
Waterborne and Dietary Bioaccumulation of Organophosphate Esters in Zooplankton Daphnia magna


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Organisms
2.2. S. obliquus Exposure to OPEs
2.3. D. magna Waterborne Exposure to OPEs
2.4. D. magna Dietary Exposure to OPEs
2.5. Sample Preparation
2.6. Chemical Analysis and Quality Assurance and Control
2.7. Toxicokinetics Model
2.8. Statistical Analysis
3. Results
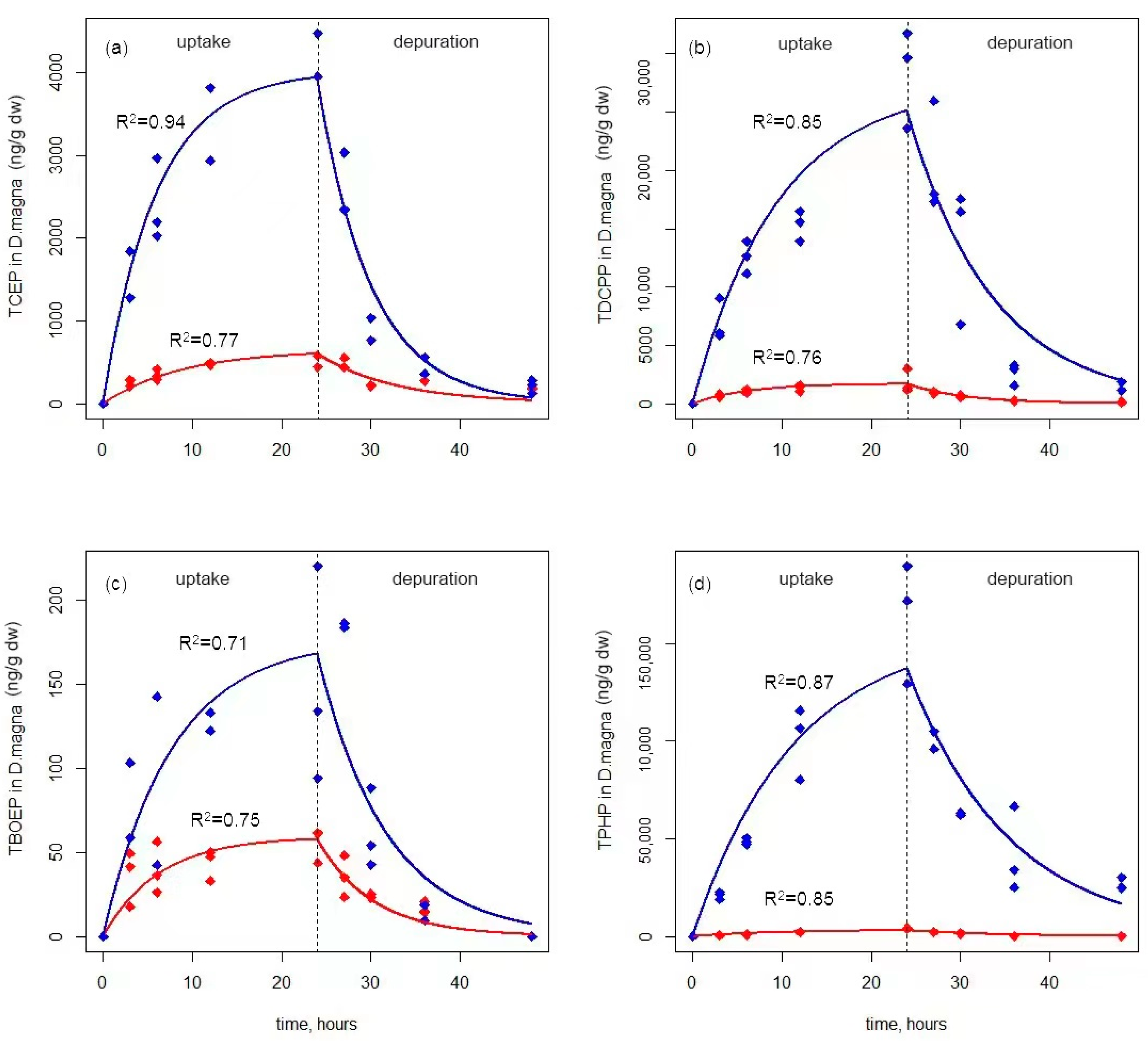
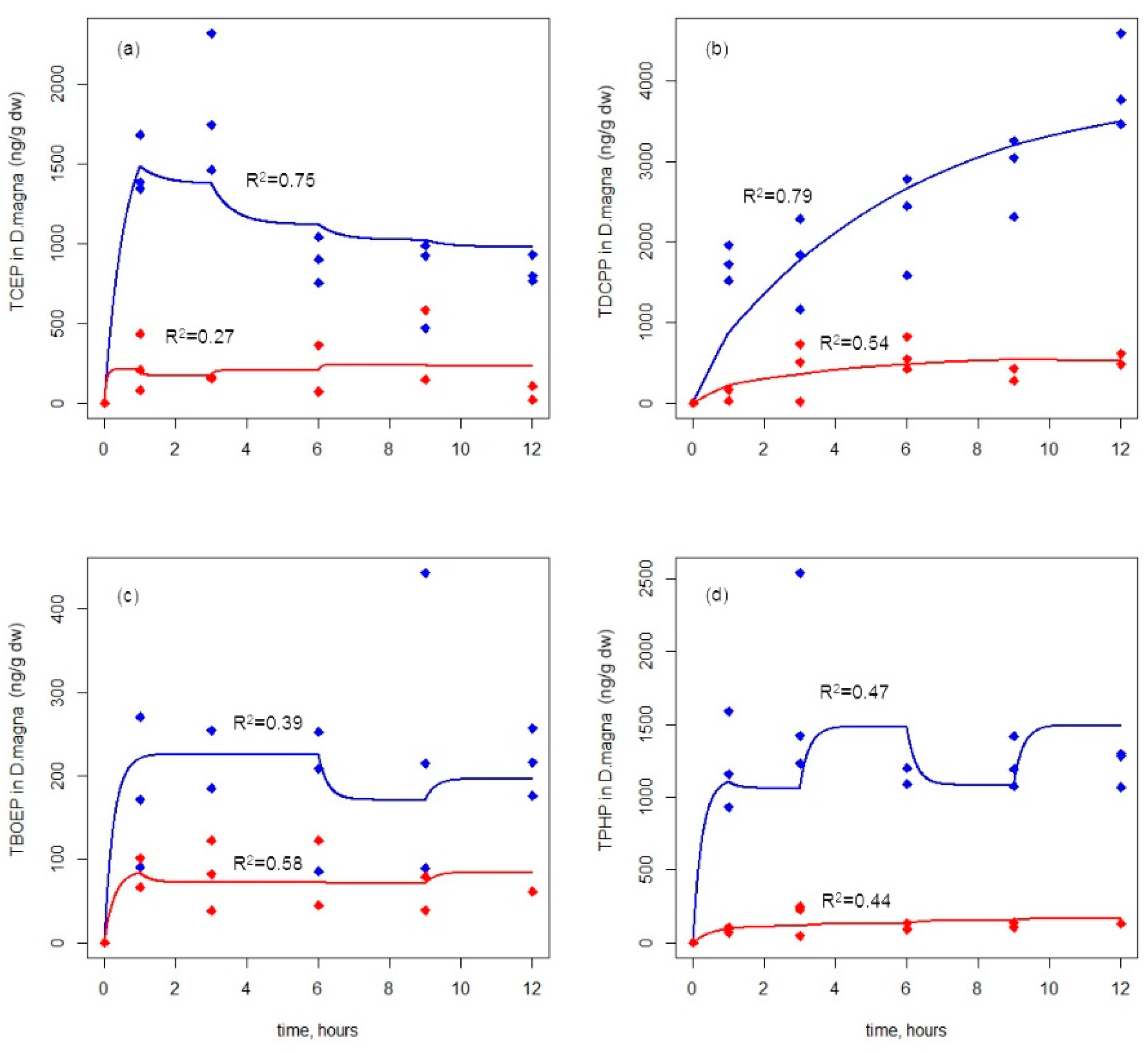
3.1. Accumulation of OPEs in D. magna via Waterborne Route
3.2. Accumulation of OPEs in D. magna via Dietary Route
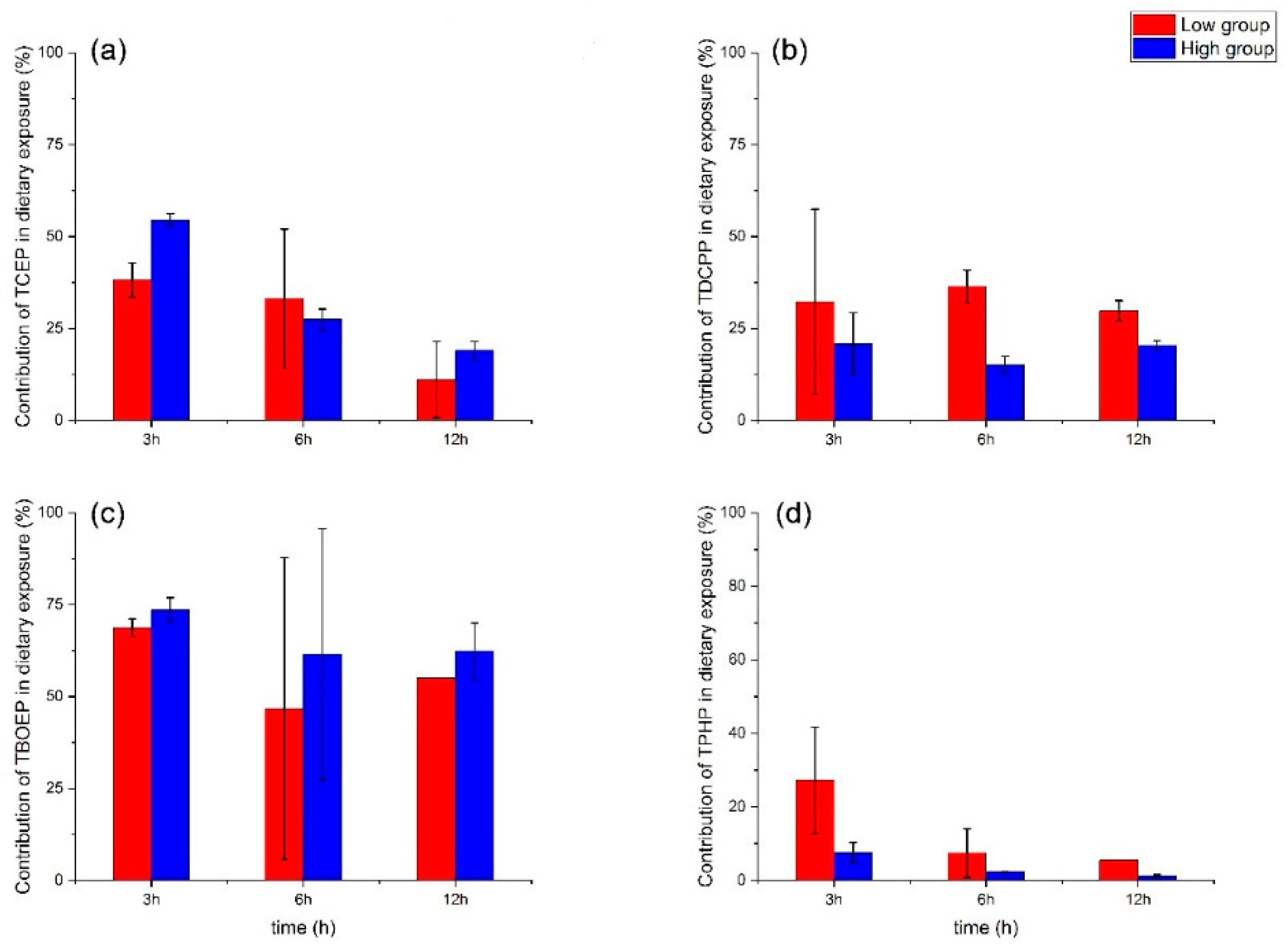
3.3. Relative Importance of Waterborne and Dietary Routes to the Accumulation of OPEs in D. magna
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- van der Veen, I.; de Boer, J. Phosphorus flame retardants: Properties, production, environmental occurrence, toxicity and analysis. Chemosphere 2012, 88, 1119–1153. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhao, F.; Liu, J.; Zhang, S.; Mu, D.; An, L.; Wan, Y.; Hu, J. Trophic transfer of organophosphorus flame retardants in a lake food web. Environ. Pollut. 2018, 242, 1887–1893. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhong, W.; Xiao, B.; Liu, Q.; Yang, L.; Covaci, A.; Zhu, L. Bioavailability and biomagnification of organophosphate esters in the food web of Taihu Lake, China: Impacts of chemical properties and metabolism. Environ. Int. 2019, 125, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Song, N.; Guo, R.; Xu, H.; Zhang, Q.; Han, Z.; Feng, M.; Li, D.; Zhang, S.; Chen, J. Occurrence and partitioning behavior of organophosphate esters in surface water and sediment of a shallow Chinese freshwater lake (Taihu Lake): Implication for eco-toxicity risk. Chemosphere 2018, 202, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.E.; Luo, X.J.; Zapata Corella, P.; Zeng, Y.H.; Mai, B.X. Organophosphorus flame retardants in a typical freshwater food web: Bioaccumulation factors, tissue distribution, and trophic transfer. Environ. Pollut. 