Maternal Obesity Alters Placental Cell Cycle Regulators in the First Trimester of Human Pregnancy: New Insights for BRCA1

 , , and
, , and
Abstract
:1. Introduction
2. Results
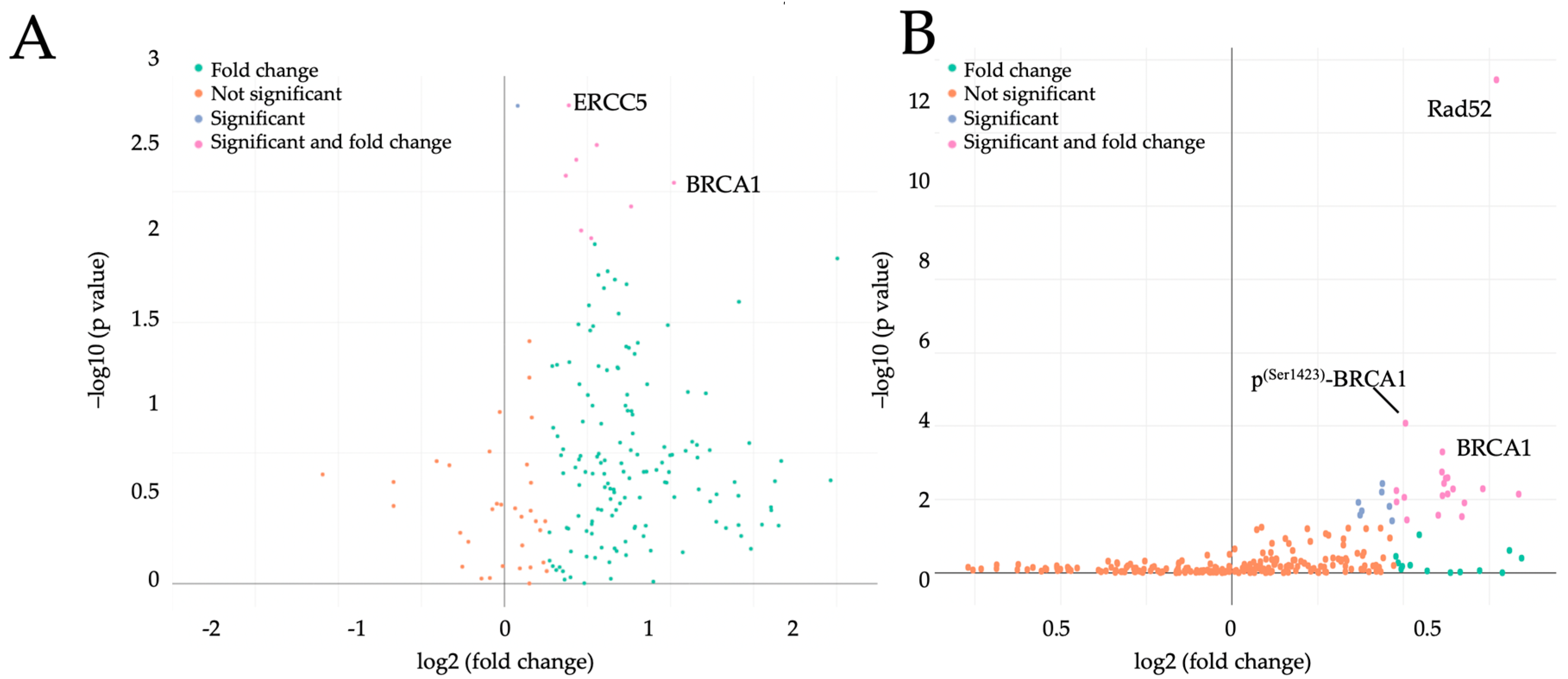
2.1. Maternal Obesity Affects Placental Cell Cycle Regulators Already in the First Trimester of Pregnancy
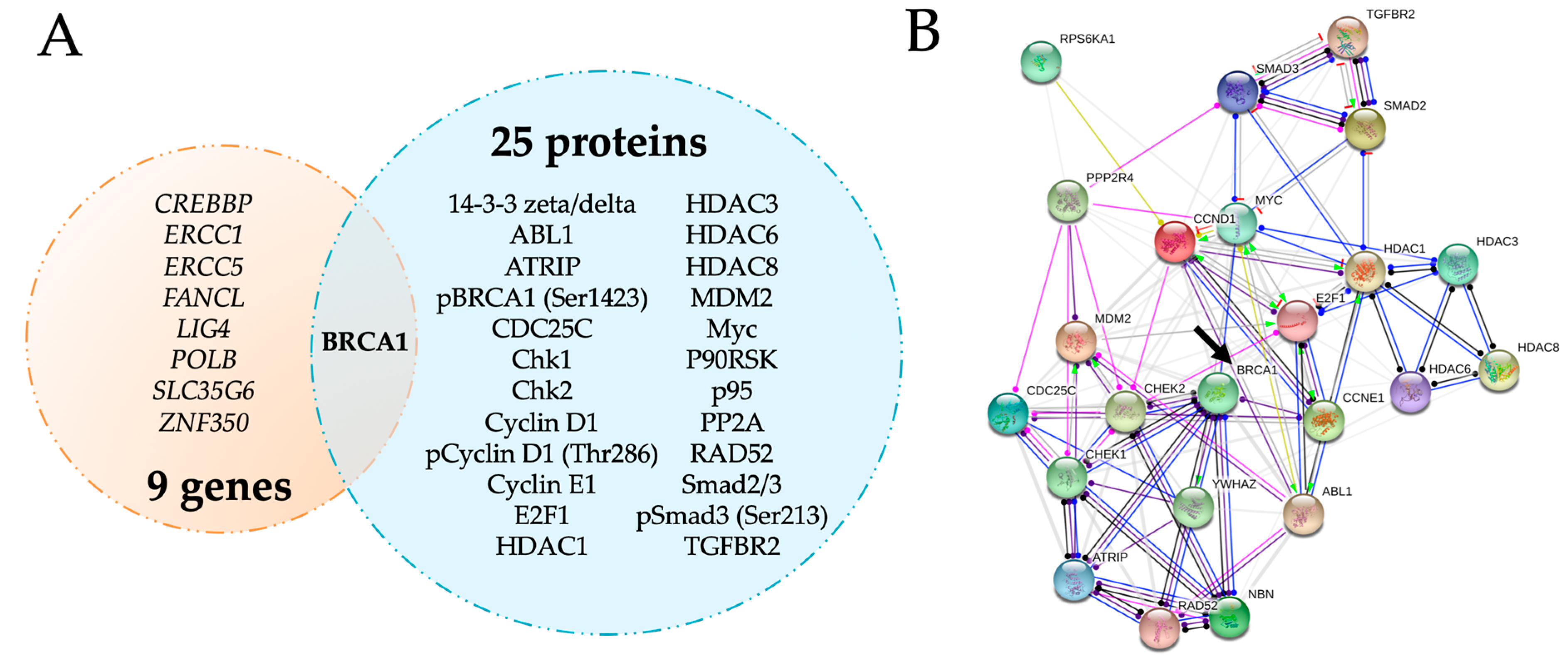
2.2. BRCA1 Is a Key Player in Cell Cycle Regulation in Early Pregnancy and Is Upregulated by Maternal Obesity
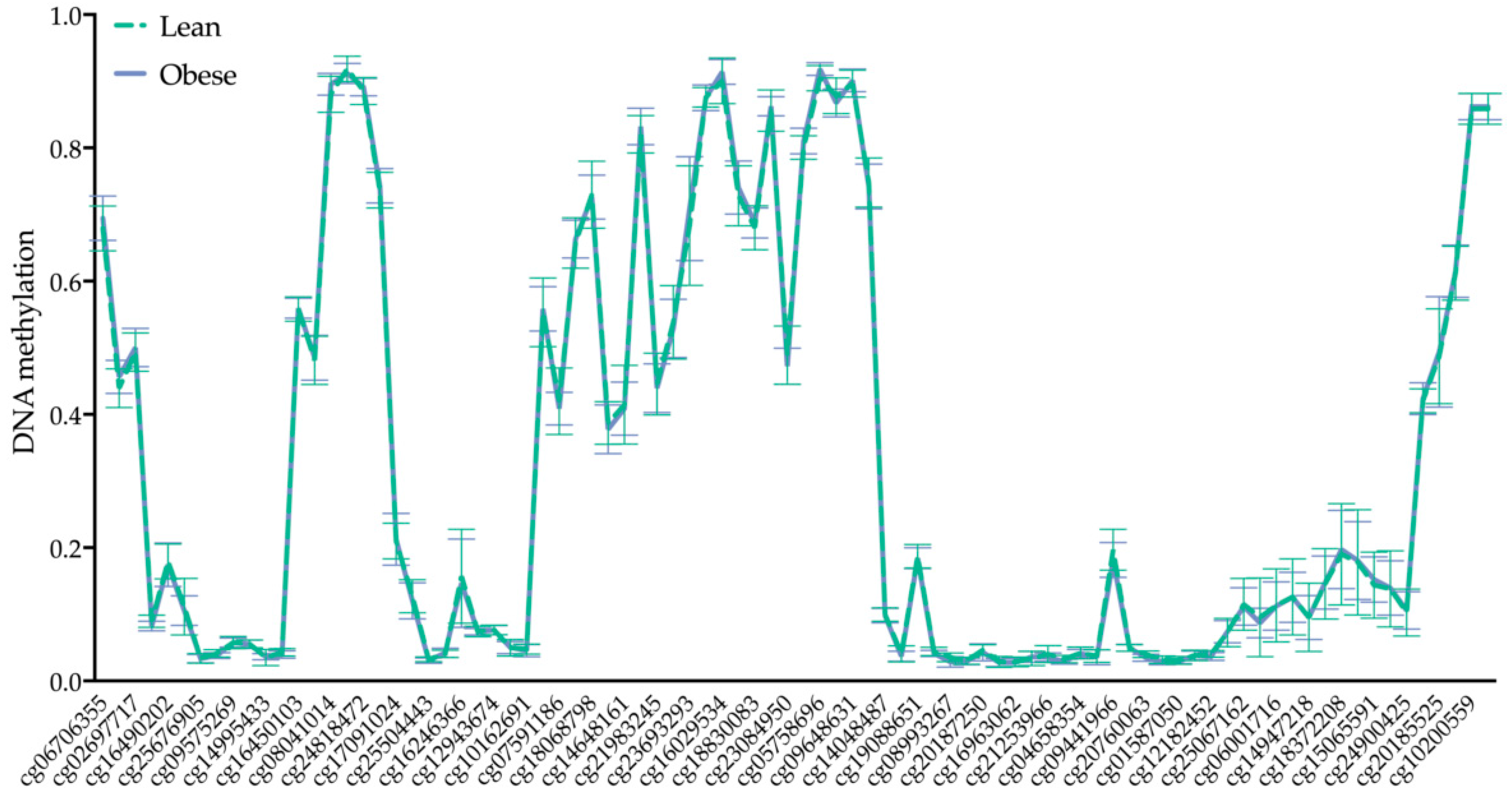
2.3. BRCA1 DNA Methylation Is Not Altered by Maternal Obesity
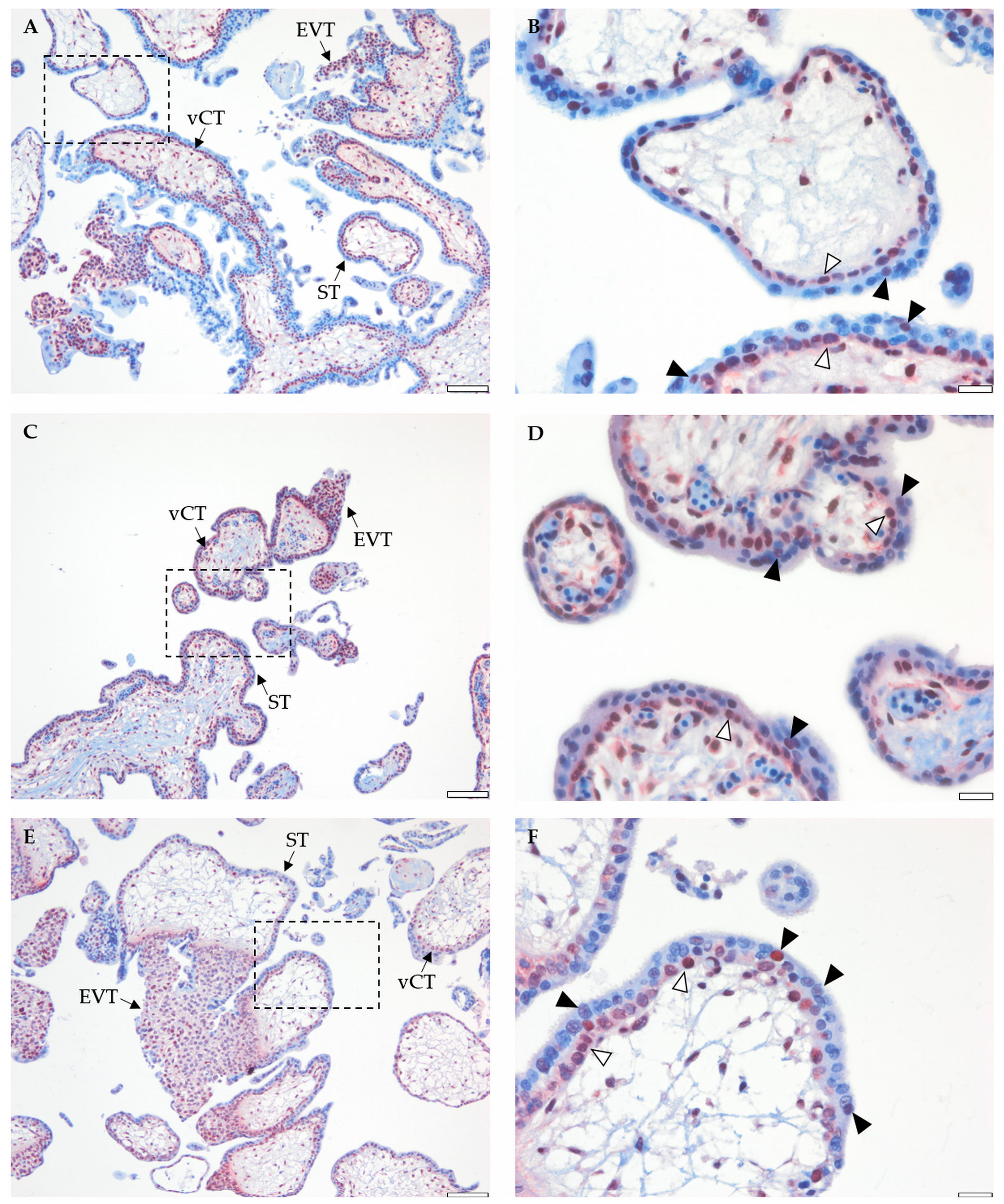
2.4. Placental BRCA1 Is Mainly Localized to vCTs and EVTs during the First Trimester of Pregnancy
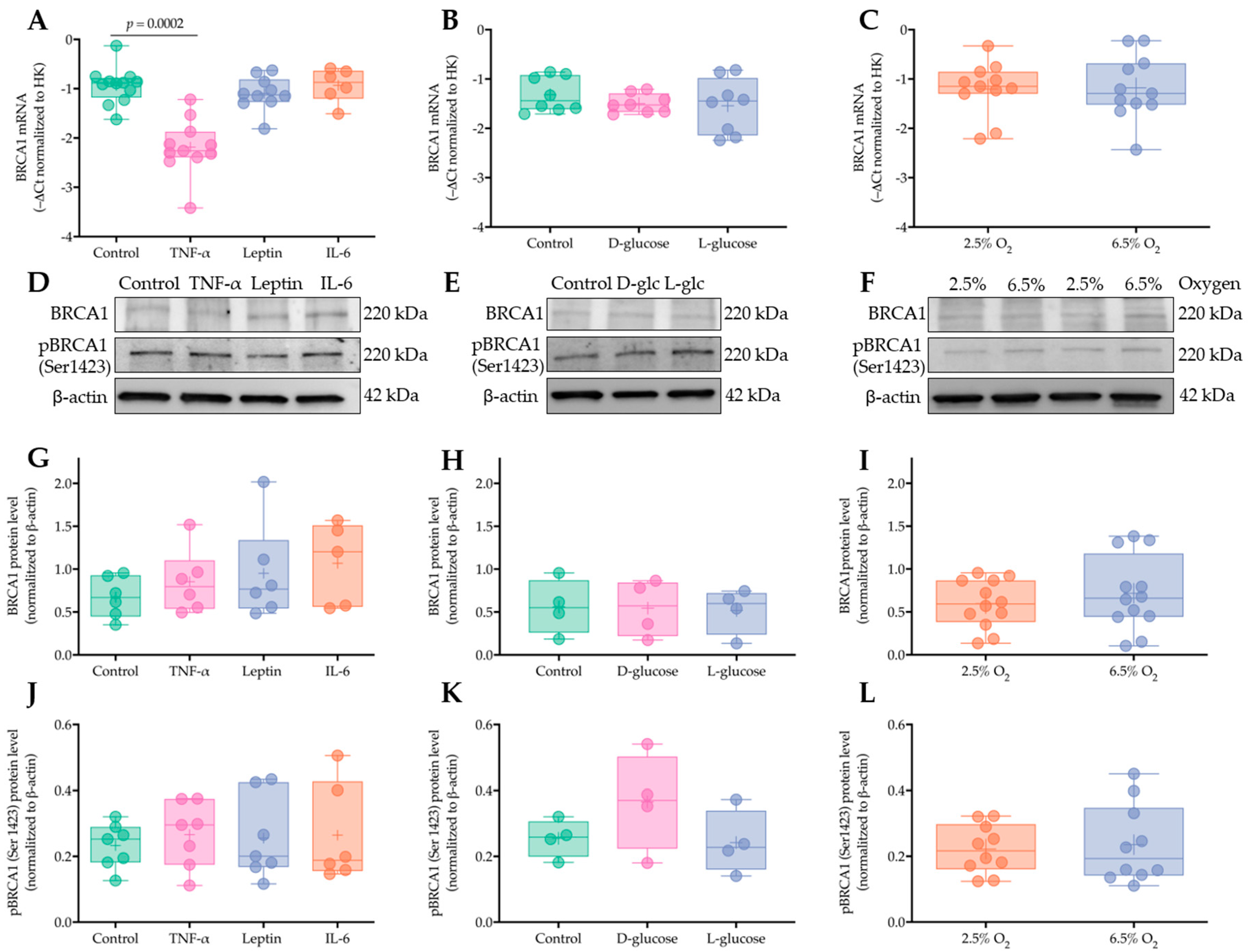
2.5. Short Term Exposure to Obesity-Associated Cytokines, Hyperglycemia, and Increased Oxygen Tension Do Not Regulate Placental BRCA1 Levels in Early Pregnancy
3. Discussion
4. Materials and Methods
4.1. Study Subjects
4.2. Human Placental Tissue Collection
4.3. First Trimester Chorionic Villous Explants
4.4. DNA/RNA Isolation and Reverse Transcription
4.5. PrimePCR Panel
4.6. Real Time PCR
4.7. Nanostring
4.8. Protein Isolation and Quantification
4.9. Cell Cycle Control Protein Array
4.10. Immunoblotting
4.11. Immunohistochemistry
4.12. DNA Methylation Profiling
4.13. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATM | Ataxia telangiectasia mutated kinase |
| BCA | Bicinchoninic acid assay |
| BMI | Body mass index |
| BRCA1 | Breast cancer 1 |
| CRL | Crown-rump length |
| DMEM | Dulbecco’s Modified Eagle Medium |
| EVTs | Extravillous trophoblasts |
| FC | Fold change |
| hCG | Human chorionic gonadotropin |
| HPRT1 | Hypoxanthine phosphoribosyltransferase 1 |
| IHC | Immunohistochemistry |
| IL-6 | Interleukin 6 |
| MVLR | Multivariate linear regression |
| PFA | Paraformaldehyde |
| PNKP | Polynucleotide kinase 3′-phosphatase |
| PPIA | Peptidylprolyl isomerase A |
| SD | Standard deviation |
| ST | Syncytiotrophoblast |
| T1D | Type-1 diabetes |
| TBP | TATA box binding protein |
| TNF-α | Tumor necrosis factor α |
| vCTs | Villous cytotrophoblasts |
| WDR45L | WD repeat domain 45B |
References
