Characterization of the Bacteriophage vB_EfaS-271 Infecting Enterococcus faecalis

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
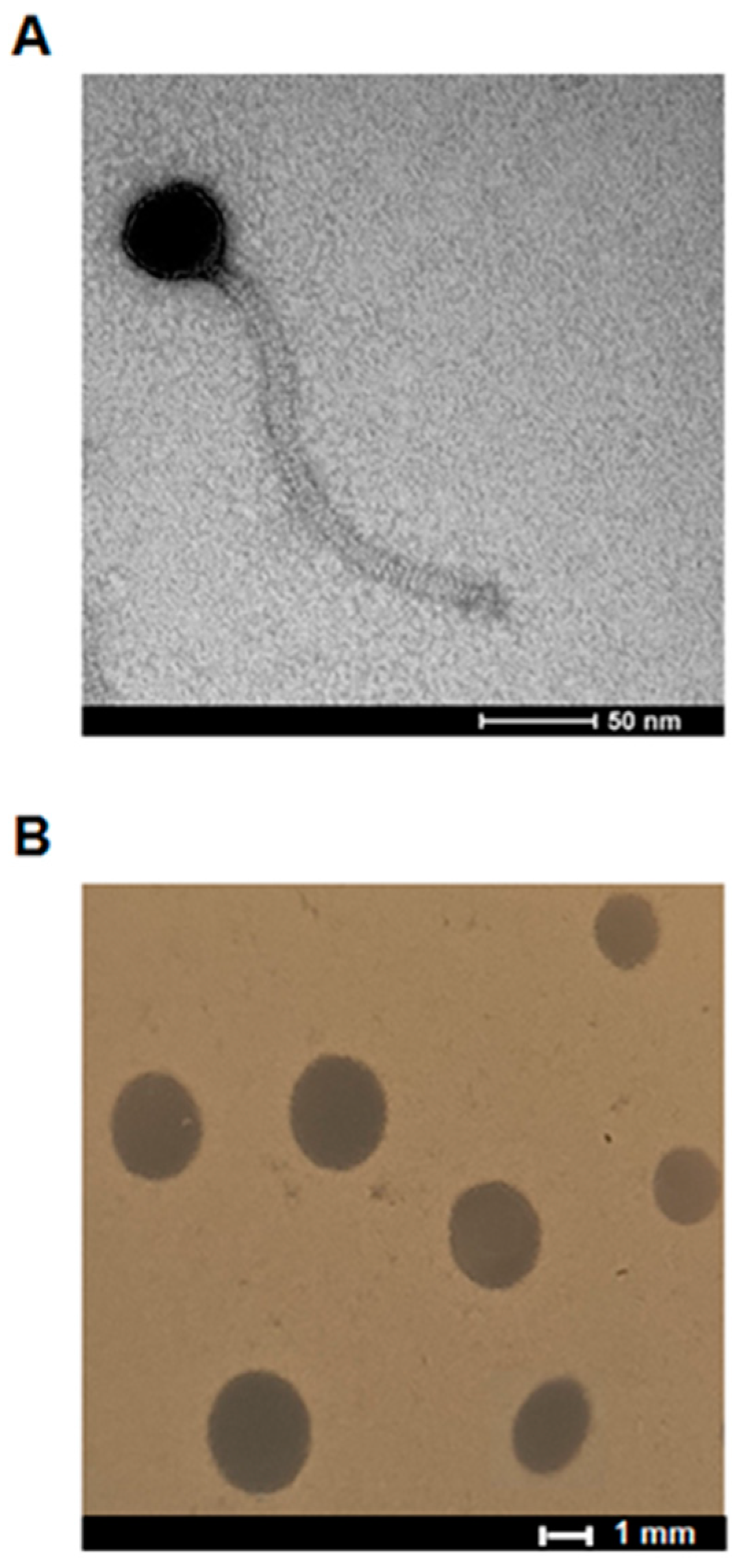
2.1. Virion and Plaque Morphology
2.2. Host Range
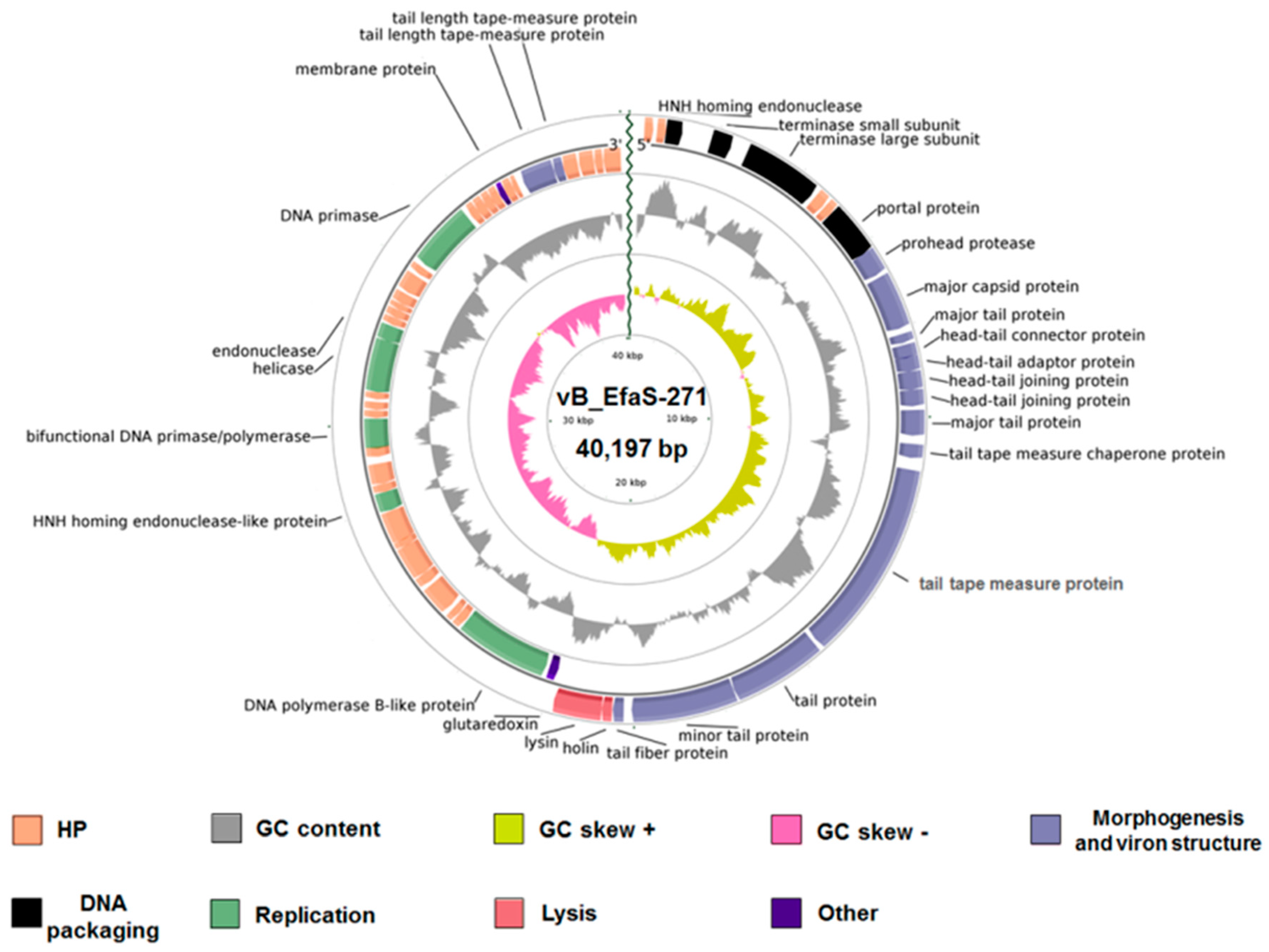
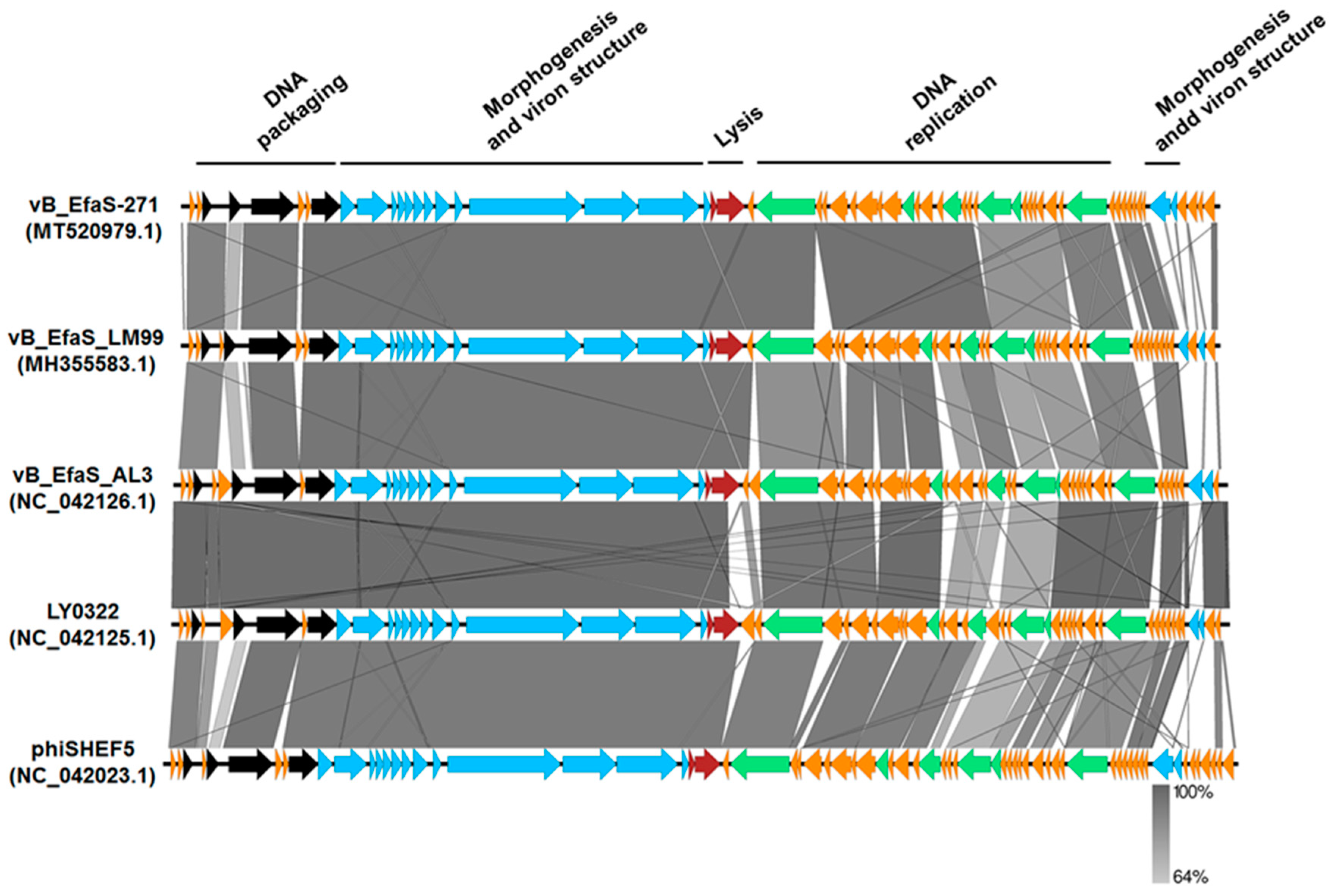
2.3. Phage Genome Analysis
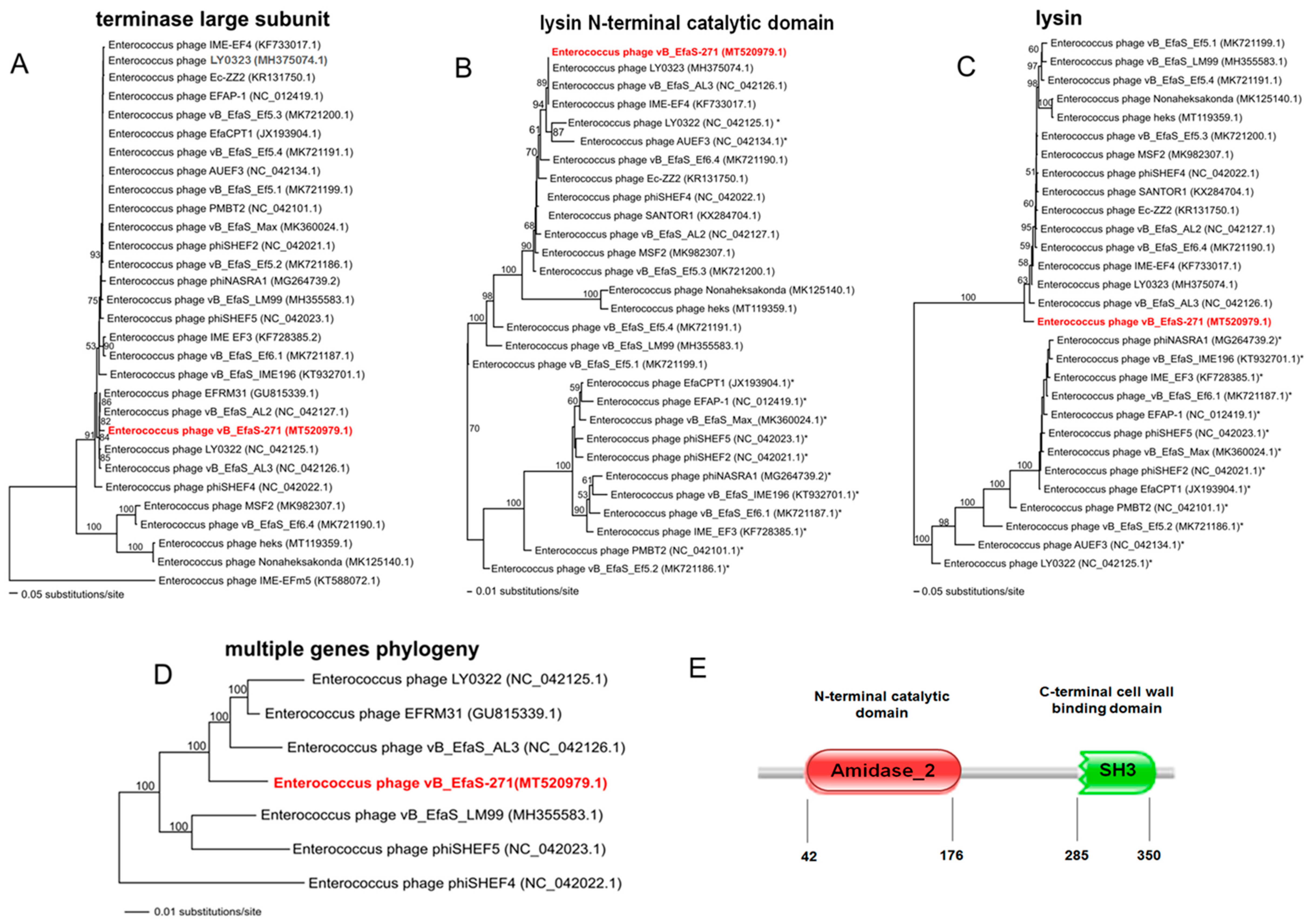
2.4. Phage Protein Analysis
2.5. Sensitivity of Virions to External Conditions
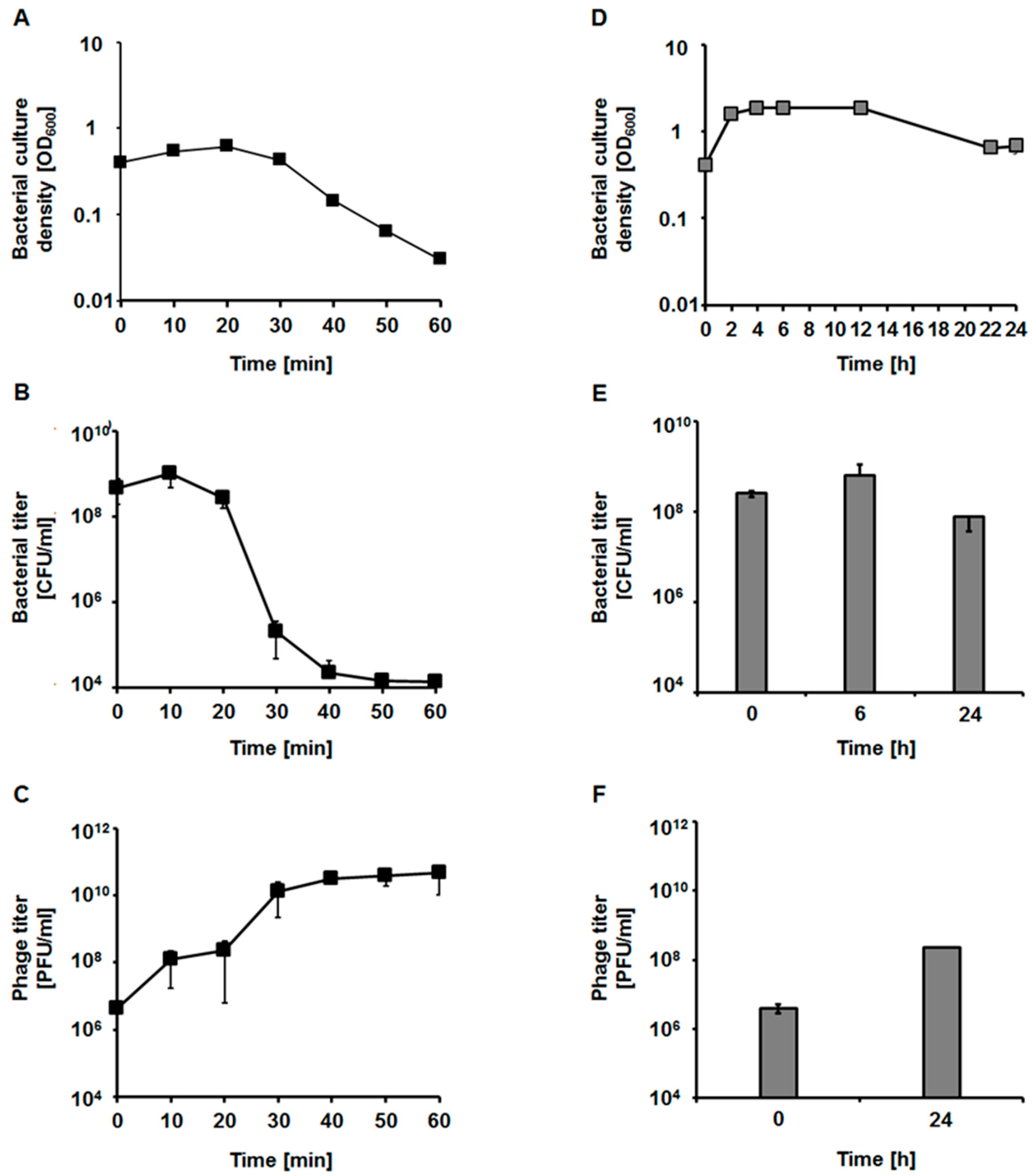
2.6. Bacteriophage Development
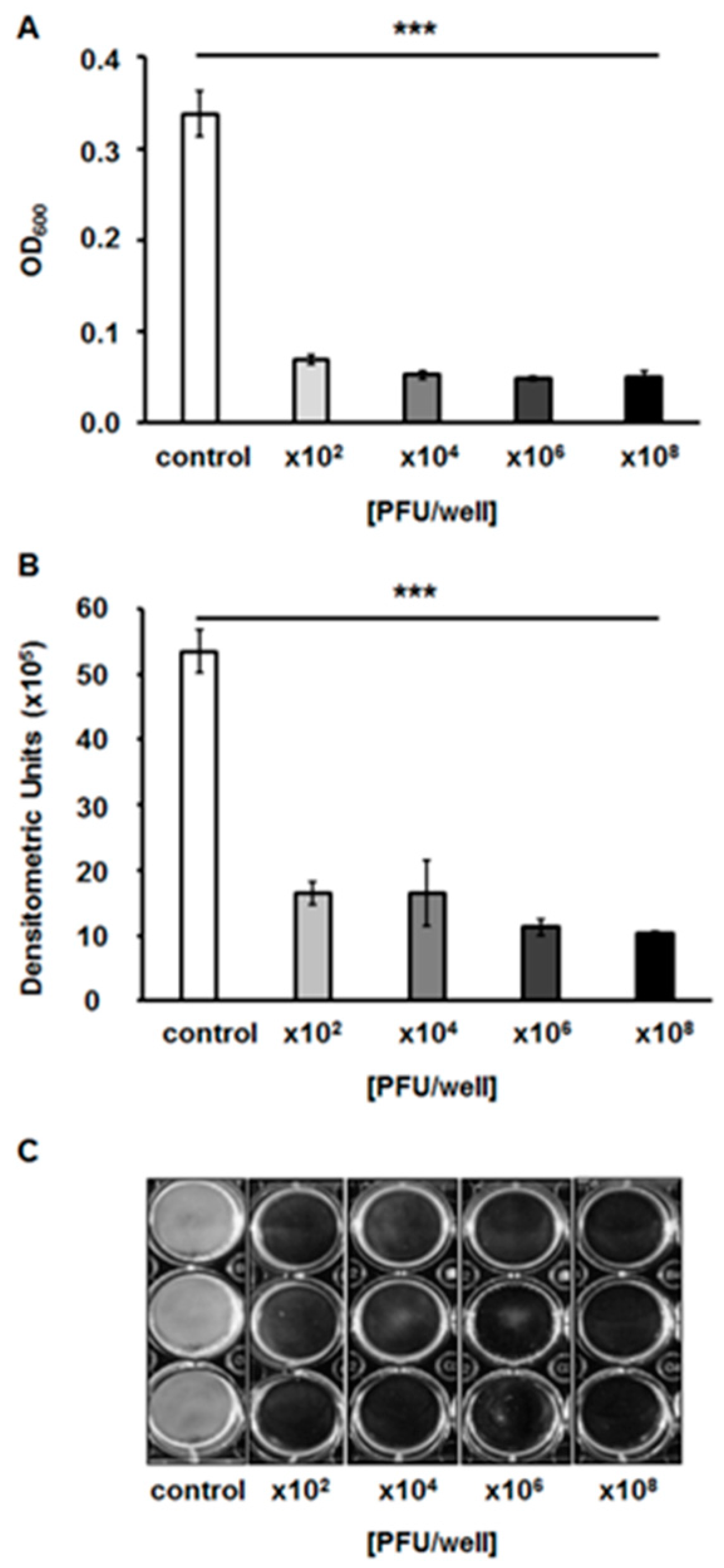
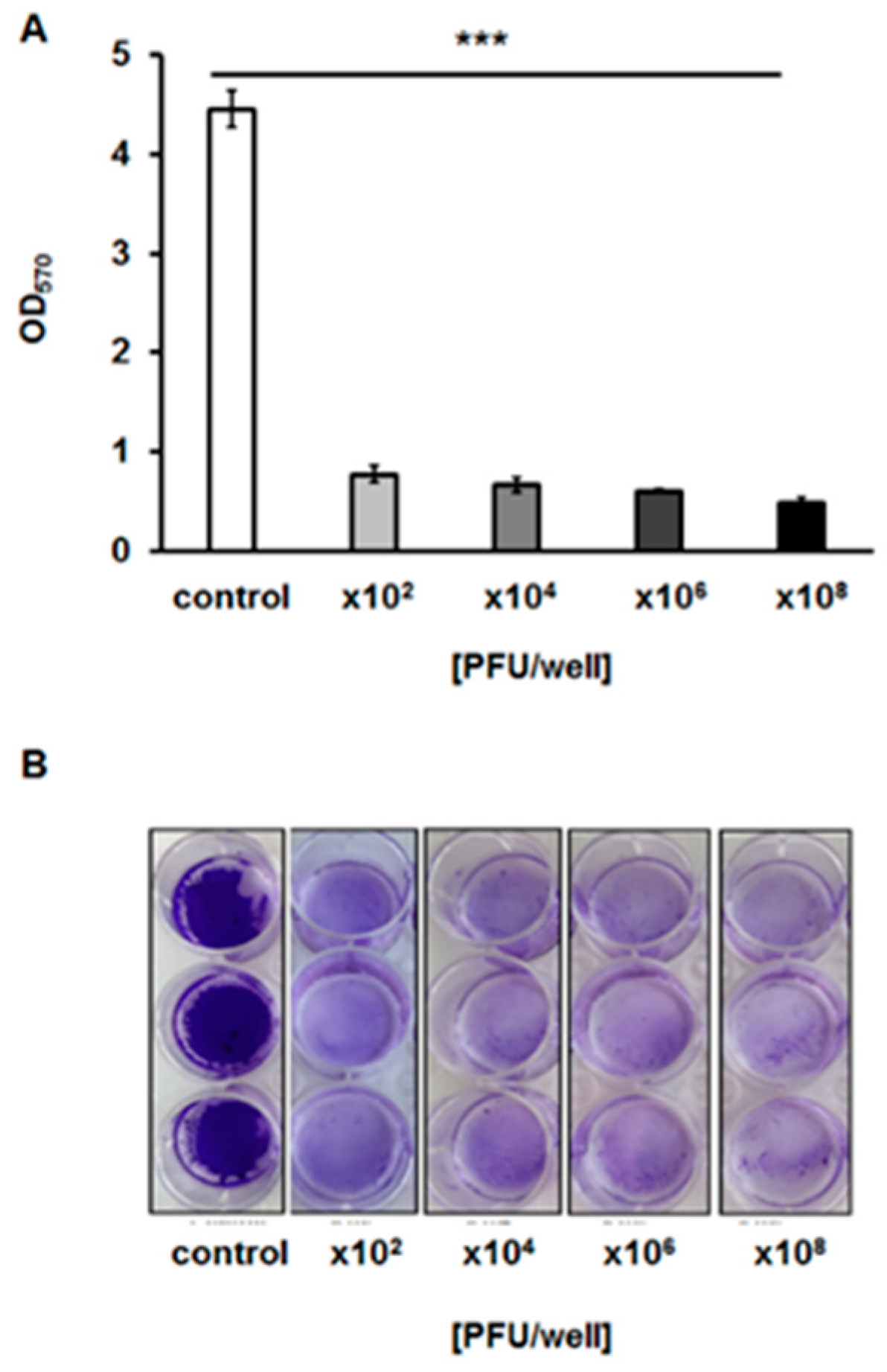
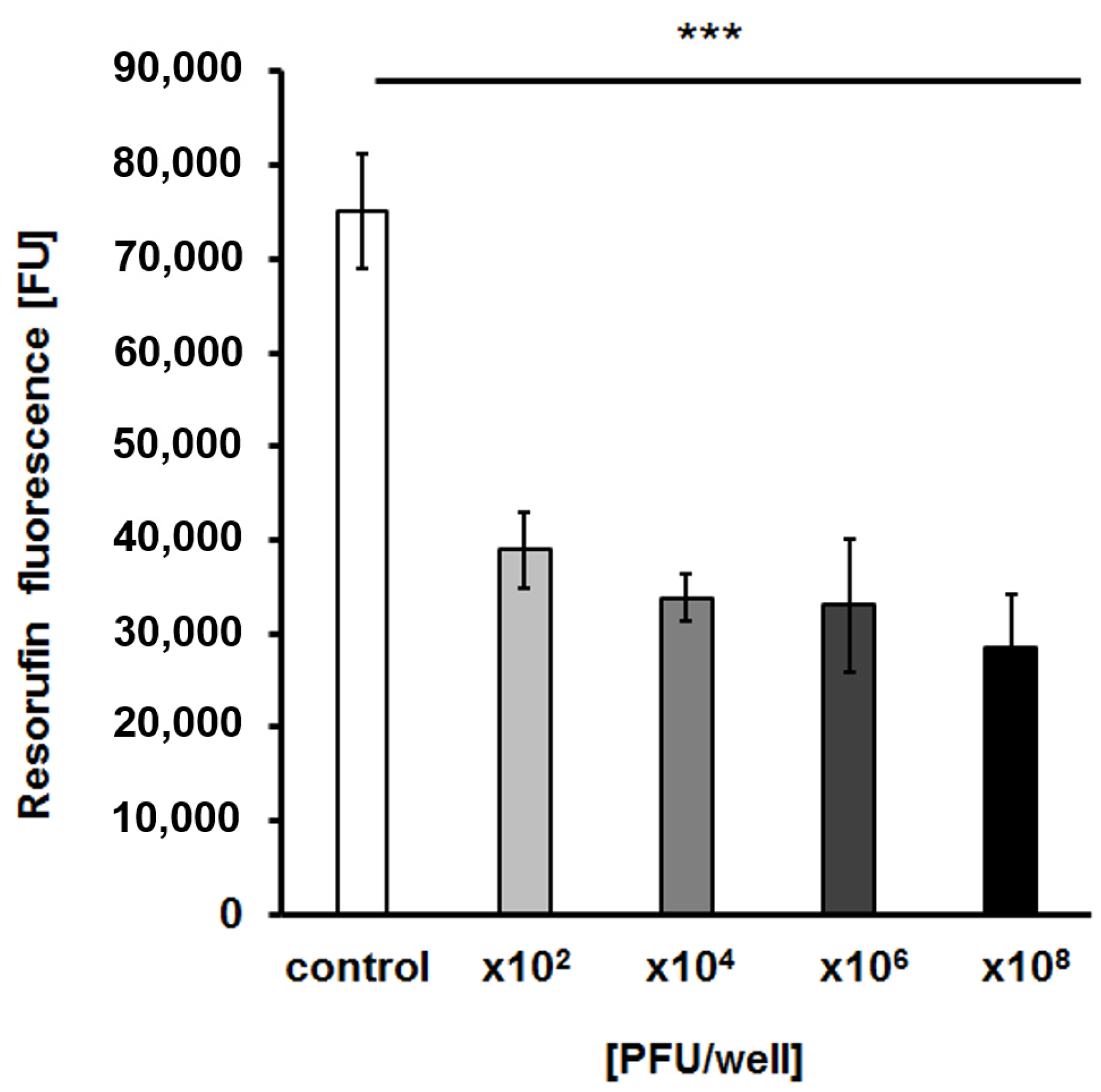
2.7. Effects of the Bacteriophage on Bacterial Biofilm
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Isolation of the vB_EfaS-271 Bacteriophage from Urban Sewage
4.3. Preparation of the vB_EfaS-271 Lysate
4.4. Determination of the Bacteriophage vB_EfaS-271 Host Range
4.5. Electron Microscopic Examination of Phage vB_EfaS-271 Morphology
4.6. Examination of the Morphology of vB_EfaS-271 Plaques
4.7. The Effects of External Factors on the Stability of vB_EfaS-271 Virions
