ADAMTS-9 in Mouse Cartilage Has Aggrecanase Activity That Is Distinct from ADAMTS-4 and ADAMTS-5


Abstract
:1. Introduction
2. Results
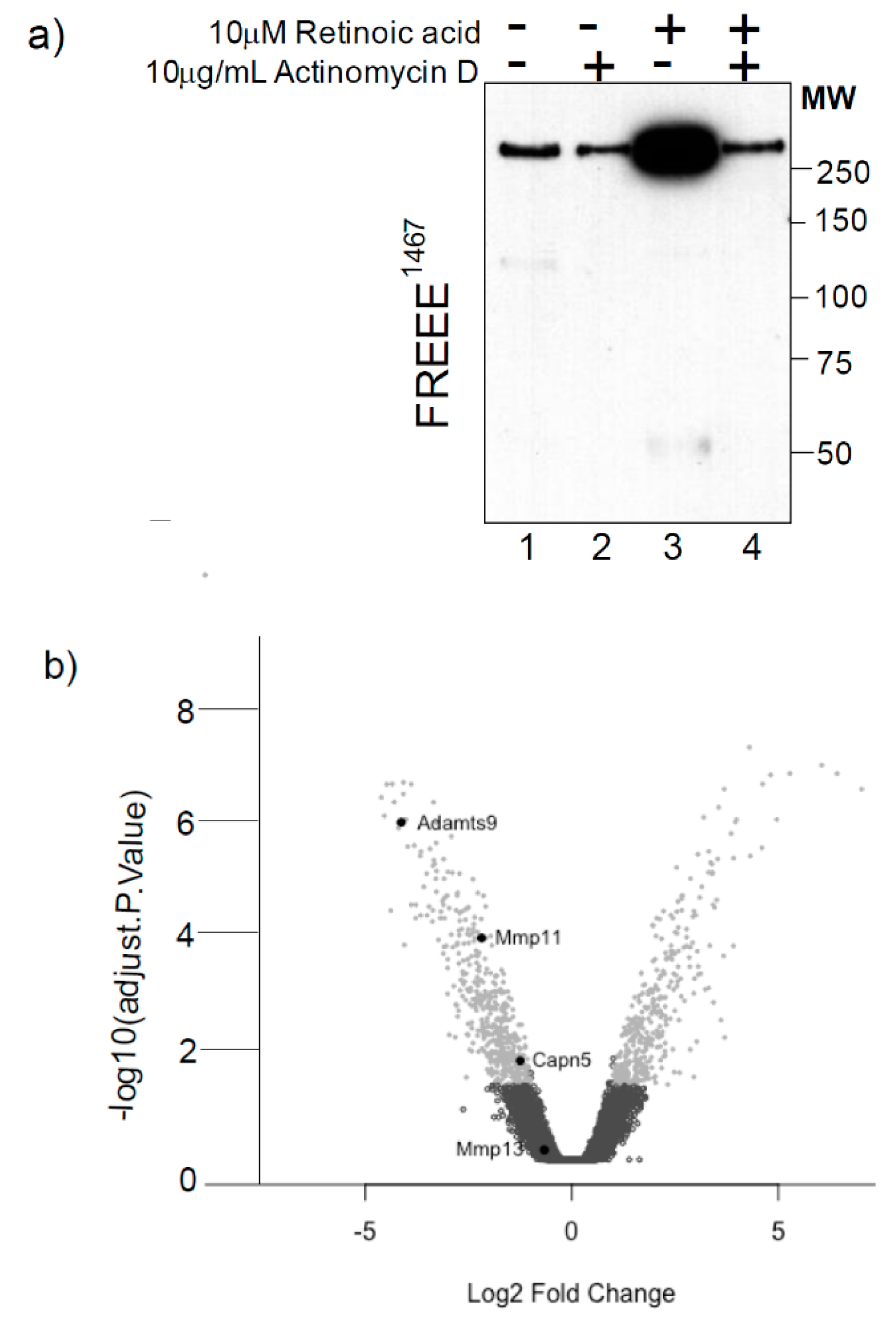
2.1. Retinoic Acid Regulates the Expression of a Novel Aggrecanase Activity via a Transcriptional Mechanism
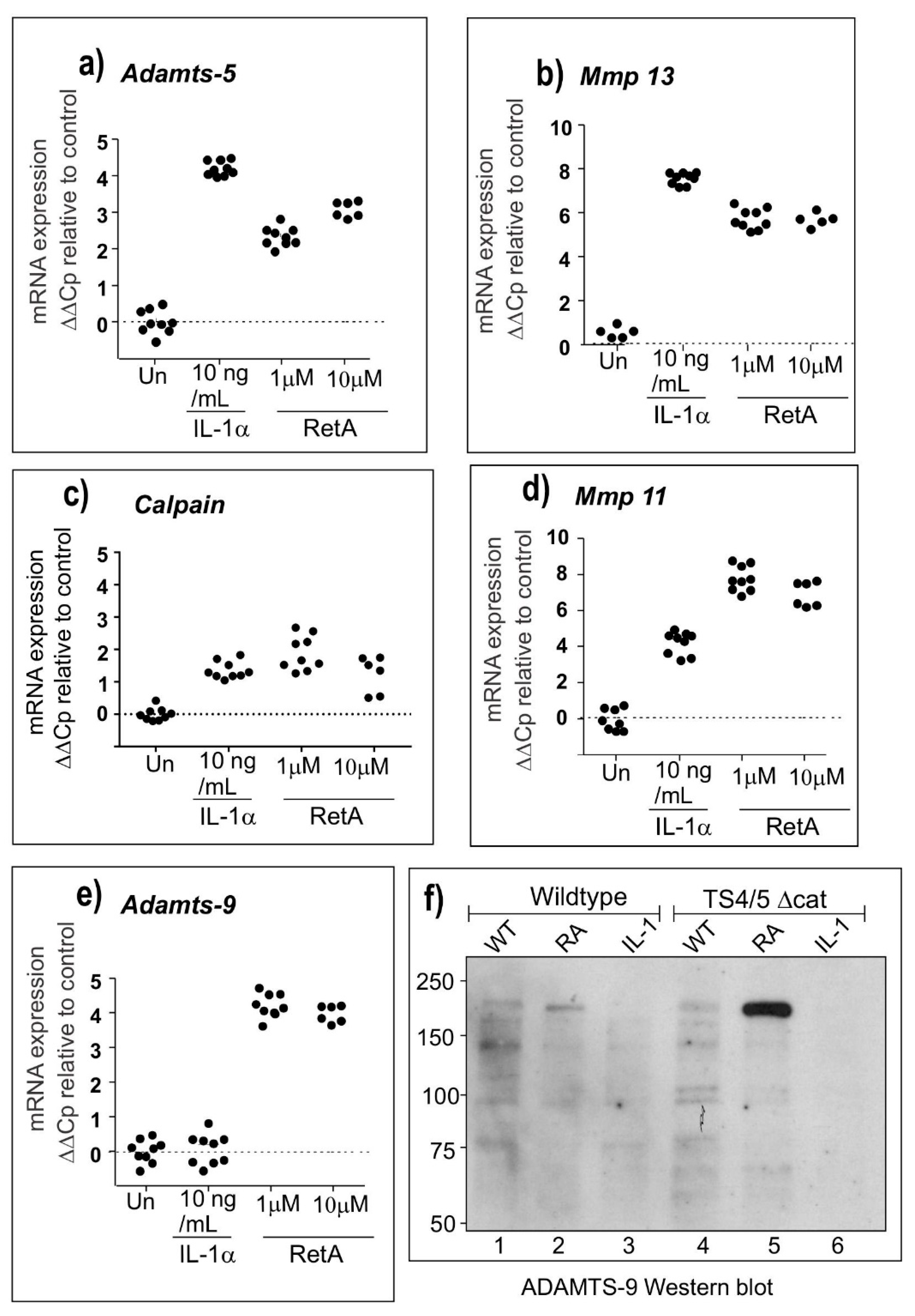
2.2. Microarray and qPCR Studies Reveal ADAMTS-9 as a Strong Candidate for the Novel Aggrecanase
2.3. The Novel Aggrecanase is Inhibited by TIMP-3
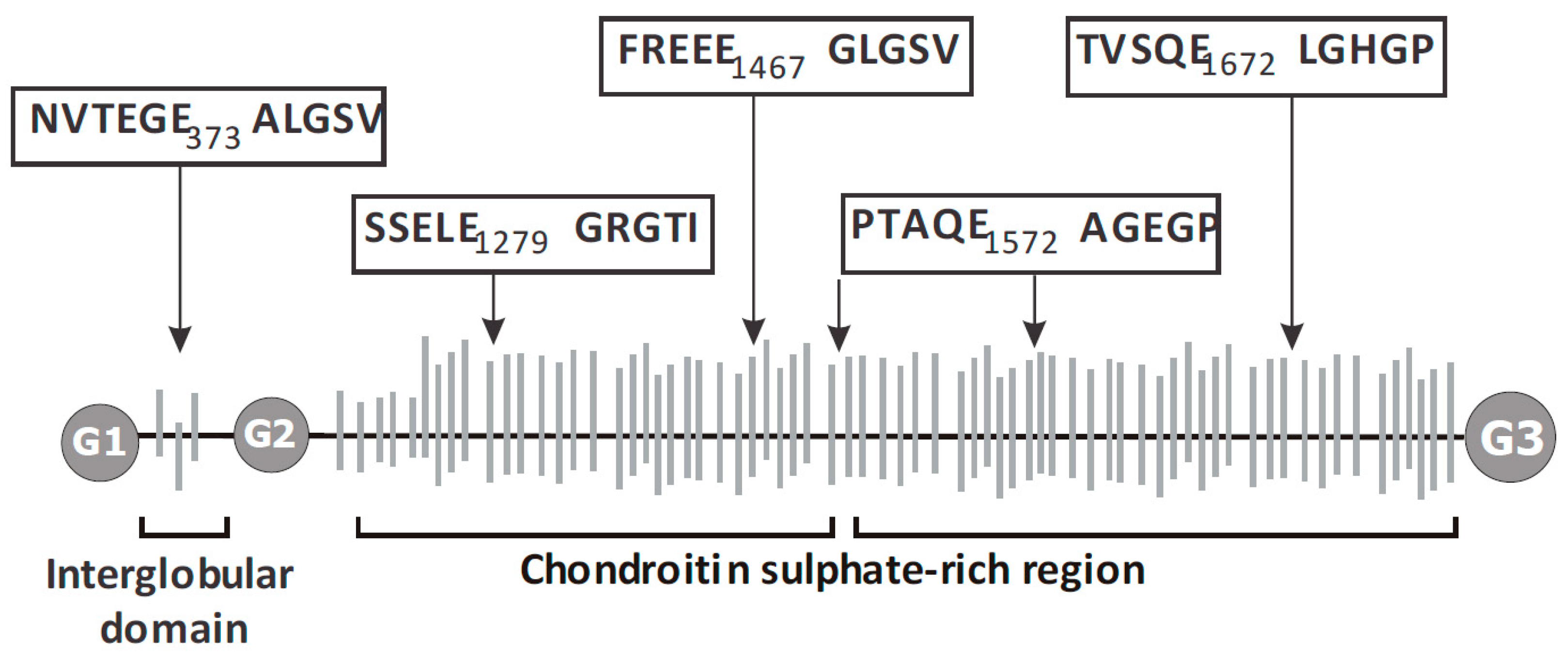
2.4. ADAMTS9 Expression and Aggrecanase Activity in the Growth Plate
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Preparation of Primary Chondrocytes
4.3. Culture of Femoral Head Cartilage Explants
4.4. Western Blotting
4.5. Microarray Analysis
4.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR) Analysis of mRNA Expression
4.7. Immunohistochemistry
4.8. Declarations Ethics Approval
4.9. Availability of Data and Materials
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tortorella, M.D.; Burn, T.C.; Pratta, M.A.; Abbaszade, I.; Hollis, J.M.; Liu, R.; Rosenfeld, S.A.; Copeland, R.A.; Decicco, C.P.; Wynn, R.; et al. Purification and Cloning of Aggrecanase-1: A Member of the ADAMTS Family of Proteins. Science 1999, 284, 1664–1666. [Google Scholar] [CrossRef] [PubMed]
- Abbaszade, I.; Liu, R.Q.; Yang, F.; Rosenfeld, S.A.; Ross, O.H.; Link, J.R.; Ellis, D.M.; Tortorella, M.D.; Pratta, M.A.; Hollis, J.M.; et al. Cloning and Characterization of ADAMTS11, an Aggrecanase from the ADAMTS Family. J. Biol. Chem. 1999, 274, 23443–23450. [Google Scholar] [CrossRef]
- Sandy, J.D.; Neame, P.J.; Boynton, R.E.; Flannery, C.R. Catabolism of aggrecan in cartilage explants. Identification of a major cleavage site within the interglobular domain. J. Biol. Chem. 1991, 266, 8683–8685. [Google Scholar] [PubMed]