2019, 255, 113286. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Tao, M.; Zhang, Q.; Kong, M.; Sun, J.; Jia, S.; Liu, C.H. Occurrence, spatial distribution and risk assessment of organophosphate esters in surface water from the lower Yangtze River Basin. Sci. Total Environ. 2020, 734, 139380. [Google Scholar] [CrossRef]
- Bekele, T.G.; Zhao, H.; Wang, Q.; Chen, J. Bioaccumulation and Trophic Transfer of Emerging Organophosphate Flame Retardants in the Marine Food Webs of Laizhou Bay, North China. Environ. Sci. Technol. 2019, 53, 13417–13426. [Google Scholar] [CrossRef] [PubMed]
- Bekele, T.G.; Zhao, H.; Wang, Q. Tissue distribution and bioaccumulation of organophosphate esters in wild marine fish from Laizhou Bay, North China: Implications of human exposure via fish consumption. J. Hazard. Mater. 2021, 401, 123410. [Google Scholar] [CrossRef]
- Sala, B.; Gimenez, J.; de Stephanis, R.; Barcelo, D.; Eljarrat, E. First determination of high levels of organophosphorus flame retardants and plasticizers in dolphins from Southern European waters. Environ. Res. 2019, 172, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Kim, U.-J.; Kannan, K. Occurrence and Distribution of Organophosphate Flame Retardants/Plasticizers in Surface Waters, Tap Water, and Rainwater: Implications for Human Exposure. Environ. Sci. Technol. 2018, 52, 5625–5633. [Google Scholar] [CrossRef] [PubMed]
- Pantelaki, I.; Voutsa, D. Organophosphate esters in inland and coastal waters in northern Greece. Sci. Total Environ. 2021, 800, 149544. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Fu, K.; Gao, K.; Li, H.; Xue, Q.; Chen, Y.; Wang, L.; Shi, J.; Fu, J.; Zhang, Q.; et al. Occurrence and Trophic Magnification of Organophosphate Esters in an Antarctic Ecosystem: Insights into the Shift from Legacy to Emerging Pollutants. J. Hazard. Mater. 2020, 396, 122742. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Lam, J.C.; Man, Y.C.; Lai, N.L.; Kwok, K.Y.; Guo, Y.; Lam, P.K.; Zhou, B. Bioconcentration, metabolism and neurotoxicity of the organophorous flame retardant 1,3-dichloro 2-propyl phosphate (TDCPP) to zebrafish. Aquat. Toxicol. 2015, 158, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Hong, X.; Chen, R.; Hou, R.; Yuan, L.; Zha, J. Triphenyl Phosphate (TPHP)-Induced Neurotoxicity in Adult Male Chinese Rare Minnows (Gobiocypris rarus). Environ. Sci. Technol. 2018, 52, 11895–11903. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Cai, Y.; Wang, Y.; Xu, S.; Ji, K.; Choi, K. Effects of tris(1,3-dichloro-2-propyl) phosphate (TDCPP) and triphenyl phosphate (TPP) on sex-dependent alterations of thyroid hormones in adult zebrafish. Ecotoxicol. Environ. Saf. 2019, 170, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Tsui, M.M.P.; Hu, C.; Lam, J.C.W.; Zhou, B.; Chen, L. Acute exposure to triphenyl phosphate (TPhP) disturbs ocular development and muscular organization in zebrafish larvae. Ecotoxicol. Environ. Saf. 2019, 179, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.L.; Li, D.Q.; Zhuo, M.N.; Liao, Y.S.; Xie, Z.Y.; Guo, T.L.; Li, J.J.; Zhang, S.Y.; Liang, Z.Q. Organophosphorus flame retardants and plasticizers: Sources, occurrence, toxicity and human exposure. Environ. Pollut. 2015, 196, 29–46. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, N.; Wu, Y.; Zhang, L. The simultaneous uptake of dietary and waterborne Cd in gastrointestinal tracts of marine yellowstripe goby Mugilogobius chulae. Environ. Pollut. 2017, 223, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Cui, Y.Q.; Guo, W.B.; Yang, L.; Miao, A.J. Waterborne and dietary accumulation of well-dispersible hematite nanoparticles by zebrafish at different life stages. Environ. Pollut. 2020, 259, 113852. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Bortvedt, A.; Harper, B.J.; Crandon, L.E.; Harper, S.L. Uptake and toxicity of CuO nanoparticles to Daphnia magna varies between indirect dietary and direct waterborne exposures. Aquat. Toxicol. 2017, 190, 78–86. [Google Scholar] [CrossRef] [PubMed]
