The ENDpoiNTs Project: Novel Testing Strategies for Endocrine Disruptors Linked to Developmental Neurotoxicity

 , , , ,
, , , , 
Abstract
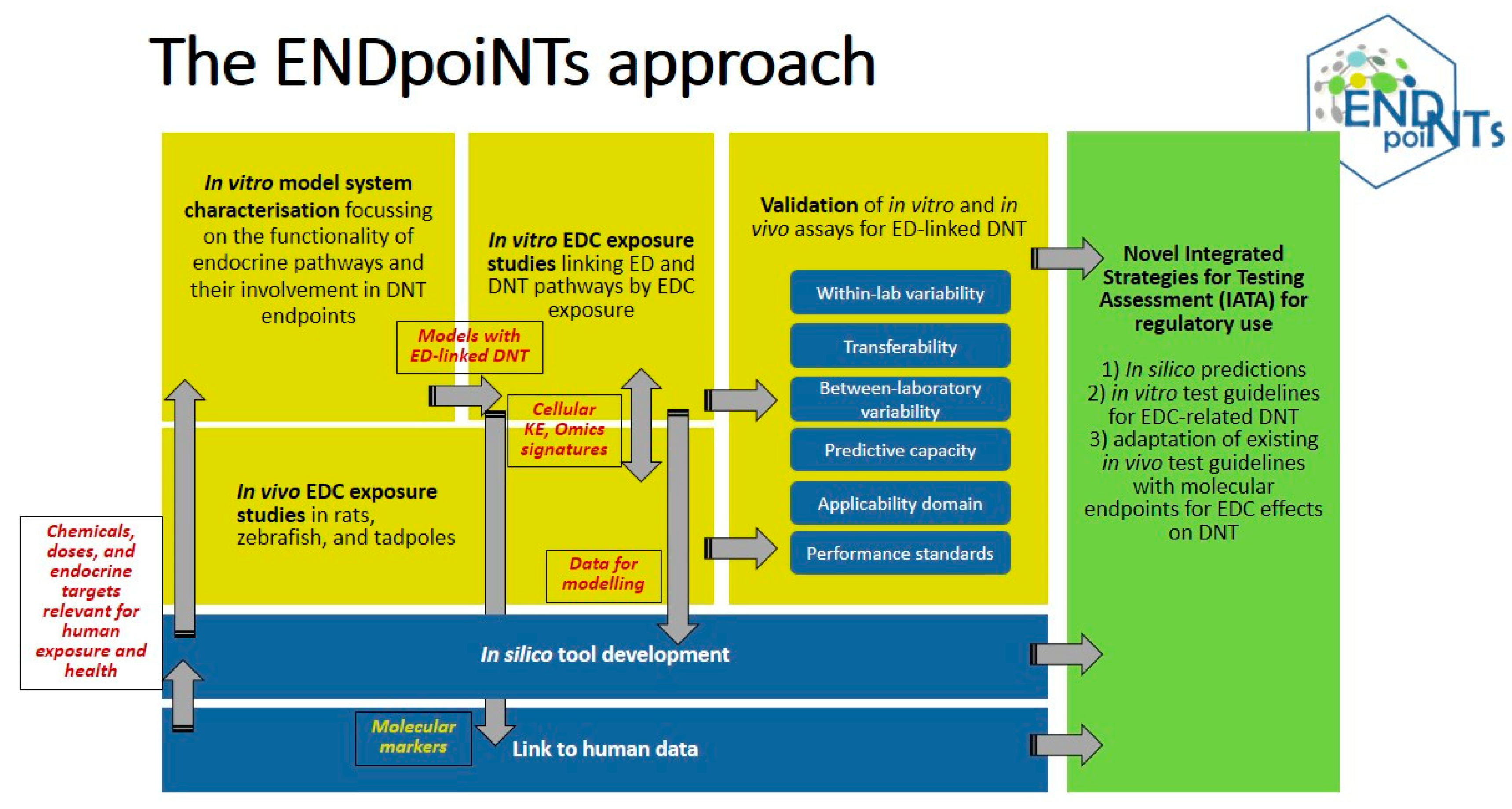
:1. Introduction
2. Human-Relevant Exposure to EDCs
3. Pathways Linking ED to DNT
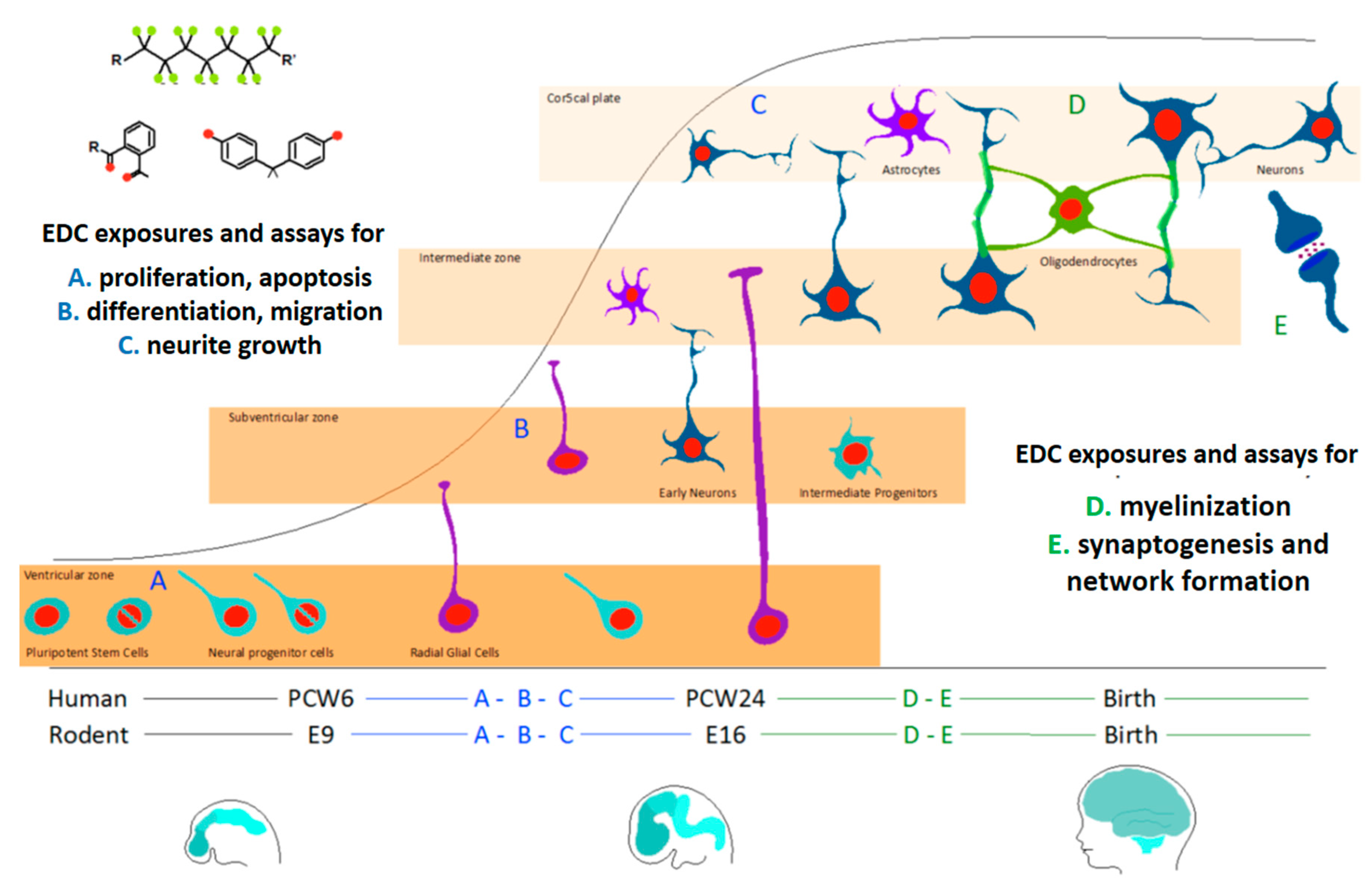
4. Relevant Models and Endpoints to Test ED-Induced DNT
5. Identification of Novel “Omics” Readouts
- (i)
- Transcriptomics. Transcriptomic analyses will assess the expression pattern changes of coding and non-coding RNAs, in the context of one or several DNT endpoints as a result of EDC exposure. Transcriptomics analyses will be performed in in vitro models and in specific brain areas of exposed rats, using RNA extracted both from bulk biological material and from single cells, to dissect population-specific pattern changes. Starting from the list of top regulated genes, functional annotation and data mining will be employed to gain knowledge on the affected cellular pathways and biological processes. Further analyses of regulatory motifs and associated master regulators will allow the reconstruction of gene networks and regulatory circuits affected by the exposures.
