Ileal Transcriptome Profiles of Japanese Quail Divergent in Phosphorus Utilization

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Results and Discussion
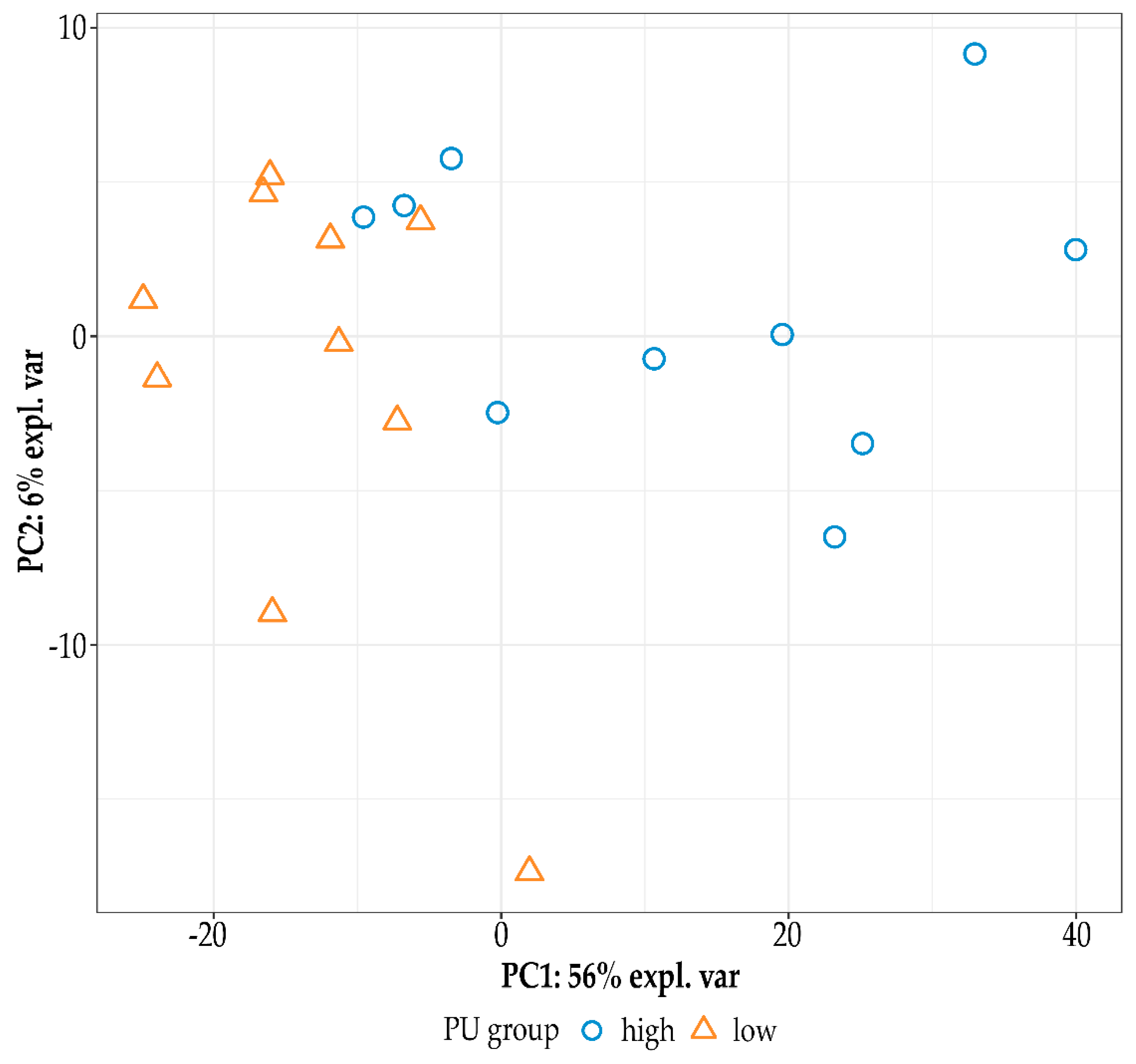
2.1. Sequencing Data Analysis and Differential Gene Expression
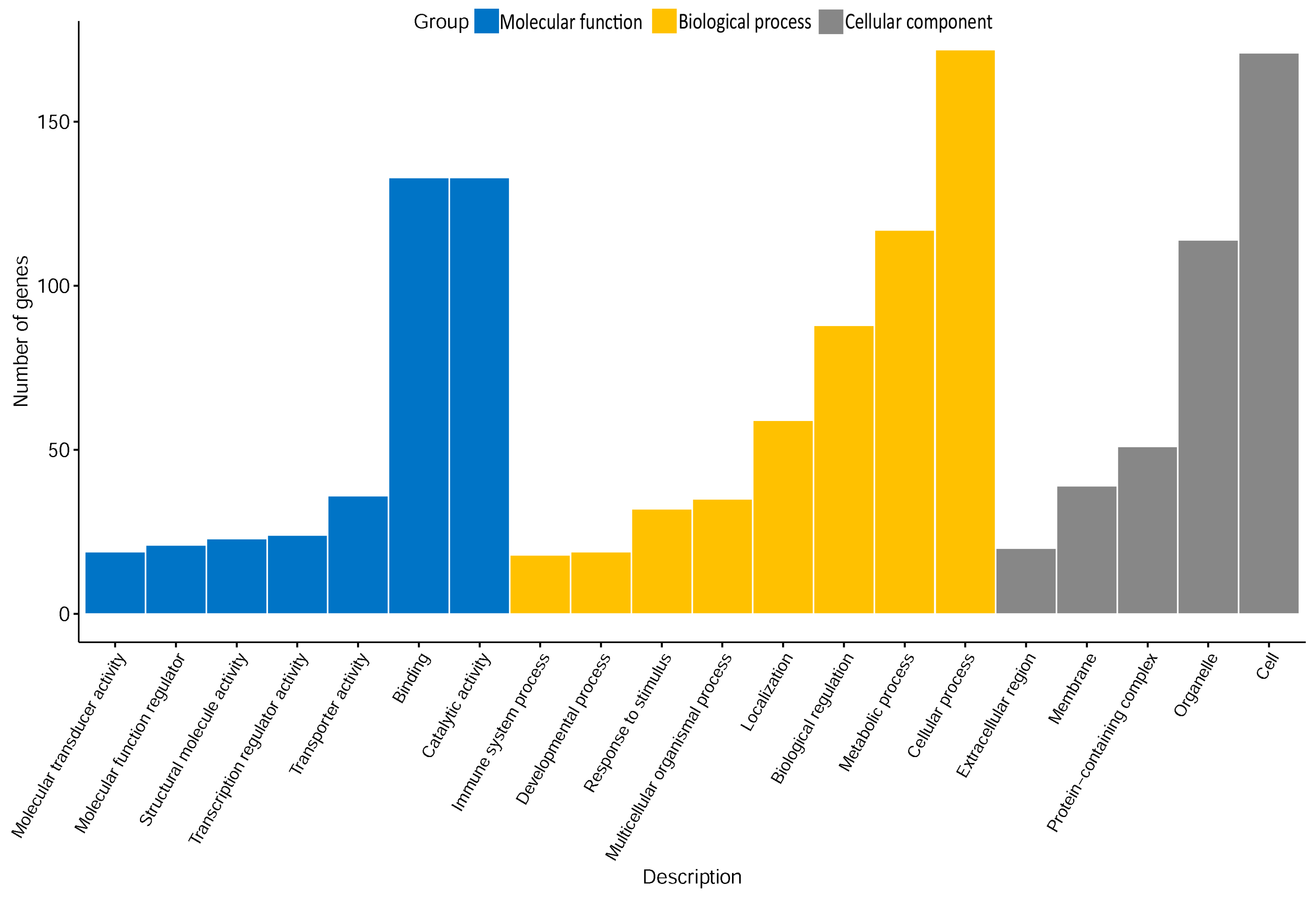
2.2. Gene Ontology (GO) Analysis
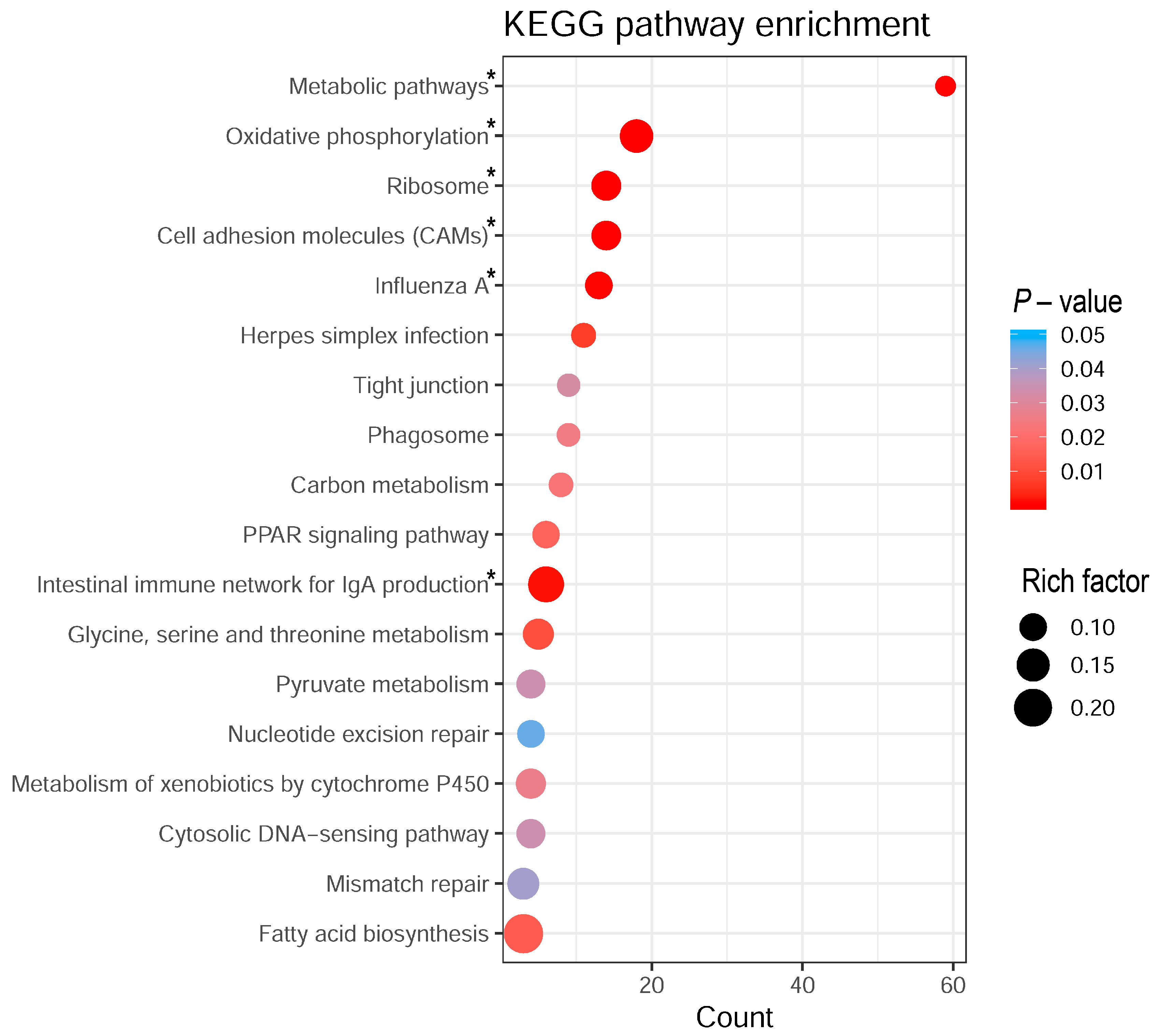
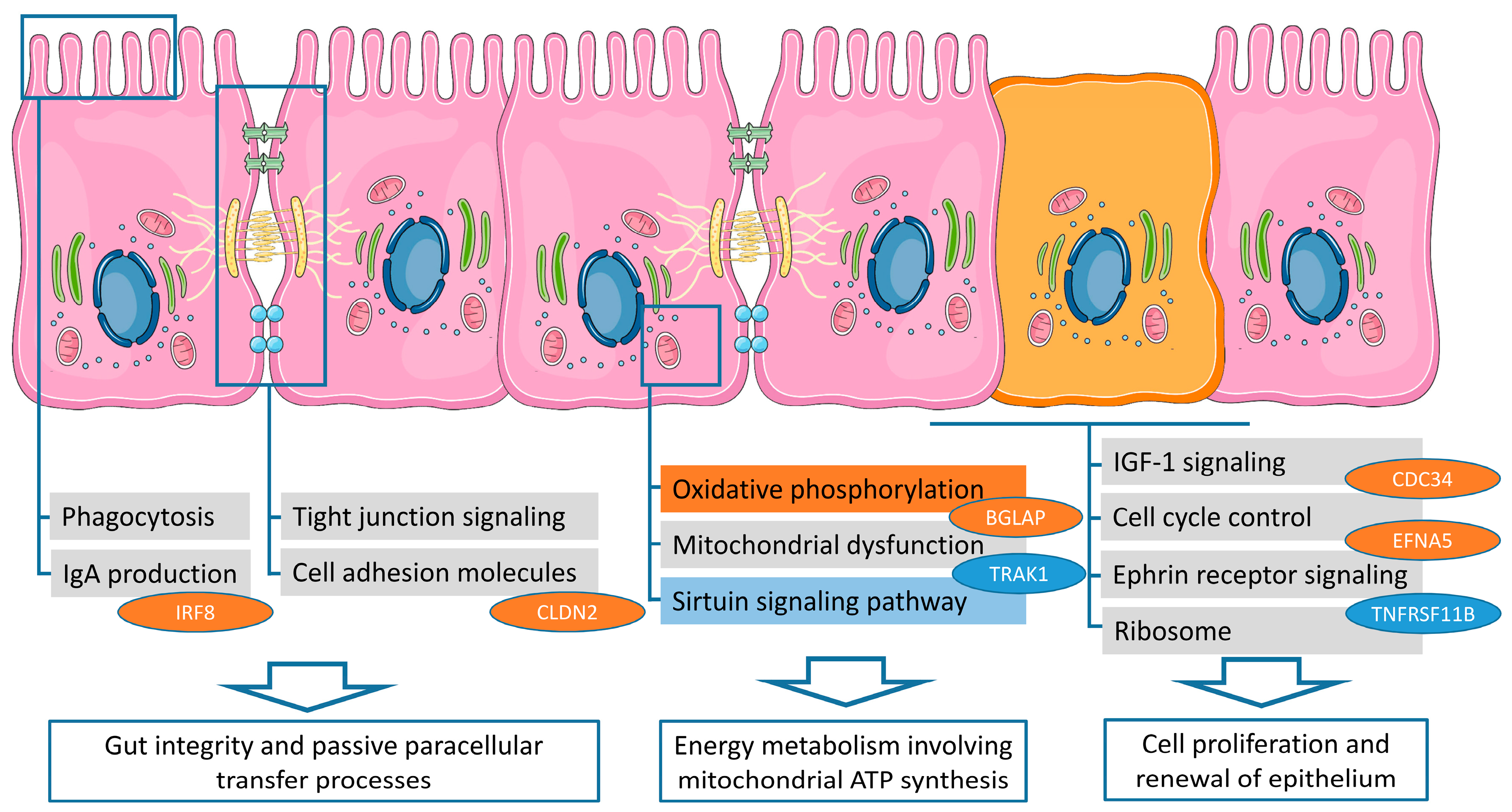
2.3. Biological Pathway Analysis
2.4. Target Genes Attributed to the Vitamin D System
3. Materials and Methods
3.1. Animals and Sample Collection
3.2. RNA Preparation, Library Construction, and RNA Sequencing
3.3. Data Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CLDN2 | Claudin 2 |
| FI | Feed intake |
| GO | Gene ontology |
| InsP6 | Inositol hexakisphosphate |
| IPA | Ingenuity pathway analyses |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| OPG | Osteoprotegerin |
| P | Phosphorus |
| PU | Phosphorus utilization |
| RANK | Receptor Activator of NF-κB |
| RANKL | Receptor Activator of NF-κB Ligand |
| VDR | Vitamin D receptor |
References
- Berndt, T.; Kumar, R. Novel mechanisms in the regulation of phosphorus homeostasis. Physiology 2009, 24, 17–25. [Google Scholar] [CrossRef]
- Just, F.; Oster, M.; Büsing, K.; Borgelt, L.; Muráni, E.; Ponsuksili, S.; Wolf, P.; Wimmers, K. Lowered dietary phosphorus affects intestinal and renal gene expression to maintain mineral homeostasis with immunomodulatory implications in weaned piglets. BMC Genom. 2018, 19, 207. [Google Scholar] [CrossRef] [Green Version]
- Gerlinger, C.; Oster, M.; Borgelt, L.; Reyer, H.; Muráni, E.; Ponsuksili, S.; Polley, C.; Vollmar, B.; Reichel, M.; Wolf, P.; et al. Physiological and Transcriptional Responses in Weaned Piglets Fed Diets with Varying Phosphorus and Calcium Levels. Nutrients 2019, 11, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Künzel, S.; Bennewitz, J.; Rodehutscord, M. Genetic parameters for bone ash and phosphorus utilization in an F2 cross of Japanese quail. Poult. Sci. 2019, 98, 4369–4372. [Google Scholar] [CrossRef] [PubMed]
