Heart Failure Disturbs Gut–Blood Barrier and Increases Plasma Trimethylamine, a Toxic Bacterial Metabolite

 ,
,
Abstract
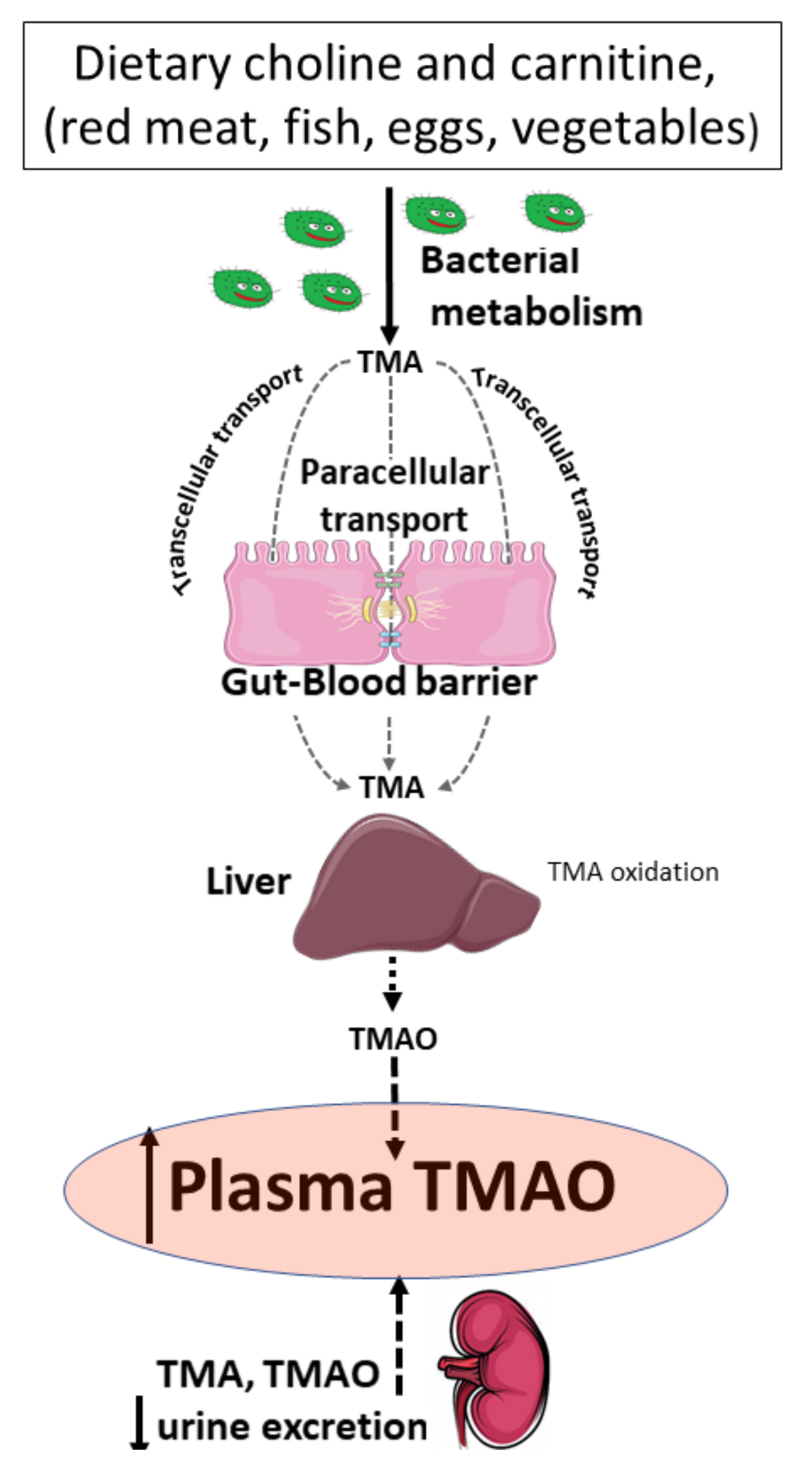
:1. Introduction
2. Results
2.1. General Metabolic and Kidney Parameters
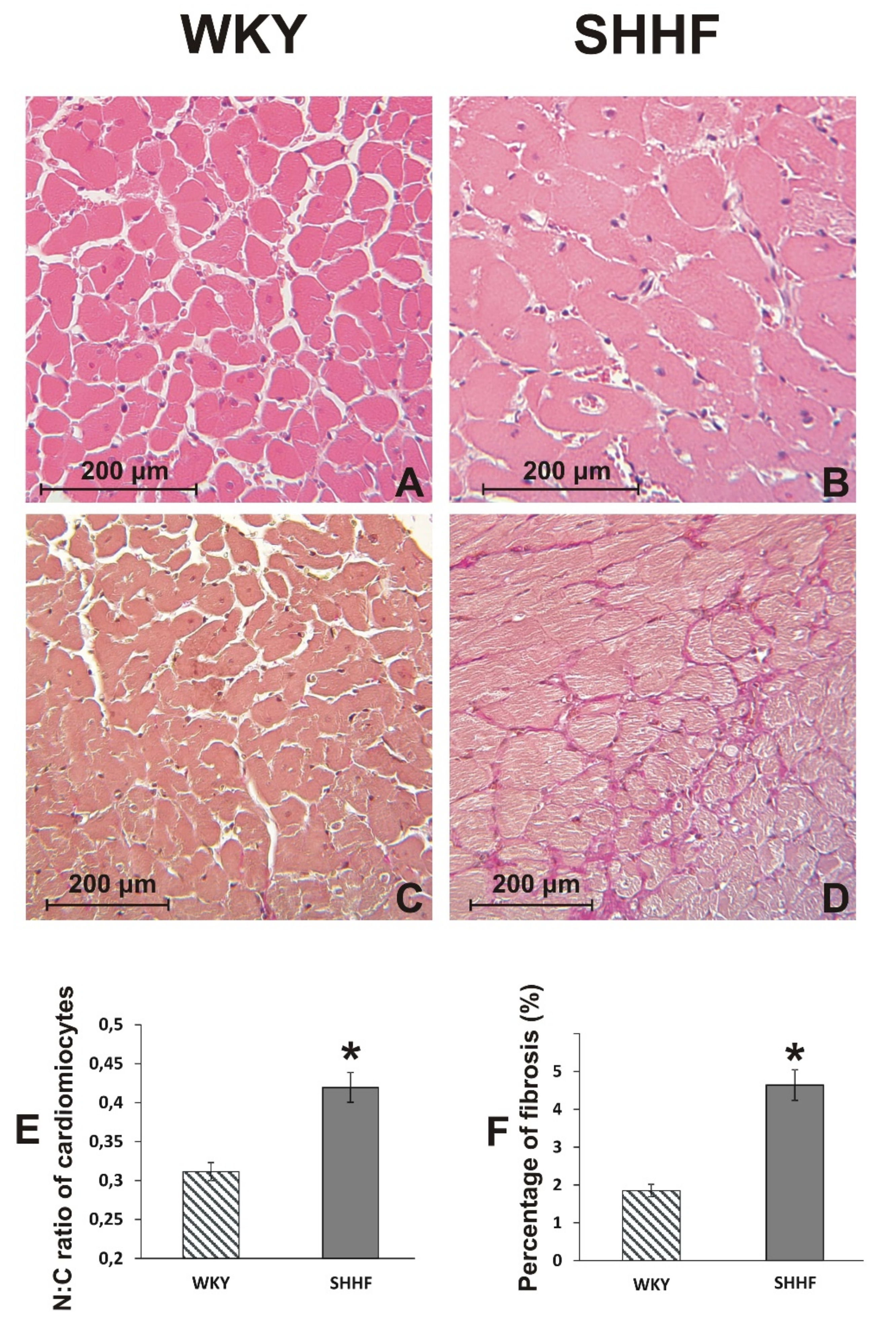
2.2. Cardiovascular Parameters
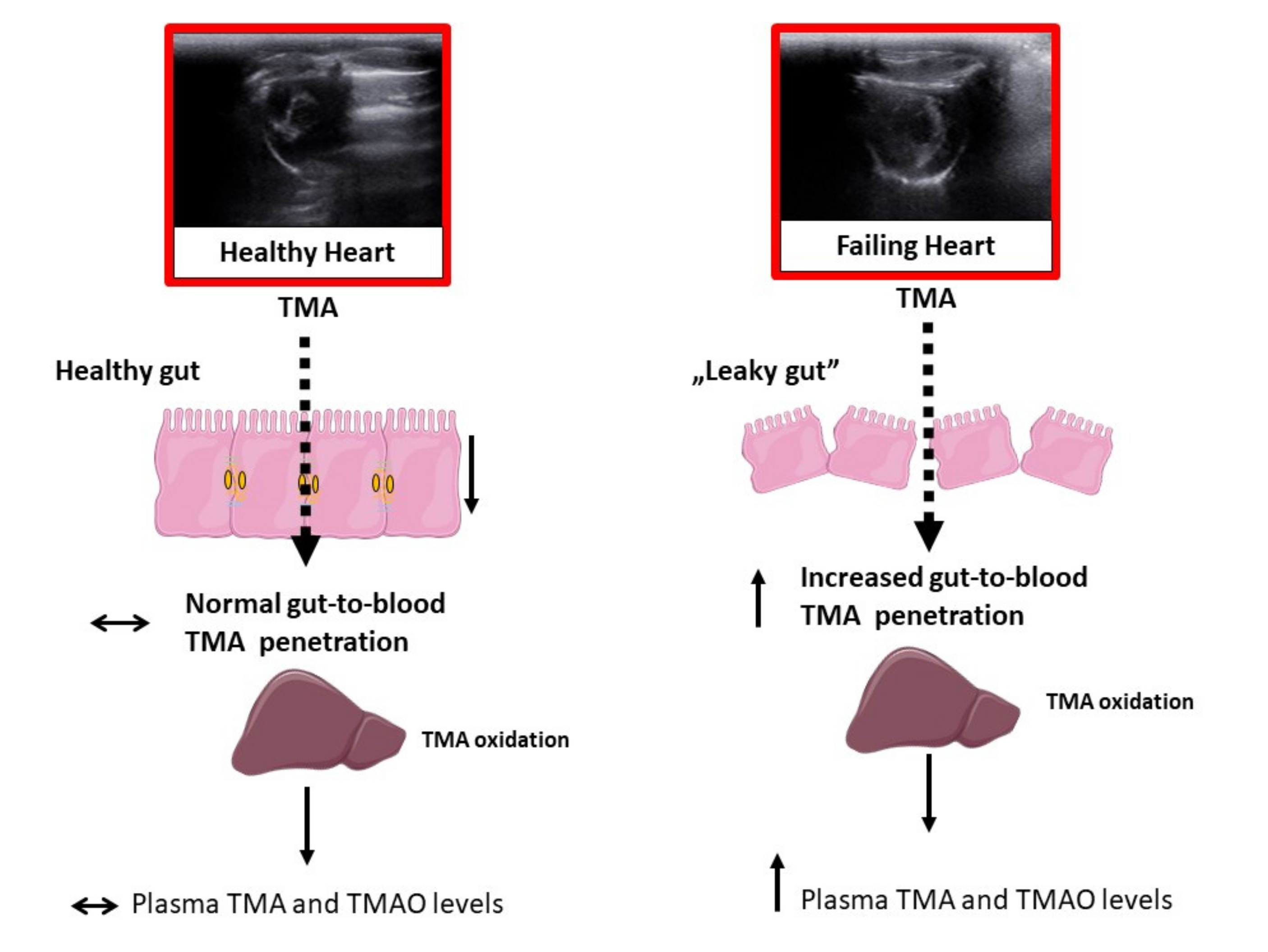
2.3. Plasma Level of TMA and TMAO in WKY Rats and SHHF Rats with Heart Failure
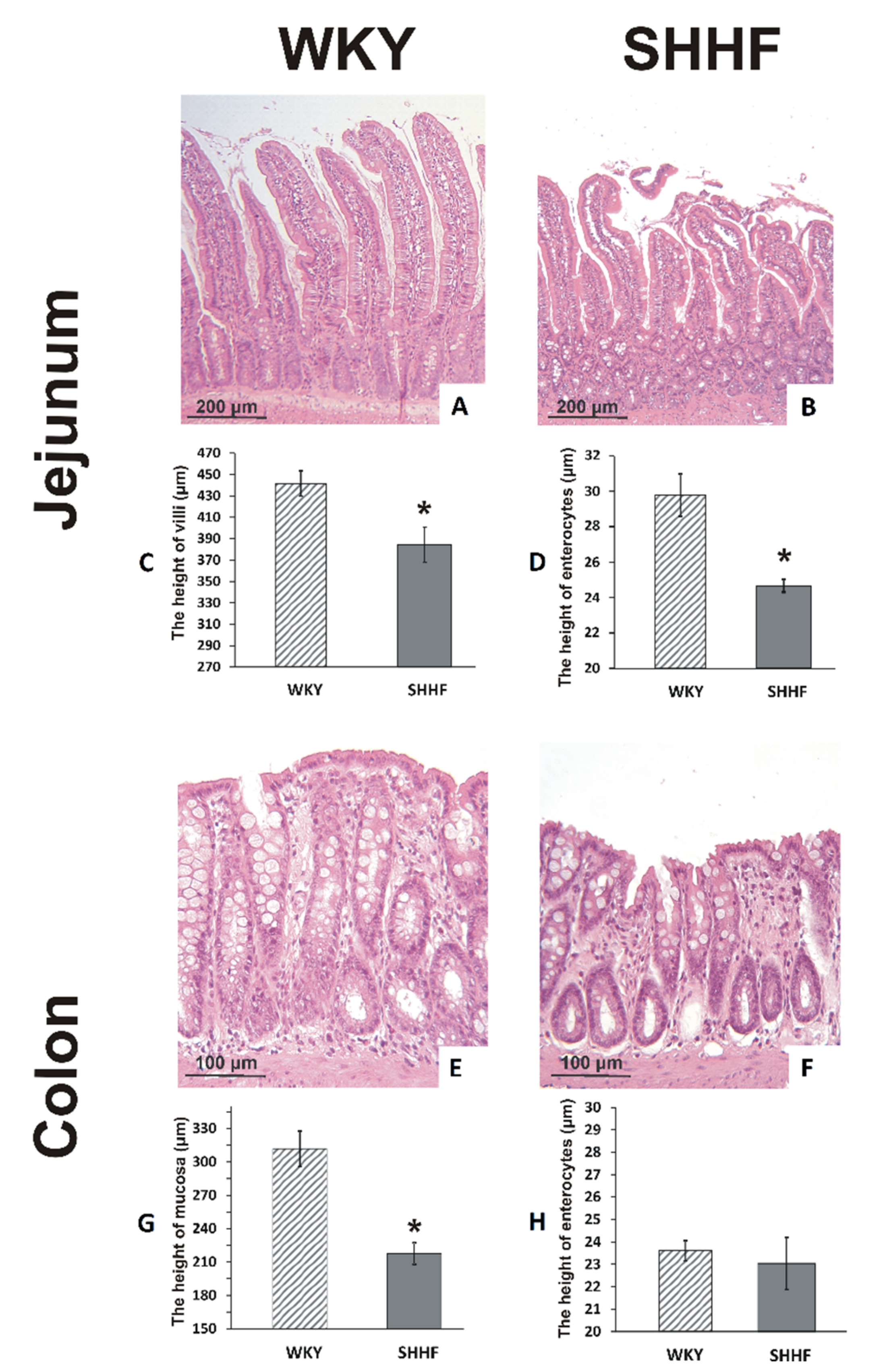
2.4. Intestines
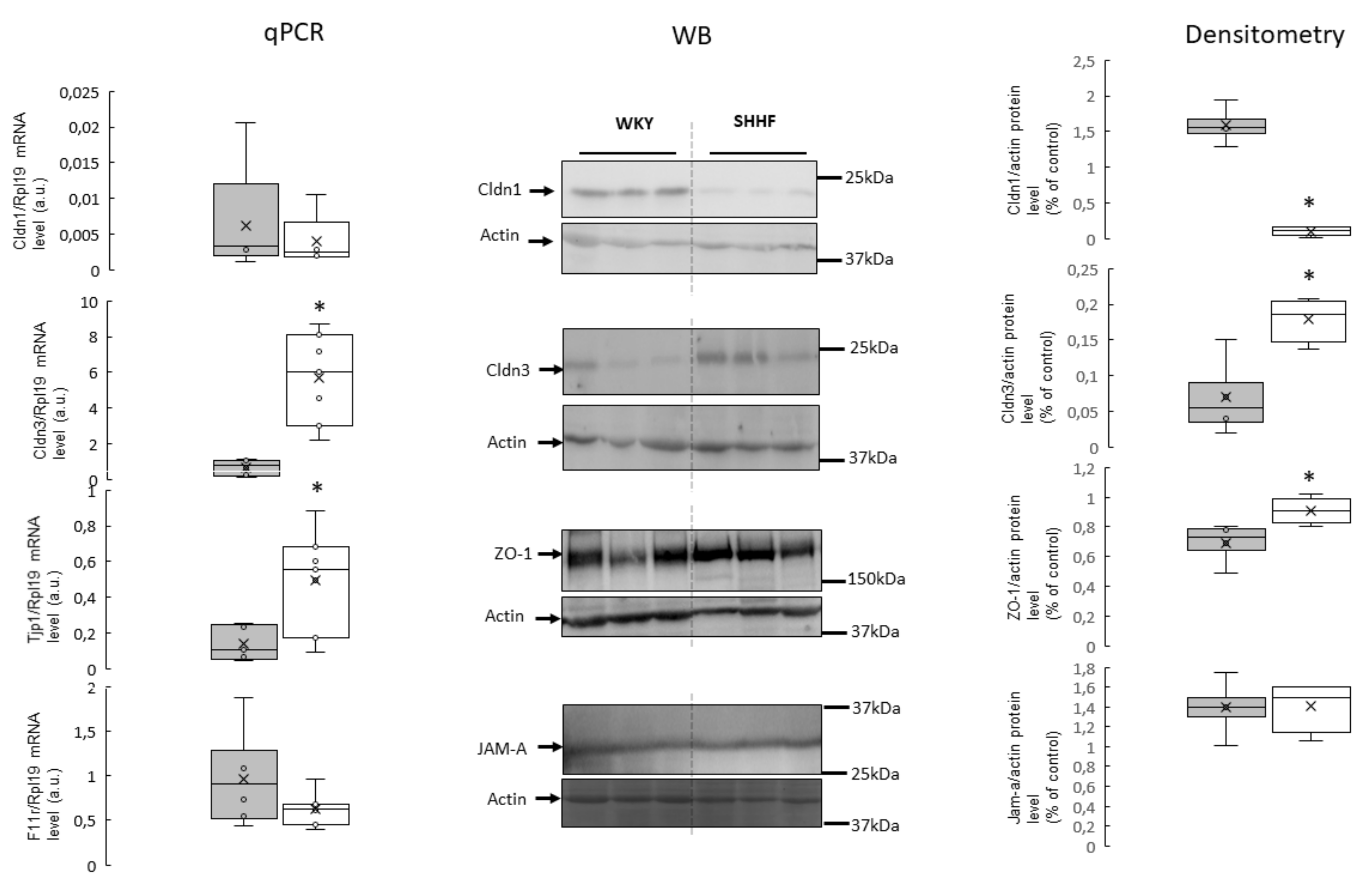
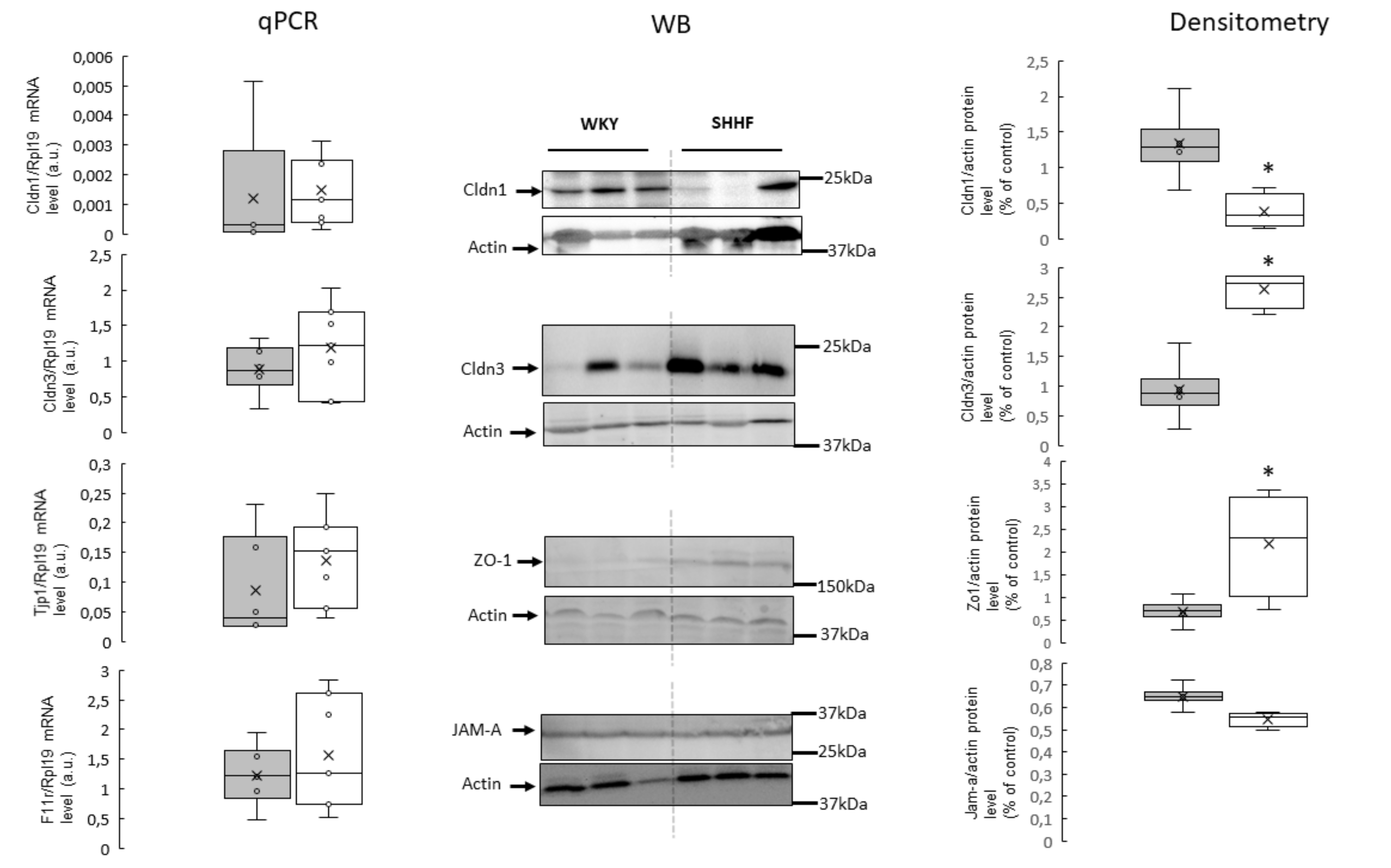
2.5. Tight Junction mRNA and Protein Levels
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Metabolic and Hemodynamic Measurements
4.3. Stools, Plasma and Urine TMA, TMAO and General Biochemistry Evaluation
4.4. Histopathological Evaluation
4.5. ELISA Test
4.6. RNA Isolation and RT-qPCR
4.7. Protein Extraction and Immunoblotting Analysis
4.8. Data Analysis and Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| TMA | trimethylamine |
| TMAO | Trimethylamine N-oxide |
| WKY | Wistar Kyoto rats |
| SHHF | Spontaneously Hypertensive-Heart-Failure rats |
| Cldn1 | claudin 1 |
| Cldn3 | claudin 3 |
| ZO-1 | zonula occludens-1 |
| JAM-A | junctional adhesion molecule A |
| ABP | Arterial blood pressure |
| LVEDP | Left ventricular end-diastolic pressure |
References
- Mensah, G.A.; Roth, G.A.; Fuster, V. The Global Burden of Cardiovascular Diseases and Risk Factors. J. Am. Coll. Cardiol. 2019, 74, 2529–2532. [Google Scholar] [CrossRef] [PubMed]
- Ahmadmehrabi, S.; Tang, W.W. Gut microbiome and its role in cardiovascular diseases. Curr. Opin. Cardiol. 2017, 32, 761. [Google Scholar] [CrossRef] [PubMed]
