Feeding the Walls: How Does Nutrient Availability Regulate Cell Wall Composition?
by
 , ,
, ,
Michael Ogden
1,2 ,
,
Rainer Hoefgen
2,
Ute Roessner
1,
Staffan Persson
1 and
Ghazanfar Abbas Khan
1,* 1
School of Biosciences, University of Melbourne, Victoria 3010, Australia
2
Max Planck Institute of Molecular Plant Physiology, Potsdam-Golm 14476, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(9), 2691; https://doi.org/10.3390/ijms19092691
Submission received: 15 July 2018
/
Revised: 27 August 2018
/
Accepted: 29 August 2018
/
Published: 10 September 2018
(This article belongs to the Special Issue Plant Cell Wall Proteins and Development)
{kind=link}
{kind=link}
Abstract
:Nutrients are critical for plants to grow and develop, and nutrient depletion severely affects crop yield. In order to optimize nutrient acquisition, plants adapt their growth and root architecture. Changes in growth are determined by modifications in the cell walls surrounding every plant cell. The plant cell wall, which is largely composed of complex polysaccharides, is essential for plants to attain their shape and to protect cells against the environment. Within the cell wall, cellulose strands form microfibrils that act as a framework for other wall components, including hemicelluloses, pectins, proteins, and, in some cases, callose, lignin, and suberin. Cell wall composition varies, depending on cell and tissue type. It is governed by synthesis, deposition and remodeling of wall components, and determines the physical and structural properties of the cell wall. How nutrient status affects cell wall synthesis and organization, and thus plant growth and morphology, remains poorly understood. In this review, we aim to summarize and synthesize research on the adaptation of root cell walls in response to nutrient availability and the potential role of cell walls in nutrient sensing.
1. Introduction
Plants are the primary producers on earth, accounting for 80% of all living biomass, and provide us with food, feed, and shelter [1]. Nutrient availability controls how plants grow and thus their ability to produce biomass [2]. The levels of macro- and micro-nutrients in the soil, and the ability of plants to access them, therefore have major implications on agriculture and ecology [3]. To maximize acquisition of nutrients, plants adjust their growth and metabolic processes [4,5,6]. By deliberately limiting their chlorophyll content and rate of photosynthetic carbon fixation to coordinate their growth with resource availability, plants achieve optimal morphology to survive [3]. Cell division and cell elongation are the major factors that drive growth and morphology in plants. Unlike animal cells, plant cells are surrounded by a protective and supportive structure, referred to as the cell wall, which contributes the bulk of a plant’s biomass. Dynamic remodeling of this structure is crucial for plant cells to divide and elongate [7,8]. Because the cell wall dictates cell and tissue morphology, and as nutrient availability drives changes in plant growth, it follows that changes to the cell wall structure should, indirectly or directly, be controlled by the nutrients available to the plant. There is mounting evidence from gene expression data that cell walls are actively regulated in response to nutrient availability [9,10,11,12,13]. However, while it is clear that reprogramming of cell wall genes is essential for plant adaptations to nutrient status, knowledge relating to molecular mechanisms controlling these changes is just starting to emerge. In the agricultural context, plant biomass and nutrient availability are the two most important variables. Research in this field is critically important and has direct links to societal and economic benefits. In this review, we summarize the current understanding of how cell walls are dynamically regulated in response to nutrient availability and how cell wall defects cause changes to nutrient sensitivity. In order to understand the relationship between cell wall and nutrients, we first introduce the basic concepts of cell wall synthesis and structure, along with nutrient transport. Unless otherwise noted, examples presented are from the model plant Arabidopsis thaliana (Arabidopsis).
1.1. Cell Walls
Plant cell walls are highly dynamic structures. They are composed primarily of polysaccharides, including cellulose, pectins, hemicelluloses, and callose. In this review, we focus only on the areas of cell wall synthesis and regulation that are directly relevant to nutrient response. There are excellent reviews that cover key topics of cell wall synthesis and regulation [7,14,15,16,17,18,19].
1.1.1. Cellulose
Cellulose is composed of β-(1→4)-d-glucan chains, which are crystallized into cellulose microfibrils through inter- and intra- molecular hydrogen bonds and Van der Waals forces. These microfibrils are the primary loadbearing polymers of cell walls and act as a framework for tethering and deposition of other wall components [19].
Cellulose is synthesized by CELLULOSE SYNTHASE A (CESA) catalytic subunits, which are organized at the plasma membrane in large multiprotein complexes called cellulose synthase complexes (CSCs). The CSC constitutes a heteromeric arrangement of 18 to 24 CESAs, with CESA1, CESA3, and a CESA6-like protein (CESA2, 5, 6 or 9) being required for primary cell wall synthesis [14,20]. Certain specialized cells, such as xylem tracheary elements, also synthesize a secondary cell wall that is deposited between the primary cell wall and the plasma membrane. During secondary cell wall synthesis, the CSC is comprised of CESA4, CESA7, and CESA8 [21,22]. Most secondary cell walls contain a significantly increased amount of lignins, which are hydrophobic aromatic polymers typically derived from phenylalanine [23].
Apart from their plasma membrane localization, CSCs are also localized at the Golgi apparatus, trans-Golgi network, small CESA compartments, or microtubule-associated cellulose synthase compartments [24]. These latter compartments may be involved in delivery or internalization of the CSCs [14]. During synthesis, newly formed cellulose microfibrils become entangled in cell walls through cross-linking with cell wall polymers. Further synthesis of cellulose pushes the CSCs forward along the plasma membrane [25,26]. CSC speed is therefore often used as a proxy for CESA catalytic activity. The direction of CSC movement, as well as its targeted delivery to the plasma membrane, is guided by cortical microtubules [27]. Several proteins are involved in guiding the CESAs along microtubules; their functions are necessary to maintain cellulose synthesis. These proteins include CELLULOSE SYNTHASE-MICROTUBULE UNCOUPLING (CMU), COMPANIONS OF CELLULOSE SYNTHASE (CCs), and CELLULOSE SYNTHASE INTERACTING PROTEIN 1(CSI1) [28,29,30]. Of particular note, CCs bind to CSCs and microtubules, and regulate cellulose synthesis under salt stress conditions by re-establishing the microtubule array following salt stress-mediated microtubule depolymerization [29]. KORRIGAN 1 (KOR1), an endo-1,4-β-d-glucanase, also functions in cellulose synthesis by interacting with the CSC at the plasma membrane and during intracellular trafficking [31]. Although the precise function of KOR1 is unknown, kor1 mutants display reduced cellulose synthesis, and KOR1 is thought to play a role in relieving tensional stress generated during microfibril synthesis, or by releasing microfibrils from the CSC during cessation of cellulose synthesis [32]. COBRA and COBRA-like proteins encode glycosylphosphatidylinositol anchored proteins, and are involved in cellulose synthesis and modify cellulose crystallinity [33]. Finally, the chitinase-like protein homologs, CHITINASE-LIKE PROTEIN 1 (CTL1) and CTL2, bind cellulose and impair CSC activity at the plasma membrane [34,35,36]. Although the exact function of CTL1 and CTL2 is unclear, CTL1 colocalizes with CESAs during secretion to the apoplast [35,36].
1.1.2. Pectins
Pectins are a diverse family of complex acidic polysaccharides that act as a hydrophilic gel in which other cell wall components are embedded. Pectin composition can vary widely in chain length and branching complexity; however, all pectins contain 1,4-linked α-d-galacturonic acid residues [19]. Pectins are synthesized in the Golgi, and require a minimum of 67 transferases, including glycosyltransferases, acetyltransferases, and methyltransferases, many of which remain unknown or uncharacterized. A large number of transferases may be required due to the many diverse linkages present in pectins [16]. Following synthesis, pectins are packaged into vesicles and trafficked to the plasma membrane for secretion to the cell wall [37]. In the cell wall, cellulose and pectins are closely linked via hydrogen bonds [38]. Pectin deposition and pectin modification play a central role in mediating cell growth [39]. Pectins are typically secreted to the cell wall in a highly methyl-esterified form, and cell wall-localized pectin methylesterases can act upon pectins to cleave methyl ester bonds to remove the methyl groups [40]. This results in the production of free carboxylic groups, which drastically alters the physical properties of pectins, where low levels of methyl-esterified pectins are typically associated with decreased cell wall extensibility and growth inhibition [41].
1.1.3. Hemicelluloses
Similar to pectins, hemicelluloses are synthesized within the Golgi, packaged into vesicles, and secreted to the apoplast [42]. Hemicelluloses are characterized as molecules containing a backbone of β-(1→4)-linked xylose, glucose, or mannose. Mixed linked glucan (MLG) is a special hemicellulose containing a backbone of unbranched (1,3)- and (1,4)-linked b-glucosyl residues and is typically found in grasses, such as cereals [43], which contain distinct differences in cell wall composition compared to dicotyledons, like Arabidopsis [44]. Within the cell wall, hemicelluloses function by crosslinking cellulose microfibrils via hydrogen bonds [42]. Cosgrove (2014) [45] proposed a model, where xyloglucans, a hemicellulose, act as an adhesive layer between cellulose microfibrils to bundle them together at biomechanical hotspots, thus performing an important role in cell wall structure and integrity by maintaining a strong network of interconnected cellulose microfibrils.
