Prostaglandin D2-Mediated DP2 and AKT Signal Regulate the Activation of Androgen Receptors in Human Dermal Papilla Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
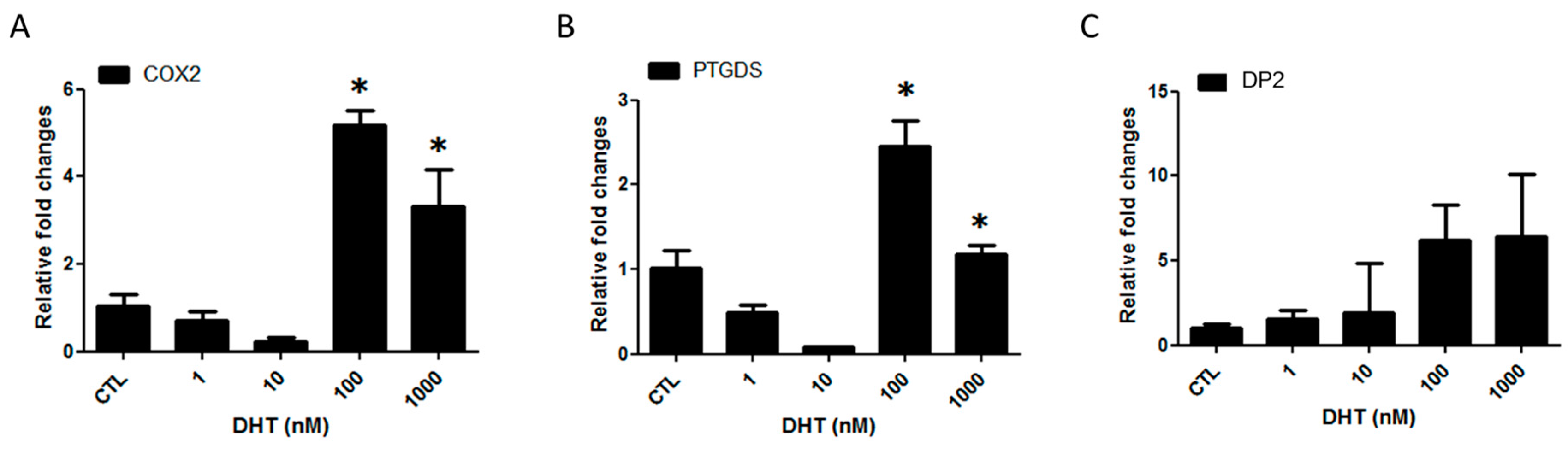
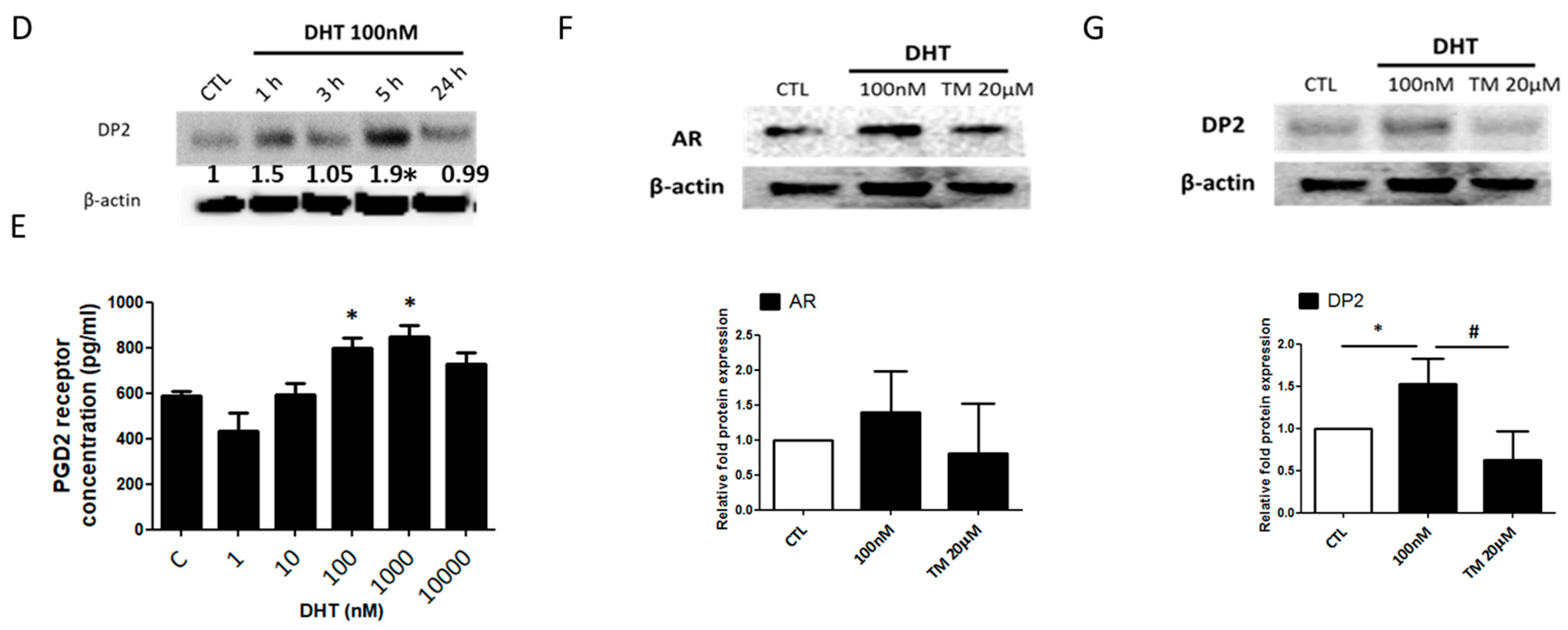
2.1. Prostaglandin D2 Receptor 2 (DP2) Antagonist Regulates Dihydrotestosterone (DHT)-Induced Prostaglandin D2 (PGD2) Pathway
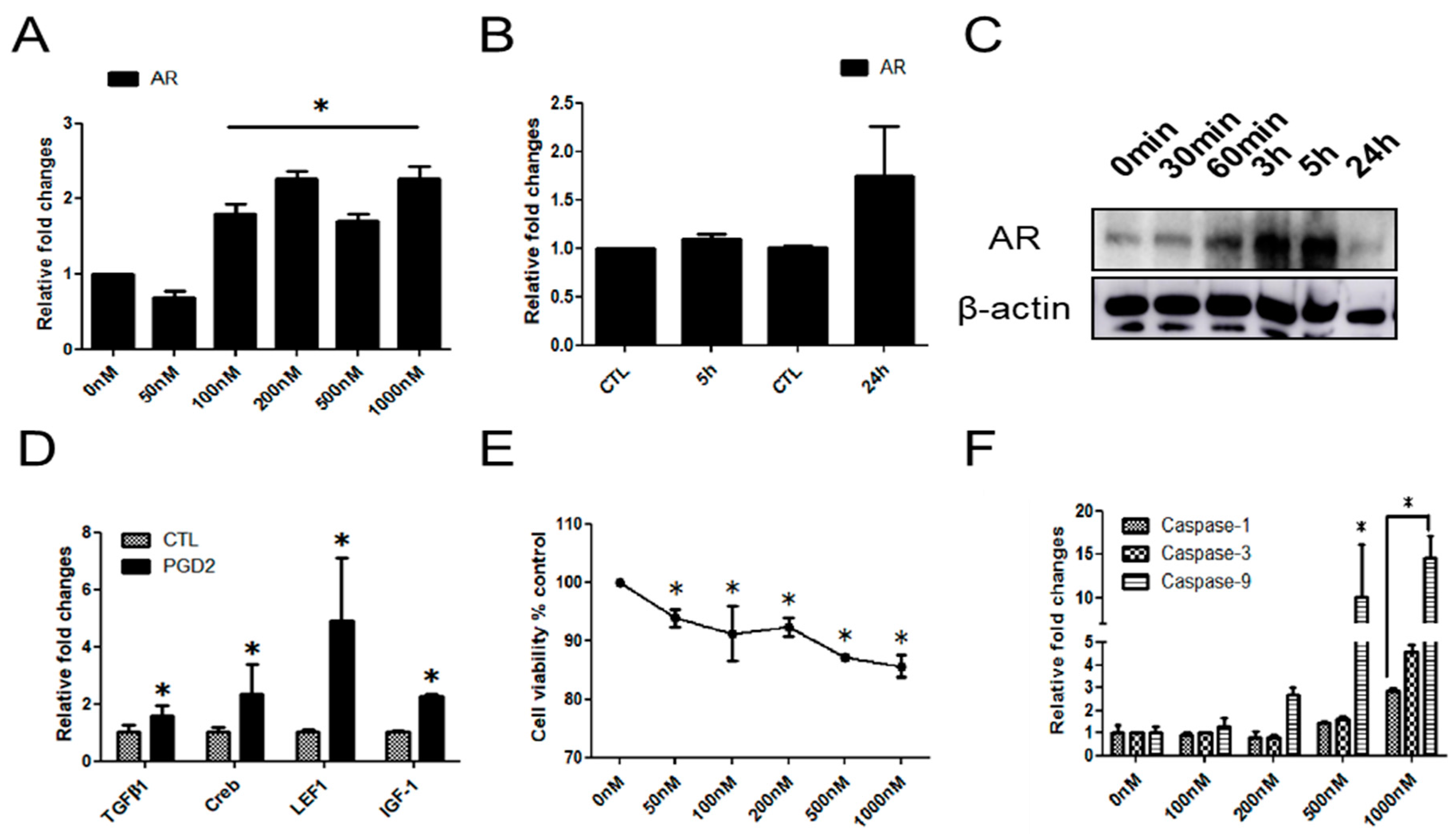
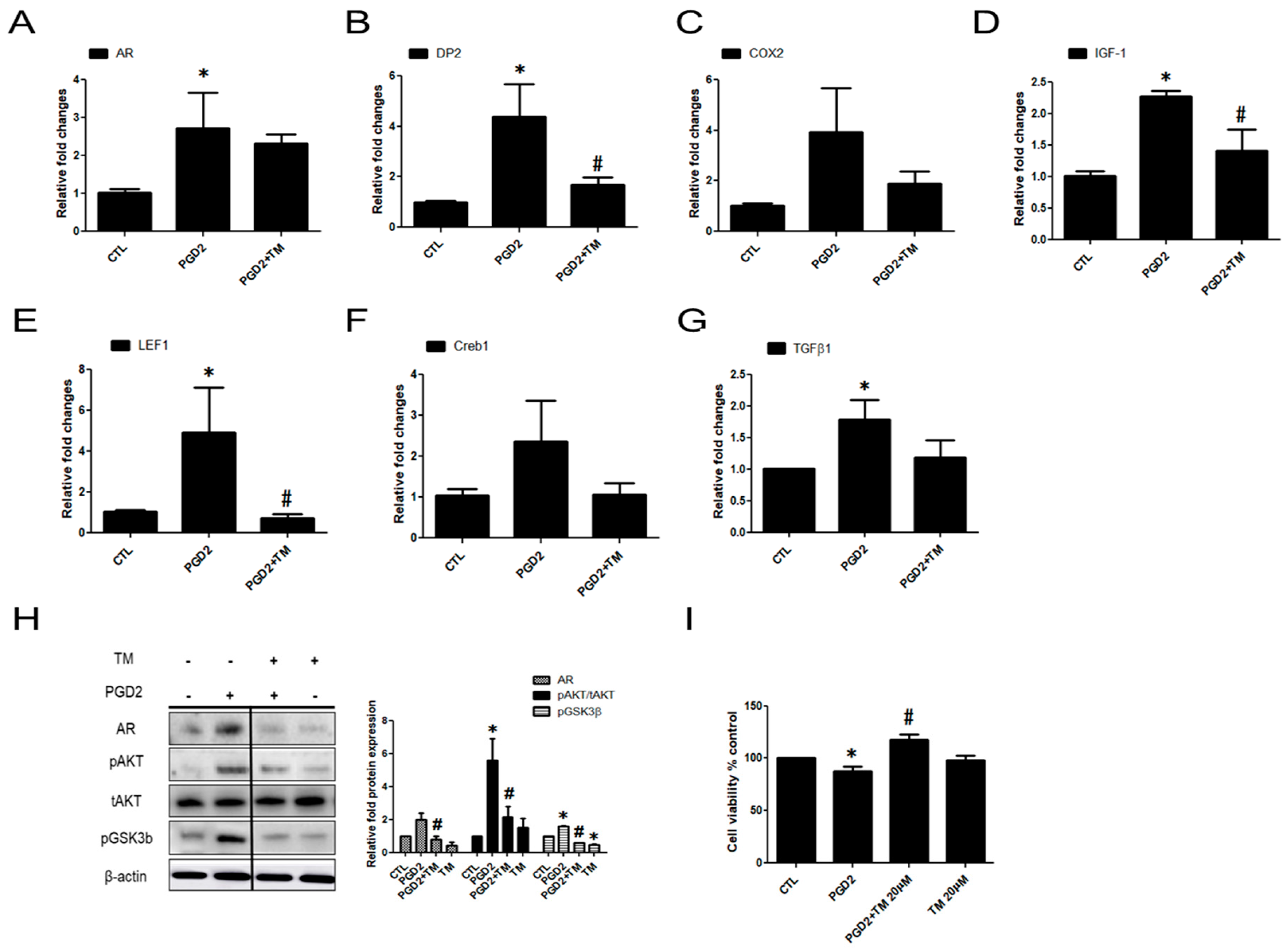
2.2. The Effects of PGD2 on AR Expression and hDPCs
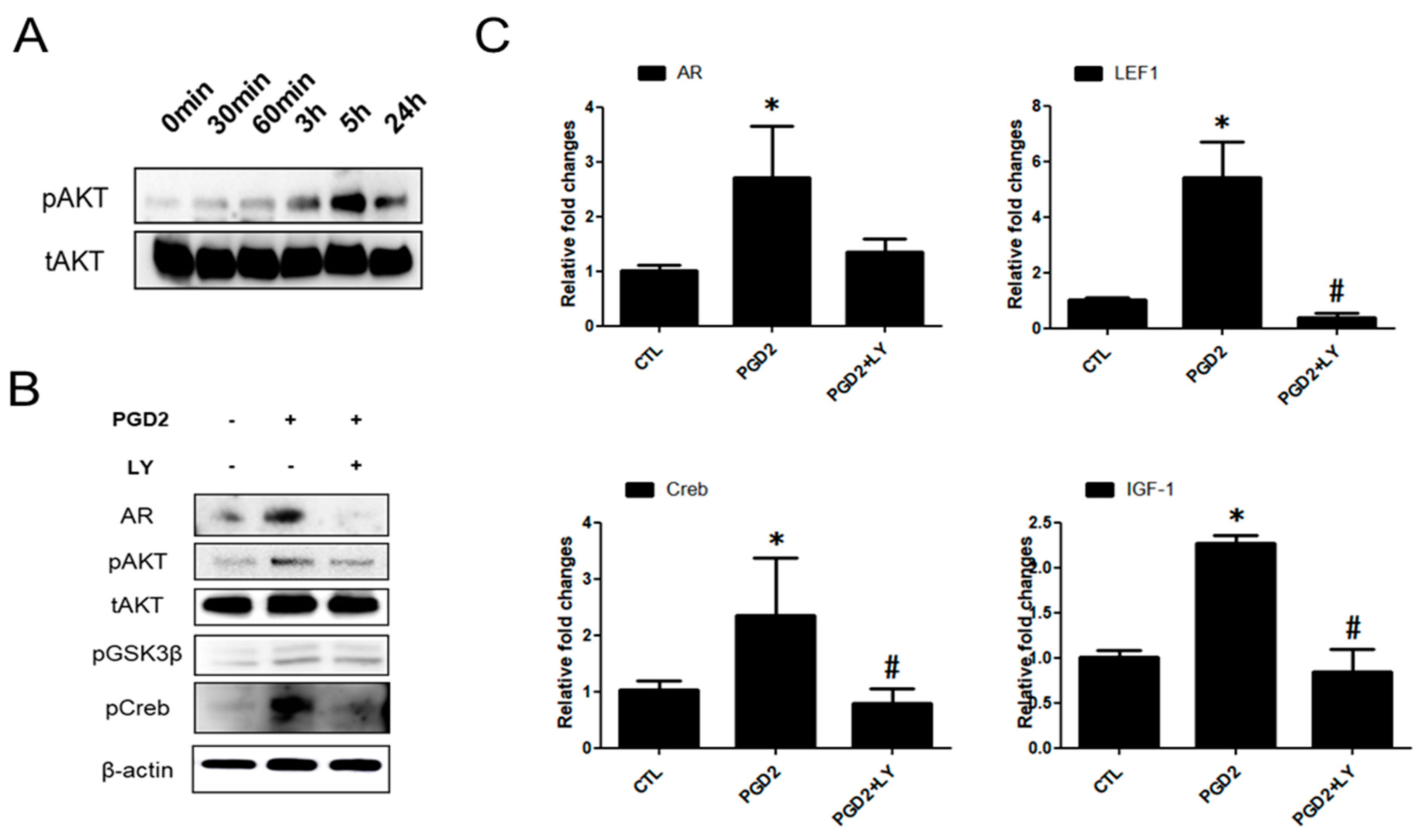
2.3. PGD2-Induced AR Expression is Regulated by AKT Signalling
2.4. PGD2-Induced AR Expression and AKT Signalling Are Regulated by a DP2 Antagonist
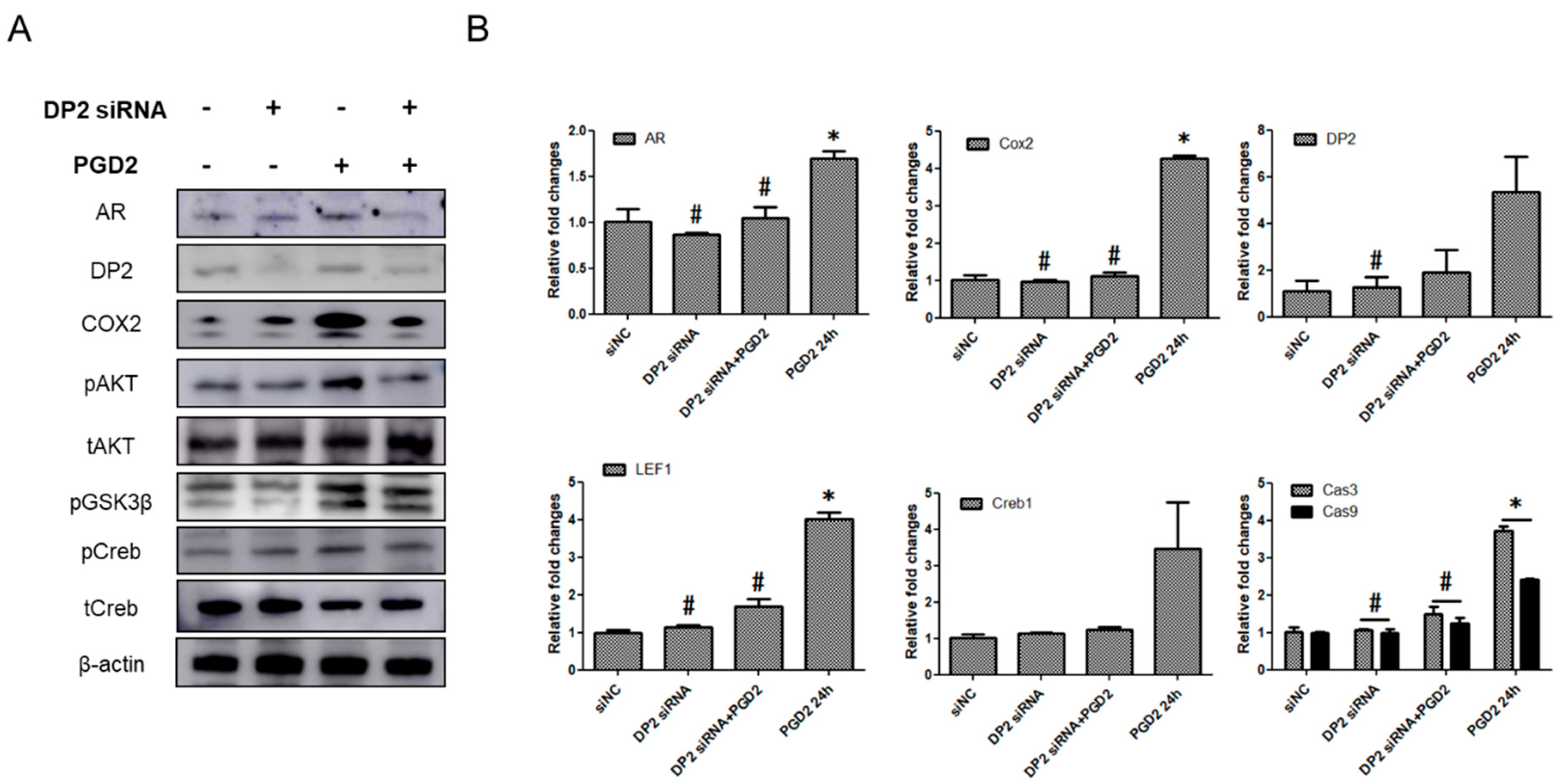
2.5. The Functions of DP2 on PGD2-Induced AR Expression
3. Discussion
4. Materials and Methods
4.1. Human Dermal Papilla Cell (hDPCs) Culture and Reagents
4.2. Cell Viability Assay
4.3. Real Time-PCR (qRT-PCR)
4.4. Western Blotting Analysis
4.5. Immunofluoresence of Androgen Receptor and DP2
4.6. ELISA
4.7. DP2 Gene Silencing Experiments
4.8. TUNEL Assay
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sinclair, R. Male pattern androgenetic alopecia. BMJ 1998, 317, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, K.D. Androgens and alopecia. Mol. Cell. Endocrinol. 2002, 198, 89–95. [Google Scholar] [CrossRef]