- Landfald, B.; Valeur, J.; Berstad, A.; Raa, J. Microbial trimethylamine-N-oxide as a disease marker: Something fishy? Microb. Ecol. Health Dis. 2017, 28, 1327309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, K.; Bielinska, K.; Gawrys-Kopczynska, M.; Ufnal, M. TMA (trimethylamine), but not its oxide TMAO (trimethylamine-oxide), exerts haemodynamic effects: Implications for interpretation of cardiovascular actions of gut microbiome. Cardiovasc. Res. 2019, 115, 1948–1949. [Google Scholar] [CrossRef] [PubMed]
- Jaworska, K.; Hering, D.; Mosieniak, G.; Bielak-Zmijewska, A.; Pilz, M.; Konwerski, M.; Gasecka, A.; Kaplon-Cieslicka, A.; Filipiak, K.; Sikora, E. TMA, A Forgotten Uremic Toxin, but Not TMAO, Is Involved in Cardiovascular Pathology. Toxins 2019, 11, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, K.; Konop, M.; Hutsch, T.; Perlejewski, K.; Radkowski, M.; Grochowska, M.; Bielak-Zmijewska, A.; Mosieniak, G.; Sikora, E.; Ufnal, M. TMA but not TMAO increases with age in rat plasma and affects smooth muscle cells viability. J. Gerontol. A Biol. Sci. Med. Sci. 2019. [Google Scholar] [CrossRef]
- Li, X.S.; Obeid, S.; Klingenberg, R.; Gencer, B.; Mach, F.; Raber, L.; Windecker, S.; Rodondi, N.; Nanchen, D.; Muller, O. Gut microbiota-dependent trimethylamine N-oxide in acute coronary syndromes: A prognostic marker for incident cardiovascular events beyond traditional risk factors. Eur. Heart J. 2017, 38, 814–824. [Google Scholar] [CrossRef]
- Qi, J.; You, T.; Li, J.; Pan, T.; Xiang, L.; Han, Y.; Zhu, L. Circulating trimethylamine N-oxide and the risk of cardiovascular diseases: A systematic review and meta-analysis of 11 prospective cohort studies. J. Cell. Mol. Med. 2018, 22, 185–194. [Google Scholar] [CrossRef]
- Senthong, V.; Wang, Z.; Fan, Y.; Wu, Y.; Hazen, S.L.; Tang, W.H. Trimethylamine N-Oxide and Mortality Risk in Patients With Peripheral Artery Disease. J. Am. Heart Assoc. 2016, 5, e004237. [Google Scholar] [CrossRef]