1.1.4. Callose
Although plant cells are surrounded by cell walls, they are also symplastically connected through plasmodesmata (PD) (analogous in function to gap junction in animal cells). These are symplastic channels that transverse the cell walls and connect the cytoplasm of adjacent cells [46]. For cell-to-cell transport of signaling molecules to occur through PD, cell wall composition is important, as the major regulation of PD transport is controlled by the deposition or removal of callose (β-1,3-glucan) in the cell walls surrounding PD. Callose deposition causes restriction of the symplastic channels, isolating the cells from each other and interrupting symplastic signaling [46]. Callose is also deposited in the cell plate in dividing cells, pollen tubes, roots, and can be induced by various biotic and abiotic stresses, including wounding [46,47,48].
1.1.5. Suberin
Suberin is a hydrophobic lipid phenolic polyester comprised of aliphatic, glycerol, and phenolic monomers, with α-ω-dicarboxylic acids and ω-hydroxy acids being the main monomers [49]. Suberin precursors are transported from the endoplasmic reticulum to the plasma membrane for secretion to the apoplast, where suberin polymerization occurs; however, the pathways that mediate these processes remain largely unknown [49]. Suberin localizes to the cell walls of specialized cells, including seed coats and root endodermal cells. Suberin deposition can occur in response to various biotic and abiotic stresses. Most importantly, suberin is deposited to the inner surface of the primary cell wall of root endodermal cells to act as a barrier for diffusion of water and nutrients to the stele [49].
1.1.6. Lignin
Lignin is a complex phenolic polymer typically deposited to secondary cell walls and commonly associated with woody tissues [50]. Lignin is mainly comprised of three monolignols (hydroxycinnamyl alcohols) synthesized from phenylalanine, including ρ-coumaryl, sinapyl, and coniferyl alcohols [23]. Within the cytoplasm, a complex array of reactions is required for monolignol biosynthesis, including deamination, hydroxylation, methylation, and reduction [50]. Monolignols are then transported to the plasma membrane and secreted to the cell wall, where polymerization and crosslinking occurs. Lignification induces unique physical characteristics to cells and tissues by enhancing rigidity through increased cross-linking of cell wall polysaccharides [51]. Lignin deposition is developmentally regulated within specialized cells and tissues, such as the endodermis and xylem [52]. Lignin deposition can also be induced in response to various biotic and abiotic stresses [53]. In the root endodermis, lignin is deposited in maturing cells, specifically to the radial and transverse domains of the cell wall, in a ring-shaped conformation known as the Casparian strip [54]. This acts as a barrier to the apoplastic diffusion of water and nutrients and is important in maintaining nutrient homeostasis. Characteristic hydrophobic properties of lignin are also essential for efficiently transporting water and minerals in water-conducting xylem cells [55].
1.1.7. Structural Proteins and Enzymes
A wide variety of structural proteins and enzymes is required to remodel structures or interactions of cell wall components, leading to wall extensibility and loosening. These proteins include xyloglucan hydrolases, β-1,4-glucanases, peroxidases, extensins, and expansins [56]. Their expression is regulated by a variety of abiotic stresses, including drought and salt stress. Several studies found that these proteins regulate cell wall stiffness through the cross-linking of different polymers. Consequently, ectopic expression of several cell wall modifying proteins leads to abiotic stress tolerance or hyper-susceptibility [57,58].
2. Importance of Cell Walls in Nutrient Transport
In addition to carbon dioxide and water, both of which are required for photosynthetic production of carbohydrates, plant growth depends on nutrient acquisition from the soil. Balanced proportions of a range of macronutrients (nitrogen (N), phosphorus (P), potassium (K), and sulfur (S)), and micronutrients (iron (Fe), copper, molybdenum, selenium, zinc, and others) are essential for optimal growth and crop yield. Plant growth under nutrient-deficient conditions typically results in nutrient-specific phenotypes, including an overall reorganization of root system architecture (RSA), and oftentimes, growth inhibition [59].
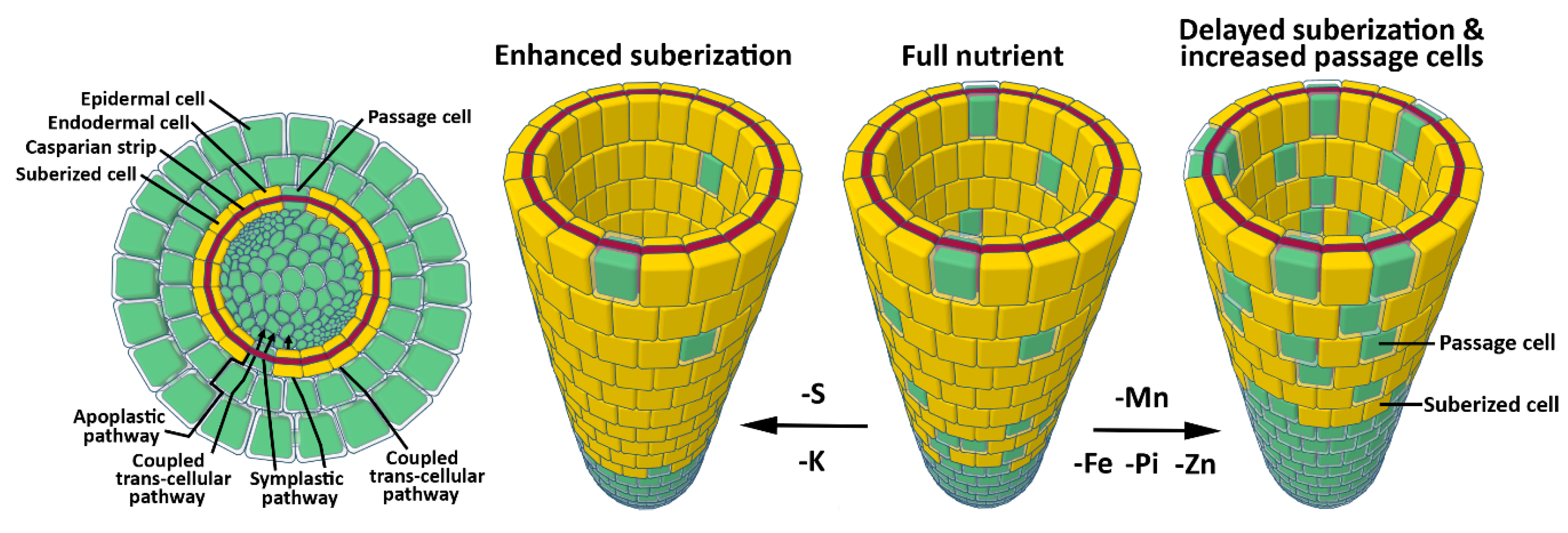
Nutrients and water are absorbed at the root-soil interface and are translocated from the soil to the stele via three distinct pathways: An apoplastic pathway, a symplastic pathway, and a coupled transcellular pathway [60]. Water and nutrients freely diffuse through the apoplastic pathway in an unregulated manner. The symplastic pathway is characterized by nutrient movement between cells via PD, with the coupled-transcellular pathway describing nutrient movement in and out of the cells, mediated by activity of influx and efflux carriers (Figure 1) [60]. Cell walls play a key role in regulating nutrient transport between soil and the stele. Casparian strips are highly lignified, hydrophobic cell walls that encompass the radial and transverse domains of root endodermal cells in the zone of maturation. Casparian strips fill the cell wall between each endodermal cell, creating an apoplastic barrier to water and nutrient diffusion. Within the maturation zone, nutrients must therefore move through the symplastic and coupled transcellular pathways to bypass Casparian strips [61]. Following Casparian strip formation, suberin is deposited to the inner surface of the primary cell wall, entirely enveloping each endodermal cell [49,60]. The so-called suberin lamellae initiates along patches of endodermal cells apical to the Casparian strip initiation site, eventually displaying homogenous cell wall distribution across the endodermis at increasing distances from the root tip. This barrier is crucial in limiting the diffusion of water and nutrients across the endodermis, thus protecting the plant from nutrient influx at toxic levels, or nutrient leaching from the stele [62]. Suberin production is tightly regulated by ethylene and abscisic acid (ABA) signaling, and its synthesis and deposition is highly plastic in response to nutrient availability. Moderate deficiency in Mn, Fe, and Zn leads to a delay in suberization, while K- and S-deficiency results in increased suberization [62]. This suggests a nutrient management strategy by the plant that modulates bidirectional movement of nutrients across the endodermis. In agreement with this, targeted degradation of suberin leads to suppression of the mutant phenotype in the Fe uptake mutant irt. Moreover, suberin degradation in the endodermis enhances the S deficiency phenotype of S uptake mutants for SULFATE TRANSPORTER 1 (sultr1;1sutr1;2) and leads to K deficiency in plants [62], demonstrating the high plasticity of suberin regulation as an adaptive response to various nutritional cues. The suberization response to nutrient availability is a new field of research that will benefit our understanding of the regulation of nutrient homeostasis within plants.
