Insight into Nephrocan Function in Mouse Endoderm Patterning

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
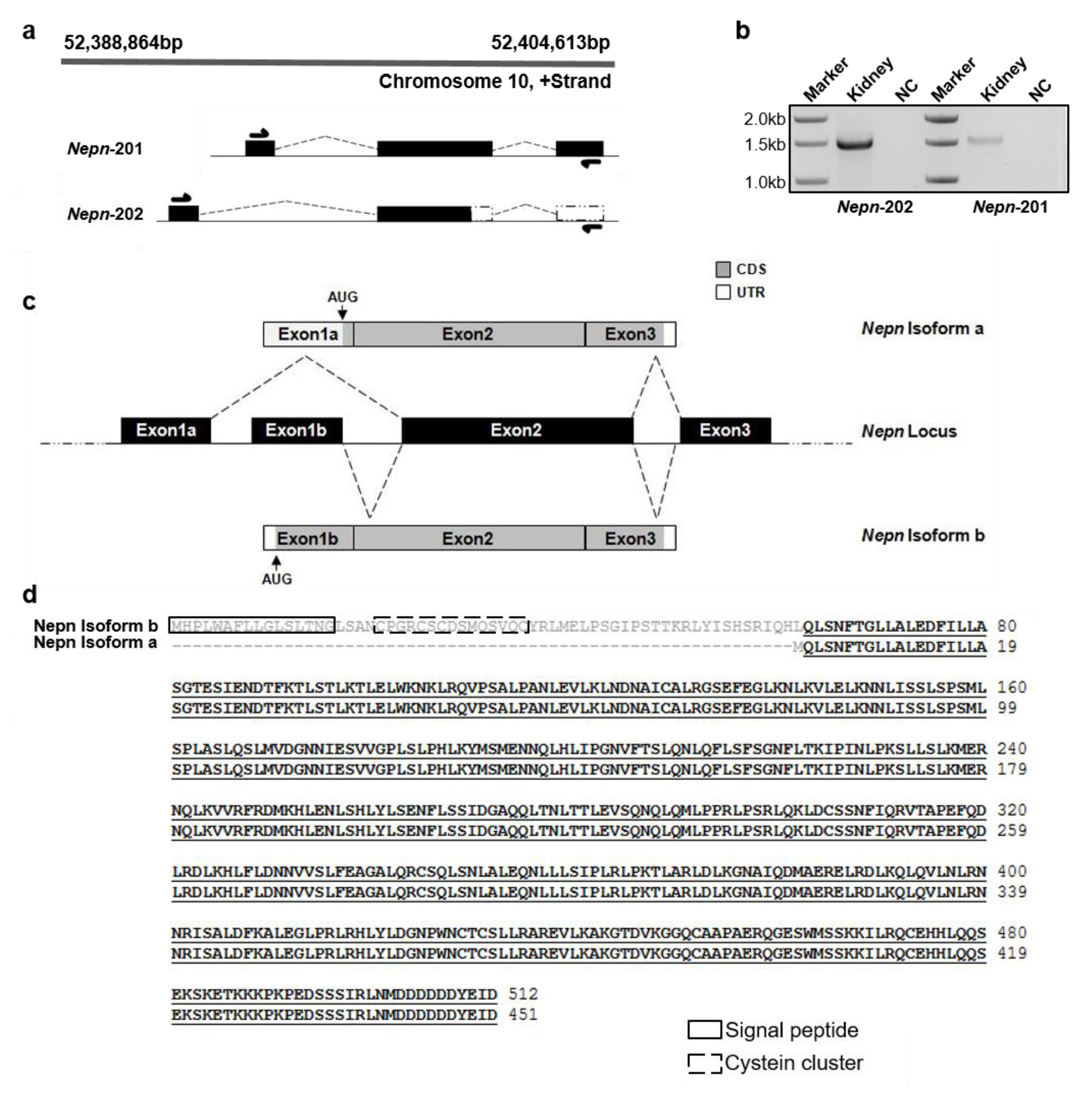
2.1. Evidence for a New Transcript Variant of Nephrocan in mouse
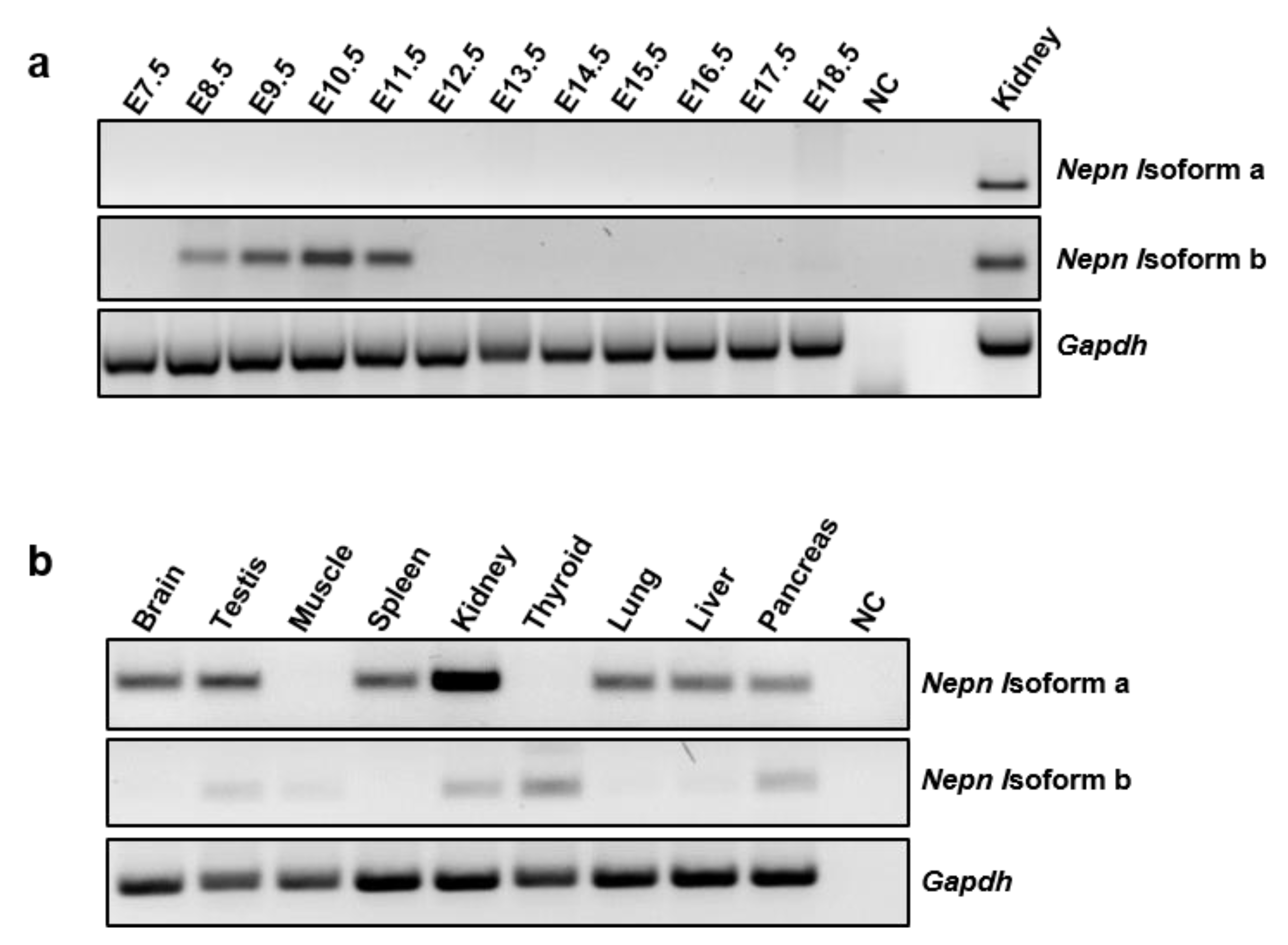
2.2. Nepn Isoforms Expression Pattern in Mouse Development and Tissues
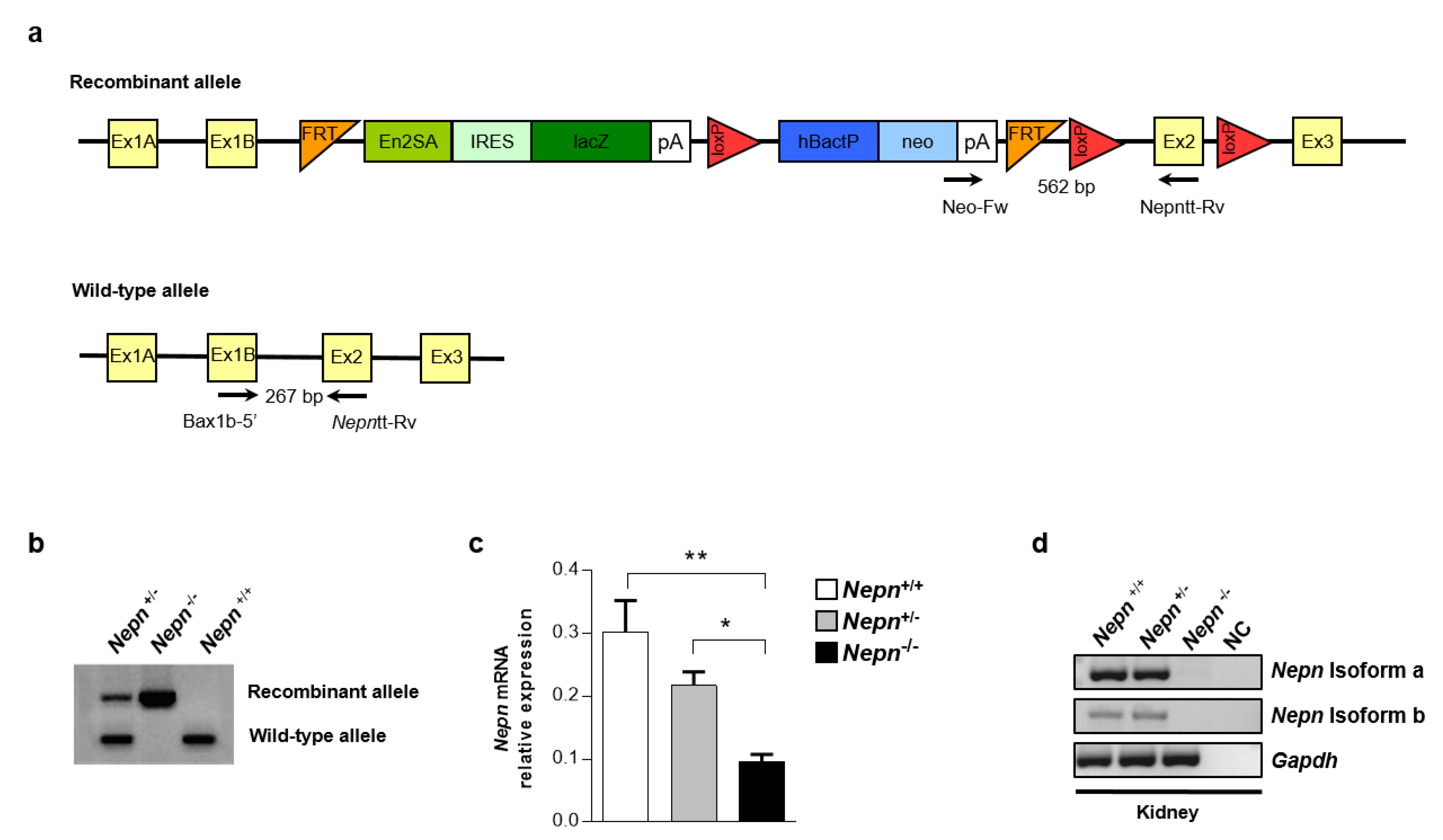
2.3. Nepn Knockout First Mouse (NepnKOfirst) Generation
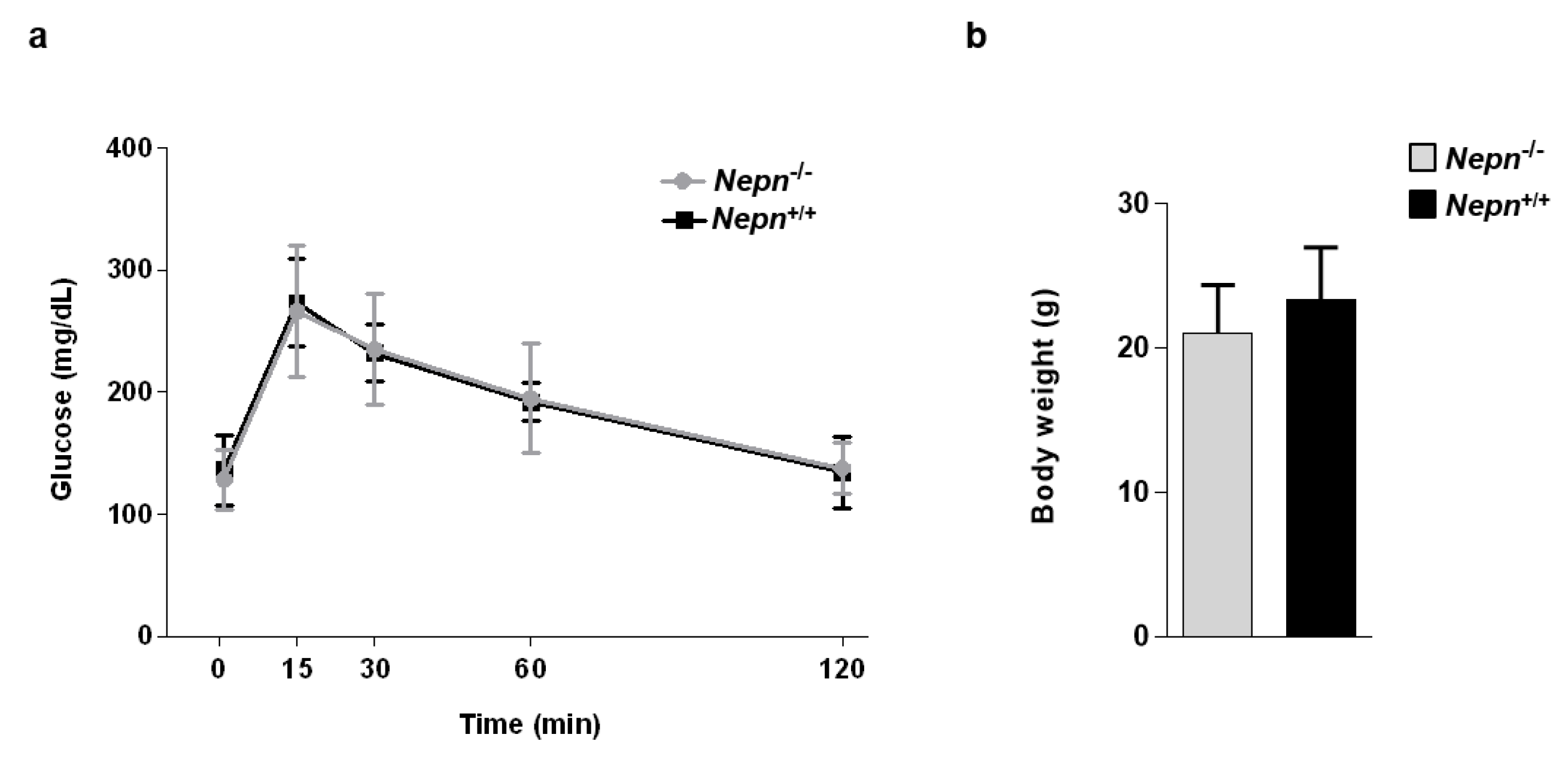
2.4. Characterization of Pancreatic and Renal Functionality
2.5. Generation of Nepn Deficient mESCs by RNA-Guided CRIPSR/Cas9
2.6. Nepn Decifiency Impairs Endoderm Lineage Commitment
3. Discussion
4. Materials and Methods
4.1. Ethics Statements and Animal Experiments
4.2. Generation of a Constitutive Nepn Knockout Mouse Line
4.3. Cell Culture and Differentiation Protocol
4.4. Vector Construction
4.5. CRISPR/Cas9 Genome Editing
4.6. RNA Extraction, RT-PCR Analysis and Quantitative Real Time RT–qPCR
4.7. Experimental Study for Renal Functionality
4.8. Intraperitoneal Glucose Tollerance Test (IPGTT)
4.9. Serum Analyses
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| B6 | C57BL/6J |
| CESA | Comitato Etico per la Sperimentazione Animale |
| DE | Definitive endoderm |
| E7.5 | Embryonic day 7.5 |
| ESCs | Embryonic stem cells |
| mESCs | Mouse embryonic stem cells |
| PS | Primitive streak |
| EMT | Epithelial-to-mesenchymal transition |
| MET | Mesenchymal-to-epithelial transition |
| VE | Visceral endoderm |
| PFE | Posterior foregut endoderm |