- Tang, W.H.; Wang, Z.; Levison, B.S.; Koeth, R.A.; Britt, E.B.; Fu, X.; Wu, Y.; Hazen, S.L. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N. Engl. J. Med. 2013, 368, 1575–1584. [Google Scholar] [CrossRef] [Green Version]
- Troseid, M.; Ueland, T.; Hov, J.R.; Svardal, A.; Gregersen, I.; Dahl, C.P.; Aakhus, S.; Gude, E.; Bjorndal, B.; Halvorsen, B. Microbiota-dependent metabolite trimethylamine-N-oxide is associated with disease severity and survival of patients with chronic heart failure. J. Intern. Med. 2015, 277, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Wang, Z.; Fan, Y.; Levison, B.; Hazen, J.E.; Donahue, L.M.; Wu, Y.; Hazen, S.L. Prognostic value of elevated levels of intestinal microbe-generated metabolite trimethylamine-N-oxide in patients with heart failure: Refining the gut hypothesis. J. Am. Coll. Cardiol. 2014, 64, 1908–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samulak, J.J.; Sawicka, A.K.; Samborowska, E.; Olek, R.A. Plasma trimethylamine-N-oxide following cessation of L-carnitine supplementation in healthy aged women. Nutrients 2019, 11, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querio, G.; Antoniotti, S.; Levi, R.; Gallo, M.P. Trimethylamine N-Oxide Does Not Impact Viability, ROS Production, and Mitochondrial Membrane Potential of Adult Rat Cardiomyocytes. Int. J. Mol. Sci. 2019, 20, 3045. [Google Scholar] [CrossRef] [Green Version]
- Jaworska, K.; Konop, M.; Bielinska, K.; Hutsch, T.; Dziekiewicz, M.; Banaszkiewicz, A.; Ufnal, M. Inflammatory bowel disease is associated with increased gut-to-blood penetration of short-chain fatty acids: A new, non-invasive marker of a functional intestinal lesion. Exp. Physiol. 2019, 104, 1226–1236. [Google Scholar] [CrossRef]
- Suzuki, T.; Heaney, L.M.; Bhandari, S.S.; Jones, D.J.; Ng, L.L. Trimethylamine N-oxide and prognosis in acute heart failure. Heart Fail. Clin. 2016, 102, 841–848. [Google Scholar]
