1. Introduction
Pancreatic cancer remains a cancer of unmet need, with a 5 year survival rate of approximately 7−9% [
1]. The survival rate has shown little to no improvement over a 30 year period. Pancreatic ductal adenocarcinoma (PDAC) has no viable screening method, and an absence of early clinical symptoms, leaving it detected usually later in the disease stage. The only curative option is surgery, and this is only viable in early stage (R0/R1) cases where the cancer has not metastasised. Even with radical surgical resection, the 5 year survival rates are not comparable with other solid cancers.
One of the most common research tools for oncology research is the cell-line model. Cell lines and cell-line panels have formed the basis of much preclinical research previously. The limitations of conventional cell lines for pancreatic cancer research has been previously discussed, showing to be a poor predictor of clinical trial outcome [
2]. In pancreatic cancer, in vitro models are often poor in predicting clinical therapeutic response [
3].
Patient-derived xenograft (PDX) models have emerged as a tool for investigation of pancreatic cancer and become more utilised as they have been shown to more closely represent patient tumours [
4]. Due to its more aggressive nature, pancreatic cancer PDX models are established with a high take rate when compared to other cancers, such as breast cancer [
5,
6]. While ovarian cancer PDX models are established with similar engraftment rates as pancreatic cancer [
7].
Pancreatic cancer cell lines, while not mimicking the tumour–stromal interaction or pancreatic cancer’s dense desmoplasia offer a model to examine direct drug effects on the tumour cells. PDX models also offer the opportunity for the development of novel cell-line models. The establishment of pancreatic cancer cell lines using finely minced tissue fragments and specialised media components has been previously reported [
8,
9]. These additional cell-line models provide valuable resources to map the genomic alterations involved in human pancreatic cancer.
This study shows the comprehensive development of patient-derived xenograft models, including detailing the tumours that failed to proliferate as PDX models. From these novel models, primary cell lines were developed and characterised as human tumour cell lines. Using microarray screening this study sought to compare the mRNA profile of adjacent normal tissue, patient tumour tissues and PDX F1 tissue. This revealed the innate heterogeneity of the adjacent normal tissue and patient pancreatic tumour but showed patient-derived xenograft models more closely clustered together. More than 1400 genes were differential expressed between the tumour and adjacent normal tissue, with more than 3800 genes differentially expressed in the tumour–PDX comparison. In this study, we focused on genes that were differentially expressed in both comparisons and associated with disease initiation and disease progression characteristics.
3. Materials and Methods
3.1. Sample Acquisition and Ethical Approval
Pancreatic cancer tissue and adjacent normal tissue (N) were obtained from patients undergoing surgical resection at St Vincent’s University Hospital. After initial macroscopic pathological confirmation, material remaining after diagnostic sampling was cold transferred in RPMI 1640 medium containing 1% Penicillin-Streptomycin, 1% fungazone to DCU. Transfer time between hospital and implantation was on average 2 h or less.
Collection of patient material was approved by St Vincent’s University Hospital Research Ethics Committee. All animal work received ethical approval from the DCU Research Ethics Committee (DCUREC/2012/202) and was licensed by Department of Health (B100-4501).
3.2. PDX Tumour Development
The tumour was cut into implant-sized pieces (<2 mm3) and rinsed with fresh serum-free RPMI media following transport. Severe combined immunodeficiency (SCID), CB17/lcr-Prkdcscid/lcrCrl mice (Charles River, UK) were implanted subcutaneously with fresh patient tumour material. Depending upon the size and type of tumour material, 3–5 mice were implanted per patient sample. Under anaesthesia (isoflourane, O2 carrier gas) a small incision was made in the skin of the left flank of the animal. The tumour piece was placed in the pocket under the skin and the wound sealed with a single staple. The animals were monitored post-surgery, and staple removal was within 10 days. Animals were monitored weekly for body weight and tumour development. Mice were monitored for tumours development for up to 1 year post implantation. Animal welfare monitoring criteria included tumour volume, tumour axis, body weight and condition. There tumour volume and tumour axis limits were set as <2000 mm3, and <20 mm respectively. A decrease in body weight of >10% resulted in increased monitoring with body weight decrease of 20% resulting in humane euthanasia.