4.8. vB_EfaS-271 Phage Adsorption Rate Assay
4.9. One-Step Growth Experiment
4.10. Lysis Profile of the Bacterial Hosts after vB_EfaS-271 Infection
4.11. Assessment of vB_EfaS-271 Phage Lytic Activity in Biofilms
4.12. Crystal Violet (CV) Staining for the Quantification of Biofilm Biomass
4.13. Detection of the Metabolic Activity of Biofilm Cells after vB_EfaS-271 Infection
4.14. Extraction, Sequencing and Bioinformatic Analysis of the vB_EfaS-271 Genome
4.15. Phylogenetic Analysis
4.16. Mass Spectrometry Analysis of vB_EfaS-271 Proteins
4.17. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BCA | Bicinchonic acid |
| BRIG | BLAST ring image generator |
| CFU | Cetyltrimethylammonium bromide |
| CTAB | Crystal violet |
| CV | Dimethyl sulfoxide |
| DNase I | Deoxyribonuclease |
| DTT | Dithiothreitol |
| EHEC | Enterohemorrhagic Escherichia coli |
| EM | Emmision |
| EPEC | Enteropathogenic Escherichia coli |
| ESP | Eschrichia coli secretion protein |
| EXC | Excitation |
| FDR | False discovery rate |
| FU | Fluorescence units |
| HPLC | High-performance liquid chromatography |
| LA | Luria-Bertani agar |