| DPB | Dorsal pancreatic bud |
| SLRP | Small leucine-rich repeat protein |
| IPGTT | Intraperitoneal glucose tolerance test |
| KO | Knock-out |
| Nepn | Nephrocan |
| WT | Wild-type |
| CRISPR/Cas9 | (CRISPR)/CRISPR-associated systems 9 |
| UTR | Untranslated region |
| RT-PCR | Reverse transcriptase-polymerase chain reaction |
| RT-qPCR | Real Time-quantitative polymerase chain reaction |
| SD | Standard Deviation |
| FRT | Flip-recombinase targets |
| PGK | Phosphoglycerate kinase 1 |
| PAM | Protospacer adjacent motif |
References
- Wells, J.M.; Melton, D.A. Vertebrate endoderm development. Annu. Rev. Cell Dev. Biol. 1999, 15, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Zorn, A.M.; Wells, J.M. Vertebrate endoderm development and organ formation. Annu. Rev. Cell Dev. Biol. 2009, 25, 221–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, P.P.; Behringer, R.R. Mouse gastrulation: The formation of a mammalian body plan. Mech. Dev. 1997, 68, 3–25. [Google Scholar] [CrossRef]
- Kubo, A.; Shinozaki, K.; Shannon, J.M.; Kouskoff, V.; Kennedy, M.; Woo, S.; Fehling, H.J.; Keller, G. Development of definitive endoderm from embryonic stem cells in culture. Development 2004, 131, 1651–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowotschin, S.; Hadjantonakis, A.K. Cellular dynamics in the early mouse embryo: From axis formation to gastrulation. Curr. Opin. Genet. Dev. 2010, 20, 420–427. [Google Scholar] [CrossRef] [Green Version]