3.3. Preservation of PDX tumours
Following humane euthanasia of the mouse, the tumour was divided for cryopreservation, formalin-fixed paraffin embedding (FFPE) and snap frozen. For cryopreservation, implant-sized tumour pieces were stored in freezing media (RMPI/FCS/DMSO—50/40/10% v/v) in a cryovial and placed in a Mr FrostyTM freezing container (ThermoFischer, Dublin Ireland) for 24 h and then transferred to the liquid nitrogen (LN2) for long-term storage. For snap freezing, a piece of tumour was minced with a scalpel, placed in a cryovial and placed directly into LN2, then stored at −80 °C. For FFPE, a central slice was preserved in 10% neutral buffered formalin overnight and processed subsequently in 70%, 90%, 100% ethanol followed 100% xylene (1 h each). Following processing the tumours were embedded in paraffin. FFPE samples of PDX tumours were reviewed by consultant histopathologist to validate maintenance of tumour phenotype in comparison with original patient tumour.
3.4. Cell Isolation from Tumour Material
A small piece of tumour (<2 mm3) was minced and placed in Collagenase/Hyaluronidase (StemCell Technologies Cambridge, UK) at 4 °C for 1 h and then transferred to 37 °C for 30 min. Following digestion, the sample was centrifuged 200 g for 5 min and the supernatant removed. The digested material was suspended with complete media (DMEM/F-12 Hams supplemented with 10% FCS 1% PenStrep). Samples were monitored for growth and fed as required. Tumour cells were isolated from first generation PDX tumours while fibroblasts were isolated directly from patient tumours. Tumour cell lines were confirmed as human by IHC.
3.5. Immunohistochemical Analysis of PDX tumours to determine Human-Mouse Tissue
3.5.1. Immunohistochemistry
All immune-histochemical (IHC) staining was performed using the DAKO Autostainer. Deparaffinisation and antigen retrieval were performed using Epitope Retrieval 3-in-1 solution (pH 6) (DAKO, Cruinn, Dublin Ireland) was used. For epitope retrieval slides were heated to 97 °C for 20 min and then cooled to 65 °C.
3.5.2. Real Envision Detection System Peroxidase/DAB+
Following either deparaffinisation or epitope retrieval as outlined above slides were immersed in 1× wash buffer. On the autostainer (DAKO), slides were blocked for 10 min with 200 µL HRP Block (DAKO). Slides were washed with 1× wash buffer and 200 µL of the primary antibody were added to the slides for 30 min. The primary antibody used was Abcam Anti-Mitochondria antibody 113–1 (ab92824), a human-specific antibody [
21]. The antibody was used at a dilution of 1 in 1000. Slides were once again washed with 1× wash buffer and then incubated with 200 µL Real EnVision (DAKO) for 30 min. Slides were washed again with wash buffer and then stained with 200 µL DAB+ substrate for 5 min and this procedure was repeated twice. All slides were then counterstained with haematoxylin (DAKO) for 5 min and were rinsed with deionised water and then with wash buffer. A negative control sample was also tested for each sample using antibody diluent without any antibody present. This was used to evaluate any non-specific staining. Following the counterstaining with haematoxylin, the slides were then dehydrated. This was achieved by immersing the slide in 70%, 90% and 100% ethanol, twice in each ethanol solutions for 3 min. The slides were then immersed into xylene, twice, for 5 min each. Once the slides were cleared, they were mounted using DPX (BDH).
3.5.3. Immunohistochemical Analysis of Cell Lines
Cells were seeded directly on Superfrost
® microscope slides and allowed to attach overnight at 37 °C, 5% CO
2. Slides were washed 3 times in PBS and cells were then fixed in cold 4% paraformaldehyde for 15 min and washed with PBS. Immunostaining was carried out using the Dako Autostainer as per
Section 3.5.2, without the initial antigen retrieval.