Within the root zone of continuous suberization, randomly distributed cells, known as passage cells, lack suberin deposition (Figure 1) [61,63,64]. These cells are closely associated with the xylem pole. They lack secondary cell wall deposition and their development is mediated by repression of cytokinin signaling in the root apical meristem (RAM) (Figure 1) [64]. The exact role of passage cells is unclear; however, it is thought that they facilitate water and nutrient transport or cell communication in older parts of the root [61,63,64]. In line with this, expression of the phosphate (Pi) efflux protein family member PHOSPHATE1 (PHO1), which is typically present in pericycle cells, expands into the endodermal passage cells [64]. Furthermore, Pi, Fe, and Zn deficiency simultaneously reduces suberization and increases passage cell occurrence [64]. Dynamic regulation of passage cell occurrence in response to nutritional status and expression of nutrient transporters in these cells supports the hypothesis that passage cells are involved in nutrient transport. Overall, these results suggest that plants tightly regulate nutrient transport across the root by modulating nutrient flux carriers and cell wall permeability (Figure 1).
3. Influence of Nutrient Availability on Cell Wall Composition
3.1. Nitrogen
All plants utilize N in the form of ammonium (NH4+) and nitrate (NO3−). Various groups have investigated the transcriptomic response of Arabidopsis plants to N availability. These studies have mainly relied upon NO3− induction or deprivation, followed by RNA extraction and Affymetrix microarray analyses [9]. Few studies specifically highlight the expression of cell wall related genes; however, it was noted that “glucose catabolic process” is one of the most consistent biological functions associated with NO3− response. Genes involved in trichoblast differentiation, which contain many cell wall related genes, were particularly enriched in NO3− responsive genes [9]. Additionally, a co-expression analysis of NO3− transporters, using the ATTED-II database, revealed that they are co-expressed with many cell wall related genes [65]. However, caution should be exercised in interpreting this data, as a recent meta-analysis of NO3−-response transcriptomic data revealed a striking inconsistency in the reproducibility between different studies, noting that the common number of differentially expressed genes between any two experiments is only 6.7% [9]. This variation in expression patterns is likely due to redundancy of large gene families, developmental differences, and changes in experimental setups between labs, as gene expression is highly sensitive to environmental conditions and plant age. This is highlighted in plants that were not subjected to changes in NO3− availability, but showed gene induction resulting from other stress related to simply moving the plants from one solution to another [66]. Transcriptomic assays would greatly benefit from a standardized experimental setup, with detail paid to the nutrient composition of media, lighting levels, plant handling and transfer between media, and the time at which tissue is harvested following treatment. By adopting a standardized experimental setup, future transcriptomic results may lead to increased reproducibility in gene expression trends.
Under N-limiting conditions, plants adapt their RSA to maximize N acquisition by exploiting a larger soil surface area. This is mostly achieved by the elongation of lateral roots (LRs), whereas high N concentrations result in an inhibition of LR growth, with little effect on primary root (PR) growth [59,67]. In crop plants, N status affects stem mechanical strength and disease resistance. These traits are regulated by cell wall organization and strength, suggesting that cell walls are modulated in response to N status. Expression of genes involved in biosynthesis of both lignin and cellulose was significantly reduced in response to high N in rice [68,69]. In support of this finding, cell wall analysis of roots showed that high N leads to a significant reduction of cellulose and lignin. This reduction in cell wall components is accompanied by reduced stem mechanical strength, increased lodging, and reduced disease resistance [68,69,70]. Similar to roots, expression of cellulose and lignin synthesis genes is reduced in filling seeds in response to high N. This leads to reduced accumulation of both cellulose and lignin in seed endosperms [71]. In line with this observation, N deficiency leads to an increase in cellulose content in the roots of rice plants [72].
In Arabidopsis, intact cell walls are essential for the adaptation of RSA in response to N availability, as genetic mutants with altered cell wall organization show impaired RSA response to N availability. Cellulose-deficient mutant roots are hypersensitive to high (60 mM) KNO3 [73]. When grown under high N conditions, cesa3, cesa6, and ctl1 mutants show a drastic reduction in PR length, an increase in root hair and LR density, and swollen roots. Further investigation of ctl1 revealed the same hypersensitive root response to high NaCl, KCl and CaCl2, whereas no response was observed under high PO4, SO4, mannitol, or sorbitol, suggesting a hypersensitive response to chloride ions [35]. Hypersensitivity to chloride in conjunction with NO3− is not unexpected, as multiple NO3− transporters are also known to function in chloride transport [74]. Furthermore, under high KNO3, ctl1 and cesa3 mutants displayed ectopic lignin deposition in the endodermis, whereas cesa6 and kor1 exhibited only minor ectopic lignin deposition compared with wild-type controls [73]. If cellulose reduction occurs in Arabidopsis under high N availability (as also observed in rice), this could explain the hypersensitive root phenotype of cellulose deficient mutants when subjected to high N, resulting from an additive effect due to high N-mediated reduction in cellulose in an already cellulose deficient mutant. These results demonstrate the complex response to N availability, inferring a dynamic regulation of cell wall synthesis and remodeling. However, the molecular mechanisms by which N status affects cell wall biosynthesis and organization are unknown.
3.2. Phosphorus
In a recent study of Pi-responsive genes, a detailed transcriptomic analysis using wild-type and mutants for LOW PHOSPHATE RESPONSE 1 (LPR1) and LPR2, and PHOSPHATE DEFICIENCY RESPONSE 2, which are required for Pi response, highlighted a robust Pi-dependent regulation of cell wall related genes [75]. These genes were sorted into four main groups, which included pectin modification, cell wall relaxation, hemicellulose/cellulose modification, and carbohydrate hydrolytic enzymes; however, it is unknown how these changes in expression impact growth. Additional studies in wild-type plants noted similar changes in expression of cell wall related genes in response to Pi starvation [10,13,76,77]. A transcriptomic analysis for seedlings subjected to Pi-deficiency revealed differential regulation of the largely uncharacterized CELLULOSE SYNTHASE-LIKE B5 (CSLB5) [10]. Further investigation revealed that cslb5 mutants produce shorter root hairs compared to wild-type under Pi-deficiency, suggesting a role of CSLB5 in Pi response [10]. The exact function of the CSLB family is unknown; however, it is thought that members are involved in the synthesis of β-1,4-linked cell wall polysaccharides [78]. Interestingly, CSLB5 is expressed in roots, which goes in line with its likely function in regulation of RSA in response to Pi starvation [78].
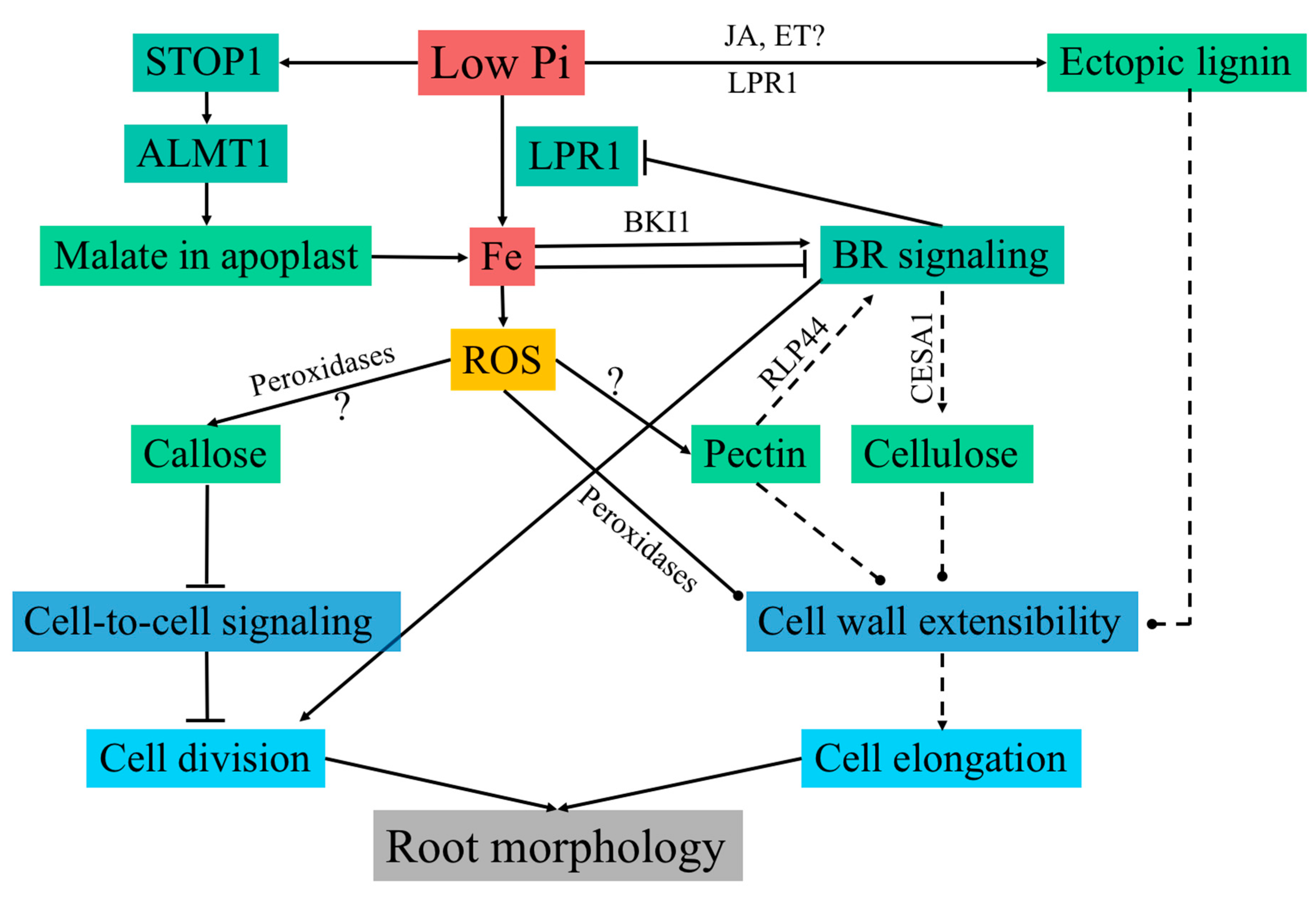