- Staud, F.; Karahoda, R. Trophoblast: The central unit of fetal growth, protection and programming. Int. J. Biochem. Cell Biol. 2018, 105, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Knofler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.; James, J. Human placenta and trophoblast development: Key molecular mechanisms and model systems. Cell. Mol. Life Sci. CMLS 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knofler, M. Critical growth factors and signalling pathways controlling human trophoblast invasion. Int. J. Dev. Biol. 2010, 54, 269–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moser, G.; Weiss, G.; Sundl, M.; Gauster, M.; Siwetz, M.; Lang-Olip, I.; Huppertz, B. Extravillous trophoblasts invade more than uterine arteries: Evidence for the invasion of uterine veins. Histochem. Cell Biol. 2017, 147, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, P.; Black, S.; Huppertz, B. Endovascular trophoblast invasion: Implications for the pathogenesis of intrauterine growth retardation and preeclampsia. Biol. Reprod. 2003, 69, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knofler, M.; Pollheimer, J. Human placental trophoblast invasion and differentiation: A particular focus on Wnt signaling. Front. Genet. 2013, 4, 190. [Google Scholar] [CrossRef] [Green Version]
- Gude, N.M.; Roberts, C.T.; Kalionis, B.; King, R.G. Growth and function of the normal human placenta. Thrombosis Res. 2004, 114, 397–407. [Google Scholar] [CrossRef]
- Hu, K.-L.; Chang, H.-M.; Zhao, H.-C.; Yu, Y.; Li, R.; Qiao, J. Potential roles for the kisspeptin/kisspeptin receptor system in implantation and placentation. Hum. Reprod. Update 2019, 25, 326–343. [Google Scholar] [CrossRef]
- Burton, G.J.; Jauniaux, E.; Charnock-Jones, D.S. The influence of the intrauterine environment on human placental development. Int. J. Dev. Biol. 2010, 54, 303–312. [Google Scholar] [CrossRef] [Green Version]
- Majali-Martinez, A.; Velicky, P.; Pollheimer, J.; Knofler, M.; Yung, H.W.; Burton, G.J.; Tabrizi-Wizsy, N.G.; Lang, U.; Hiden, U.; Desoye, G.; et al. Endothelin-1 down-regulates matrix metalloproteinase 14 and 15 expression in human first trimester trophoblasts via endothelin receptor type B. Hum. Reprod. 2017, 32, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Majali-Martinez, A.; Barth, S.; Lang, U.; Desoye, G.; Cervar-Zivkovic, M. Temporal changes of the endothelin system in human cytotrophoblasts during the first trimester of pregnancy. Physiol. Res. 2018, 67, S247–S255. [Google Scholar] [CrossRef]