3.6. Mutational Analysis
Mass spectrometry-based single-nucleotide polymorphism genotyping technology (Agena Biosciences, Hamburg, Germany) was used for identification of hotspot, potentially clinically relevant nonsynonymous somatic mutations as previously described [
22]. The genes were further subdivided by pathway, and include MAPK, PI3K and related pathway genes (
Supplementary Table S3). Assays were designed using strict assay design parameters optimized for sensitive mutation detection. The panel consisted of 31 multiplex assays capable of detecting 378 somatic hotspot mutations in 49 genes. DNA was extracted from fresh frozen tumour material using QIAmp DNA mini kit (Qiagen, Hilden, Germany) and quantified by Qubit dsDNA. 10 ng of DNA was added to each PCR reaction and DNA was amplified using custom designed PCR primer pools. Unincorporated nucleotides were inactivated using shrimp alkaline phosphatase (SAP), and a single base extension reaction was performed using extension primers that hybridise immediately adjacent to the mutations of interest. Salts were removed by adding a cation-exchange resin, before the multiplexed reactions were spotted onto SpectroCHIP II arrays. Matrix chips were analysed on an Agena MassArray Matrix-assisted laser desorption/ionisation Time of Flight (MALDI-TOF) system.
3.7. Sample Preparation for Microarray
Snap-frozen sections of PDX tumours, original patient material and adjacent normal tissue were prepared for RNA extraction by grinding samples under LN2 using a mortar and pestle. Briefly, tissue sample was placed in a metal mortar with a small volume of LN2 and ground quickly. When powdered the powder was transferred to a clean Eppendorf tube. This was done before the LN2 completely evaporated and while the powder was still frozen.
Once homogenised, RNA samples were prepared using Trizol, according to manufacturer’s guidelines. Briefly, powdered tissue was lysed in 1 mL of Trizol reagent. Sample was allowed to homogenise at room temperature for 15 min. Per 1 mL of Trizol reagent used, 200 µL of chloroform was added to the Eppendorf (Hamburg, Germany), sample was mixed and allowed to sit for 5 min. At 4 °C samples were spun at 12,000× g for 15 min. The aqueous phase was then separated using a micropipette and places in a new Eppendorf. 500 µL of isopropanol was added to the aqueous phase, allowed to sit at room temperature for 10 min, after which it was spun at 12,000× g at 4 °C for 10 min. The RNA pellet was then washed using 75% ethanol and spun at 4 °C at 7500× g for 5 min. Ethanol was removed from pellet, which were allowed to air dry, and were then reconstituted in nuclease free water.
Samples were quantified using a Nanodrop (Thermo Scientific, Dublin Ireland) and quality determined using Agilent Bioanalyser. Samples with a RNA Integrity Number (RIN) number greater than 8 were most suitable for microarray analysis.
3.8. Microarray GeneChip Human Gene 2.0 ST array Processing and Hybridization
Preparation of cRNA, hybridization, and scanning of microarrays was performed according to the manufacturer’s protocol (Affymetrix, Thermo-Fischer, Santa Clara, Ca, USA). In brief, 300 ng of total RNA extracted from homogenised patient samples isolated using Trizol was converted into double-stranded cDNA by reverse transcription. Biotin-labelled cRNA was generated by converting the cDNA sample using the Genechip WT plus reagent kit (Affymetrix, Thermo-Fischer, Santa Clara, Ca, USA). Labelled cRNA was hybridized to the Affymetrix GeneChip® Human Gene 2.0 ST Array while rotating at 60 rpm for 16 h at 45 °C. After hybridization, the microarray was washed using the Affymetrix Fluidics Station according to the manufacturer’s protocol. The chips were scanned in an Affymetrix 3000 7G scanner.
Differential gene analysis expression was carried out using Applied Biosystems Transcriptome Analysis Console (TAC) software 4.0.2. Resulting gene lists were filtered for +/- 2-fold changes, a p-value < 0.05 and an overall FDR F-Test: <0.005.