| LB | Luria-Bertani broth |
| LIN | Linezolid |
| MS | Mass spectrometry |
| M.O.I. | Multiplicity of infection |
| NCBI | National center for biotechnology information |
| NGS | Next generation sequencing |
| OD | Optical density |
| ORF | Open reading frame |
| PBS | Phosphate-buffered saline |
| PFU | Plaque forming units |
| RNaseA | Ribonuclease A |
| SD | Standard deviation |
| Stx | Shiga toxin |
| TEM | Transmission electron microscopy |
| TFA | Trifluoroacetic acid |
| TerL | Large subunit of terminase |
| TM | Tris-Magnesium sulfate buffer |
| TSA | Tryptic soy agar |
| TSB | Tryptic soy broth |
| VFDB | Virulence Factors of the Pathogenic Bacteria database |
References
- Comeau, A.M.; Hatfull, G.F.; Krisch, H.M.; Lindell, D.; Mann, N.H.; Prangishvili, D. Exploring the prokaryotic virosphere. Res. Microbiol. 2008, 159, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Keen, E.C. A century of phage research: Bacteriophages and the shaping of modern biology. BioEssays 2015, 37, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R.; Hendrix, R.W. Bacteriophage lambda: Early pioneer and still relevant. Virology 2015, 479, 310–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Genet. 