- Acloque, H.; Adams, M.S.; Fishwick, K.; Bronner-Fraser, M.; Nieto, M.A. Epithelial-mesenchymal transitions: The importance of changing cell state in development and disease. J. Clin. Investig. 2009, 119, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Burtscher, I.; Lickert, H. Foxa2 regulates polarity and epithelialization in the endoderm germ layer of the mouse embryo. Development 2009, 136, 1029–1038. [Google Scholar] [CrossRef] [Green Version]
- Kwon, G.S.; Viotti, M.; Hadjantonakis, A.K. The endoderm of the mouse embryo arises by dynamic widespread intercalation of embryonic and extraembryonic lineages. Dev. Cell 2008, 15, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, Y.; Sheng, G. Epithelial to mesenchymal transition during gastrulation: An embryological view. Dev. Growth Differ. 2008, 50, 755–766. [Google Scholar] [CrossRef]
- Lewis, S.L.; Tam, P.P. Definitive endoderm of the mouse embryo: Formation, cell fates, and morphogenetic function. Dev. Dyn. 2006, 235, 2315–2329. [Google Scholar] [CrossRef]
- Liu, P.; Wakamiya, M.; Shea, M.J.; Albrecht, U.; Behringer, R.R.; Bradley, A. Requirement for Wnt3 in vertebrate axis formation. Nat. Genet. 1999, 22, 361–365. [Google Scholar] [CrossRef]
- Schier, A.F. Nodal signaling in vertebrate development. Annu. Rev. Cell Dev. Biol. 2003, 19, 589–621. [Google Scholar] [CrossRef]