- Hiden, U.; Glitzner, E.; Ivanisevic, M.; Djelmis, J.; Wadsack, C.; Lang, U.; Desoye, G. MT1-MMP expression in first-trimester placental tissue is upregulated in type 1 diabetes as a result of elevated insulin and tumor necrosis factor-alpha levels. Diabetes 2008, 57, 150–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.W.; Wakeland, A.K.; Parast, M.M. Trophoblast lineage specification, differentiation and their regulation by oxygen tension. J. Endocrinol. 2018, 236, R43–R56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.J.; Jauniaux, E.; Murray, A.J. Oxygen and placental development; parallels and differences with tumour biology. Placenta 2017, 56, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Hoch, D.; Gauster, M.; Hauguel-de Mouzon, S.; Desoye, G. Diabesity-associated oxidative and inflammatory stress signalling in the early human placenta. Mol. Asp. Med. 2019, 66, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Pantham, P.; Aye, I.L.; Powell, T.L. Inflammation in maternal obesity and gestational diabetes mellitus. Placenta 2015, 36, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Xu, X.; Yan, Y. Estimated global overweight and obesity burden in pregnant women based on panel data model. PLoS ONE 2018, 13, e0202183. [Google Scholar] [CrossRef] [Green Version]
- Jeyabalan, A. Epidemiology of preeclampsia: Impact of obesity. Nutr. Rev. 2013, 71 (Suppl. 1), S18–S25. [Google Scholar] [CrossRef] [Green Version]
- Bautista-Castano, I.; Henriquez-Sanchez, P.; Aleman-Perez, N.; Garcia-Salvador, J.J.; Gonzalez-Quesada, A.; Garcia-Hernandez, J.A.; Serra-Majem, L. Maternal obesity in early pregnancy and risk of adverse outcomes. PLoS ONE 2013, 8, e80410. [Google Scholar] [CrossRef] [Green Version]
- Lowe, W.L., Jr.; Bain, J.R.; Nodzenski, M.; Reisetter, A.C.; Muehlbauer, M.J.; Stevens, R.D.; Ilkayeva, O.R.; Lowe, L.P.; Metzger, B.E.; Newgard, C.B.; et al. Maternal BMI and Glycemia Impact the Fetal Metabolome. Diabetes Care 2017, 40, 902–910. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M.; Shankar, K. Obesity and pregnancy: Mechanisms of short term and long term adverse consequences for mother and child. BMJ (Clin. Res.) 2017, 356, j1. [Google Scholar] [CrossRef] [PubMed]