- Beck, P.; Piepho, H.P.; Rodehutscord, M.; Bennewitz, J. Inferring relationships between Phosphorus utilization, feed per gain, and bodyweight gain in an F2 cross of Japanese quail using recursive models. Poult. Sci. 2016, 95, 764–773. [Google Scholar] [CrossRef] [PubMed]
- Minvielle, F. What are quail good for in a chicken-focused world? World’s Poult. Sci. J. 2009, 65, 601–608. [Google Scholar] [CrossRef]
- Rodehutscord, M.; Dieckmann, A. Comparative studies with three-week-old chickens, turkeys, ducks, and quails on the response in phosphorus utilization to a supplementation of monobasic calcium phosphate. Poult. Sci. 2005, 84, 1252–1260. [Google Scholar] [CrossRef]
- Reyer, H.; Oster, M.; Wittenburg, D.; Muráni, E.; Ponsuksili, S.; Wimmers, K. Genetic Contribution to Variations of Blood Calcium, Phosphorus and Alkaline Phosphatase Activity in Pigs. Front. Genet. 2019, 10, 590. [Google Scholar] [CrossRef]
- Eeckhout, W.; De Paepe, M. Total phosphorus, phytate-phosphorus and phytase activity in plant feedstuffs. Anim. Feed Sci. Technol. 1994, 47, 19–29. [Google Scholar] [CrossRef]
- Zeller, E.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Hydrolysis of phytate and formation of inositol phosphate isomers without or with supplemented phytases in different segments of the digestive tract of broilers. J. Nutr. Sci. 2015, 4, e1. [Google Scholar] [CrossRef] [Green Version]
- Palacios, M.C.; Haros, M.; Sanz, Y.; Rosell, C.M. Selection of lactic acid bacteria with high phytate degrading activity for application in whole wheat breadmaking. LWT-Food Sci. Technol. 2008, 41, 82–92. [Google Scholar] [CrossRef]
- Sommerfeld, V.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Interactive effects of phosphorus, calcium, and phytase supplements on products of phytate degradation in the digestive tract of broiler chickens. Poult. Sci. 2018, 97, 1177–1188. [Google Scholar] [CrossRef] [PubMed]