- Suzuki, T.; Heaney, L.M.; Jones, D.J.; Ng, L.L. Trimethylamine N-oxide and Risk Stratification after Acute Myocardial Infarction. Clin. Chem. 2017, 63, 420–428. [Google Scholar] [CrossRef]
- Ufnal, M.; Zadlo, A.; Ostaszewski, R. TMAO: A small molecule of great expectations. Nutrition 2015, 31, 1317–1323. [Google Scholar] [CrossRef]
- Sandek, A.; Bauditz, J.; Swidsinski, A.; Buhner, S.; Weber-Eibel, J.; von Haehling, S.; Schroedl, W.; Karhausen, T.; Doehner, W.; Rauchhaus, M. Altered intestinal function in patients with chronic heart failure. J. Am. Coll. Cardiol. 2007, 50, 1561–1569. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, A.; Banan, A.; Fields, J.; Keshavarzian, A. Intestinal barrier: An interface between health and disease. J. Gastroenterol. Hepatol. 2003, 18, 479–497. [Google Scholar] [CrossRef]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight junction proteins and signaling pathways in cancer and inflammation: A functional crosstalk. Front. Physiol. 2019, 9, 1942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.; Ganapathy, A.S.; Suchanec, E.; Laidler, L.; Ma, T.; Nighot, P. Intestinal epithelial tight junction barrier regulation by autophagy-related protein atg6/beclin 1. Am. J. Physiol. Cell Physiol. 2019, 316, C753–C765. [Google Scholar] [CrossRef] [PubMed]
- Dhawan, P.; Singh, A.B.; Deane, N.G.; No, Y.; Shiou, S.-R.; Schmidt, C.; Neff, J.; Washington, M.K.; Beauchamp, R.D. Claudin-1 regulates cellular transformation and metastatic behavior in colon cancer. J. Clin. Investig. 2005, 115, 1765–1776. [Google Scholar] [CrossRef] [Green Version]
- Kinugasa, T.; Akagi, Y.; Yoshida, T.; Ryu, Y.; Shiratuchi, I.; Ishibashi, N.; Shirouzu, K. Increased claudin-1 protein expression contributes to tumorigenesis in ulcerative colitis-associated colorectal cancer. Anticancer Res. 2010, 30, 3181–3186. [Google Scholar] [PubMed]