- Velicky, P.; Meinhardt, G.; Plessl, K.; Vondra, S.; Weiss, T.; Haslinger, P.; Lendl, T.; Aumayr, K.; Mairhofer, M.; Zhu, X.; et al. Genome amplification and cellular senescence are hallmarks of human placenta development. PLoS Genet. 2018, 14, e1007698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lassance, L.; Haghiac, M.; Leahy, P.; Basu, S.; Minium, J.; Zhou, J.; Reider, M.; Catalano, P.M.; Hauguel-de Mouzon, S. Identification of early transcriptome signatures in placenta exposed to insulin and obesity. Am. J. Obstet. Gynecol. 2015, 212, 647.e1–647.e611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korgun, E.T.; Celik-Ozenci, C.; Acar, N.; Cayli, S.; Desoye, G.; Demir, R. Location of cell cycle regulators cyclin B1, cyclin A, PCNA, Ki67 and cell cycle inhibitors p21, p27 and p57 in human first trimester placenta and deciduas. Histochem. Cell Biol. 2006, 125, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Genbacev, O.; McMaster, M.T.; Fisher, S.J. A repertoire of cell cycle regulators whose expression is coordinated with human cytotrophoblast differentiation. Am. J. Pathol. 2000, 157, 1337–1351. [Google Scholar] [CrossRef] [Green Version]
- Mitsuya, K.; Parker, A.N.; Liu, L.; Ruan, J.; Vissers, M.C.M.; Myatt, L. Alterations in the placental methylome with maternal obesity and evidence for metabolic regulation. PLoS ONE 2017, 12, e0186115. [Google Scholar] [CrossRef] [Green Version]
- Unek, G.; Ozmen, A.; Isenlik, B.S.; Korgun, E.T. The proliferation mechanism of normal and pathological human placentas. Histol. Histopathol. 2017, 32, 339–349. [Google Scholar] [CrossRef]
- Wlodarczyk, M.; Nowicka, G. Obesity, DNA Damage, and Development of Obesity-Related Diseases. Int. J. Mol. Sci. 2019, 20, 1146. [Google Scholar] [CrossRef]
- Lopez-Jaramillo, P.; Barajas, J.; Rueda-Quijano, S.M.; Lopez-Lopez, C.; Felix, C. Obesity and Preeclampsia: Common Pathophysiological Mechanisms. Front. Physiol. 2018, 9, 1838. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Z.; Kitagawa, M.; Takemura, T.; Hirokawa, K. Effect of maternal age on incidences of apoptotic and proliferative cells in trophoblasts of full-term human placenta. Mol. Hum. Reprod. 2001, 7, 1179–1185. [Google Scholar] [CrossRef] [Green Version]