- Vincent, S.D.; Dunn, N.R.; Hayashi, S.; Norris, D.P.; Robertson, E.J. Cell fate decisions within the mouse organizer are governed by graded Nodal signals. Genes Dev. 2003, 17, 1646–1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winnier, G.; Blessing, M.; Labosky, P.A.; Hogan, B.L. Bone morphogenetic protein-4 is required for mesoderm formation and patterning in the mouse. Genes Dev. 1995, 9, 2105–2116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Angelis, M.T.; Russo, F.; D’Angelo, F.; Federico, A.; Gemei, M.; Del Vecchio, L.; Ceccarelli, M.; De Felice, M.; Falco, G. Novel pancreas organogenesis markers refine the pancreatic differentiation roadmap of embryonic stem cells. Stem. Cell Rev. 2014, 10, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Iozzo, R.V. Matrix proteoglycans: From molecular design to cellular function. Annu. Rev. Biochem. 1998, 67, 609–652. [Google Scholar] [CrossRef] [Green Version]
- Mochida, Y.; Parisuthiman, D.; Kaku, M.; Hanai, J.; Sukhatme, V.P.; Yamauchi, M. Nephrocan, a novel member of the small leucine-rich repeat protein family, is an inhibitor of transforming growth factor-beta signaling. J. Biol. Chem. 2006, 281, 36044–36051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, J.; Wei, W.; Saund, R.S.; Xiang, P.; Cunningham, T.J.; Yi, Y.; Alder, O.; Lu, D.Y.; Savory, J.G.; Krentz, N.A.; et al. A regulatory network controls nephrocan expression and midgut patterning. Development 2014, 141, 3772–3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sander, J.D.; Joung, J.K. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat. Biotechnol. 2014, 32, 347–355. [Google Scholar] [CrossRef]