4. Discussion
Patients ages and diagnosis characteristics are broadly representative of the earlier-stage pancreatic cancer patient population at large. In Ireland, the median age of diagnosis is 70–74 years for the entire pancreatic patient population [
23]. In this study, the average age of 65 years for women and 67 years for men shows that patients who are eligible for surgery are somewhat younger than the pancreatic patient population as a whole. Only 10–20% of patients diagnosed with pancreatic cancer are eligible for surgery, and the biobank developed here is representative of that patient population [
24,
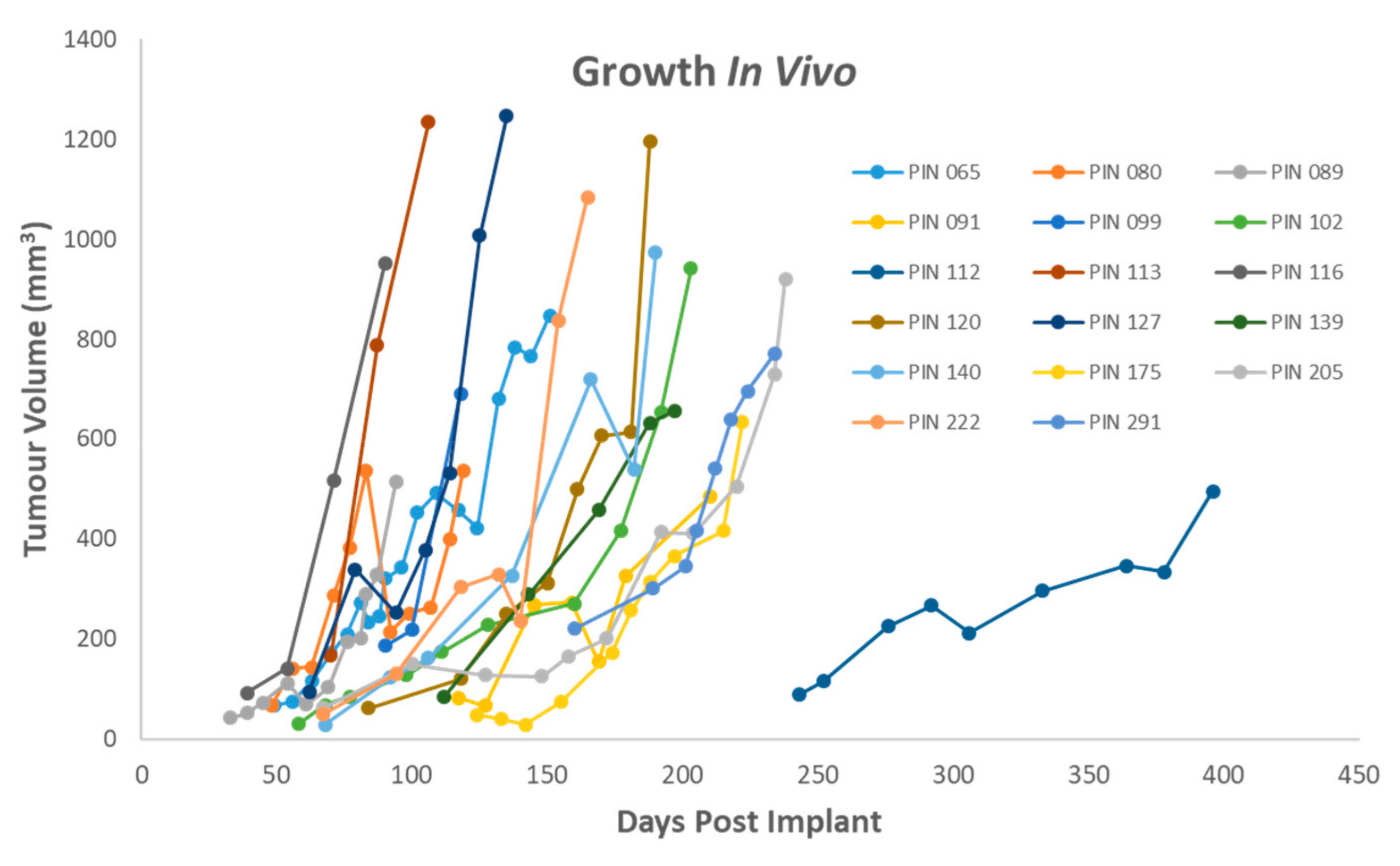
25]. The ability of a patient tumour to grow a tumour in vivo was not correlated with any single characteristic, though PDAC samples, which formed the bulk of the samples received, formed tumours at a greater rate than neuroendocrine tumours (78% compared to 20%).