- Sawaya, M.E.; Keane, R.W.; Blume-Peytavi, U.; Mullins, D.L.; Nusbaum, B.P.; Whiting, D.; Nicholson, D.W. Androgen responsive genes as they affect hair growth. Eur. J. Dermatol. 2001, 11, 304–308. [Google Scholar] [PubMed]
- Magro, C.M.; Rossi, A.; Poe, J.; Manhas-Bhutani, S.; Sadick, N. The role of inflammation and immunity in the pathogenesis of androgenetic alopecia. J. Drugs Dermatol. 2011, 10, 1404–1411. [Google Scholar] [PubMed]
- Mahe, Y.F.; Michelet, J.F.; Billoni, N.; Jarrousse, F.; Buan, B.; Commo, S.; Saint-Leger, D.; Bernard, B.A. Androgenetic alopecia and microinflammation. Int. J. Dermatol. 2000, 39, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Garza, L.A.; Liu, Y.; Yang, Z.; Alagesan, B.; Lawson, J.A.; Norberg, S.M.; Loy, D.E.; Zhao, T.; Blatt, H.B.; Stanton, D.C.; et al. Prostaglandin D2 inhibits hair growth and is elevated in bald scalp of men with androgenetic alopecia. Sci. Transl. Med. 2012, 4, 126ra34. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, S. Increased DHT levels in androgenic alopecia have been selected for to protect men from prostate cancer. Med. Hypotheses 2014, 82, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.M.; Loy, D.E.; Lawson, J.A.; Katseff, A.S.; Fitzgerald, G.A.; Garza, L.A. Prostaglandin D2 inhibits wound-induced hair follicle neogenesis through the receptor, Gpr44. J. Investig. Dermatol. 