- (ii)
- Epigenomics. Epigenetic processes regulate temporal and spatial patterns of transcription and play a critical role in cell differentiation and tissue organization during development [56]. Epigenetic patterns at specific loci can change in response to environmental factors [57] and this can potentially affect health, depending on cell type and developmental stage. Epidemiological data provides increasing evidence for associations between chemical exposures and epigenetic changes [20] and an increasing number of experimental studies show that early-life exposures to EDCs and other neurotoxic compounds induce epigenetic changes, in particular DNA methylation changes, which, in some cases, have been linked to modifications in brain morphology and to adversities later in life (such as changes in anxiety-like, exploratory, and social behaviors) [20,57]. Therefore, changes in epigenetic patterns might serve as biomarkers for adverse effects on developmental processes induced by EDCs, as well as other environmental factors. In this context, ENDpoiNTs aims to assess epigenomic changes in selected in vitro and in vivo models and couple them to chemical exposures and later key events/adverse outcomes. The focus will be on DNA methylation and non-coding RNAs (including miRNA) patterns since these modifications are currently considered as the most promising epigenetic biomarkers for disease states. They are easily measured in accessible human tissue (blood, saliva, buccal epithelia) and are the most studied patterns in relation to environmental exposures [20].
- (iii)
- Metabolomics. Metabolomics provides a functional readout of the physiologic state of an organism as determined by the sum of its genetic predisposition, regulation, protein abundance, and environmental influences. In ENDpoiNTs, targeted and non-targeted metabolomic approaches will be employed in exposed rats (hippocampus and MPO), zebrafish, and various in vitro models to link disruption of endocrine pathways with DNT endpoints. The aim is to understand the molecular mechanism of ED-induced DNT using exploratory and hypothesis-driven metabolomic pathway analyses, and to relate the affected molecular pathways to phenotypical, developmental, behavioral, and cognitive changes in the in vitro and in vivo assays.
- (a)
- signatures that can be used as “fingerprints” for exposures with a specific endocrine mode of action;
- (b)
- signatures that can predict certain KE and AO with relevance for DNT;
- (c)
- panels of epigenomic, transcriptomic, and metabolomic markers which can be used to predict EDC-induced developmental neurotoxicity in both in vitro and in vivo models.
6. Exposure Modelling Using PBTK Models and In Silico Models (QSARs)
7. Linking Novel Test System Results to Population-Based Human Data Using Metrics of Risk Assessment
8. AOP Development, Incorporating Fundamental Neurodevelopmental Processes
9. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DAPI | 4′,6-diamidino-2-phenylindole |
| AOPs | adverse outcome pathways |
| AO | adverse outcome |
| AR | androgen receptors |
| BrdU | bromodeoxyuridine |
| CNS | central nervous system |
| DNT | developmental neurotoxicity |
| DCX | doublecortin |
| EDCs | endocrine-disrupting chemicals |
| ER | estrogen receptors |
| GFAP | glial fibrillary acidic protein |
| HI | hazard index |
| HQ | hazard quotients |
| HCIA | high content image acquisition |
| hiPSCs | human induced pluripotent stem cells |
| KE | key event |
| LDH | lactate dehydrogenase |
| LXRs | liver X/oxysterols receptors |
| MPO | medial preoptic area medial preoptic area |
| MEA | microelectrode arrays |
| MAP2 | microtubule-associated protein 2 |
| MIE | molecular initiating event |
| MBP | myelin basic protein |
| NPCs | neural progenitor cells |
| OECD | Organisation for Economic Co-operation and Development |
| PPARs | peroxisome proliferator-activated receptors |
| PBTK | physiologically based toxicokinetic |
| POD | point of departure |
| PCBs | polychlorinated biphenyls |
| PBDEs | polybrominated diphenyl ethers |
| PSD95 | postsynaptic density protein 95 |
| PR | progesterone receptor |
| PGE2 | prostaglandin E2 |
| QSAR | quantitative structure–activity relationship |
| RAR | retinoic acid receptors |
| RXR | retinoic X receptors |
| SMRI | similar mixture risk indicator |
| SOPs | standard operating procedures |
| SVZ | subventricular zone |
| TGs | test guidelines |
| TH | thyroid hormone |
| TR | thyroid receptors |
| VDR | vitamin D3 receptor |
References
- WHO (World Health Organization)/UNEP (United Nations Environment Programme). The State-of-the-Science of Endocrine Disrupting Chemicals—2012; Bergman, Å., Heindel, J.J., Jobling, S., Kidd, K.A., Zoeller, R.T., Eds.; UNEP/WHO: Geneva, Switzerland, 2013; Available online: http://www.who.int/ceh/publications/endocrine/en/index.html (accessed on 12 March 2020).
- Demeneix, B.; Slama, R. Endocrine Disruptors: From Scientific Evidence to Human Health Protection. In Report Commissioned by the PETI Committee of the European Parliament; Policy Department for Citizen’s Rights and Constitutional Affairs, 2019; Available online: http://www.europarl.europa.eu/RegData/etudes/STUD/2019/608866/IPOL_STU(2019)608866_EN.pdf (accessed on 10 February 2020).