- Kumar, T.R.; Larson, M.; Wang, H.; McDermott, J.; Bronshteyn, I. Transgenic mouse technology: Principles and methods. Methods Mol. Biol. 2009, 590, 335–362. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.H.; Silva, F.J.; Tsark, W.M.; Mann, J.R. A Cre/loxP-deleter transgenic line in mouse strain 129S1/SvImJ. Genesis 2002, 32, 199–202. [Google Scholar] [CrossRef]
- Ayala, J.E.; Samuel, V.T.; Morton, G.J.; Obici, S.; Croniger, C.M.; Shulman, G.I.; Wasserman, D.H.; McGuinness, O.P.; Consortium, N.M.M.P.C. Standard operating procedures for describing and performing metabolic tests of glucose homeostasis in mice. Dis. Model. Mech. 2010, 3, 525–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauck, M.A.; Meier, J.J. Incretin hormones: Their role in health and disease. Diabetes Obes. Metab. 2018, 20, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, A.; Romarís, M.; Rasmussen, L.M.; Heinegård, D.; Twardzik, D.R.; Border, W.A.; Ruoslahti, E. Interaction of the small interstitial proteoglycans biglycan, decorin and fibromodulin with transforming growth factor beta. Biochem. J. 1994, 302, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Kodama, Y.; Matsumoto, T. Bone matrix decorin binds transforming growth factor-beta and enhances its bioactivity. J. Biol. Chem. 1994, 269, 32634–32638. [Google Scholar]
- Qin, P.; Haberbusch, J.M.; Zhang, Z.; Soprano, K.J.; Soprano, D.R. Pre-B cell leukemia transcription factor (PBX) proteins are important mediators for retinoic acid-dependent endodermal and neuronal differentiation of mouse embryonal carcinoma P19 cells. J. Biol. Chem. 2004, 279, 16263–16271. [Google Scholar] [CrossRef] [Green Version]