- Prasad, S.; Mingrino, R.; Kaukinen, K.; Hayes, K.L.; Powell, R.M.; MacDonald, T.T.; Collins, J.E. Inflammatory processes have differential effects on claudins 2, 3 and 4 in colonic epithelial cells. Lab. Investig. 2005, 85, 1139–1162. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Gröne, J.; Weber, B.; Staub, E.; Heinze, M.; Klaman, I.; Pilarsky, C.; Hermann, K.; Castanos-Velez, E.; Röpcke, S.; Mann, B. Differential expression of genes encoding tight junction proteins in colorectal cancer: Frequent dysregulation of claudin-1,-8 and-12. Int. J. Colorectal Dis. 2007, 22, 651–659. [Google Scholar] [CrossRef]
- Tamura, A.; Kitano, Y.; Hata, M.; Katsuno, T.; Moriwaki, K.; Sasaki, H.; Hayashi, H.; Suzuki, Y.; Noda, T.; Furuse, M. Megaintestine in claudin-15–deficient mice. Gastroenterology 2008, 134, 523–534.e3. [Google Scholar] [CrossRef]
- Xing, T.; Benderman, L.J.; Sabu, S.; Parker, J.; Yang, J.; Lu, Q.; Ding, L.; Chen, Y.-H. Hepatology, Tight junction protein Claudin-7 is essential for intestinal epithelial stem cell self-renewal and differentiation. Cell. Mol. Gastroenterol. 2019, 9, 641–659. [Google Scholar]
- Xing, T.; Camacho Salazar, R.; Chen, Y.-H. Animal models for studying epithelial barriers in neonatal necrotizing enterocolitis, inflammatory bowel disease and colorectal cancer. Tissue Barriers 2017, 5, e1356901. [Google Scholar] [CrossRef]
- Kirschner, N.; Poetzl, C.; von den Driesch, P.; Wladykowski, E.; Moll, I.; Behne, M.J.; Brandner, J.M. Alteration of tight junction proteins is an early event in psoriasis: Putative involvement of proinflammatory cytokines. Am. J. Pathol. 2009, 175, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.-M.; Li, X.-M.; Qin, B.