- Bal-Price, A.; Pistollato, F.; Sachana, M.; Bopp, S.K.; Munn, S.; Worth, A. Strategies to improve the regulatory assessment of developmental neurotoxicity (DNT) using in vitro methods. Toxicol. Appl. Pharmacol. 2018, 354, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P.; Landrigan, P.J. Developmental neurotoxicity of industrial chemicals. Lancet 2006, 368, 2167–2178. [Google Scholar] [CrossRef]
- Grandjean, P.; Landrigan, P.J. Neurobehavioural effects of developmental toxicity. Lancet Neurol. 2014, 13, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Bornehag, C.G.; Reichenberg, A.; Hallerback, M.U.; Wikstrom, S.; Koch, H.M.; Jonsson, B.A.; Swan, S.H. Prenatal exposure to acetaminophen and children’s language development at 30 months. Eur. Psychiatry 2018, 51, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Boucher, O.; Muckle, G.; Bastien, C.H. Prenatal exposure to polychlorinated biphenyls: A neuropsychologic analysis. Environ. Health Perspect. 2009, 117, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.M.; Yolton, K.; Stacy, S.L.; Erar, B.; Papandonatos, G.D.; Bellinger, D.C.; Lanphear BPChen, A. Prenatal environmental chemical exposures and longitudinal patterns of child neurobehavior. Neurotoxicology 2017, 62, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Engel, S.M.; Miodovnik, A.; Canfield, R.L.; Zhu, C.; Silva, M.J.; Calafat, A.M.; Wolff, M.S. Prenatal phthalate exposure is associated with childhood behavior and executive functioning. Environ. Health Perspect. 2010, 118, 565–571. [Google Scholar] [CrossRef]
- Vuong, A.M.; Yolton, K.; Poston, K.L.; Xie, C.; Webster, G.M.; Sjödin, A.; Braun, J.M.; Dietrich, K.N.; Lanphear, B.P.; Chen, A. Childhood polybrominated diphenyl ether (PBDE) exposure and executive function in children in the HOME Study. Int. J. Hyg. Environ. Health 2018, 221, 87–94. [Google Scholar] [CrossRef]
- Mallozzi, M.; Bordi, G.; Garo, C.; Caserta, D. The effect of maternal exposure to endocrine disrupting chemicals on fetal and neonatal development: A review on the major concerns. Birth Defects Res. C Embryo Today 2016, 108, 224–242. [Google Scholar] [CrossRef]
- Andre, S.M.; Markowski, V.P. Learning deficits expressed as delayed extinction of a conditioned running response following perinatal exposure to vinclozolin. Neurotoxicol. Teratol. 2006, 28, 482–488. [Google Scholar] [CrossRef]
- Dingemans, M.M.; van den Berg, M.; Westerink, R.H. Neurotoxicity of brominated flame retardants: (in) direct effects of parent and hydroxylated polybrominated diphenyl ethers on the (developing) nervous system. Environ. Health Perspect. 2011, 119, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and transgenerational effects of low doses of perinatal di-(2-ethylhexyl) phthalate (DEHP) on social behaviors in mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef] [PubMed]
- Roegge, C.S.; Seo, B.W.; Crofton, K.M.; Schantz, S.L. Gestational-lactational exposure to Aroclor 1254 impairs radial-arm maze performance in male rats. Toxicol. Sci. 2000, 57, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Zhang, J.; Wang, Y.M.; Ye, Y.P.; Luo, Q. Perinatal exposure to bisphenol-A impairs learningmemory by concomitant down-regulation of N-methyl-D-aspartate receptors of hippocampus in male offspring mice. Horm. Behav. 2010, 58, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.E.; Desesso, J.M.; Hurtt, M.E.; Cappon, G.D. Postnatal growth and morphological development of the brain: A species comparison. Birth Defects Res. B Dev. Reprod. Toxicol. 2006, 77, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Silbereis, J.C.; Pochareddy, S.; Zhu, Y.; Li, M.; Sestan, N. The Cellular and Molecular Landscapes of the Developing Human Central Nervous System. Neuron 2016, 89, 248–268. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.M. Estradiol and the developing brain. Physiol. Rev. 2008, 88, 91–124. [Google Scholar] [CrossRef] [Green Version]
- Tanner, E.M.; Hallerbäck, M.U.; Wikström, S.; Lindh, C.; Kiviranta, H.; Gennings, C.; Bornehag, C.G. Early prenatal exposure to suspected endocrine disruptor mixtures is associated with lower IQ at age seven. Environ. Int. 2020, 134, 105185. [Google Scholar] [CrossRef]
- Jacobs, M.N.; Marczylo, E.L.; Guerrero-Bosagna, C.; Ruegg, J. Marked for Life: Epigenetic Effects of Endocrine Disrupting Chemicals. Ann. Rev. Environ. Resour. 2017, 42, 105–160. [Google Scholar] [CrossRef]
- Bayless, D.W.; Shah, N.M. Genetic dissection of neural circuits underlying sexually dimorphic social behaviours. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150109. [Google Scholar] [CrossRef] [Green Version]
- Küppers, E.; Ivanova, T.; Karolczak, M.; Lazarov, N.; Föhr, K.; Beyer, C. Classical and nonclassical estrogen action in the developing midbrain. Horm. Behav. 2001, 40, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Hines, M. Sex steroids and human behavior: Prenatal androgen exposure and sex-typical play behavior in children. Ann. N. Y. Acad. Sci. 2003, 1007, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Bakker, J. The Sexual Differentiation of the Human Brain: Role of Sex Hormones Versus Sex Chromosomes. Curr. Top. Behav. Neurosci. 2019, 43, 45–67. [Google Scholar] [PubMed] [Green Version]
- Denley, M.C.S.; Gatford, N.J.F.; Sellers, K.J.; Srivastava, D.P. Estradiol and the Development of the Cerebral Cortex: An Unexpected Role? Front. Neurosci. 2018, 12, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, J. Thyroid hormone receptors in brain development and function. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Willoughby, K.A.; McAndrews, M.P.; Rovet, J.F. Effects of maternal hypothyroidism on offspring hippocampus and memory. Thyroid 2014, 24, 576–584. [Google Scholar] [CrossRef]
- Korevaar, T.I.; Muetzel, R.; Medici, M.; Chaker, L.; Jaddoe, V.W.; de Rijke, Y.B.; Steegers, E.A.; Visser, T.J.; White, T.; Peeters, R.P.; et al. Association of maternal thyroid function during early pregnancy with offspring IQ and brain morphology in childhood: A population-based prospective cohort study. Lancet Diabetes Endocrinol. 2016, 4, 35–43. [Google Scholar] [CrossRef]
- Remaud, S.; Ortiz, F.C.; Perret-Jeanneret, M.; Aigrot, M.S.; Gothie, J.D.; Fekete, C.; Kvárta-Papp, Z.; Gereben, B.; Langui, D.; Angulo, M.C.; et al. Transient hypothyroidism favors oligodendrocyte generation providing functional remyelination in the adult mouse brain. Elife 2017, 6, e29996. [Google Scholar] [CrossRef]
- Dach, K.; Bendt, F.; Huebenthal, U.; Giersiefer, S.; Lein, P.J.; Heuer, H.; Fritsche, E. BDE-99 impairs differentiation of human and mouse NPCs into the oligodendroglial lineage by species-specific modes of action. Sci. Rep. 2017, 7, 44861. [Google Scholar] [CrossRef]
- López-Juárez, A.; Remaud, S.; Hassani, Z.; Jolivet, P.; Simons, J.P.; Sontag, T.; Yoshikawa, K.; Price, J.; Morvan-Dubois, G.; Demeneix, B.A. Thyroid hormone signaling acts as a neurogenic switch by repressing Sox2 in the adult neural stem cell niche. Cell Stem Cell 2012, 10, 531–543. [Google Scholar] [CrossRef] [Green Version]
- McGowan, P.O.; Matthews, S.G. Prenatal Stress, Glucocorticoids, and Developmental Programming of the Stress Response. Endocrinology 2018, 159, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Gkikas, D.; Tsampoula, M.; Politis, P.K. Nuclear receptors in neural stem/progenitor cell homeostasis. Cell. Mol. Life Sci. 2017, 74, 4097–4120. [Google Scholar] [CrossRef] [PubMed]
- Maden, M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat. Rev. Neurosci. 2007, 8, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.K. Progesterone receptors and neural development: A gap between bench and bedside? Endocrinology 2008, 149, 2743–2749. [Google Scholar] [CrossRef] [Green Version]
- González-Orozco, J.C.; Camacho-Arroyo, I. Progesterone Actions during Central Nervous System Development. Front. Neurosci. 2019, 13, 503. [Google Scholar] [CrossRef] [Green Version]
- Kesby, J.P.; Eyles, D.W.; Burne, T.H.; McGrath, J.J. The effects of vitamin D on brain development and adult brain function. Mol. Cell. Endocrinol. 2011, 347, 121–127. [Google Scholar] [CrossRef]
- Courtney, R.; Landreth, G.E. LXR Regulation of Brain Cholesterol: From Development to Disease. Trends Endocrinol. Metab. 2016, 27, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Wright, C.L.; Burks, S.R.; McCarthy, M.M. Identification of Prostaglandin E2 Receptors Mediating Perinatal Masculinization of Adult Sex Behavior and Neuroanatomical Correlates. Dev. Neurobiol. 2008, 68, 1406–1419. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.Z.; Kriebel, D.; Herbert, M.R.; Bornehag, C.G.; Swan, S.H. Prenatal paracetamol exposure and child neurodevelopment: A review. Horm. Behav. 2018, 101, 125–147. [Google Scholar] [CrossRef]
- Bal-Price, A.; Hogberg, H.T.; Crofton, K.M.; Daneshian, M.; FitzGerald, R.E.; Fritsche, E.; Heinonen, T.; Bennekou, S.H.; Klima, S.; Sachana, M.; et al. Recommendation on test readiness criteria for new approach methods in toxicology: Exemplified for developmental neurotoxicity. ALTEX 2018, 35, 306–352. [Google Scholar] [CrossRef]
- Fritsche, E.; Grandjean, P.; Crofton, K.M.; Aschner, M.; Goldberg, A.; Heinonen, T.; Hessel, E.V.; Hogberg, H.T.; Bennekou, S.H.; Lein, P.J.; et al. Consensus statement on the need for innovation, transition and implementation of developmental neurotoxicity (DNT) testing for regulatory purposes. Toxicol. Appl. Pharmacol. 2018, 354, 3–6. [Google Scholar] [CrossRef] [PubMed]
- Fritsche, E.; Crofton, K.M.; Hernandez, A.F.; Bennekou, S.H.; Leist, M.; Bal-Price, A.; Reaves, E.; Wilks, M.F.; Terron, A.; Sachana, M.; et al. OECD/EFSA workshop on developmental neurotoxicity (DNT): The use of non-animal test methods for regulatory purposes. ALTEX 2017, 34, 311–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aschner, M.; Ceccatelli, S.; Daneshian, M.; Fritsche, E.; Hasiwa, N.; Hartung, T.; Hogberg, H.T.; Leist, M.; Li, A.; Mundy, W.R.; et al. Reference compounds for alternative test methods to indicate developmental neurotoxicity (DNT) potential of chemicals: Example lists and criteria for their selection and use. ALTEX 2017, 34, 49–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporale, N.; Testa, G. At the Intersection of Epigenetics and Regeneration: An Analysis of the Experimental Outlook of Organoid Technology. In Epigenetics and Regeneration; Palacios, D., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 11, pp. 385–402. [Google Scholar]
- Attoff, K.; Gliga, A.; Lundqvist, J.; Norinder, U.; Forsby, A. Whole genome microarray analysis of neural progenitor C17.2 cells during differentiation and validation of 30 neural mRNA biomarkers for estimation of developmental neurotoxicity. PLoS ONE 2017, 12, e0190066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gothié, J.D.; Sébillot, A.; Luongo, C.; Legendre, M.; Van, C.N.; Le Blay, K.; Perret-Jeanneret, M.; Remaud, S.; Demeneix, B.A. Adult neural stem cell fate is determined by thyroid hormone activation of mitochondrial metabolism. Mol. Metab. 2017, 6, 1551–1561. [Google Scholar] [CrossRef] [PubMed]
- Lubetzki, C.; Demerens, C.; Anglade, P.; Villarroya, H.; Frankfurter, A.; Lee, V.M.; Zalc, B. Even in culture, oligodendrocytes myelinate solely axons. Proc. Natl. Acad. Sci. USA 1993, 90, 6820–6824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.P.; Lynch, B.S.; Curry-Chisolm, I.M.; Shafer, T.J.; Strickland, J.D. Assaying spontaneous network activity and cellular viability using multi-well microelectrode arrays. Methods Mol. Biol. 2017, 1601, 153–170. [Google Scholar]
- Schmuck, M.R.; Temme, T.; Dach, K.; de Boer, D.; Barenys, M.; Bendt, F.; Mosig, A.; Fritsche, E. Omnisphero: A high-content image analysis (HCA) approach for phenotypic developmental neurotoxicity (DNT) screenings of organoid neurosphere cultures in vitro. Arch. Toxicol. 2017, 91, 2017–2028. [Google Scholar] [CrossRef]
- López-Tobón, A.; Villa, C.E.; Cheroni, C.; Trattaro, S.; Caporale, N.; Conforti, P.; Iennaco, R.; Lachgar, M.; Rigoli, M.T.; de la Cruz, B.M.; et al. Longitudinal dissection in brain organoids at single cell resolution uncovers the developmental role of GSK3 in human corticogenesis. BioRxiv 2018. [Google Scholar] [CrossRef]
- Gatewood, J.D.; Wills, A.; Shetty, S.; Xu, J.; Arnold, A.P.; Burgoyne, P.S.; Rissman, E.F. Sex chromosome complement and gonadal sex influence aggressive and parental behaviors in mice. J. Neurosci. 2006, 26, 2335–2342. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, M.M.; Arnold, A.P. Reframing sexual differentiation of the brain. Nat. Neurosci. 2011, 14, 677–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krebs, A.; Waldmann, T.; Wilks, M.F.; Van Vugt-Lussenburg, B.M.; Van der Burg, B.; Terron, A.; Steger-Hartmann, T.; Ruegg, J.; Rovida, C.; Pedersen, E.; et al. Template for the description of cell-based toxicological test methods to allow evaluation and regulatory use of the data. ALTEX 2019, 36, 682–699. [Google Scholar] [CrossRef]
- Bale, T.L. Epigenetic and transgenerational reprogramming of brain development. Nat. Rev. Neurosci. 2015, 16, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Marczylo, E.L.; Jacobs, M.N.; Gant, T.W. Environmentally induced epigenetic toxicity: Potential public health concerns. Crit. Rev. Toxicol. 2016, 46, 676–700. [Google Scholar] [CrossRef] [PubMed]
- Lioy, P.J.; Hauser, R.; Gennings, C.; Koch, H.M.; Mirkes, P.E.; Schwetz, B.A.; Kortenkamp, A. Assessment of phthalates/phthalate alternatives in children’s toys and childcare articles: Review of the report including conclusions and recommendation of the Chronic Hazard Advisory Panel of the Consumer Product Safety Commission. J. Expo. Sci. Environ. Epidemiol. 2015, 25, 343–353. [Google Scholar] [CrossRef] [PubMed]
- AOP-Wiki. Available online: https://aopwiki.org/aops (accessed on 17 February 2020).
- Organisation for Economic Co-operation and Development. Users’ Handbook Supplement to the Guidance Document for Developing and Assessing AOPs. ENV/JM/MONO (2016)12. Available online: http://www.oecd.org/officialdocuments/publicdisplaydocumentpdf/?cote=env/jm/mono(2016)12&doclanguage=en (accessed on 17 February 2020).



{kind=link}
{kind=link}
| Nr. | Chemical Name | Abrev. |
|---|---|---|
| 1 | Benzyl butyl phthalate | BBzP |
| 2 | Dibutyl benzene-1,2-dicarboxylate | DBP |
| 3 | Di-ethylphthalic acid ester | DEP |
| 4 | Bis (2-ethylhexyl) phthalate | DEHP |
| 5 | Di-isodecylphthalic acid ester | DIDP |
| 6 | Di-isononyl phthalate | DINP |
| 7 | Monobutyl phthalate | MBP |
| 8 | Mono-ethylphthalic acid ester | MEP |
| 9 | Monobenzyl phthalate | MBzP |
| 10 | Mono (2-ethylhexyl)phthalate | MEHP |
| 11 | Mono-isodecylphthalic acid ester | MIDP |
| 12 | Monoisononyl phthalate | MINP |
| 13 | Bisphenol A | BPA |
| 14 | Bisphenol F | BPF |
| 15 | Perfluorohexanesulfonic acid | PFHxS |
| 16 | Perfluorooctanoic acid | PFOA |
| 17 | Perfluorooctanesulfonic acid | PFOS |
| 18 | 2,2′,3,4,4′,5,5′-Heptachlorobiphenyl | PCB-180 |
| 19 | 3,5,6-trichloro-2-pyridinol | TCP |
| 20 | Dichlordiphenyldichlorethylen | 4,4′-DDE |
| 21 | 3-Phenoxybenzoic acid | 3-PBA |
| 22 | Aldicarb | |
| 23 | Carbaryl | |
| 24 | Cypermethrin | |
| 25 | Endosulfan | |
| 26 | Methomyl | |
| 27 | Permethrin |
| Target Name | Chemicals | Type of Interaction |
|---|---|---|
| Estrogen receptor alpha and beta | BPA | Agonistic |
| Estradiol 17-beta-dehydrogenase 1 | Endosulfan | Binding |
| Androgen receptor | BPA, endosulfan, 4,4′-DDE, PFHxS | Antagonistic |
| Thyroid hormone receptor beta | BPA, cypermethrin, endosulfan, PFHxS, permethrin | Agonistic |
| Glucocorticoid receptor | BPA, MEHP | Agonistic |
| Retinoic acid receptor alpha, beta, gamma | 3-PBA, MEP, MIDP, MINP | Agonistic/not predicted |
| Progesterone receptor | BPA, MEHP | Not predicted |
| Peroxisome proliferator-activated receptor alpha, gamma, delta | Permethrin | Agonistic |
| Vitamin D3 receptor | Permethrin | Agonistic |
| Oxysterols receptor LXR-alpha | PFHxS, PFOA, PFOS | Agonistic |
| Prostaglandin E2 receptor EP1, EP2, EP3, EP4 subtype | MBzP |
| Assay | Endpoint | Readout | Ref |
|---|---|---|---|
| Mouse C17.2 NPC line | Proliferation | LDH release | [47] |
| Differentation | bIII-tubulin staining | ||
| Neurite growth | Quantification of neurite length in cells stained with bIII-tubulin and DAPI | ||
| Mouse primary NPC (SVZ) | Proliferation | Sphere size, BrdU, Ki67/PH3 stainings | [48] |
| Differentiation | DCX staining | ||
| Mouse primary neuron-oligodendrocyte co-cultures | Differentiation | MBP staining | [49] |
| Myelination | MBP staining | ||
| Rat NPCs (different brain regions) | Proliferation | Sphere Size, BrdU staining | |
| Differentiation | GFAP (HCIA), O4 staining (HCIA) | ||
| Migration | HCIA of the bIII-tubulin or O4 stained cells which migrate from the sphere core stained with DAPI | ||
| Neurite growth | DAPI staining, MAP2, bIII-tubulin staining (HCIA) | ||
| Myelination | qRT-PCR for expression of markers of oligodendrocyte differentiation and maturation | ||
| Network formation and activity | Synapsin/PSD95 staining | ||
| Rat primary cortical cultures | Network formation and activity | MEA | [50] |
| Human primary NPC | Proliferation | Sphere Size, BrdU staining | [42] |
| Differentiation | GFAP staining (HCIA), O4 staining (HCIA), bIII-tubulin staining (HCIA) | [42] | |
| Migration | HCIA of the bIII-tubulin or O4 stained cells which migrate from the sphere core stained with DAPI | [42] | |
| Neurite growth | DAPI staining, MAP2 staining, bIII-tubulin staining | [51] | |
| Myelination | rtRT-PCR for expression of markers of oligodendrocyte differentiation (and maturation?) | [42] | |
| Network formation and activity | MEA | ||
| Human iPSC-derived NPC (different brain regions) | Proliferation | Sphere Size, BrdU | |
| Differentiation | GFAP staining | ||
| Migration | HCIA of the bIII-tubulin or O4 stained cells which migrate from the sphere core stained with DAPI | [42] | |
| Neurite growth | DAPI staining, bIII-tubulin staining | ||
| Network formation and activity | Synapsin/PSD95 staining, MEA | ||
| Human iPSC-derived NCC cMINC (UKN2) | Migration | HCIA of viable cells (stained with Hoechst and calcein) which migrate to a previously unpopulated plate area | [42] |
| Human iPSC-derived NCC PeriTox (UKN5) | Neurite growth | HCIA of cells stained with Hoechst and calcein | [42] |
| Human immortalized primary NPC NeuriTox (UKN4) | Neurite growth | HCIA of cells stained with Hoechst and calcein | [42] |
| Human iPSC-derived neuronal co-cultures | Network formation and activity | MEA | |
| Human iPSC-derived cortical brain organoids | Network formation and activity | MEA | |
| Molecular changes | Single cell transcriptomics, epigenomics | [52] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupu, D.; Andersson, P.; Bornehag, C.-G.; Demeneix, B.; Fritsche, E.; Gennings, C.; Lichtensteiger, W.; Leist, M.; Leonards, P.E.G.; Ponsonby, A.-L.; et al. The ENDpoiNTs Project: Novel Testing Strategies for Endocrine Disruptors Linked to Developmental Neurotoxicity. Int. J. Mol. Sci. 2020, 21, 3978. https://doi.org/10.3390/ijms21113978
Lupu D, Andersson P, Bornehag C-G, Demeneix B, Fritsche E, Gennings C, Lichtensteiger W, Leist M, Leonards PEG, Ponsonby A-L, et al. The ENDpoiNTs Project: Novel Testing Strategies for Endocrine Disruptors Linked to Developmental Neurotoxicity. International Journal of Molecular Sciences. 2020; 21(11):3978. https://doi.org/10.3390/ijms21113978
Chicago/Turabian StyleLupu, Diana, Patrik Andersson, Carl-Gustaf Bornehag, Barbara Demeneix, Ellen Fritsche, Chris Gennings, Walter Lichtensteiger, Marcel Leist, Pim E. G. Leonards, Anne-Louise Ponsonby, and et al. 2020. "The ENDpoiNTs Project: Novel Testing Strategies for Endocrine Disruptors Linked to Developmental Neurotoxicity" International Journal of Molecular Sciences 21, no. 11: 3978. https://doi.org/10.3390/ijms21113978