2020, 18, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Hampton, H.G.; Watson, B.N.J.; Fineran, P.C. The arms race between bacteria and their phage foes. Nature 2020, 577, 327–336. [Google Scholar] [CrossRef]
- Górski, A.; Międzybrodzki, R.; Węgrzyn, G.; Jończyk-Matysiak, E.; Borysowski, J.; Weber-Dąbrowska, B. Phage therapy: Current status and perspectives. Med. Res. Rev. 2019, 40, 459–463. [Google Scholar] [CrossRef]
- Boyd, E.F. Bacteriophage-encoded bacterial virulence factors and phage–pathogenicity island interactions. Adv. Appl. Microbiol. 2012, 82, 91–118. [Google Scholar] [CrossRef]
- Jones, D.T.; Shirley, M.; Wu, X.; Keis, S. Bacteriophage infections in the industrial acetone butanol (AB) fermentation process. J. Mol. Microbiol. Biotechnol. 2000, 2, 21–26. [Google Scholar]
- Czyz, A.; Los, M.; Wróbel, B.; Węgrzyn, G. Inhibition of spontaneous induction of lambdoid prophages in Escherichia coli cultures: Simple procedures with possible biotechnological applications. BMC Biotechnol. 2001, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Gabig-Cimińska, M.; Los, M.; Holmgren, A.; Albers, J.; Czyż, A.; Hintsche, R.; Węgrzyn, G.; Enfors, S.-O. Detection of bacteriophage infection and prophage induction in bacterial cultures by means of electric DNA chips. Anal. Biochem. 2004, 324, 84–91. [Google Scholar] [CrossRef]
- Jurczak-Kurek, A.; Gąsior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of bacteriophages: Morphological and biological properties of a large group of phages isolated from urban sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef]
- Brandl, K.; Plitas, G.; Mihu, C.N.; Ubeda, C.; Jia, T.; Fleisher, M.; Schnabl, B.; DeMatteo, R.P.; Pamer, E.G. Vancomycin-resistant enterococci exploit antibiotic-induced innate immune deficits. Nature 2008, 455, 804–807. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.E.V.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The inner of the two Muc2 mucin-dependent mucus layers in colon is devoid of bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Garbajosa, P.; Regt, M.; Bonten, M.; Baquero, F.; Coque, T.M.; Canton, R.; Harmsen, H.J.M.; Willems, R.J. High-density fecal Enterococcus