-z.; Liu, B. Effect of tight junction protein of intestinal epithelium and permeability of colonic mucosa in pathogenesis of injured colonic barrier during chronic recovery stage of rats with inflammatory bowel disease. Asian Pac. J. Trop. Med. 2016, 9, 148–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeissig, S.; Bürgel, N.; Günzel, D.; Richter, J.; Mankertz, J.; Wahnschaffe, U.; Kroesen, A.J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Changes in expression and distribution of claudin 2, 5 and 8 lead to discontinuous tight junctions and barrier dysfunction in active Crohn’s disease. Gut 2007, 56, 61–72. [Google Scholar] [CrossRef]
- Sandek, A.; Swidsinski, A.; Schroedl, W.; Watson, A.; Valentova, M.; Herrmann, R.; Scherbakov, N.; Cramer, L.; Rauchhaus, M.; Grosse-Herrenthey, A. Intestinal blood flow in patients with chronic heart failure: A link with bacterial growth, gastrointestinal symptoms, and cachexia. J. Am. Coll. Cardiol. 2014, 64, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, K.; Huc, T.; Samborowska, E.; Dobrowolski, L.; Bielinska, K.; Gawlak, M.; Ufnal, M. Hypertension in rats is associated with an increased permeability of the colon to TMA, a gut bacteria metabolite. PLoS ONE 2017, 12, e0189310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, M.R.; Savory, J. Biochemistry of renal failure. Ann. Clin. Lab. Sci. 1981, 11, 292–299. [Google Scholar]
- Mackay, R.J.; McEntyre, C.J.; Henderson, C.; Lever, M.; George, P.M. Trimethylaminuria: Causes and diagnosis of a socially distressing condition. Clin. Biochem. Rev. 2011, 32, 33. [Google Scholar]
- Tomasova, L.; Dobrowolski, L.; Jurkowska, H.; Wrobel, M.; Huc, T.; Ondrias, K.; Ostaszewski, R.; Ufnal, M. Intracolonic hydrogen sulfide lowers blood pressure in rats. Nitric Oxide: Biol. Chem. 2016, 60, 50–58. [Google Scholar] [CrossRef]