Under low Pi conditions in Arabidopsis, PR growth is arrested, while growth of secondary roots and root hairs is promoted [59,79]. PR growth arrest is mainly a result of rapid reduction in root cell elongation followed by an arrest of cell division in the stem cell niche. These changes in the RSA are dependent on the external supply of Fe. This suggests an indirect surveillance of Pi status via its interaction with transition metals [80]. During Pi deficiency, LPR1, which is a cell wall localized ferroxidase, modulates Fe deposition in the cell walls of elongating cells and the stem cell niche [48]. Interestingly, in these cells, Fe deposition is promoted by malate secretion to the cell walls. Under low Pi conditions, the transcription factor SENSITIVE TO PROTON TOXICITY 1 (STOP1) targets the malate efflux channel ALUMINUM-ACTIVATED MALATE TRANSPORTER 1 (ALMT1), triggering malate secretion to the cell walls of the root transition zone. Malate deposition promotes Fe3+ accumulation via LPR1 activity in the elongating cells [81,82], coinciding with the generation of reactive oxygen species (ROS) and callose deposition [48]. Cell type-specific callose deposition disrupts symplastic communication through plasmodesmata as shown by impaired movement of the transcription factor SHORT ROOT between the stele and the endodermis [48]. Symplastic communication is essential for meristem maintenance, and its disruption is likely the main factor leading to root meristem exhaustion in response to low Pi (Figure 2) [83]; however, the mechanism behind the rapid inhibition of cell elongation, which occurs well before meristem size reduction, is unclear [48,81]. LPR1 mediated Fe accumulation leads to the generation of ROS, which coincides with the initial sites of Fe deposition in the apoplast [48]. Ferroxidase activity of LPR1 leads to Fe3+ redox cycling in the cell wall, which creates an overall oxidative environment and ROS generation [48]. ROS may serve as a substrate for the cell wall localized class III peroxidases to catalyze cross-linking between cell wall components (Figure 2) [18], leading to cell wall stiffness and reduced extensibility. Atomic force microscopy revealed an increase in cell wall stiffness in the elongation zone within 30 min following seedling transfer to Pi-deficient media, with the change in stiffness being dependent on the activity of class III peroxidases [81]. Pharmacological inhibition of peroxidases restored root growth in response to low Pi, confirming that increased cell wall rigidity is linked to inhibited root growth under Pi-deficient conditions [81]. Changes in cell wall stiffness need to happen in conjunction with changes in cell turgor in order to coordinate cell expansion [84,85]. This may be achieved through conformational changes in lipids [86], which typically result in changes in cellular trafficking and vacuolar organization [87,88], which are crucial factors for the regulation of intracellular turgor [85].
Varying Fe concentration leads to activation or inhibition of the brassinosteroid (BR) signaling pathway, which in turn regulates the expression of LPR1 [89]. This feedback loop may be important in mediating distinct root morphologies as an adaptive response to soil microenvironments with varying concentrations of either Pi or Fe [89]. BRASSINAZOLE RESISTANT 1 (BZR1), a key regulator of BR mediated regulation of gene expression [90], represses the expression of LPR1 by directly binding to its promoter [89]. In line with this, BZR1 constitutive mutants (BZR1-D) phenocopy the lpr1 root phenotype in response to low Pi [91]. Low Fe activates, and high Fe inhibits, BR signaling via translational modulation of its key repressor brassinosteroid kinase inhibitor 1 (BKI1) through an unknown mechanism [89]. Fe concentration-dependent modulation of BR signaling is required for the root growth response in low Pi conditions (Figure 2). Interestingly, BR signaling is known to play a role both in cell proliferation and cell elongation via changes in cell wall composition. Mutants affected in the BR signaling pathway show significant transcriptional changes to various cell wall remodeling enzymes and structural proteins [92]. More importantly, BRASSINOSTEROID INSENSITIVE 2 (BIN2), a key inhibitor of the BR pathway, directly phosphorylates CESA1, leading to a decrease in CESA motility and crystalline cellulose synthesis [93]. BES1 and BZR1, which are the key transcription factors in the BR pathway, positively impact cellulose synthesis by directly binding to CESA promoters [94]. Along with the direct regulation of cellulose synthesis, BR is known to influence microtubule rearrangement [95], which is required to guide CSCs during cellulose synthesis. CSI1, CMU, and CCs are the proteins required for the association of CSC with microtubules [28,29,30]. Investigation of these proteins in context of BR and phosphate starvation response may prove to be an important tool in investigating changes in cellulose synthesis in low Pi stress. Overall, the BR pathway positively impacts cellulose synthesis, and it could be a strong candidate for the regulation of cellulose in response to low Pi.
The cellulose content of rice changes in responses to low Pi [96]; however, it is unknown whether Pi deficiency affects cellulose content in Arabidopsis. Furthermore, rice mutants affected in CELLULOSE SYNTHASE-LIKE F6 (CSLF6) show a constitutively active Pi starvation response and increased Pi transport, even when seedlings are grown on high Pi [97]. CSLF6 is involved in the synthesis of MLG, which typically affects cell wall strength and cell expansion [43]. Interestingly, MLG binds to cellulose in vitro, forming a thick hydrogel at the surface of adsorption [98]. It is suggested that MLG may act as a transient binding surface where other cell wall polysaccharides can attach during periods of cell expansion [43]. Activation of the Pi starvation response in cslf6 mutants implies an interdependent regulation of cell wall composition and Pi signaling.
Pi deficiency also leads to lignin accumulation in root cell walls; however, lignin deposition was not observed in the lpr1lpr2 double mutant, suggesting that lignin deposition may be regulated by the same pathway as callose deposition under Pi-deficiency [99]. Lignin deposition is typically induced due to defects in cell wall integrity through the action of jasmonates and ethylene [100,101]. Both jasmonates and ethylene are induced in response to low Pi and possibly mediate the observed ectopic lignin deposition [102,103]. Lignin associates with cellulose and is involved in cell wall strengthening, implicating it in reduced extensibility of cell wall in low Pi conditions [104]. Under Pi-deficiency, the concentration of unesterified pectins significantly increases in cell walls in the elongation zone and the RAM, with a notable increase at the quiescent center and the border between the meristem and the stem cell niche [75]. The sites of increased pectin deposition colocalize to the sites of Fe3+ accumulation and callose deposition, indicating that pectins may play a role in cell wall organization in the root elongation inhibition response. Free carboxyl groups in unesterified pectins have a high affinity to bind to Fe3+, Al3+, and Ca2+, and pectins are known to dimerize via Ca2+-pectate crosslinked complexes called egg-boxes [105]. Increase in egg-boxes causes cell wall stiffening and reduced growth [106]. It is tempting to speculate that the so-called egg-box structures can be formed with the Fe3+, leading to an increase in cell wall stiffness. In addition, Fe binding with pectin may promote the release of Pi from Fe-Pi complexes during low Pi stress [107], which would be a fascinating adaptation to maximize Pi absorption. Interestingly, changes in pectin esterification are also known to trigger BR signaling via RECEPTOR-LIKE PROTEIN 44 (RLP44), which is required for the feedback regulation of cell wall modifications and coordination developmental output with cell wall integrity [108]. Dual control of BR signaling via pectin composition and Fe is perhaps required to modulate cell wall composition, driving RSA organization, while maintaining cell wall integrity.
Data summarized here demonstrate that cell wall modulation is essential for root response to low Pi, and that significant progress has been made in dissecting the complex genetic networks involved in this process; however, challenges remain in linking Fe accumulation and ROS generation with the modulation of cell wall chemistry. For example, what molecular mechanisms and pathways are responsible for recruiting callose synthesizing enzymes to the sites of Fe accumulation?
3.3. Other Nutrients
Apart from N and P, little is known about cell wall regulation by other nutrients. A study by Armengaud et al. (2004) [11] revealed that cell wall related genes, including extensins, xyloglucan glucosyltransferases, arabinogalactan proteins, and peroxidases, were enriched in genes that respond to low potassium (K). A meta-analysis of transcriptomic studies revealed that cell wall related genes are enriched in low S responsive genes [12]. As observed in the meta-analysis for genes responding to NO3− availability, the reproducibility in consistent gene response was low between the datasets, with only 418 (20%) of the differentially expressed genes shared by a minimum of two experiments. It was found that the gene ontology term “cell wall” had a two-fold enrichment, genes coding apoplast-localized proteins were over-represented, and a predicted glycosyl hydrolase (At3g60140) was significantly up-regulated across all microarray datasets, implicating an S-dependent regulation of cell wall related genes.