- de Wit, H.A.; Kainz, M.J.; Lindholm, M. Methylmercury bioaccumulation in invertebrates of boreal streams in Norway: Effects of aqueous methylmercury and diet retention. Environ. Pollut. 2012, 164, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, Y.; Li, J.; Zhou, D.; Guo, R.; Ji, R.; Chen, J. The bioaccumulation, elimination, and trophic transfer of BDE-47 in the aquatic food chain of Chlorella pyrenoidosa-Daphnia magna. Environ. Pollut. 2020, 258, 113720. [Google Scholar] [CrossRef]
- Wang, C.; Chang, X.L.; Shi, Q.; Zhang, X. Uptake and Transfer of (13)C-Fullerenols from Scenedesmus obliquus to Daphnia magna in an Aquatic Environment. Environ. Sci. Technol. 2018, 52, 12133–12141. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, X.; Lim, D.J.; Laserna, A.K.C.; Li, S.F.Y. Uptake and toxic effects of triphenyl phosphate on freshwater microalgae Chlorella vulgaris and Scenedesmus obliquus: Insights from untargeted metabolomics. Sci. Total Environ. 2019, 650, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, Y.; Yang, K.; Zhu, L.; Lin, D. Nanoparticle TiO2 size and rutile content impact bioconcentration and biomagnification from algae to daphnia. Environ. Pollut. 2019, 247, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yuan, S.; Su, G.; Li, M.; Wang, Q.; Zhu, G.; Letcher, R.J.; Li, Y.; Han, Z.; Liu, C. Whole-Life-Stage Characterization in the Basic Biology of Daphnia magna and Effects of TDCIPP on Growth, Reproduction, Survival, and Transcription of Genes. Environ. Sci. Technol. 2017, 51, 13967–13975. [Google Scholar] [CrossRef]
- Li, W.; Yuan, S.; Sun, Q.; Liu, C. Toxicity of tris(2-chloroethyl) phosphate in Daphnia magna after lifetime exposure: Changes in growth, reproduction, survival and gene transcription. Ecotoxicol. Environ. Saf. 2020, 200, 110769. [Google Scholar] [CrossRef]