- Jaworska, K.; Huc, T.; Gawrys, M.; Onyszkiewicz, M.; Samborowska, E.; Ufnal, M. An In Vivo Method for Evaluating the Gut-Blood Barrier and Liver Metabolism of Microbiota Products. J. Vis. Exp. 2018, 58456. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group/ Parameter | WKY | SHHF |
|---|---|---|
| Energy and water balance | ||
| Body mass (g) | 427.4 ± 12.9 | 469.8 ± 25.1 |
| Tibia length (cm) | 3.58 ± 0.15 | 3.82 ± 0.12 |
| 24 h food intake (g) | 23.43 ± 1.24 | 21.94 ± 1.44 |
| 24 h water intake (mL) | 33.3 ± 1.8 | 37.6 ± 2.1 |
| 24 h stool output (g) | 9.5 ± 0.8 | 11.2 ± 1.8 |
| 24 h urine output (g) | 14.6 ± 2.1 | 15.8 ± 0.7 |
| Electrolyte balance | ||
| Plasma sodium (mM) | 137.45 ± 0.87 | 143.97 ± 1.3 * |
| Plasma potassium (mM) | 4.47 ± 0.13 | 4.78 ± 0.18 |
| Plasma creatinine (mM) | 0.07 ± 0.007 | 0.07 ± 0.002 |
| Hormones | ||
| Aldosterone (ng/mL) | 8.13 ± 1.01 | 8.16 ± 1.34 |
| Vasopressin (pg/mL) | 1.516.9 ± 132.1 | 3.222.4 ± 345.4 * |
| NT-proBNP (pg/mL) | 30.98 ± 2.70 | 47.60 ± 1.31 * |
| Heart mass | ||
| Heart mass | 1.07 ± 0.04 | 1.85 ± 0.16 * |
| Arterial blood pressure and heart rate | ||
| Systolic (mmHg) | 106.9 ± 4.9 | 109.4 ± 4.9 |
| Diastolic (mmHg) | 66.7 ± 4.1 | 80.1 ± 4.5 * |
| HR (beats/min) | 309 ± 22 | 312 ± 35 |
| Echocardiographic parameters | ||
| LV EDV (mL) | 0.57 ± 0.03 | 0.28 ± 0.04 * |
| LV ESV (mL) | 0.13 ± 0.03 | 0.11 ± 0.03 |
| IVSs (cm) | 0.25 ± 0.02 | 0.40 ± 0.01 * |
| IVSd (cm) | 0.20 ± 0.01 | 0.29 ± 0.02 * |
| LVPWs (cm) | 0.29 ± 0.02 | 0.40 ± 0.03 * |
| LVPWd (cm) | 0.20 ± 0.01 | 0.30 ± 0.02 * |
| SV (mL) | 0.44 ± 0.02 | 0.18 ± 0.03 * |
| EF (%) | 76.9 ± 3.5 | 65.0 ± 3.9 * |
| Left ventricle hemodynamic parameters (direct measurements) | ||
| LVEDP (mmHg) | 5.23 ± 0.63 | 4.73 ± 0.67 |
| dP/dt (mmHg/ms) | 7.02 ± 0.36 | 6.28 ± 0.08 |
| -dP/dt (mmHg/ms) | 3.53 ± 0.45 | 2.43 ± 0.10 |
| ECG | ||
| RR (ms) | 200.2 ± 16.6 | 207.7 ± 7.2 |
| QT (ms) | 50.0 ± 1.5 | 63.8 ± 3.9 * |
| QTc (ms) | 112.5 ± 3.4 | 141.4 ± 8.2 * |