The micronutrient boron forms so-called borate bridges by crosslinking the pectin polysaccharide rhamnogalacturonan-II [109]. These crosslinks modify cell wall porosity, mechanical properties, and are critical for plant growth. Consequently, boron deficiency leads to highly de-structured cell walls and severe growth defects [109].
As we understand more of the complex genetic networks in cell wall synthesis, it will be possible to identify more genes that regulate these networks in response to nutrient availability. A combination of in vivo and in vitro methods and systems biology approach [110] revealed a complex transcriptional network regulating secondary cell wall synthesis. Interestingly, this network contains several genes that are transcriptionally regulated in response to N, Fe, and S imbalance, suggesting that secondary cell wall synthesis is regulated by nutrient availability. Investigations revealed that the transcription factor REVOLUTA (REV) has a number of upstream factors that are regulated by Fe deprivation, suggesting that REV plays an important role in regulating secondary cell walls during Fe deficiency. Further experiments demonstrated that REV is a key transcription factor involved in the regulation of lignin biosynthesis in response to Fe deficiency [110]. These results demonstrate that cell wall synthesis is tightly regulated in response to nutritional cues and is underpinned by complex genetic networks.
4. Conclusions and Perspectives
Cell wall synthesis, composition, and remodeling is crucial for the meticulous modulation of RSA in response to nutrient imbalance. For example, Fe-mediated callose deposition in the cell walls of the root meristem and elongation zone is crucial for adapting the RSA in response to P deficiency (although it remains unclear how Fe regulates callose deposition in roots) [46]. Moreover, how other cell wall polymers are modulated in response to P imbalance is unknown. Modulation and reorganization of other cell wall polymers is expected, as a drastic reduction in cell division and cell elongation is observed shortly after exposure to low Pi. Apart from P, imbalances in other nutrients results in RSA reorganization [57], which is likely mediated by regulation of cell wall synthesis and/or architecture. This hypothesis is mainly supported by transcriptomics data due to technical advances, simplicity, and reduced cost of this type of analysis. It will be crucial for future studies to investigate changes in cell wall chemistry and cell wall composition using polymer-specific antibodies and mass spectrometry to understand the role of cell walls in nutrient-specific RSA modulation.
Experiments with rice show that cell wall polymers are reduced in response to excess N, which is mediated through transcriptional regulation of genes involved in cell wall biosynthesis [68,69]. It can be expected that cell walls are regulated in a similar manner in Arabidopsis given the sensitivity of cellulose deficient mutants to high N and Cl. If this is the case, Arabidopsis could prove to be a powerful tool for dissecting the molecular mechanisms of N- and Cl-mediated cell wall regulation. Investigating the relationship between N availability and cell wall regulation is important in an agricultural context, as high N directly affects important agronomical traits, such as lodging and disease resistance, through cell wall changes.
Researchers typically apply a reductionist approach by modulating the concentration of a single nutrient. However, in the natural environment, plants are typically exposed to changes in various nutrients simultaneously. Emerging evidence highlights the complexity and diversity of growth responses by subjecting plants to multiple nutrient deficiencies simultaneously, where the output is not an additive effect of each individual nutrient deficiency phenotype, but rather a unique phenotype altogether [111]. As soils contain a heterogeneous distribution of nutrients, and in order to understand their complex response pathways, it will be important to study multiple concurrent changes in nutrient concentrations.
Taken as a whole, the recent works referenced here demonstrate that cell wall synthesis, composition, and remodeling are crucial for both the transport and signaling of important nutrients. A robust body of knowledge now exists about how plants control water and nutrient transport across the root by manipulating cell wall permeability. It is clear that Arabidopsis root cell walls are highly plastic in response to the nutritional status of the soil [58]. It can be expected that this plasticity in root permeability helps plants to locally adjust nutrient transport in soil microenvironments with varying nutrient concentrations. Knowledge gained in this field has significant potential for engineering crops with increased nutrient use efficiency by tailoring RSA and endodermis permeability in accordance with soil nutritional status, while also improving growth and disease resistance.
Author Contributions
M.O., S.P., and G.A.K. conceived the topic and wrote the manuscript. R.H. and U.R. proofread and provided valuable comments to improve the manuscript.
Funding
M.O. is supported by the Melbourne-Potsdam PhD Programme. G.A.K. is funded by a Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung early postdoc mobility fellowship under the grant number P2LAP3_168408. S.P. is funded by an ARC future fellowship grant (FT160100218) and acknowledges a UoM IRRTF (RNC) grant.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prinzenberg, A.E.; Barbier, H.; Salt, D.E.; Stich, B.; Reymond, M. Relationships between Growth, Growth Response to Nutrient Supply, and Ion Content Using a Recombinant Inbred Line Population in Arabidopsis thaliana. Plant Physiol. 2010, 154, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, Z.; Amtmann, A. Food for thought: How nutrients regulate root system architecture. Curr. Opin. Plant Biol. 2017, 39, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-Y.; Ried, M.K.; Hothorn, M.; Poirier, Y. Control of plant phosphate homeostasis by inositol pyrophosphates and the SPX domain. Curr. Opin. Biotechnol. 2018, 49, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Bouguyon, E.; Gojon, A.; Nacry, P. Nitrate sensing and signaling in plants. Semin. Cell Dev. Biol. 2012, 23, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Somssich, M.; Khan, G.A.; Persson, S. Cell wall heterogeneity in root development of Arabidopsis. Front. Plant Sci. 2016, 7, 1242. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef] [PubMed]
- Canales, J.; Moyano, T.C.; Villarroel, E.; Gutiérrez, R.A. Systems analysis of transcriptome data provides new hypotheses about Arabidopsis root response to nitrate treatments. Front. Plant Sci. 2014, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-D.; Liao, Y.-Y.; Yang, T.J.W.; Pan, C.-Y.; Buckhout, T.J.; Schmidt, W. Coexpression-based clustering of Arabidopsis root genes predicts functional modules in early phosphate deficiency signaling. Plant Physiology 2011, 155, 1383–1402. [Google Scholar] [CrossRef] [PubMed]
- Armengaud, P. The Potassium-Dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [PubMed]
- Henríquez-Valencia, C.; Arenas-M, A.; Medina, J.; Canales, J. Integrative transcriptomic analysis uncovers novel gene modules that underlie the sulfate response in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Wege, S.; Khan, G.A.; Jung, J.-Y.; Vogiatzaki, E.; Pradervand, S.; Aller, I.; Meyer, A.J.; Poirier, Y. The EXS domain of PHO1 participates in the response of shoots to phosphate deficiency via a root-to-shoot signal. Plant Physiol. 2016, 170, 385–400. [Google Scholar] [CrossRef] [PubMed]
- McFarlane, H.E.; Döring, A.; Persson, S. The cell biology of cellulose synthesis. Annu. Rev. Plant Biol. 2014, 65, 69–94. [Google Scholar] [CrossRef] [PubMed]
- Albenne, C.; Canut, H.; Jamet, E. Plant cell wall proteomics: The leadership of Arabidopsis thaliana. Front. Plant Sci. 2013, 4, 111. [Google Scholar] [CrossRef] [PubMed]
- Atmodjo, M.A.; Hao, Z.; Mohnen, D. Evolving views of pectin biosynthesis. Annu. Rev. Plant Biol. 2013, 64, 747–779. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Hématy, K.; Höfte, H. Growth control and cell wall signaling in plants. Annu. Rev. Plant Biol. 2012, 63, 381–407. [Google Scholar] [CrossRef] [PubMed]
- Lampugnani, E.R.; Khan, G.A.; Somssich, M.; Persson, S. Building a plant cell wall at a glance. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed]
- Persson, S.; Paredez, A.; Carroll, A.; Palsdottir, H.; Doblin, M.; Poindexter, P.; Khitrov, N.; Auer, M.; Somerville, C.R. Genetic evidence for three unique components in primary cell-wall cellulose synthase complexes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 15566–15571. [Google Scholar] [CrossRef] [PubMed]
- Taylor, N.G.; Laurie, S.; Turner, S.R. Multiple cellulose synthase catalytic subunits are required for cellulose synthesis in Arabidopsis. Plant Cell 2000, 12, 2529–2539. [Google Scholar] [CrossRef] [PubMed]