- Tasheva, E.S.; Klocke, B.; Conrad, G.W. Analysis of transcriptional regulation of the small leucine rich proteoglycans. Mol. Vis. 2004, 10, 758–772. [Google Scholar]
- Chia, C.Y.; Madrigal, P.; Denil, S.L.I.J.; Martinez, I.; Garcia-Bernardo, J.; El-Khairi, R.; Chhatriwala, M.; Shepherd, M.H.; Hattersley, A.T.; Dunn, N.R.; et al. GATA6 Cooperates with EOMES/SMAD2/3 to Deploy the Gene Regulatory Network Governing Human Definitive Endoderm and Pancreas Formation. Stem. Cell Rep. 2019, 12, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; He, A.; Pu, W.T. Regulation of GATA4 transcriptional activity in cardiovascular development and disease. Curr. Top Dev. Biol. 2012, 100, 143–169. [Google Scholar] [CrossRef]
- Morrisey, E.E.; Tang, Z.; Sigrist, K.; Lu, M.M.; Jiang, F.; Ip, H.S.; Parmacek, M.S. GATA6 regulates HNF4 and is required for differentiation of visceral endoderm in the mouse embryo. Genes Dev. 1998, 12, 3579–3590. [Google Scholar] [CrossRef] [Green Version]
- Xuan, S.; Sussel, L. GATA4 and GATA6 regulate pancreatic endoderm identity through inhibition of hedgehog signaling. Development 2016, 143, 780–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C (T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Lucci, V.; Di Palma, T.; Zannini, M. Neuropilin-2 Is a Newly Identified Target of PAX8 in Thyroid Cells. PLoS ONE 2015, 10, e0128315. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n = 86 | Nepn+/+ | Nepn+/− | Nepn−/− | p-value |