- Giraudo, M.; Douville, M.; Houde, M. Chronic toxicity evaluation of the flame retardant tris (2-butoxyethyl) phosphate (TBOEP) using Daphnia magna transcriptomic response. Chemosphere 2015, 132, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, H.; Dang, Y.; Liu, C. Effects of triphenyl phosphate on growth, reproduction and transcription of genes of Daphnia magna. Aquat. Toxicol. 2018, 195, 58–66. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Yang, R.; Yang, L.; Sun, B.; Zhu, L. Uptake Kinetics, Accumulation, and Long-Distance Transport of Organophosphate Esters in Plants: Impacts of Chemical and Plant Properties. Environ. Sci. Technol. 2019, 53, 4940–4947. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, J.; Zhu, L. Prediction of acute toxicity of cadmium and lead to zebrafish larvae by using a refined toxicokinetic-toxicodynamic model. Aquat. Toxicol. 2015, 169, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Gao, Y.; Bian, X.; Feng, J.; Ma, W.; Zhu, L. Predicting the survival of zebrafish larvae exposed to fluctuating pulses of lead and cadmium. Chemosphere 2019, 223, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.X. Uptake, absorption efficiency and elimination of DDT in marine phytoplankton, copepods and fish. Environ. Pollut. 2005, 136, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Hansman, R.L.; Metian, M.; Pouil, S.; Oberhansli, F.; Teyssie, J.L.; Swarzenski, P.W. A double-tracer radioisotope approach to assess simultaneous bioaccumulation of caesium in the olive flounder Paralichthys olivaceus. J. Environ. Radioact. 2018, 190–191, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Bekele, T.G.; Zhao, H.; Wang, Y.; Jiang, J.; Tan, F. Measurement and prediction of bioconcentration factors of organophosphate flame retardants in common carp (Cyprinus carpio). Ecotoxicol. Environ. Saf. 2018, 166, 270–276. [Google Scholar] [CrossRef]
- Wan, W.; Huang, H.; Lv, J.; Han, R.; Zhang, S. Uptake, Translocation, and Biotransformation of Organophosphorus Esters in Wheat (Triticum aestivum L.). Environ. Sci. Technol. 2017, 51, 13649–13658. [Google Scholar] [CrossRef] [PubMed]