- Atanassov, I.I.; Pittman, J.K.; Turner, S.R. Elucidating the mechanisms of assembly and subunit interaction of the cellulose synthase complex of Arabidopsis secondary cell walls. J. Biol. Chem. 2009, 284, 3833–3841. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, R.; Lindeboom, J.J.; Paredez, A.R.; Emons, A.M.C.; Ehrhardt, D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009, 11, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Diotallevi, F.; Mulder, B. The cellulose synthase complex: A polymerization driven supramolecular motor. Biophys. J. 2007, 92, 2666–2673. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.L.W.; Strumillo, J.; Zimmer, J. Crystallographic snapshot of cellulose synthesis and membrane translocation. Nature 2012, 493, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredez, A.R.; Somerville, C.R.; Ehrhardt, D.W. Visualization of cellulose synthase demonstrates functional association with microtubules. Science 2006, 312, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Bringmann, M.; Landrein, B.; Schudoma, C.; Hamant, O.; Hauser, M.-T.; Persson, S. Cracking the elusive alignment hypothesis: The microtubule–cellulose synthase nexus unraveled. Trends Plant Sci. 2012, 17, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Endler, A.; Kesten, C.; Schneider, R.; Zhang, Y.; Ivakov, A.; Froehlich, A.; Funke, N.; Persson, S. A mechanism for sustained cellulose synthesis during salt stress. Cell 2015, 162, 1353–1364. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Schneider, R.; Kesten, C.; Zhang, Y.; Somssich, M.; Zhang, Y.; Fernie, A.R.; Persson, S. Cellulose-microtubule uncoupling proteins prevent lateral displacement of microtubules during cellulose synthesis in Arabidopsis. Dev. Cell 2016, 38, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Vain, T.; Crowell, E.F.; Timpano, H.; Biot, E.; Desprez, T.; Mansoori, N.; Trindade, L.M.; Pagant, S.; Robert, S.; Höfte, H.; et al. The cellulase KORRIGAN is part of the cellulose synthase complex. Plant Physiol. 2014, 165, 1521–1532. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C. Cellulose synthesis in higher plants. Annu. Rev. Cell Dev. Biol. 2006, 22, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Shang-Guan, K.; Zhang, B.; Liu, X.; Yan, M.; Zhang, L.; Shi, Y.; Zhang, M.; Qian, Q.; Li, J.; et al. Brittle Culm1, a COBRA-like protein, functions in cellulose assembly through binding cellulose microfibrils. PLoS Genet. 2013, 9, e1003704. [Google Scholar] [CrossRef] [PubMed]
- Mouille, G.; Robin, S.; Lecomte, M.; Pagant, S.; Höfte, H. Classification and identification of Arabidopsis cell wall mutants using Fourier-Transform InfraRed (FT-IR) microspectroscopy. Plant J. 2003, 35, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Porco, S.; Vandenbussche, F.; Gille, S.; de Pessemier, J.; van Der Straeten, D.; Verbruggen, N.; Bush, D.R. Dissecting the role of CHITINASE-LIKE1 in nitrate-dependent changes in root architecture. Plant Physiol. 2011, 157, 1313–1326. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, C.; Bauer, S.; Hématy, K.; Saxe, F.; Ibáñez, A.B.; Vodermaier, V.; Konlechner, C.; Sampathkumar, A.; Rüggeberg, M.; Aichinger, E. vChitinase-like1/pom-pom1 and its homolog CTL2 are glucan-interacting proteins important for cellulose biosynthesis in Arabidopsis. Plant Cell 2012, 24, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Brandizzi, F. The plant secretory pathway: An essential factory for building the plant cell wall. Plant Cell Physiol. 2014, 55, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Park, Y.B.; Cosgrove, D.J.; Hong, M. Cellulose-pectin spatial contacts are inherent to never-dried Arabidopsis primary cell walls: evidence from solid-state nuclear magnetic resonance. Plant Physiol. 2015, 168, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Diffuse Growth of Plant Cell Walls. Plant Physiol. 2018, 176, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Greiner, S. Growth control by cell wall pectins. Protoplasma 2012, 249, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, P.; McCann, M.C.; Roberts, K. Restricted cell elongation in Arabidopsis hypocotyls is associated with a reduced average pectin esterification level. BMC Plant Biol. 2007, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Gille, S.; Liu, L.; Mansoori, N.; de Souza, A.; Schultink, A.; Xiong, G. Hemicellulose biosynthesis. Planta 2013, 238, 627–642. [Google Scholar] [CrossRef] [PubMed]
- Vega-Sánchez, M.E.; Verhertbruggen, Y.; Christensen, U.; Chen, X.; Sharma, V.; Varanasi, P.; Jobling, S.A.; Talbot, M.; White, R.G.; Joo, M. Loss of cellulose synthase-like f6 function affects mixed-linkage glucan deposition, cell wall mechanical properties, and defense responses in vegetative tissues of rice. Plant Physiol. 2012, 159, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Burton, R.A.; Fincher, G.B. Evolution and development of cell walls in cereal grains. Front. Plant Sci. 2014, 5, 456. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, D.J. Re-constructing our models of cellulose and primary cell wall assembly. Curr. Opin. Plant Biol. 2014, 22, 122–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavaliev, R.; Ueki, S.; Epel, B.L.; Citovsky, V. Biology of callose (β-1,3-glucan) turnover at plasmodesmata. Protoplasma 2011, 248, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-Y.; Kim, J.-Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.; Toev, T.; Heisters, M.; Teller, J.; Moore, K.L.; Hause, G.; Dinesh, D.C.; Bürstenbinder, K.; Abel, S. Iron-dependent callose deposition adjusts root meristem maintenance to phosphate availability. Dev. Cell 2015, 33, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, S.J.; Delude, C.; Domergue, F.; Rowland, O. Suberin: Biosynthesis, regulation, and polymer assembly of a protective extracellular barrier. Plant Cell Rep. 2015, 34, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, H.; Kamiya, A.; Nagata, T.; Katahira, M.; Watanabe, T. Direct evidence for α ether linkage between lignin and carbohydrates in wood cell walls. Sci. Rep. 2018, 8, 6538. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Rubio, M.C.; Alassimone, J.; Geldner, N. A mechanism for localized lignin deposition in the endodermis. Cell 2013, 153, 402–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, J.C.M.S.; Bonine, C.A.V.; de Oliveira Fernandes Viana, J.; Dornelas, M.C.; Mazzafera, P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. Casparian strip diffusion barrier in Arabidopsis is made of a lignin polymer without suberin. Proc. Natl. Acad. Sci. USA 2012, 109, 10101–10106. [Google Scholar] [CrossRef] [PubMed]
- Boyce, C.K.; Zwieniecki, M.A.; Cody, G.D.; Jacobsen, C.; Wirick, S.; Knoll, A.H.; Holbrook, N.M. Evolution of xylem lignification and hydrogel transport regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 17555–17558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipchinsky, A. How do expansins control plant growth? A model for cell wall loosening via defect migration in cellulose microfibrils. Acta Physiol. Plant 2013, 35, 3277–3284. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2014, 5, 771. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; McFarlane, H.E.; Persson, S. The impact of abiotic factors on cellulose synthesis. J. Exp. Bot. 2016, 67, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Gruber, B.D.; Giehl, R.F.H.; Friedel, S.; von Wiren, N. Plasticity of the Arabidopsis root system under nutrient deficiencies. Plant Physiol. 2013, 163, 161–179. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M. The endodermis as a checkpoint for nutrients. New Phytol. 2016, 213, 1604–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geldner, N. The endodermis. Annu. Rev. Plant Biol. 2013, 64, 531–558. [Google Scholar] [CrossRef] [PubMed]
- Barberon, M.; Vermeer, J.E.M.; De Bellis, D.; Wang, P.; Naseer, S.; Andersen, T.G.; Humbel, B.M.; Nawrath, C.; Takano, J.; Salt, D.E. Adaptation of Root Function by Nutrient-Induced Plasticity of Endodermal Differentiation. Cell 2016, 164, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.E.; Trontin, C.; Duan, L.; Dinneny, J.R. Beyond the Barrier: Communication in the Root through the Endodermis. Plant Physiol. 2014, 166, 551–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, T.G.; Naseer, S.; Ursache, R.; Wybouw, B.; Smet, W.; de Rybel, B.; Vermeer, J.E.M.; Geldner, N. Diffusible repression of cytokinin signalling produces endodermal symmetry and passage cells. Nature 2018, 555, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Landi, S.; Esposito, S. Nitrate uptake affects cell wall synthesis and modeling. Front. Plant Sci. 2017, 8, 3289. [Google Scholar] [CrossRef] [PubMed]
- Menz, J.; Li, Z.; Schulze, W.X.; Ludewig, U. Early nitrogen-deprivation responses in Arabidopsis roots reveal distinct differences on transcriptome and (phospho-) proteome levels between nitrate and ammonium nutrition. Plant J. 2016, 88, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-H.; Yu, J.-Q.; Hu, D.-G. Nitrate: A crucial signal during lateral roots development. Front. Plant Sci. 2017, 8, 485. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wu, L.; Ding, Y.; Yao, X.; Wu, X.; Weng, F.; Li, G.; Liu, Z.; Tang, S.; Ding, C. Nitrogen fertilizer application affects lodging resistance by altering secondary cell wall synthesis in japonica rice (Oryza sativa). J. Plant Res. 2017, 130, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ruan, R.W.; Yuan, X.H.; Hu, D.; Yang, H.; Li, Y.; Yi, Z.L. Effects of nitrogen fertilizer and planting density on the lignin synthesis in the culm in relation to lodging resistance of buckwheat. Plant Prod. Sci. 2015, 18, 218–227. [Google Scholar] [CrossRef]