2013, 133, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Muller-Decker, K.; Leder, C.; Neumann, M.; Neufang, G.; Bayerl, C.; Schweizer, J.; Marks, F.; Furstenberger, G. Expression of cyclooxygenase isozymes during morphogenesis and cycling of pelage hair follicles in mouse skin: Precocious onset of the first catagen phase and alopecia upon cyclooxygenase-2 overexpression. J. Investig. Dermatol. 2003, 121, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Uller, L.; Mathiesen, J.M.; Alenmyr, L.; Korsgren, M.; Ulven, T.; Hogberg, T.; Andersson, G.; Persson, C.G.; Kostenis, E. Antagonism of the prostaglandin D2 receptor CRTH2 attenuates asthma pathology in mouse eosinophilic airway inflammation. Respir. Res. 2007, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Randall, V.A.; Hibberts, N.A.; Hamada, K. A comparison of the culture and growth of dermal papilla cells from hair follicles from non-balding and balding (androgenetic alopecia) scalp. Br. J. Dermatol. 1996, 134, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Winiarska, A.; Mandt, N.; Kamp, H.; Hossini, A.; Seltmann, H.; Zouboulis, C.C.; Blume-Peytavi, U. Effect of 5α-dihydrotestosterone and testosterone on apoptosis in human dermal papilla cells. Skin Pharmacol. Physiol. 2006, 19, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Cha, H.J.; Lim, K.M.; Lee, O.K.; Bae, S.; Kim, C.H.; Lee, K.H.; Lee, Y.N.; Ahn, K.J.; An, S. Analysis of the microRNA expression profile of normal human dermal papilla cells treated with 5α-dihydrotestosterone. Mol. Med. Rep. 2015, 12, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Inui, S.; Itami, S. Molecular basis of androgenetic alopecia: From androgen to paracrine mediators through dermal papilla. J. Dermatol. Sci. 2011, 61, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hibino, T.; Nishiyama, T. Role of TGF-β2 in the human hair cycle. J. Dermatol. Sci. 2004, 35, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, Y.; Denda, S.; Soma, T.; Raftery, L.; Momoi, T.; Hibino, T. A potential suppressor of TGF-β delays catagen progression in hair follicles. J. Investig. Dermatol. Symp. Proc. 2003, 8, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Price, V.H. Androgenetic alopecia in women. J. Investig. Dermatol. Symp. Proc. 2003, 8, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dickkopf 1 promotes regression of hair follicles. J. Investig. Dermatol. 2012, 132, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Montserrat-de la Paz, S.; Bermudez, B.; Lopez, S.; Naranjo, M.C.; Romero, Y.; Bando-Hidalgo, M.J.; Abia, R.; Muriana, F.J. Exogenous fatty acids and niacin on acute prostaglandin D2 production in human myeloid cells. J. Nutr. Biochem. 2017, 39, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Fu, M.; D’Amico, M.; Albanese, C.; Zhou, J.N.; Brownlee, M.; Lisanti, M.P.; Chatterjee, V.K.; Lazar, M.A.; Pestell, R.G. Inhibition of cellular proliferation through IκB kinase-independent and peroxisome proliferator-activated receptor γ-dependent repression of cyclin D1. Mol. Cell. Biol. 2001, 21, 3057–3070. [Google Scholar] [CrossRef] [PubMed]
- Kitz, K.; Windischhofer, W.; Leis, H.J.; Huber, E.; Kollroser, M.; Malle, E. 15-Deoxy-Δ12,14-prostaglandin J2 induces Cox-2 expression in human osteosarcoma cells through MAPK and EGFR activation involving reactive oxygen species. Free Radic. Biol. Med. 2011, 50, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef] [PubMed]
- Su, H.Y.; Hickford, J.G.; Bickerstaffe, R.; Palmer, B.R. Insulin-like growth factor 1 and hair growth. Dermatol. Online J. 1999, 5, 1. [Google Scholar] [PubMed]