- Sommerfeld, V.; van Kessel, A.G.; Classen, H.L.; Schollenberger, M.; Kühn, I.; Rodehutscord, M. Phytate degradation in gnotobiotic broiler chickens and effects of dietary supplements of phosphorus, calcium, and phytase. Poult. Sci. 2019, 98, 5562–5570. [Google Scholar] [CrossRef] [PubMed]
- Pontoppidan, K.; Glitsoe, V.; Guggenbuhl, P.; Quintana, A.P.; Nunes, C.S.; Pettersson, D.; Sandberg, A.S. In vitro and in vivo degradation of myo-inositol hexakisphosphate by a phytase from Citrobacter braakii. Arch. Anim. Nutr. 2012, 66, 431–444. [Google Scholar] [CrossRef] [PubMed]
- Maenz, D.D.; Classen, H.L. Phytase activity in the small intestinal brush border membrane of the chicken. Poult. Sci. 1998, 77, 557–563. [Google Scholar] [CrossRef]
- Huber, K.; Zeller, E.; Rodehutscord, M. Modulation of small intestinal phosphate transporter by dietary supplements of mineral phosphorus and phytase in broilers. Poult. Sci. 2015, 94, 1009–1017. [Google Scholar] [CrossRef]
- Wubuli, A.; Reyer, H.; Muráni, E.; Ponsuksili, S.; Wolf, P.; Oster, M.; Wimmers, K. Tissue-Wide Gene Expression Analysis of Sodium/Phosphate Co-Transporters in Pigs. Int. J. Mol. Sci. 2019, 20, 5576. [Google Scholar] [CrossRef] [Green Version]
- Huber, K.; Hempel, R.; Rodehutscord, M. Adaptation of epithelial sodium-dependent phosphate transport in jejunum and kidney of hens to variations in dietary phosphorus intake. Poult. Sci. 2006, 85, 1980–1986. [Google Scholar] [CrossRef]
- Lim, H.S.; Namkung, H.; Paik, I.K. Effects of phytase supplementation on the performance, egg quality, and phosphorous excretion of laying hens fed different levels of dietary calcium and nonphytate phosphorous. Poult. Sci. 2003, 82, 92–99. [Google Scholar] [CrossRef]