- Sandy, J.D.; Flannery, C.R.; Neame, P.J.; Lohmander, L.S. The structure of aggrecan fragments in human synovial fluid. Evidence for the involvement in osteoarthritis of a novel proteinase which cleaves the Glu 373-Ala 374 bond of the interglobular domain. J. Clin. Investig. 1992, 89, 1512–1516. [Google Scholar] [CrossRef] [PubMed]
- Lohmander, L.S.; Neame, P.J.; Sandy, J.D. The structure of aggrecan fragments in human synovial fluid: Evidence that aggrecanase mediates cartilage degradation in inflammatory joint disease, joint injury, and osteoarthritis. Arthritis Rheum. 1993, 36, 1214–1222. [Google Scholar] [CrossRef] [PubMed]
- Ilic, M.Z.; Handley, C.J.; Robinson, H.C.; Mok, M.T. Mechanism of catabolism of aggrecan by articular cartilage. Arch. Biochem. Biophys. 1992, 294, 115–122. [Google Scholar] [CrossRef]
- Loulakis, P.; Shrikhande, A.; Davis, G.; Maniglia, C.A. N-terminal sequence of proteoglycan fragments isolated from medium of interleukin-1-treated articular-cartilage cultures. Putative site(s) of enzymic cleavage. Biochem. J. 1992, 284 Pt 2, 589–593. [Google Scholar] [CrossRef] [Green Version]
- Kuno, K.; Okada, Y.; Kawashima, H.; Nakamura, H.; Miyasaka, M.; Ohno, H.; Matsushima, K. ADAMTS-1 cleaves a cartilage proteoglycan, aggrecan. FEBS Lett. 2000, 478, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Collins-Racie, L.A.; Flannery, C.R.; Zeng, W.; Corcoran, C.; Annis-Freeman, B.; Agostino, M.J.; Arai, M.; DiBlasio-Smith, E.; Dorner, A.J.; Georgiadis, K.E.; et al. ADAMTS-8 exhibits aggrecanase activity and is expressed in human articular cartilage. Matrix Biol. 2004, 23, 219–230. [Google Scholar] [CrossRef]
- Somerville, R.P.T.; Longpre, J.-M.; Jungers, K.A.; Engle, J.M.; Ross, M.; Evanko, S.; Wight, T.N.; Leduc, R.; Apte, S.S. Characterization of ADAMTS-9 and ADAMTS-20 as a Distinct ADAMTS Subfamily Related to Caenorhabditis elegans GON-1. J. Biol. Chem. 2003, 278, 9503–9513. [Google Scholar] [CrossRef]