faecium colonization in hospitalized patients is associated with the presence of the polyclonal subcluster CC17. Eur. J. Clin. Microbiol. Infect. Dis. 2011, 31, 519–522. [Google Scholar] [CrossRef]
- Tedim, A.P.; Ruiz-Garbajosa, P.; Corander, J.; Rodríguez, C.M.; Cantón, R.; Willems, R.J.; Baquero, F.; Coque, T.M. Population biology of intestinal Enterococcus isolates from hospitalized and nonhospitalized individuals in different age groups. Appl. Environ. Microbiol. 2015, 81, 1820–1831. [Google Scholar] [CrossRef] [Green Version]
- Ubeda, C.; Taur, Y.; Jenq, R.R.; Equinda, M.J.; Son, T.; Samstein, M.; Viale, A.; Socci, N.D.; Brink, M.R.V.D.; Kamboj, M.; et al. Vancomycin-resistant Enterococcus domination of intestinal microbiota is enabled by antibiotic treatment in mice and precedes bloodstream invasion in humans. J. Clin. Investig. 2010, 120, 4332–4341. [Google Scholar] [CrossRef]
- Ho, C.; Lau, A.; Cimon, K.; Farrah, K.; Gardam, M. Screening, isolation, and decolonization strategies for vancomycin-eesistant enterococci or extended spectrum beta-lactamase producing organisms: A systematic review of the clinical evidence and health services impact. CADTH Technol. Overv. 2012, 3, 3202. [Google Scholar]