- Lean, S.C.; Heazell, A.E.P.; Dilworth, M.R.; Mills, T.A.; Jones, R.L. Placental Dysfunction Underlies Increased Risk of Fetal Growth Restriction and Stillbirth in Advanced Maternal Age Women. Sci. Rep. 2017, 7, 9677. [Google Scholar] [CrossRef] [PubMed]
- Varol, U.; Kucukzeybek, Y.; Alacacioglu, A.; Somali, I.; Altun, Z.; Aktas, S.; Oktay Tarhan, M. BRCA genes: BRCA 1 and BRCA 2. Apoptosis 2018, 13, 19. [Google Scholar]
- Krieger, K.L.; Hu, W.-F.; Ripperger, T.; Woods, N.T. Functional Impacts of the BRCA1-mTORC2 Interaction in Breast Cancer. Int. J. Mol. Sci. 2019, 20, 5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, K.I.; Gorski, J.J.; Barros, E.M.; Irwin, G.W.; Manti, L.; Powell, A.J.; Pellagatti, A.; Lukashchuk, N.; McCance, D.J.; McCluggage, W.G.; et al. Identification of a BRCA1-mRNA splicing complex required for efficient DNA repair and maintenance of genomic stability. Mol. Cell 2014, 54, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.X. BRCA1: Cell cycle checkpoint, genetic instability, DNA damage response and cancer evolution. Nucleic Acids Res. 2006, 34, 1416–1426. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Lu, L.-Y.; Yu, X. The role of BRCA1 in DNA damage response. Protein Cell 2010, 1, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316. [Google Scholar] [CrossRef]
- Bartek, J.; Lukas, J. Chk1 and Chk2 kinases in checkpoint control and cancer. Cancer Cell 2003, 3, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, T. BRCA1 phosphorylation: Biological consequences. Cancer Biol. Ther. 2006, 5, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Okada, S.; Ouchi, T. Cell cycle differences in DNA damage-induced BRCA1 phosphorylation affect its subcellular localization. J. Biol. Chem. 2003, 278, 2015–2020. [Google Scholar] [CrossRef] [Green Version]
- West, R.C.; McWhorter, E.S.; Ali, A.; Goetzman, L.N.; Russ, J.E.; Gonzalez-Berrios, C.L.; Anthony, R.V.; Bouma, G.J.; Winger, Q.A. HMGA2 is regulated by LIN28 and BRCA1 in human placental cells. Biol. Reprod. 2019, 100, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, R.C.; Russ, J.E.; Bouma, G.J.; Winger, Q.A. BRCA1 regulates HMGA2 levels in the Swan71 trophoblast cell line. Mol. Reprod. Dev. 2019, 86, 1663–1670. [Google Scholar] [CrossRef] [PubMed]