PDAC samples which were treatment naïve formed tumours in vivo 78% of the time. This is somewhat higher that previously reported rates of 61% (sub-cutaneous) and 62% (orthotopic) for primary pancreatic cancer samples, though these models were established in nude mice rather than SCID mice [
26,
27]. The 78% reported here is also higher than 43% detailed by Pergolini et al., though Pergolini et al. reported cryopreserving tumours first rather than direct implantation which potential reflects the differences seen here [
28]. This suggests that for initial passaging, direct implantation gives a better take rate with cryopreservation being suitable for later passages. This is potentially due to the selection pressures the tumour undergoes in vivo and the biological processes involved in engraftment.
From seven tumours collected where the patient had received neo-adjuvant treatment, one formed a sub-cutaneous tumour in vivo. Dorado et al. examined three experimental techniques for the establishment of pancreatic PDX models, subcutaneous, orthotopic and intraperitoneal. Of the bank of 11 samples collected in that study, one had received neo-adjuvant therapy and formed a sub-cutaneous tumour, but did not form a tumour intraperitoneal nor at the orthotopic site [
29].
Previously, our group examined 10 primary patient tumour samples and nine matched normal adjacent specimens and 10 matched F1 and F2 generation PDX tumours using high-resolution mass spectrometry. MS identification allowed for the isolation of the human only protein, representing the tumour cells, and these were tracked from patient tumour, to F1 and F2 generation PDX tumours. Between patient primary and F1 tumour 32 proteins were upregulated and 113 downregulated. In comparison, between F1 and F2 generation of PDX tumour, only eight human-specific proteins were differentially expressed when analysed by quantitative label-free differential analysis. This demonstrates the fidelity of tumour phenotype once engrafted [
30]. This is in line with previously published data the shows, once established, xenografts tend to be robust with stable gene expression profile between early and late passage PDX tumours [
31]. This bank of pancreatic PDX tumours models the subset of pancreatic cancer patients who are eligible for surgical resection. Mutational analyses by Sequenom MassArray MALDI-TOF showed 80% of samples examined possessed a KRAS mutation. This is in line with previously reported clinical KRAS mutation rates [
32]. These PDX samples are a truer representation of human pancreatic cancer than pancreatic cancer cell lines or cell-line derived xenograft models, which often fail to recapitulate the stroma and desmoplasia of pancreatic cancer. In recent years, given the unmet need of pancreatic cancer patients, banks of patient-derived xenograft tumours have been established. The bank established in this study displays a higher engraftment rate for PDAC tumours than many others previously reported. Two recent reports showed similar engraftment rates (72% and 71%), although this was observed in more immune compromised NSG and NSG nude mice [
33,
34].
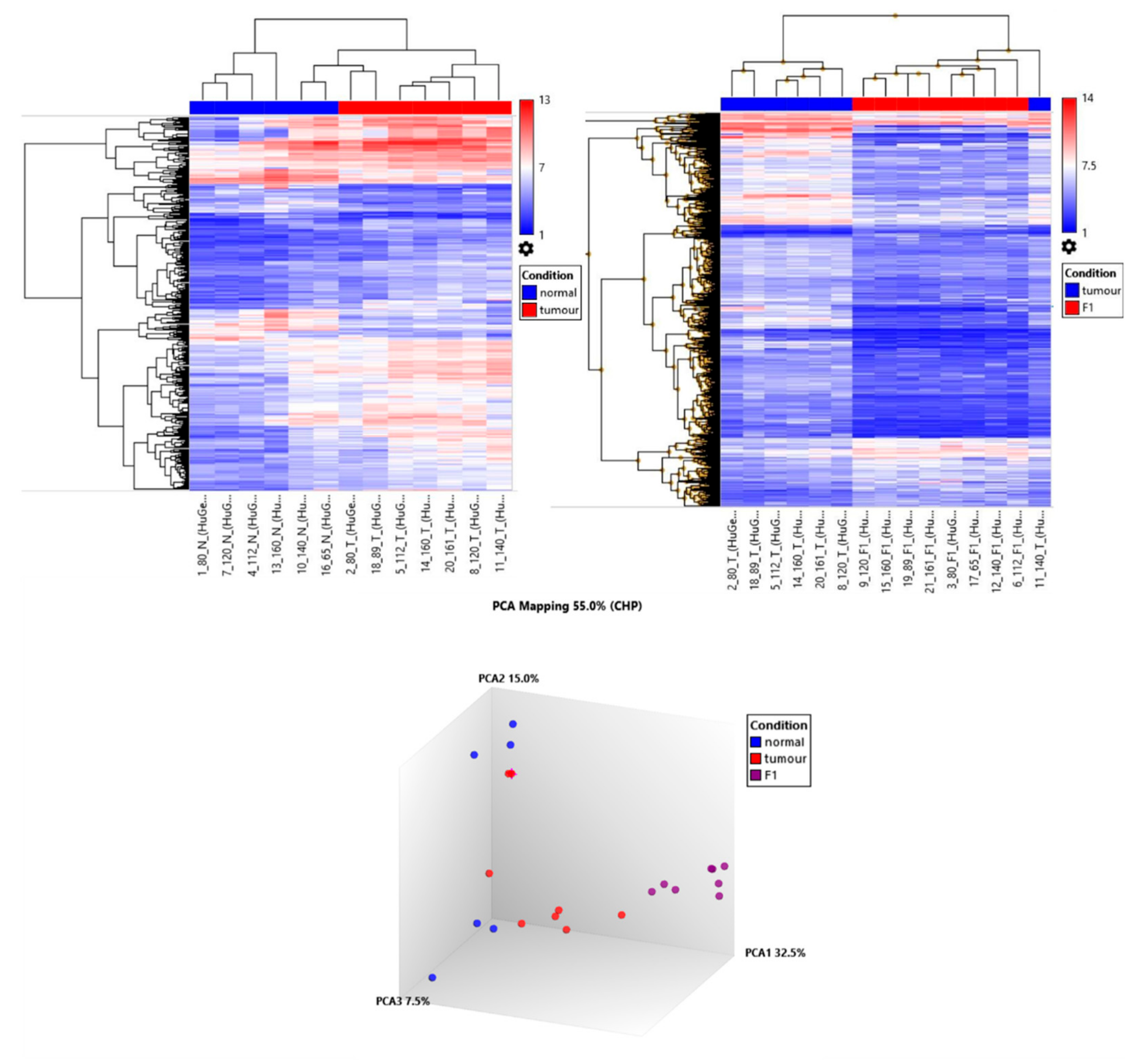
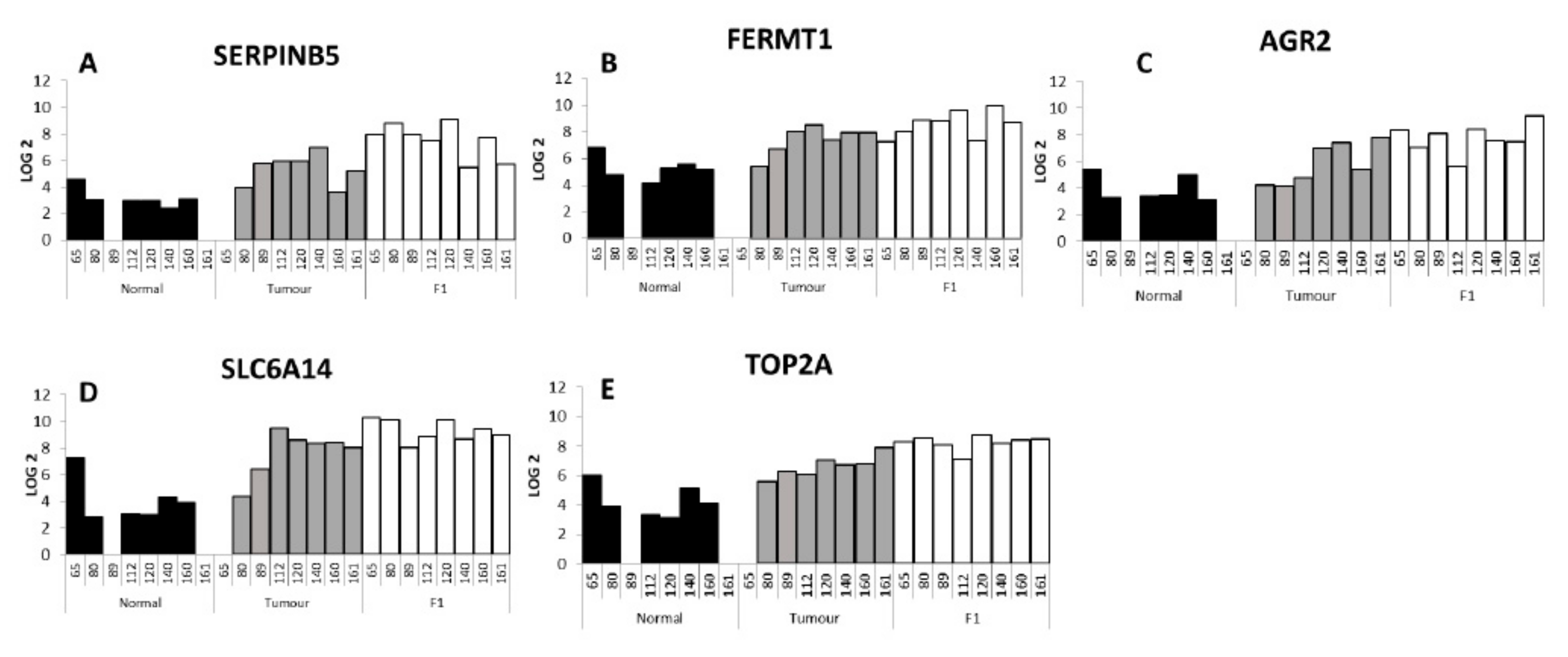
Eight PDX tumour samples were interrogated by Affymetrix Microarray technology in matched comparison to patient tumour and adjacent normal tissue. Primary patient tumour cluster together with PDX tumours clustering together but separate from the primary tumour. RNASeq data in ovarian PDX models demonstrate this differential clustering is predominantly due to the loss of human stroma and the growth of murine stroma [
35]. By comparing the adjacent normal to the tumour and the tumour to the F1 PDX tumour we aimed to map the genes associated with tumour growth, proliferation and tumour in engraftment. Gene ontology analysis of the 89 gene increased in tumour (vs. adjacent normal) and further increased in PDX tumour (vs. patient tumour) showed the biological processed enriched for genes associated with cell cycle, cell cycle process as well as mitotic cell cycle processes (
Table 8 and
Table 9). A selection of the genes shortlisted and described here, have previously been associated with pancreatic cancer and validated as having an effect on proliferation, invasion and migration.
Serpin family B member 5,
SERPINB5, had the largest fold change in the T vs. F1 analysis and in the top ten differentially expressed genes in the T vs. N analysis. The SERPIN superfamily of proteins is known to consist of 37 members in humans, with 13 members of the SERPINA family and 13 members of the SERPINB family [
36]. SERPINA3 has been associated with endometrial cancers [
37] and undifferentiated carcinoma with osteoclast-like giant cells, a rare pancreatic cancer [
38]. In our differential gene expression list 3 of the 39 SERPIN family members were statistically significantly changed,
SERPINA1, SERPINB5 and
SERPINE1.
SERPINA1 was increased in T vs. N (3.6-fold), but decreased in T vs. F1 (5.0-fold).
SERPINE1 was decreased in T vs. N analysis (3.6-fold) and further decreased in T vs. F1 comparison (4.9-fold).
SERPINB5 has been previously associated with pancreatic cancer. In a meta-analysis of publicly available datasets of transcriptome data for human pancreas specimens and a GEM mouse models Bhasin et al. included
SERPINB5 as one the of the genes in a five gene panel for discriminating PDAC and early precursor lesions. Other genes included in this panel were
TMPRSS4, AHNAK2, POSTN, ECT2 [
39]. The other four genes in the panel were also detected as being significantly changed in our analysis.