- Arukwe, A.; Carteny, C.C.; Eggen, T.; Moder, M. Novel aspects of uptake patterns, metabolite formation and toxicological responses in Salmon exposed to the organophosphate esters-Tris(2-butoxyethyl)- and tris(2-chloroethyl) phosphate. Aquat. Toxicol. 2018, 196, 146–153. [Google Scholar] [CrossRef]
- Wang, G.; Shi, H.; Du, Z.; Chen, H.; Peng, J.; Gao, S. Bioaccumulation mechanism of organophosphate esters in adult zebrafish (Danio rerio). Environ Pollut 2017, 229, 177–187. [Google Scholar] [CrossRef]
- Tang, B.; Poma, G.; Bastiaensen, M.; Yin, S.S.; Luo, X.J.; Mai, B.X.; Covaci, A. Bioconcentration and biotransformation of organophosphorus flame retardants (PFRs) in common carp (Cyprinus carpio). Environ. Int. 2019, 126, 512–522. [Google Scholar] [CrossRef]
- Mata, M.C.; Castro, V.; Quintana, J.B.; Rodil, R.; Beiras, R.; Vidal-Linan, L. Bioaccumulation of organophosphorus flame retardants in the marine mussel Mytilus galloprovincialis. Sci. Total Environ. 2022, 805, 150384. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, L.; Wang, W.X. Bioaccumulation and trophic transfer of dioxins in marine copepods and fish. Environ. Pollut. 2011, 159, 3390–3397. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Jeon, J.; Choi, Y.; Kim, S.D. Characterizing biotransformation products and pathways of the flame retardant triphenyl phosphate in Daphnia magna using non-target screening. Sci. Total Environ. 2020, 708, 135106. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Qing, X.; Wang, J.; He, T.; Fan, R.; Huang, Y. Bioaccumulation and potential risk of organophosphate flame retardants in coral reef fish from the Nansha Islands, South China Sea. Chemosphere 2022, 287, 132125. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Du, Y.; Zheng, F.; Wang, J.; Wang, Z.; Ji, R.; Chen, J. Bioaccumulation and elimination of bisphenol a (BPA) in the alga Chlorella pyrenoidosa and the potential for trophic transfer to the rotifer Brachionus calyciflorus. Environ. Pollut. 