- Gauster, M.; Maninger, S.; Siwetz, M.; Deutsch, A.; El-Heliebi, A.; Kolb-Lenz, D.; Hiden, U.; Desoye, G.; Herse, F.; Prokesch, A. Downregulation of p53 drives autophagy during human trophoblast differentiation. Cell. Mol. Life Sci. CMLS 2018, 75, 1839–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldman-Wohl, D.; Yagel, S. United we stand not dividing: The syncytiotrophoblast and cell senescence. Placenta 2014, 35, 341–344. [Google Scholar] [CrossRef]
- Henderson, B.R. The BRCA1 Breast Cancer Suppressor: Regulation of Transport, Dynamics, and Function at Multiple Subcellular Locations. Scientifica 2012, 2012, 796808. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Macias, V.; Al-Alem, U.; Deaton, R.J.; Kadjaksy-Balla, A.; Gann, P.H.; Rauscher, G.H. BRCA1 protein expression and subcellular localization in primary breast cancer: Automated digital microscopy analysis of tissue microarrays. PLoS ONE 2017, 12, e0184385. [Google Scholar] [CrossRef] [Green Version]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef] [Green Version]
- Nadhan, R.; Vaman, J.V.; Sengodan, S.K.; Hemalatha, S.K.; Nirmala, C.; Sadasivan, S.; Aysha, P.V.; Yesodharan, S.; Krishnapriya, R.S.; Amritha Krishna, B.V.; et al. BRCA1 promoter hypermethylation in human placenta: A hidden link with beta-hCG expression. In Carcinogenesis; Oxford University Press: Oxford, UK, 2019. [Google Scholar] [CrossRef]
- Pendeloski, K.P.T.; Ono, E.; Torloni, M.R.; Mattar, R.; Daher, S. Maternal obesity and inflammatory mediators: A controversial association. Am. J. Reprod. Immunol. 2017, 77, e12674. [Google Scholar] [CrossRef] [Green Version]
- Silva, C.; Nunes, C.; Correia-Branco, A.; Araújo, J.R.; Martel, F. Insulin Exhibits an Antiproliferative and Hypertrophic Effect in First Trimester Human Extravillous Trophoblasts. Reprod. Sci. 2017, 24, 582–594. [Google Scholar] [CrossRef]
- Higgins, L.; Greenwood, S.L.; Wareing, M.; Sibley, C.P.; Mills, T.A. Obesity and the placenta: A consideration of nutrient exchange mechanisms in relation to aberrant fetal growth. Placenta 2011, 32, 1–7. [Google Scholar] [CrossRef]
- Scott-Drechsel, D.E.; Rugonyi, S.; Marks, D.L.; Thornburg, K.L.; Hinds, M.T. Hyperglycemia slows embryonic growth and suppresses cell cycle via cyclin D1 and p21. Diabetes 2013, 62, 234–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordeleau, L.; Lipscombe, L.; Lubinski, J.; Ghadirian, P.; Foulkes, W.D.; Neuhausen, S.; Ainsworth, P.; Pollak, M.; Sun, P.; Narod, S.A. Diabetes and breast cancer among women with BRCA1 and BRCA2 mutations. Cancer 2011, 117, 1812–1818. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Murase, T. Obesity-associated insulin resistance adversely affects skin function. PLoS ONE 2019, 14, e0223528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jauniaux, E.; Watson, A.L.; Hempstock, J.; Bao, Y.P.; Skepper, J.N.; Burton, G.J. Onset of maternal arterial blood flow and placental oxidative stress. A possible factor in human early pregnancy failure. Am. J. Pathol. 2000, 157, 2111–2122. [Google Scholar] [CrossRef]
- Li, L.; Cohen, M.; Wu, J.; Sow, M.H.; Nikolic, B.; Bischof, P.; Irminger-Finger, I. Identification of BARD1 splice-isoforms involved in human trophoblast invasion. Int. J. Biochem. Cell Biol. 2007, 39, 1659–1672. [Google Scholar] [CrossRef]
- Benowitz, N.L.; Bernert, J.T.; Caraballo, R.S.; Holiday, D.B.; Wang, J. Optimal serum cotinine levels for distinguishing cigarette smokers and nonsmokers within different racial/ethnic groups in the United States between 1999 and 2004. Am. J. Epidemiol. 2009, 169, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Strutz, J.; Cvitic, S.; Hackl, H.; Kashofer, K.; Appel, H.M.; Thüringer, A.; Desoye, G.; Koolwijk, P.; Hiden, U. Gestational diabetes alters microRNA signatures in human feto-placental endothelial cells depending on fetal sex. Clin. Sci. 2018, 132, 2437–2449. [Google Scholar] [CrossRef]
- Maksimovic, J.; Gordon, L.; Oshlack, A. SWAN: Subset-quantile within array normalization for illumina infinium HumanMethylation450 BeadChips. Genome Biol. 2012, 13, R44. [Google Scholar] [CrossRef] [Green Version]
- Phipson, B.; Maksimovic, J.; Oshlack, A. missMethyl: An R package for analyzing data from Illumina’s HumanMethylation450 platform. Bioinformatics 2016, 32, 286–288. [Google Scholar] [CrossRef]