- Matsuyama, N. The effect of ample nitrogen fertilizer on cell-wall materials and its significance to rice blast disease. Jpn. J. Phytopathol. 1975, 41, 56–61. [Google Scholar] [CrossRef]
- Midorikawa, K.; Kuroda, M.; Terauchi, K.; Hoshi, M.; Ikenaga, S.; Ishimaru, Y.; Abe, K.; Asakura, T. Additional nitrogen fertilization at heading time of rice down-regulates cellulose synthesis in seed endosperm. PLoS ONE 2014, 9, e98738. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, L.; Wang, F.; Deng, M.; Yi, K. Modulating the root elongation by phosphate/nitrogen starvation in an OsGLU3 dependant way in rice. Plant Signal. Behav. 2012, 7, 1144–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, C.; Porco, S.; Verbruggen, N.; Bush, D.R. Chitinase-like protein ctl1 plays a role in altering root system architecture in response to multiple environmental conditions. Plant Physiol. 2010, 152, 904–917. [Google Scholar] [CrossRef] [PubMed]
- Wege, S.; Gilliham, M.; Henderson, S.W. Chloride: Not simply a “cheap osmoticum,” but a beneficial plant macronutrient. J. Exp. Bot. 2017, 68, 3057–3069. [Google Scholar] [CrossRef] [PubMed]
- Hoehenwarter, W.; Mönchgesang, S.; Neumann, S.; Majovsky, P.; Abel, S.; Müller, J. Comparative expression profiling reveals a role of the root apoplast in local phosphate response. BMC Plant Biol. 2016, 16, 106. [Google Scholar] [CrossRef] [PubMed]
- Misson, J.; Raghothama, K.G.; Jain, A.; Jouhet, J.; Block, M.A.; Bligny, R.; Ortet, P.; Creff, A.; Somerville, S.; Rolland, N. A genome-wide transcriptional analysis using Arabidopsis thaliana Affymetrix gene chips determined plant responses to phosphate deprivation. Proc. Natl. Acad. Sci. USA 2005, 102, 11934–11939. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Ma, L.; Hou, X.; Wang, M.; Wu, Y.; Liu, F.; Deng, X.W. Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves. Plant Physiol. 2003, 132, 1260–1271. [Google Scholar] [CrossRef] [PubMed]
- Richmond, T.A.; Somerville, C.R. The cellulose synthase superfamily. Plant Physiol. 2000, 124, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Linkohr, B.I.; Williamson, L.C.; Fitter, A.H.; Leyser, H.M.O. Nitrate and phosphate availability and distribution have different effects on root system architecture of Arabidopsis. Plant J. 2002, 29, 751–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, J.T.; Lahner, B.; Yakubova, E.; Salt, D.E.; Raghothama, K.G. The Effect of Iron on the Primary Root Elongation of Arabidopsis during Phosphate Deficiency. Plant Physiol. 2008, 147, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Balzergue, C.; Dartevelle, T.; Godon, C.; Laugier, E.; Meisrimler, C.; Teulon, J.-M.; Creff, A.; Bissler, M.; Brouchoud, C.; Hagège, A. Low phosphate activates STOP1-ALMT1 to rapidly inhibit root cell elongation. Nat Commun. 2017, 8, 15300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Macías, J.; Ojeda-Rivera, J.O.; Gutiérrez-Alanís, D.; Yong-Villalobos, L.; Oropeza-Aburto, A.; Raya-González, J.; Jiménez-Domínguez, G.; Chávez-Calvillo, G.; Rellán-Álvarez, R.; Herrera-Estrella, L. Malate-dependent Fe accumulation is a critical checkpoint in the root developmental response to low phosphate. Proc. Natl. Acad. Sci. USA 2017, 114, E3563–E3572. [Google Scholar] [CrossRef] [PubMed]
- Vatén, A.; Dettmer, J.; Wu, S.; Stierhof, Y.-D.; Miyashima, S.; Yadav, S.R.; Roberts, C.J.; Campilho, A.; Bulone, V.; Lichtenberger, R. Callose biosynthesis regulates symplastic trafficking during root development. Dev. Cell 2011, 21, 1144–1155. [Google Scholar]
- Guerriero, G.; Hausman, J.-F.; Cai, G. No Stress! Relax! Mechanisms Governing Growth and Shape in Plant Cells. Int. J. Mol. Sci. 2014, 15, 5094–5114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karnik, R.; Waghmare, S.; Zhang, B.; Larson, E.; Lefoulon, C.; Gonzalez, W.; Blatt, M.R. Commandeering channel voltage sensors for secretion, cell turgor, and volume control. Trends Plant Sci. 2017, 22, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y. Phosphate starvation and membrane lipid remodeling in seed plants. Prog. Lipid Res. 2013, 52, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Helms, J.B.; Zurzolo, C. Lipids as targeting signals: lipid rafts and intracellular trafficking. Traffic 2004, 5, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, S.; Sasser, T.; Rajalekshmi, S.; Fratti, R.A. SNAREs, HOPS and regulatory lipids control the dynamics of vacuolar actin during homotypic fusion in S. cerevisiae. J. Cell Sci. 2012, 125, 1683–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Fridman, Y.; Holland, N.; Ackerman-Lavert, M.; Zananiri, R.; Jaillais, Y.; Henn, A.; Savaldi-Goldstein, S. Interdependent nutrient availability and steroid hormone signals facilitate root growth plasticity. Dev. Cell 2018, 46, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, Z.-Y.; Mora-Garcia, S.; Li, J.; Yoshida, S.; Asami, T.; Chory, J. BES1 accumulates in the nucleus in response to brassinosteroids to regulate gene expression and promote stem elongation. Cell 2002, 109, 181–191. [Google Scholar] [CrossRef]
- Singh, A.P.; Fridman, Y.; Friedlander-Shani, L.; Tarkowska, D.; Strnad, M.; Savaldi-Goldstein, S. Activity of the brassinosteroid transcription factors BRASSINAZOLE RESISTANT1 and BRASSINOSTEROID INSENSITIVE1-ETHYL METHANESULFONATE-SUPPRESSOR1/BRASSINAZOLE RESISTANT2 blocks developmental reprogramming in response to low phosphate availability. Plant Physiol. 2014, 166, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Li, L.; Zola, J.; Aluru, M.; Ye, H.; Foudree, A.; Guo, H.; Anderson, S.; Aluru, S.; Liu, P. A brassinosteroid transcriptional network revealed by genome-wide identification of BESI target genes in Arabidopsis thaliana. Plant J. 2011, 65, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, C.; Ketelaar, K.; Schneider, R.; Villalobos, J.A.; Somerville, C.R.; Persson, S.; Wallace, I.S. BRASSINOSTEROID INSENSITIVE2 negatively regulates cellulose synthesis in Arabidopsis by phosphorylating cellulose synthase 1. Proc. Natl. Acad. Sci. USA 2017, 114, 3533–3538. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yang, C.; Wang, X. Brassinosteroids can regulate cellulose biosynthesis by controlling the expression of CESA genes in Arabidopsis. J. Exp. Bot. 2011, 62, 4495–4506. [Google Scholar] [CrossRef] [PubMed]
- Catterou, M.; Dubois, F.; Schaller, H.; Aubanelle, L.; Vilcot, B.; Sangwan-Norreel, B.S.; Sangwan, R.S. Brassinosteroids, microtubules and cell elongation in Arabidopsis thaliana. II. Effects of brassinosteroids on microtubules and cell elongation in the bul1 mutant. Planta 2001, 212, 673–683. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-W.; Xu, L.; Wu, Y.-R.; Chen, X.-A.; Liu, Y.; Zhu, S.-H.; Ding, W.-N.; Wu, P.; Yi, K.-K. OsGLU3, a putative membrane-bound endo-1,4-beta-glucanase, is required for root cell elongation and division in rice (Oryza sativa L.). Mol. Plant 2012, 5, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Fang, C.; Yuan, H.; Wang, S.; Wu, Y.; Liu, X.; Zhang, Y.; Luo, J. Interaction between carbon metabolism and phosphate accumulation is revealed by a mutation of a cellulose synthase-like protein, CSLF6. J. Exp. Bot. 2015, 66, 2557–2567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiemle, S.N.; Zhang, X.; Esker, A.R.; Toriz, G.; Gatenholm, P.; Cosgrove, D.J. Role of (1,3)(1,4)-β-glucan in cell walls: Interaction with cellulose. Biomacromolecules 2014, 15, 1727–1736. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.; Schmidt, S.; Chutia, R.; Müller, J.; Böttcher, C.; Strehmel, N.; Scheel, D.; Abel, S. Non-targeted profiling of semi-polar metabolites in Arabidopsis root exudates uncovers a role for coumarin secretion and lignification during the local response to phosphate limitation. J. Exp. Bot. 2016, 67, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.; Karafyllidis, I.; Wasternack, C.; Turner, J.G. The Arabidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses. Plant Cell 2002, 14, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Caño-Delgado, A.; Penfield, S.; Smith, C.; Catley, M.; Bevan, M. Reduced cellulose synthesis invokes lignification and defense responses in Arabidopsis thaliana. Plant J. 2003, 34, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Khan, G.A.; Vogiatzaki, E.; Glauser, G.; Poirier, Y. Phosphate deficiency induces the jasmonate pathway and enhances resistance to insect herbivory. Plant Physiol. 2016, 171, 632–644. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Liu, D. Ethylene and plant responses to phosphate deficiency. Front. Plant Sci. 2015, 6, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodish, H.; Berk, A.; Zipursky, S.L.; Matsudaira, P.; Baltimore, D.; Darnell, J. The dynamic plant cell wall. In Molecular Cell Biology, 4th ed.; W. H. Freeman: New York, NY, USA, 2000. [Google Scholar]
- Grant, G.T.; Morris, E.R.; Rees, D.A.; Smith, P.J.C.; Thom, D. Biological interactions between polysaccharides and divalent cations: The egg-box model. FEBS Lett. 1973, 32, 195–198. [Google Scholar] [CrossRef] [Green Version]