2017, 227, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Hou, R.; Xu, Y.; Wang, Z. Review of OPFRs in animals and humans: Absorption, bioaccumulation, metabolism, and internal exposure research. Chemosphere 2016, 153, 78–90. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Measured Exposure Concentration (μg/L) | Waterborne Exposure | Dietary Exposure | ||||||
|---|---|---|---|---|---|---|---|---|
| ku1 (L kg−1 h−1) | kd1 (h−1) | t 1/2 (h) | BCF (L kg−1) | ku2 (h−1) | kd2 (h−1) | BMF | ||
| TCEP | 21.72 | 3.39 | 0.12 | 6.03 | 29.48 | 5.82 | 16.41 | 0.35 |
| 116.67 | 5.78 | 0.17 | 4.13 | 34.40 | 1.05 | 1.71 | 0.61 | |
| TDCPP | 17.07 | 16.04 | 0.16 | 4.36 | 100.86 | 0.06 | 0.38 | 0.17 |
| 124.22 | 23.22 | 0.11 | 6.60 | 221.14 | 0.03 | 0.15 | 0.22 | |
| TBOEP | 16.71 | 0.57 | 0.16 | 4.28 | 3.52 | 0.53 | 3.59 | 0.15 |
| 135.67 | 0.17 | 0.13 | 5.33 | 1.29 | 0.25 | 3.82 | 0.07 | |
| TPHP | 16.29 | 21.15 | 0.10 | 6.66 | 203.37 | 0.09 | 2.05 | 0.04 |
| 99.06 | 138.80 | 0.09 | 7.88 | 1577.27 | 0.29 | 3.88 | 0.07 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Zhang, H.; Ding, J.; He, W.; Zhu, L.; Feng, J. Waterborne and Dietary Bioaccumulation of Organophosphate Esters in Zooplankton Daphnia magna. Int. J. Environ. Res. Public Health 2022, 19, 9382. https://doi.org/10.3390/ijerph19159382
Liu W, Zhang H, Ding J, He W, Zhu L, Feng J. Waterborne and Dietary Bioaccumulation of Organophosphate Esters in Zooplankton Daphnia magna. International Journal of Environmental Research and Public Health. 2022; 19(15):9382. https://doi.org/10.3390/ijerph19159382
Chicago/Turabian StyleLiu, Wenxin, Hong Zhang, Jiaqi Ding, Wanyu He, Lin Zhu, and Jianfeng Feng. 2022. "Waterborne and Dietary Bioaccumulation of Organophosphate Esters in Zooplankton Daphnia magna" International Journal of Environmental Research and Public Health 19, no. 15: 9382. https://doi.org/10.3390/ijerph19159382