- Angel, R.; Saylor, W.W.; Mitchell, A.D.; Powers, W.; Applegate, T.J. Effect of dietary phosphorus, phytase, and 25-hydroxycholecalciferol on broiler chicken bone mineralization, litter phosphorus, and processing yields. Poult. Sci. 2006, 85, 1200–1211. [Google Scholar] [CrossRef]
- Pike, J.W.; Meyer, M.B.; Martowicz, M.L.; Bishop, K.A.; Lee, S.M.; Nerenz, R.D. Emerging regulatory paradigms for control of gene expression by 1,25-dihydroxyvitamin D3. J. Steroid. Biochem. Mol. Biol. 2010, 121, 130–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oster, M.; Just, F.; Büsing, K.; Wolf, P.; Polley, C.; Vollmar, B.; Murani, E.; Ponsuksili, S.; Wimmers, K. Toward improved phosphorus efficiency in monogastrics—Interplay of serum, minerals, bone, and immune system after divergent dietary phosphorus supply in swine. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R917–R925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, X.D.; Suo, H.Q.; Lu, L.; Hu, Y.X.; Zhang, L.Y.; Luo, X.G. Effects of sodium, 1, 25-dihydroxyvitamin D3 and parathyroid hormone fragment on inorganic P absorption and Type IIb sodium-phosphate cotransporter expression in ligated duodenal loops of broilers. Poult. Sci. 2017, 96, 2344–2350. [Google Scholar] [CrossRef] [PubMed]
- Nemere, I.; Larsson, D. Does PTH have a direct effect on intestine? J. Cell. Biochem. 2002, 86, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Gentili, C.; Morelli, S.; Boland, R.; de Boland, A.R. Parathyroid hormone activation of map kinase in rat duodenal cells is mediated by 3′, 5′-cyclic AMP and Ca2+. Biochim. Biophys. Acta. Mol. Cell Res. 2001, 1540, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Miao, Z.; Zhang, G.; Zhang, J.; Li, J.; Yang, Y. Effect of early dietary energy restriction and phosphorus level on subsequent growth performance, intestinal phosphate transport, and AMPK activity in young broilers. PLoS ONE 2017, 12, e0186828. [Google Scholar] [CrossRef] [Green Version]