- Wu, C.C.; Shyu, R.Y.; Wang, C.H.; Tsai, T.C.; Wang, L.K.; Chen, M.L.; Jiang, S.Y.; Tsai, F.M. Involvement of the prostaglandin D2 signal pathway in retinoid-inducible gene 1 (RIG1)-mediated suppression of cell invasion in testis cancer cells. Biochim. Biophys. Acta 2012, 1823, 2227–2236. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.G.; Phipps, R.P. Prostaglandin D2, its metabolite 15-d-PGJ2, and peroxisome proliferator activated receptor-γ agonists induce apoptosis in transformed, but not normal, human T lineage cells. Immunology 2002, 105, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Janicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Soma, T.; Ogo, M.; Suzuki, J.; Takahashi, T.; Hibino, T. Analysis of apoptotic cell death in human hair follicles in vivo and in vitro. J. Investig. Dermatol. 1998, 111, 948–954. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Cheng, H.; Roberts, T.M.; Zhao, J.J. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat. Rev. Drug Discov. 2009, 8, 627–644. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, D.J.; Dedhar, S.; Wu, H.; Nelson, C.C. PTEN and GSK3β: Key regulators of progression to androgen-independent prostate cancer. Oncogene 2006, 25, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Jasmin, B.J.; Gramolini, A.O.; Adatia, F.A.; Angus, L.; Boudreau-Lariviere, C.; Chan, R.Y.; Krupa, A.M.; Lunde, J.A.; Mankal, F.A.; Wu, J. Nerve-derived