- Zeng, W.; Corcoran, C.; Collins-Racie, L.A.; Lavallie, E.R.; Morris, E.A.; Flannery, C.R. Glycosaminoglycan-binding properties and aggrecanase activities of truncated ADAMTSs: Comparative analyses with ADAMTS-5, -9, -16 and -18. Biochim. Biophys. Acta 2006, 1760, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Stanton, H.; Golub, S.B.; Rogerson, F.M.; Last, K.; Little, C.B.; Fosang, A.J. Investigating ADAMTS-mediated aggrecanolysis in mouse cartilage. Nat. Protoc. 2011, 6, 388–404. [Google Scholar] [CrossRef] [PubMed]
- Rogerson, F.M.; Stanton, H.; East, C.J.; Golub, S.B.; Tutolo, L.; Farmer, P.J.; Fosang, A.J. Evidence of a novel aggrecan-degrading activity in cartilage: Studies of mice deficient in both ADAMTS-4 and ADAMTS-5. Arthritis Rheum. 2008, 58, 1664–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashiwagi, M.; Tortorella, M.; Nagase, H. Brew K TIMP-3 is a potent inhibitor of aggrecanase-1 (ADAM-TS4) and aggrecanase 2 (ADAM-TS5). J. Biol. Chem. 2001, 276, 12501–12504. [Google Scholar] [CrossRef] [PubMed]
- Struglics, A.; Lohmander, L.S.; Last, K.; Akikusa, J.; Allen, R.; Fosang, A.J. Aggrecanase cleavage in juvenile idiopathic arthritis patients is minimally detected in the aggrecan interglobular domain but robust at the aggrecan C-terminus. Arthritis Rheum. 2012, 64, 4151–4161. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, D.R.; Nelson, C.M.; Dixon, L.J.; Silver, D.L.; Wylie, J.D.; Lindner, V.; Sasaki, T.; Cooley, M.A.; Argraves, W.S.; Apte, S.S. ADAMTS metalloproteases generate active versican fragments that regulate interdigital web regression. Dev. Cell 2009, 17, 687–698. [Google Scholar] [CrossRef]