- Rodehutscord, M. Interactions between minerals and phytate degradation in poultry–challenges for phosphorus digestibility assays. In Phytate Destruction—Consequences for Precision Animal Nutrition; Walk, C.L., Kühn, I., Stein, H.H., Kidd, M.T., Rodehutscord, M., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 167–178. [Google Scholar] [CrossRef]
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [Green Version]
- Rath, E.; Moschetta, A.; Haller, D. Mitochondrial function—Gatekeeper of intestinal epithelial cell homeostasis. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 497–516. [Google Scholar] [CrossRef]
- Croom, W.J.; Brake, J.; Coles, B.A.; Havenstein, G.B.; Christensen, V.L.; McBride, B.W.; Peebles, E.D.; Taylor, I.L. Is intestinal absorption capacity rate-limiting for performance in poultry? J. Appl. Poult. Res. 1999, 8, 242–252. [Google Scholar] [CrossRef]
- Zhou, W.; Ramachandran, D.; Mansouri, A.; Dailey, M.J. Glucose stimulates intestinal epithelial crypt proliferation by modulating cellular energy metabolism. J. Cell Physiol. 2018, 233, 3465–3475. [Google Scholar] [CrossRef]
- Kita, K.; Nagao, K.; Okumura, J. Nutritional and tissue specificity of IGF-I and IGFBP-2 gene expression in growing chickens-a review. Asian-Australas J. Anim. Sci. 2005, 18, 747–754. [Google Scholar] [CrossRef]
- Lorenzo-Zuñiga, V.; Rodriguez-Ortigosa, C.M.; Bartoli, R.; Martinez-Chantar, M.L.; Martinez-Peralta, L.; Pardo, A.; Ojanguren, I.; Quiroga, J.; Planas, R.; Prieto, J. Insulin-like growth factor I improves intestinal barrier function in cirrhotic rats. Gut 2006, 55, 1306–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knöpfel, T.; Himmerkus, N.; Günzel, D.; Bleich, M.; Hernando, N.; Wagner, C.A. Paracellular transport of phosphate along the intestine. Am. J. Physiol. Gastrointest Liver Physiol. 2019, 317, G233–G241. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.; Yang, H.; Kang, H.S.; Ahn, C.; Hong, E.J.; An, B.S.; Jeung, E.B. Alteration of tight junction gene expression by calcium-and vitamin D-deficient diet in the duodenum of calbindin-null mice. Int. J. Mol. Sci. 2013, 14, 22997–23010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, F.; Thom, U.; Albrecht, E.; Weikard, R.; Nolte, W.; Kuhla, B.; Kuehn, C. Heat stress directly impairs gut integrity and recruits distinct immune cell populations into the bovine intestine. Proc. Natl. Acad. Sci. USA 2019, 116, 10333–10338. [Google Scholar] [CrossRef] [Green Version]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2015, 120, 692–701. [Google Scholar] [CrossRef]
- Kong, J.; Zhang, Z.; Musch, M.W.; Ning, G.; Sun, J.; Hart, J.; Bissonnette, M.; Li, Y.C. Novel role of the vitamin D receptor in maintaining the integrity of the intestinal mucosal barrier. Am. J. Physiol. Gastrointest Liver Physiol. 2008, 294, G208–G216. [Google Scholar] [CrossRef] [Green Version]
- Fujita, H.; Sugimoto, K.; Inatomi, S.; Maeda, T.; Osanai, M.; Uchiyama, Y.; Yamamoto, Y.; Wada, T.; Kojima, T.; Yokozaki, H.; et al. Tight Junction Proteins Claudin-2 and-12 Are Critical for Vitamin D-dependent Ca2+ Absorption between Enterocytes. Mol. Biol. Cell 2008, 19, 1912–1921. [Google Scholar] [CrossRef] [Green Version]
- Wikipathways. Available online: https://www.wikipathways.org/index.php/Pathway:WP2877 (accessed on 15 January 2020).
- Vidal, K.; Serrant, P.; Schlosser, B.; van den Broek, P.; Lorget, F.; Donnet-Hughes, A. Osteoprotegerin production by human intestinal epithelial cells: A potential regulator of mucosal immune responses. Am. J. Physiol. Gastrointest Liver Physiol. 2004, 287, G836–G844. [Google Scholar] [CrossRef] [Green Version]
- Chikatsu, N.; Takeuchi, Y.; Fukumoto, S.; Yano, K.; Fijita, N.; Tsuruo, T.; Fujita, T. Clonal endothelial cells produce humoral factors that inhibit osteoclast-like cell formation in vitro. Endocr. J. 2002, 49, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Baud’huin, M.; Duplomb, L.; Teletchea, S.; Lamoureux, F.; Ruiz-Velasco, C.; Maillasson, M.; Redini, F.; Heymann, M.F.; Heymann, D. Osteoprotegerin: Multiple partners for multiple functions. Cytokine Growth Factor Rev. 2013, 24, 401–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of bone resorption and formation by RANKL reverse signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Sims, N.A. RANKL/OPG; Critical role in bone physiology. Rev. Endocr. Metab. Disord. 2015, 16, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Lin, S.; Ranilla, M.J.; Baldwin, R.L. Transcriptomic profiling of duodenal epithelium reveals temporally dynamic impacts of direct duodenal starch-infusion during dry period of dairy cattle. Front. Vet. Sci. 2019, 6, 214. [Google Scholar] [CrossRef] [PubMed]
- Mills, A.D.; Faure, J.M. Divergent selection for duration of tonic immobility and social reinstatement behavior in Japanese quail (Coturnix coturnix japonica) chicks. J. Comp. Psychol. 1991, 105, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Rodehutscord, M. WPSA: Working Group No 2: Nutrition of the European Federation of Branches of WPSA. Determination of phosphorus availability in poultry. World’s Poult. Sci. J. 2013, 69, 687–698. [Google Scholar]
- VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs-und Forschungsanstalten). Die chemische Untersuchung von Futtermitteln. Handbuch der Landwirtschaftlichen Versuchs-und Untersuchungsmethodik Bd III; VDLUFA-Verlag: Darmstadt, Germany, 2006. [Google Scholar]
- Shastak, Y.; Rodehutscord, M. Determination and estimation of phosphorus availability in growing poultry and their historical development. World’s Poultry Sci. J. 2013, 69, 569–586. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storey, J.D.; Bass, A.J.; Dabney, A.; Robinson, D.; Qvalue: Q-Value Estimation for False Discovery Rate Control. R Package Version 2.18.0. Available online: http://github.com/jdstorey/qvalue (accessed on 14 January 2020).
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wie, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. 2), W316–W322. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canonical Pathways | P-Value 1 | z-Score 2 | Molecules |
|---|---|---|---|
| Mitochondrial Dysfunction * | <0.001 | ATP5F1D, ATP5MG, CASP9, COX5B, COX6A1, COX7B, MAOB, MT-ND6, NDUFA11, NDUFA13, NDUFB1, NDUFB6, NDUFB9, NDUFV1, SDHC, TRAK1, UQCR11 | |
| Oxidative Phosphorylation * | <0.001 | 3.6 | ATP5F1D, ATP5MG, COX5B, COX6A1, COX7B, NDUFA1, NDUFA13, NDUFB1, NDUFB6, NDUFB9, NDUFV1, SDHC, UQCR11 |
| Sirtuin Signaling Pathway * | <0.001 | −2.3 | ATP5F1D, GABARAPL1, GLS, HIST1H1T, MT-ND6, NDUFA11, NDUFA13, NDUFB1, NDUFB6, NDUFB9, NDUFV1, NFKB2, OGG1, PCK1, SDHC, TOMM6, TP53BP1, XPC |
| Fcγ Receptor-mediated Phagocytosis in Macrophages and Monocytes | 0.003 | 1.4 | ARPC3, ARPC5L, FGR, HMOX1, NCF1, NCK2, PIK3CG, RPS6KB2 |
| Cell Cycle Control of Chromosomal Replication | 0.003 | CDK10, CDK13, CDK4, POLE, RPA3, TOP2A | |
| Tight Junction Signaling | 0.003 | AFDN, CDK4, CLDN2, CLDN23, CLDN4, MYH6, NFKB2, PATJ, PPM1J, TJP1, TNFRSF11B | |
| IGF-1 Signaling | 0.005 | −1.3 | CASP9, IGF1R, IGFBP2, IGFBP7, PIK3CG, RASD1, RPS6KB2, SOCS3 |
| Ephrin Receptor Signaling | 0.006 | −0.7 | ARPC3, ARPC5L, ATF2, EFNA5, EPHB6, GNAL, GNG13, NCK2, PIK3CG, RASD1, SORBS1 |
| Sertoli Cell-Sertoli Cell Junction Signaling | 0.007 | AFDN, ATF2, CLDN2, CLDN23, CLDN4, MPP6, PRKG1, RASD1, SORBS1, SPTBN1, TJP1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oster, M.; Reyer, H.; Trakooljul, N.; Weber, F.M.; Xi, L.; Muráni, E.; Ponsuksili, S.; Rodehutscord, M.; Bennewitz, J.; Wimmers, K. Ileal Transcriptome Profiles of Japanese Quail Divergent in Phosphorus Utilization. Int. J. Mol. Sci. 2020, 21, 2762. https://doi.org/10.3390/ijms21082762
Oster M, Reyer H, Trakooljul N, Weber FM, Xi L, Muráni E, Ponsuksili S, Rodehutscord M, Bennewitz J, Wimmers K. Ileal Transcriptome Profiles of Japanese Quail Divergent in Phosphorus Utilization. International Journal of Molecular Sciences. 2020; 21(8):2762. https://doi.org/10.3390/ijms21082762
Chicago/Turabian StyleOster, Michael, Henry Reyer, Nares Trakooljul, Frank M. Weber, Lu Xi, Eduard Muráni, Siriluck Ponsuksili, Markus Rodehutscord, Jörn Bennewitz, and Klaus Wimmers. 2020. "Ileal Transcriptome Profiles of Japanese Quail Divergent in Phosphorus Utilization" International Journal of Molecular Sciences 21, no. 8: 2762. https://doi.org/10.3390/ijms21082762