|---|---|---|---|---|
| Observed frequency | 29.1% | 53.5% | 17.4% | 0.2 |
| Expected frequency | 25% | 50% | 25% |
| Urinary Parameters | Nepn+/+ | Nepn−/− |
|---|---|---|
| Urinary Volume (mL) | 1.86 ± 0.37 | 1.63 ± 0.25 |
| Creatinine excretion (µmol/g body weight) | 0.24 ± 0.03 | 0.23 ± 0.01 |
| Na+/creatinine | 22.75 ± 4.16 | 27.92 ± 4.41 |
| K+/creatinine | 20.36 ± 3.95 | 18.14 ± 2.23 |
| Cl−/creatinine | 62.91 ± 6.43 | 72.34 ± 3.43 |
| Creatinine clearence | 113.16 ± 14.91 | 97.56 ± 11.33 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Addeo, M.; Buonaiuto, S.; Guerriero, I.; Amendola, E.; Visconte, F.; Marino, A.; De Angelis, M.T.; Russo, F.; Roberto, L.; Marotta, P.; et al. Insight into Nephrocan Function in Mouse Endoderm Patterning. Int. J. Mol. Sci. 2020, 21, 8. https://doi.org/10.3390/ijms21010008
Addeo M, Buonaiuto S, Guerriero I, Amendola E, Visconte F, Marino A, De Angelis MT, Russo F, Roberto L, Marotta P, et al. Insight into Nephrocan Function in Mouse Endoderm Patterning. International Journal of Molecular Sciences. 2020; 21(1):8. https://doi.org/10.3390/ijms21010008
Chicago/Turabian StyleAddeo, Martina, Silvia Buonaiuto, Ilaria Guerriero, Elena Amendola, Feliciano Visconte, Antonio Marino, Maria Teresa De Angelis, Filomena Russo, Luca Roberto, Pina Marotta, and et al. 2020. "Insight into Nephrocan Function in Mouse Endoderm Patterning" International Journal of Molecular Sciences 21, no. 1: 8. https://doi.org/10.3390/ijms21010008