- Shiadeh, S.M.J.; Pormohammad, A.; Hashemi, A.; Lak, P. Global prevalence of antibiotic resistance in blood-isolated Enterococcus faecalis and Enterococcus faecium: A systematic review and meta-analysis. Infect. Drug Resist. 2019, 12, 2713–2725. [Google Scholar] [CrossRef] [Green Version]
- Olawale, K.; Fadiora, S.; Taiwo, S. Prevalence of hospital acquired enterococci infections in two primary-care hospitals in Osogbo, Southwestern Nigeria. Afr. J. Infect. Dis. 2011, 5, 2. [Google Scholar] [CrossRef]
- McBride, S.J.; Upton, A.; Roberts, S.A. Clinical characteristics and outcomes of patients with vancomycin-susceptible Enterococcus faecalis and Enterococcus faecium bacteraemia—A five-year retrospective review. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 29, 107–114. [Google Scholar] [CrossRef]
- Vergis, E.N.; Hayden, M.K.; Chow, J.W.; Snydman, D.R.; Zervos, M.J.; Linden, P.K.; Wagener, M.M.; Schmitt, B.; Muder, R.R. Determinants of vancomycin resistance and mortality rates in enterococcal bacteremia. a prospective multicenter study. Ann. Intern. Med. 2001, 135, 484–492. [Google Scholar] [CrossRef]
- Deshpande, L.M.; Fritsche, T.R.; Moet, G.J.; Biedenbach, D.J.; Jones, R.N. Antimicrobial resistance and molecular epidemiology of vancomycin-resistant enterococci from North America and Europe: A report from the SENTRY antimicrobial surveillance program. Diagn. Microbiol. Infect. Dis. 2007, 58, 163–170. [Google Scholar] [CrossRef]
- Hidron, A.I.; Edwards, J.R.; Patel, J.; Horan, T.C.; Sievert, D.M.; Pollock, D.A.; Fridkin, S.K.; National Healthcare Safety Network Team; Participating National Healthcare Safety Network Facilities. NHSN Annual Update: Antimicrobial-Resistant Pathogens Associated with Healthcare-Associated Infections: Annual Summary of Data Reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2006–2007. Infect. Control. Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef] [Green Version]
- Reik, R.; Tenover, F.C.; Klein, E.Y.; McDonald, L.C. The burden of vancomycin-resistant enterococcal infections in US hospitals, 2003 to 2004. Diagn. Microbiol. Infect. Dis. 2008, 62, 81–85. [Google Scholar] [CrossRef]
- Wisplinghoff, H.; