8st + 2m + 2sat |
| Ae. speltoides Var. Aucheri | 14 | 52 ± 0.12 | 1.31 ± 0.09 | 7m + 1sat |
| Ae. speltoides Var. Ligustica | 14 | 69.35 ± 0.15 | 1.67 ± 0.19 | 5sm + 2m + 5sat |
| Ae. speltoides Taush. | 14 | 56.10 ± 0.02 | 1.47 ± 0.24 | 7m |
| Ae. triuncialis | 28 | 108.32 ± 0.01 | 3.81 ± 1.63 | 12st + 2sm |
| Ae. triaristata | 28 | 81.99 ± 0.10 | 2.17 ± 0.05 | 9sm + 5m + 1B chr |
| Ae. umbellulata | 14 | 29.21 ± 0.62 | 2.94 ± 0.08 | 3sm + 2st + 2t + 1sat |
| Mean of Square | ||||||
|---|---|---|---|---|---|---|
| Turkey | Iran | |||||
| S.O.V | D | T | H | D | T | H |
| Plant Species | 0.548 ** | 2.861 ** | 0.096 ns | 1.222 ** | 5.328 ** | 1.444 ns |
| Error | 0.119 | 0.227 | 0.104 | 0.184 | 0.593 | 0.995 |
| Species | 2n | MTCL ± Se (µm) | AR + Se | KF | DNA Content ± Se | Genome |
|---|---|---|---|---|---|---|
| Ae. biuncialis | 28 | 108.32 ± 0.63 | 2.54 ± 0.92 | 10sm + 4st + 3sat | 20.23 ± 0.35 | UUMM |
| Ae. olumnaris | 28 | 103 ± 0.21 | 2.12 ± 0.29 | 10sm + 3m + 1st + 2sat | 21.75 ± 0.36 | UUMM |
| Ae. crassa (4x) | 28 | 156 ± 0.21 | 1.46 ± 0.03 | 11m + 3sm + 2sat | 20.44 ± 0.29 | DDMM |
| Ae. crassa (6x) | 42 | 217.39 ± 0.14 | 1.36 ± 0.03 | 20m + 1sm + 3sat | 33.97 ± 0.06 | DDMM |
| Ae. cylindrica (4x) | 28 | 105.97 ± 0.19 | 2.37 ± 0.53 | 8sm + 5st + 1m + 1sat | 18.61 ± 0.07 | CCDD |
| Ae. cylindrica (6x) | 42 | 237 ± 0.12 | 2.29 ± 0.19 | 12sm + 8st + 2m + 2sat | 32.58 ± 0.25 | CCDD |
| Ae. speltoides var. Aucheri | 14 | 56.22 ± 0.37 | 1.37 ± 0.04 | 6m + 1sm + 1sat | 10.22 ± 0.02 | SS |
| Ae. speltoides var. Ligustica | 14 | 82.12 ± 0.17 | 1.52 ± 0.22 | 5sm + 2m + 5sat | 11.56 ± 0.01 | SS |
| Ae. speltoides var. Taush. | 14 | 63.18 ± 0.33 | 1.38 ± 0.18 | 7m | 10.20 ± 0.36 | SS |
| Ae. triuncialis | 28 | 108.52 ± 0.16 | 2.45 ± 0.91 | 14st | 19.27 ± 0.04 | UUCC |
| Ae. umbellulata | 28 | 25.95 ± 0.09 | 2.84 ± 0.17 | 5st + 1sm + 1t + 1sat | 10.58 ± 0.56 | UU |
| Species | 2n | MTCL ± Se (µm) | AR ± Se | KF | DNA Content ± Se | Genome |
|---|---|---|---|---|---|---|
| Ae. biuncialis | 28 | 98.69 ± 0.61 | 2.15 ± 0.69 | 13sm + 1m + 3sat | 19.89 ± 0.29 | UUMM |
| Ae. columnaris | 28 | 103.32 ± 0.79 | 2.12 ± 1.17 | 10sm + 3m + 1st + 2sat | 20.17 ± 0.16 | UUMM |
| Ae. crassa (4x) | 28 | 153.35 ± 0.49 | 1.61 ± 0.05 | 11m + 3sm + 2sat | 20.08 ± 0.01 | DDMM |
| Ae. crassa (6x) | 42 | 206.11 ± 0.03 | 1.13 ± 0.04 | 10m + 1sm + 3sat | 31.59 ± 0.03 | DDMM |
| Ae. cylindrica (4x) | 28 | 81.57 ± 0.10 | 2.39 ± 0.21 | 8sm + 5st + 1m + 1sat | 18.09 ± 0.05 | CCDD |
| Ae. cylindrica (6x) | 42 | 245 ± 0.08 | 2.30 ± 0.12 | 12sm + 8st + 2m + 2sat | 31.81 ± 0.08 | CCDD |
| Ae. speltoides var. Aucheri | 14 | 52 ± 0.12 | 1.31 ± 0.09 | 7m + 1sat | 10.15 ± 0.04 | SS |
| Ae. speltoides var. Ligustica | 14 | 69.35 ± 0.15 | 1.67 ± 0.19 | 5sm + 2m + 5sat | 10.95 ± 0.12 | SS |
| Ae. speltoides Taush. | 14 | 56.10 ± 0.02 | 1.47 ± 0.24 | 7m | 10.16 ± 0.27 | SS |
| Ae. triuncialis | 28 | 108.32 ± 0.01 | 3.81 ± 1.63 | 12st + 2sm | 19.05 ± 0.03 | UUCC |
| Ae. triaristata | 28 | 81.99 ± 0.10 | 2.17 ± 0.05 | 9sm + 5m + 1B chr | 21.65 ± 0.05 | UUMM |
| Ae. umbellulata | 14 | 29.21 ± 0.62 | 2.94 ± 0.08 | 3sm + 2st + 2t + 1sat | 10.09 ± 0.04 | UU |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najafi, S.; Ulker, M.; Oral, E.; Tuncturk, R.; Tuncturk, M.; Sayyed, R.Z.; Perveen, K.; Poczai, P.; Cseh, A. Estimation of Nuclear DNA Content in Some Aegilops Species: Best Analyzed Using Flow Cytometry. Genes 2022, 13, 1980. https://doi.org/10.3390/genes13111980
Najafi S, Ulker M, Oral E, Tuncturk R, Tuncturk M, Sayyed RZ, Perveen K, Poczai P, Cseh A. Estimation of Nuclear DNA Content in Some Aegilops Species: Best Analyzed Using Flow Cytometry. Genes. 2022; 13(11):1980. https://doi.org/10.3390/genes13111980
Chicago/Turabian StyleNajafi, Solmaz, Mehmet Ulker, Erol Oral, Ruveyde Tuncturk, Murat Tuncturk, R. Z. Sayyed, Kahkashan Perveen, Peter Poczai, and Andras Cseh. 2022. "Estimation of Nuclear DNA Content in Some Aegilops Species: Best Analyzed Using Flow Cytometry" Genes 13, no. 11: 1980. https://doi.org/10.3390/genes13111980