- The R Development Core Team. R: A Language and Environment for Statistical Computing; Scientific Research Publishing: Wuhan, China, 2018; Volume 1, p. 34. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Baptiste, A. gridExtra: Miscellaneous Functions for “Grid” Graphics. R Package Version 2.3; 2017; Available online: https://cran.r-project.org/web/packages/gridExtra/index.html (accessed on 10 December 2019).
- Plotly Technologies, Inc. Collaborative Data Science; Plotly Technologies, Inc.: Montréal, QC, Canada, 2015. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | PCR Panel and Protein Array | Methylation | IHC | Explants | ||||
|---|---|---|---|---|---|---|---|---|
| Lean | Obese | p | Lean | Obese | p | Lean | Lean | |
| Sample size (n) | 7 | 6 | 15 | 15 | 4 | 11 | ||
| Gestational age (days) | 49.0 ± 0.0 | 49.8 ± 1.8 | 0.3 | 57.3 ± 13.5 | 56.1 ± 13.1 | 0.7 | 53.0 ± 17.5 | 51.9 ± 11.9 |
| Maternal age (years) | 33.0 ± 5.8 | 36.7 ± 5.3 | 0.3 | 27.8 ± 7.0 | 30.3 ± 6.0 | 0.3 | 31.7 ± 9 | 28.2 ± 7.4 |
| Maternal BMI (kg/m2) | 20.3 ± 1.6 | 30.3 ± 2.3 | <0.0001 | 22.2 ± 1.6 | 34.3 ± 3.4 | <0.0001 | 24.5 ± 3.3 | 21.9 ± 3.3 |
| Fetal sex | 4 m; 3 f | 3 m; 3 f | 6 m; 9 f | 9 m; 6 f | 1 m; 3 f | 6 m; 5 f | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoch, D.; Bachbauer, M.; Pöchlauer, C.; Algaba-Chueca, F.; Tandl, V.; Novakovic, B.; Megia, A.; Gauster, M.; Saffery, R.; Glasner, A.; et al. Maternal Obesity Alters Placental Cell Cycle Regulators in the First Trimester of Human Pregnancy: New Insights for BRCA1. Int. J. Mol. Sci. 2020, 21, 468. https://doi.org/10.3390/ijms21020468
Hoch D, Bachbauer M, Pöchlauer C, Algaba-Chueca F, Tandl V, Novakovic B, Megia A, Gauster M, Saffery R, Glasner A, et al. Maternal Obesity Alters Placental Cell Cycle Regulators in the First Trimester of Human Pregnancy: New Insights for BRCA1. International Journal of Molecular Sciences. 2020; 21(2):468. https://doi.org/10.3390/ijms21020468
Chicago/Turabian StyleHoch, Denise, Martina Bachbauer, Caroline Pöchlauer, Francisco Algaba-Chueca, Veronika Tandl, Boris Novakovic, Ana Megia, Martin Gauster, Richard Saffery, Andreas Glasner, and et al. 2020. "Maternal Obesity Alters Placental Cell Cycle Regulators in the First Trimester of Human Pregnancy: New Insights for BRCA1" International Journal of Molecular Sciences 21, no. 2: 468. https://doi.org/10.3390/ijms21020468