- Enomoto, H.; Nelson, C.M.; Somerville, R.P.; Mielke, K.; Dixon, L.J.; Powell, K.; Apte, S.S. Cooperation of two ADAMTS metalloproteases in closure of the mouse palate identifies a requirement for versican proteolysis in regulating palatal mesenchyme proliferation. Development 2010, 137, 4029–4038. [Google Scholar] [CrossRef] [Green Version]
- Dubail, J.; Aramaki-Hattori, N.; Bader, H.L.; Nelson, C.M.; Katebi, N.; Matuska, B.; Olsen, B.R.; Apte, S.S. A new Adamts9 conditional mouse allele identifies its non-redundant role in interdigital web regression. Genesis 2014, 52, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Mead, T.J.; Du, Y.; Nelson, C.M.; Gueye, N.A.; Drazba, J.; Dancevic, C.M.; Vankemmelbeke, M.; Buttle, D.J.; Apte, S.S. ADAMTS9-Regulated Pericellular Matrix Dynamics Governs Focal Adhesion-Dependent Smooth Muscle Differentiation. Cell Rep. 2018, 23, 485–498. [Google Scholar] [CrossRef]
- Demircan, K.; Topcu, V.; Takigawa, T.; Akyol, S.; Yonezawa, T.; Ozturk, G.; Ugurcu, V.; Hasgul, R.; Yigitoglu, M.R.; Akyol, O.; et al. ADAMTS4 and ADAMTS5 knockout mice are protected from versican but not aggrecan or brevican proteolysis during spinal cord injury. Biomed. Res. Int. 2014, 2014, 693746. [Google Scholar] [CrossRef]
- Kumagishi, K.; Nishida, K.; Yamaai, T.; Momota, R.; Miyaki, S.; Hirohata, S.; Naito, I.; Asahara, H.; Ninomiya, Y.; Ohtsuka, A. A disintegrin and metalloproteinase with thrombospondin motifs 9 (ADAMTS9) expression by chondrocytes during endochondral ossification. Arch. Histol. Cytol. 2009, 72, 175–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanton, H.; Rogerson, F.M.; East, C.J.; Golub, S.B.; Lawlor, K.E.; Meeker, C.T.; Little, C.B.; Last, K.; Farmer, P.J.; Campbell, I.K.; et al. ADAMTS5 is the major aggrecanase in mouse cartilage in vivo and in vitro. Nature 2005, 434, 648–652. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Lohr, T.A.; Elefante, L.; Shearin, J.; Matico, R.; Su, J.L.; Xue, Y.; Liu, F.; Genell, C.; Miller, R.E.; et al. Translational development of an ADAMTS-5 antibody for osteoarthritis disease modification. Osteoarthr. Cartil. 2015, 23, 1254–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patwari, P.; Kurz, B.; Sandy, J.D.; Grodzinsky, A.J. Mannosamine inhibits aggrecanase-mediated changes in the physical properties and biochemical composition of articular cartilage. Arch. Biochem. Biophys. 2000, 374, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Paulsson, M.; Mörgelin, M.; Wiedemann, H.; Beardmore-Gray, M.; Dunham, D.G.; Hardingham, T.E.; Heinegård, D.; Timpl, R.; Engel, J. Extended and globular protein domains in cartilage proteoglycans. Biochem. J. 1987, 245, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Buckwalter, J.A.; Rosenberg, L.C. Electron microscopic studies of cartilage proteoglycans. Direct evidence for the variable length of the chondroitin sulfate-rich region of proteoglycan subunit core protein. J. Biol. Chem. 1982, 257, 9830–9839. [Google Scholar]
- Flannery, C.; Stanescu, V.; Mörgelin, M.; Boynton, R.; Gordy, J.; Sandy, J. Variability in the G3 domain content of bovine aggrecan from cartilage extracts and chondrocyte cultures. Arch. Biochem. Biophys. 1992, 15, 52–60. [Google Scholar] [CrossRef]
- Dudhia, J.; Davidson, C.M.; Wells, T.M.; Vynios, D.H.; Hardingham, T.E.; Bayliss, M.T. Age-related changes in the content of the C-terminal region of aggrecan in human articular cartilage. Biochem. J. 1996, 313, 933–940. [Google Scholar] [CrossRef] [Green Version]
- Sandy, J.D.; Thompson, V.; Doege, K.; Verscharen, C. The intermediates of aggrecanase- dependent cleavage of aggrecan in rat chondrosarcoma cells treated with interleukin-1. Biochem. J. 2000, 351 Pt 1, 1–166. [Google Scholar] [CrossRef]
- Sandy, J.D.; Verscharen, C. Analysis of aggrecan in human knee cartilage and synovial fluid indicates that aggrecanase (ADAMTS) activity is responsible for the catabolic turnover and loss of whole aggrecan whereas other protease activity is required for C-terminal processing in vivo. Biochem. J. 2001, 358 Pt 3, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Ilic, M.Z.; Robinson, H.C.; Handley, C.J. Characterization of Aggrecan Retained and Lost from the Extracellular Matrix of Articular Cartilage. Involvement of carboxyl-terminal processing in the catabolism of aggrecan. J. Biol. Chem. 1998, 273, 17451–17458. [Google Scholar] [CrossRef]
- Struglics, A.; Hansson, M.; Lohmander, L.S. Human aggrecanase generated synovial fluid fragment levels are elevated directly after knee injuries due to proteolysis both in the inter globular and chondroitin sulfate domains. Osteoarthr. Cartil. 2011, 19, 1047–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melin Furst, C.; Morgelin, M.; Vadstrup, K.; Heinegard, D.; Aspberg, A.; Blom, A.M. The C-type lectin of the aggrecan G3 domain activates complement. PLoS ONE 2013, 8, e61407. [Google Scholar] [CrossRef] [PubMed]
- Kosasih, H.J.; Last, K.; Rogerson, F.M.; Golub, S.B.; Gauci, S.J.; Russo, V.C.; Stanton, H.; Wilson, R.; Lamande, S.R.; Holden, P.; et al. A Disintegrin and Metalloproteinase with Thrombospondin Motifs-5 (ADAMTS-5) Forms Catalytically Active Oligomers. J. Biol. Chem. 2016, 291, 3197–3208. [Google Scholar] [CrossRef]
- Fosang, A.J.; Last, K.; Stanton, H.; Golub, S.B.; Little, C.B.; Brown, L.; Jackson, D.C. Neoepitope antibodies against MMP-cleaved and aggrecanase-cleaved aggrecan. Methods Mol. Biol. 2010, 622, 312–347. [Google Scholar]
- East, C.J.; Stanton, H.; Golub, S.B.; Rogerson, F.M.; Fosang, A.J. ADAMTS-5 deficiency does not block aggrecanolysis at preferred cleavage sites in the chondroitin sulphate-rich region of aggrecan. J. Biol. Chem. 2007, 282, 8632–8640. [Google Scholar] [CrossRef] [PubMed]
- Fosang, A.J.; Neame, P.J.; Hardingham, T.E.; Murphy, G.; Hamilton, J.A. Cleavage of cartilage proteoglycan between G1 and G2 domains by stromelysins. J. Biol. Chem. 1991, 266, 15579–15582. [Google Scholar]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| Mmp-13 F | TCAAGGCTATGCACACTGGT |
| Mmp-13 R | CACTATGGTCTTTTCAATGCCTAA |
| Mmp-11 F | CAGGCCAAAAGGTACACAGC |
| Mmp-11 R | CCTTTGAGGTTCCGTGTCTC |
| Adamts-5 F | ATGCAGCCATCCTGTTCAC |
| Adamts-5 R | CATTCCCAGGGTGTCACAT |
| Adamts-9 F | ACAGCCATCAGAGAGTGCAA |
| Adamts-9 R | TCCTACACAGTACTTCCCACCAT |
| Calpain 5 F | CTAGCCTCCGCTCCAGTG |
| Calpain 5 R | AAGAAGGGGAGGCACCTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rogerson, F.M.; Last, K.; Golub, S.B.; Gauci, S.J.; Stanton, H.; Bell, K.M.; Fosang, A.J. ADAMTS-9 in Mouse Cartilage Has Aggrecanase Activity That Is Distinct from ADAMTS-4 and ADAMTS-5. Int. J. Mol. Sci. 2019, 20, 573. https://doi.org/10.3390/ijms20030573
Rogerson FM, Last K, Golub SB, Gauci SJ, Stanton H, Bell KM, Fosang AJ. ADAMTS-9 in Mouse Cartilage Has Aggrecanase Activity That Is Distinct from ADAMTS-4 and ADAMTS-5. International Journal of Molecular Sciences. 2019; 20(3):573. https://doi.org/10.3390/ijms20030573
Chicago/Turabian StyleRogerson, Fraser M., Karena Last, Suzanne B. Golub, Stephanie J. Gauci, Heather Stanton, Katrina M. Bell, and Amanda J. Fosang. 2019. "ADAMTS-9 in Mouse Cartilage Has Aggrecanase Activity That Is Distinct from ADAMTS-4 and ADAMTS-5" International Journal of Molecular Sciences 20, no. 3: 573. https://doi.org/10.3390/ijms20030573