| PR (ms) | 52.5 ± 2.2 | 57.9 ± 2.1 |
| QRS width (ms) | 20.6 ± 1.2 | 28.9 ± 2.1 * |
| QRS amplitude (mV) | 2.61 ± 0.40 | 1.62 ± 0.17 * |
| Intestinal blood flow | ||
| Intestinal blood flow (mL/min) | 8.56 ± 1.35 | 5.28 ± 0.75 * |
| Group/ Parameter | WKY | SHHF |
|---|---|---|
| Stool TMA (µM) | 375.3 ± 25.7 | 304.1 ± 20.8 |
| Stool TMAO (µM) | LQQ | LQQ |
| Plasma TMA (µM) | 97.42 ± 6.7 | 153.0 ± 20.3 * |
| Plasma TMAO (µM) | 4.57 ± 0.39 | 6.67 ± 0.69 * |
| Liver TMA oxidation (plasma TMAO/TMA ratio) | 0.049 ± 0.005 | 0.048 ± 0.006 |
| Gut permeability (plasma TMA+TMAO/stool TMA) | 0.28 ± 0.03 | 0.54 ± 0.08 * |
| Urine TMA (µM) | 594.20 ± 32.73 | 731.41 ± 98.99 * |
| Urine TMAO (µM) | 432.63 ± 53.61 | 667.61 ± 64.13 * |
| 24 h urine output TMA + TMAO (µmol/24 h) | 16.07 ± 1.98 | 21.09 ± 1.55 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drapala, A.; Szudzik, M.; Chabowski, D.; Mogilnicka, I.; Jaworska, K.; Kraszewska, K.; Samborowska, E.; Ufnal, M. Heart Failure Disturbs Gut–Blood Barrier and Increases Plasma Trimethylamine, a Toxic Bacterial Metabolite. Int. J. Mol. Sci. 2020, 21, 6161. https://doi.org/10.3390/ijms21176161
Drapala A, Szudzik M, Chabowski D, Mogilnicka I, Jaworska K, Kraszewska K, Samborowska E, Ufnal M. Heart Failure Disturbs Gut–Blood Barrier and Increases Plasma Trimethylamine, a Toxic Bacterial Metabolite. International Journal of Molecular Sciences. 2020; 21(17):6161. https://doi.org/10.3390/ijms21176161
Chicago/Turabian StyleDrapala, Adrian, Mateusz Szudzik, Dawid Chabowski, Izabella Mogilnicka, Kinga Jaworska, Katarzyna Kraszewska, Emilia Samborowska, and Marcin Ufnal. 2020. "Heart Failure Disturbs Gut–Blood Barrier and Increases Plasma Trimethylamine, a Toxic Bacterial Metabolite" International Journal of Molecular Sciences 21, no. 17: 6161. https://doi.org/10.3390/ijms21176161