trophic factors and DNA elements controlling expression of genes encoding synaptic proteins in skeletal muscle fibers. Can. J. Appl. Physiol. 1998, 23, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Liao, R.S.; Ma, S.; Miao, L.; Li, R.; Yin, Y.; Raj, G.V. Androgen receptor-mediated non-genomic regulation of prostate cancer cell proliferation. Transl. Androl. Urol. 2013, 2, 187–196. [Google Scholar] [PubMed]
- Liao, X.; Thrasher, J.B.; Holzbeierlein, J.; Stanley, S.; Li, B. Glycogen synthase kinase-3β activity is required for androgen-stimulated gene expression in prostate cancer. Endocrinology 2004, 145, 2941–2949. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Haroun, S.; Parent, J.L.; de Brum-Fernandes, A.J. Prostaglandin D2 induces apoptosis of human osteoclasts through ERK1/2 and AKT signaling pathways. Bone 2014, 60, 112–121. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Ishikawa, T.; Watanabe, S. Prostaglandin D2 induces the production of human β-defensin-3 in human keratinocytes. Biochem. Pharmacol. 2010, 79, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.; Lee, Y.; Hyun, M.Y.; Choi, S.Y.; Jeong, K.H.; Park, Y.M.; Kang, H.; Park, K.Y.; Armstrong, C.A.; Johnson, A.; et al. Mycophenolate antagonizes IFN-γ-induced catagen-like changes via β-catenin activation in human dermal papilla cells and hair follicles. Int. J. Mol. Sci. 2014, 15, 16800–16815. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.S.; Park, E.K.; Kwack, M.H.; Sung, Y.K. Effects of dexamethasone, a synthetic glucocorticoid, on human periodontal ligament stem cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 991–995. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, K.H.; Jung, J.H.; Kim, J.E.; Kang, H. Prostaglandin D2-Mediated DP2 and AKT Signal Regulate the Activation of Androgen Receptors in Human Dermal Papilla Cells. Int. J. Mol. Sci. 2018, 19, 556. https://doi.org/10.3390/ijms19020556
Jeong KH, Jung JH, Kim JE, Kang H. Prostaglandin D2-Mediated DP2 and AKT Signal Regulate the Activation of Androgen Receptors in Human Dermal Papilla Cells. International Journal of Molecular Sciences. 2018; 19(2):556. https://doi.org/10.3390/ijms19020556
Chicago/Turabian StyleJeong, Kwan Ho, Ji Hee Jung, Jung Eun Kim, and Hoon Kang. 2018. "Prostaglandin D2-Mediated DP2 and AKT Signal Regulate the Activation of Androgen Receptors in Human Dermal Papilla Cells" International Journal of Molecular Sciences 19, no. 2: 556. https://doi.org/10.3390/ijms19020556