TMPRSS4 showed fold changes of 9.5 and 2.1 in the N vs. T and T vs. F1 respectively. Similarly,
AHNAK2 fold changes were 2.6 (N vs. T) and 2 (T vs. F1),
POSTN 15.5 (N vs. T) and -69.9 (T vs. F1),
ECT2 2.6 (N vs. T) and 2.7 (T vs. F1). RNA-Seq analysis by Mao et al. of 10 PDAC tumour samples and adjacent benign tissue also detected
SERPINB5. Their analysis showed
SERPINB5 to be overexpressed in PDAC tumour in comparison to adjacent normal tissue [
40]. Also in 2010, Mardin et al. identified through the analysis of the invasive and metastatic potential of 16 PDAC cell lines that
SERPINB5 expression may correspond with invasive tumours [
41]. Previous proteomic analysis of the same sample cohort also found
SERPINB5 to be increased at the protein level when human-specific proteins were identified with significantly increased expression in PDX F1 tumours compared to PDAC tumour tissues, as determined by quantitative label-free mass spectrometric analysis [
30].
Fermitin family member 1,
FERMT1, encoded protein is involved in integrin signalling and linkage of the actin cytoskeleton to the extracellular matrix and has been shown to be significantly over-expressed in colon cancer [
42]. FERMT1 has been shown to interact directly with β-catenin and activated the Wnt/β-catenin signalling pathway by decreasing the phosphorylation level of β-catenin in colon cancer. This activation was seen to promote EMT and led to a more aggressive and invasive phenotype in colon cancer [
43]. In oesophageal cancer, overexpression of
FERMT1 by lenti-viral vector increased proliferation and radiation resistance in vitro [
44]. A study by Fukuhisa et al. examined the role of EPS8 in PDAC, and showed that EPS8 was overexpressed in PDAC clinical specimens, and linked with proliferation and invasion in vitro.
FERMT1 is a putative downstream gene target of EPS8 [
45]. While changes in
EPS8 were not seen in our data set,
FERMT1 changes were seen. To date,
FERMT1 changes have not previously been directly shown in pancreatic cancer.
Ramachandran et al. showed that Anterior Gradient 2,
AGR2, gene was expressed in pancreatic cancer tissues and cell lines, though this is not expressed in chronic pancreatitis samples. In vitro,
AGR2 silencing by siRNA knockdown showed a decrease in cancer cell proliferation and invasion and increased gemcitabine sensitivity of resistant cells [
46]. In breast cancer patients,
AGR2 was associated with poorer survival and increased metastasis [
47]. In pancreatic cancer, the invasive potential of pancreatic cancer cells in vitro was proportional to the
AGR2 expression level [
48].
Solute carrier family 6 member, SLC6A14 encoded protein is a sodium- and chloride-dependent neutral and basic amino acid transporter [
49]. In breast cancer mouse models,
SLC6A14 knock out mice showed a marked delay in tumour formation and tumour size, and analysis of the tumour material revealed evidence of amino acid starvation in the slc6a14
-/- mice suggesting a key role from SLC6A14 in both tumour development and growth [
50]. In 2019, Cheng et al. used three publicly available datasets to identified differentially regulated genes and validated these genes in a retrospective clinical study. This highlighted
SLC6A14 as a potential prognostic biomarker of pancreatic cancer. Interestingly
CENPF and
SERBINB5 were also included in this panel of prognostic genes. Both of these genes were also identified in our panel [
51]. SLC6A14 has been investigated as a potential novel therapeutic target for pancreatic cancer using α-methyltryptophan as a pharmacological inhibitor of SLC6A14 and showed reduced proliferation and clonogenic survival [
52].
Topoisomerase IIα (
TOP2A) gene encodes for a nuclear enzyme that regulates the topological structure of DNA and is involved in chromosome condensation, chromatid separation and torsional stress relief during DNA transcription and replication. TOP2A has been associated with many solid cancers such as breast and prostate cancers [
53,
54,
55]. However, in a study of 24,262 patients with diverse tumour types, only 4% of tumours had
TOP2A amplification [
56]. Pancreatic patients with higher TOP2A levels were thought to have poor prognosis. TOP2A has been shown in pancreatic cancer cell lines to be associated with proliferation and migration through activation of the β-catenin signalling pathway [
57].

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}