- Chebli, Y.; Geitmann, A. Cellular growth in plants requires regulation of cell wall biochemistry. Curr. Opin. Cell Biol. 2017, 44, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Abel, S. Phosphate scouting by root tips. Curr. Opin. Plant Biol. 2017, 39, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; van der Does, D.; Ladwig, F.; Sticht, C.; Kolbeck, A.; Schürholz, A.-K.; Augustin, S.; Keinath, N.; Rausch, T.; Greiner, S. A receptor-like protein mediates the response to pectin modification by activating brassinosteroid signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 15261–15266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, M.A.; Ishii, T.; Albersheim, P.; Darvill, A.G. Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharide. Annu. Rev. Plant Biol. 2004, 55, 109–139. [Google Scholar] [CrossRef] [PubMed]
- Taylor-Teeples, M.; Lin, L.; de Lucas, M.; Turco, G.; Toal, T.W.; Gaudinier, A.; Young, N.F.; Trabucco, G.M.; Veling, M.T.; Lamothe, R.; et al. An Arabidopsis gene regulatory network for secondary cell wall synthesis. Nature 2015, 517, 571–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellermeier, F.; Armengaud, P.; Seditas, T.J.; Danku, J.; Salt, D.E.; Amtmann, A. Analysis of the root system architecture of Arabidopsis provides a quantitative readout of crosstalk between nutritional signals. Plant Cell 2014, 26, 1480–1496. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Plasticity in the permeability of endodermal cell walls in response to nutrient imbalance. (Left Panel) Cellular schematic cross section of a fully differentiated Arabidopsis root. Suberized endodermal cells (yellow) and the lignified endodermal Casparian strip (red) block nutrient transport via the apoplastic and coupled-trans-cellular pathways. Passage cells are unsuberized endodermal cells that allow symplastic and coupled trans-cellular transport. (Right Panel) Magnified view of the endodermal cell layer under varying nutrient availability. K and S deficiency causes an increase in suberization, while deficiency in Fe, Zn, and P causes a delay in suberization and increased number of passage cells. Mn deficiency also delays suberization, but its impact on passage cell occurrence is unknown. This plasticity in root cell wall permeability is an excellent demonstration of how plants modulate nutrient transport in response to varying nutrient concentrations in soil. Fe: iron; K: potassium; Mn: manganese; Pi: phosphate; S: sulfur; Zn: zinc. Schematics were modeled using the online software Tinkercad (www.tinkercad.com).
Figure 1.
Plasticity in the permeability of endodermal cell walls in response to nutrient imbalance. (Left Panel) Cellular schematic cross section of a fully differentiated Arabidopsis root. Suberized endodermal cells (yellow) and the lignified endodermal Casparian strip (red) block nutrient transport via the apoplastic and coupled-trans-cellular pathways. Passage cells are unsuberized endodermal cells that allow symplastic and coupled trans-cellular transport. (Right Panel) Magnified view of the endodermal cell layer under varying nutrient availability. K and S deficiency causes an increase in suberization, while deficiency in Fe, Zn, and P causes a delay in suberization and increased number of passage cells. Mn deficiency also delays suberization, but its impact on passage cell occurrence is unknown. This plasticity in root cell wall permeability is an excellent demonstration of how plants modulate nutrient transport in response to varying nutrient concentrations in soil. Fe: iron; K: potassium; Mn: manganese; Pi: phosphate; S: sulfur; Zn: zinc. Schematics were modeled using the online software Tinkercad (www.tinkercad.com).

Figure 2.
Model of cell wall regulation in response to Pi starvation. Low Pi causes LPR1-dependent Fe accumulation in the apoplast, which is required for the inhibition of primary root growth. Low Pi also leads to activation of the transcription factor STOP1 through an unknown mechanism. STOP1 induces the malate transporter ALMT1 by directly binding to its promoter. Subsequently, ALMT1 secretes malate in the apoplast, which is required for Fe accumulation in the transition zone. Apoplast-localized LPR1 expresses ferroxidase activity and is thought to generate ROS via Fe redox cycling. Cell-type-specific ROS generation is required for callose deposition and impairs symplastic connectivity, leading to inhibition of cell division in the RAM. In a feedback regulation, Fe modulates LPR1 expression via BR signaling in a concentration-dependent manner. Low Fe concentration activates, and high Fe concentration inhibits, BR signaling by translational regulation of BKI1, which is a key inhibitor of BR signaling. BR signaling is known to regulate cellulose synthesis by the transcriptional and post-translational regulation of CESAs. However, it is unknown whether cellulose synthesis is affected in response to low Pi. Additional cell wall modifications include ectopic lignin deposition, possibly via changes in jasmonate and ethylene signaling, peroxidase-dependent cell wall stiffening, and cell-type-specific pectin deposition. Changes in pectin are known to trigger BR signaling via RLP44 to maintain cell wall integrity. Dual control of BR signaling through pectin and Fe may be required to achieve essential changes in cell walls required for growth modulation in response to low Pi, while maintaining cell wall integrity. Cell wall modifications through changes in cellulose, pectin, and lignin, impact cell wall extensibility. These modifications could be responsible for the rapid inhibition of cell expansion in response to low Pi. Solid lines denote known interactions and dashed lines denote inferred interactions. Arrows are positive interactions and terminated lines indicate negative interactions. Round heads suggest interactions responsible for a change in cell wall architecture. STOP1: SENSITIVE TO PROTON TOXICITY 1; ALMT1: ALUMINUM-ACTIVATED MALATE TRANSPORTER 1; ROS: reactive oxygen species; LPR1: LOW PHOSPHATE RESPONSE 1; BR: brassinosteroid.
Figure 2.
Model of cell wall regulation in response to Pi starvation. Low Pi causes LPR1-dependent Fe accumulation in the apoplast, which is required for the inhibition of primary root growth. Low Pi also leads to activation of the transcription factor STOP1 through an unknown mechanism. STOP1 induces the malate transporter ALMT1 by directly binding to its promoter. Subsequently, ALMT1 secretes malate in the apoplast, which is required for Fe accumulation in the transition zone. Apoplast-localized LPR1 expresses ferroxidase activity and is thought to generate ROS via Fe redox cycling. Cell-type-specific ROS generation is required for callose deposition and impairs symplastic connectivity, leading to inhibition of cell division in the RAM. In a feedback regulation, Fe modulates LPR1 expression via BR signaling in a concentration-dependent manner. Low Fe concentration activates, and high Fe concentration inhibits, BR signaling by translational regulation of BKI1, which is a key inhibitor of BR signaling. BR signaling is known to regulate cellulose synthesis by the transcriptional and post-translational regulation of CESAs. However, it is unknown whether cellulose synthesis is affected in response to low Pi. Additional cell wall modifications include ectopic lignin deposition, possibly via changes in jasmonate and ethylene signaling, peroxidase-dependent cell wall stiffening, and cell-type-specific pectin deposition. Changes in pectin are known to trigger BR signaling via RLP44 to maintain cell wall integrity. Dual control of BR signaling through pectin and Fe may be required to achieve essential changes in cell walls required for growth modulation in response to low Pi, while maintaining cell wall integrity. Cell wall modifications through changes in cellulose, pectin, and lignin, impact cell wall extensibility. These modifications could be responsible for the rapid inhibition of cell expansion in response to low Pi. Solid lines denote known interactions and dashed lines denote inferred interactions. Arrows are positive interactions and terminated lines indicate negative interactions. Round heads suggest interactions responsible for a change in cell wall architecture. STOP1: SENSITIVE TO PROTON TOXICITY 1; ALMT1: ALUMINUM-ACTIVATED MALATE TRANSPORTER 1; ROS: reactive oxygen species; LPR1: LOW PHOSPHATE RESPONSE 1; BR: brassinosteroid.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ogden, M.; Hoefgen, R.; Roessner, U.; Persson, S.; Khan, G.A. Feeding the Walls: How Does Nutrient Availability Regulate Cell Wall Composition? Int. J. Mol. Sci. 2018, 19, 2691. https://doi.org/10.3390/ijms19092691
AMA Style
Ogden M, Hoefgen R, Roessner U, Persson S, Khan GA. Feeding the Walls: How Does Nutrient Availability Regulate Cell Wall Composition? International Journal of Molecular Sciences. 2018; 19(9):2691. https://doi.org/10.3390/ijms19092691
Chicago/Turabian StyleOgden, Michael, Rainer Hoefgen, Ute Roessner, Staffan Persson, and Ghazanfar Abbas Khan. 2018. "Feeding the Walls: How Does Nutrient Availability Regulate Cell Wall Composition?" International Journal of Molecular Sciences 19, no. 9: 2691. https://doi.org/10.3390/ijms19092691
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.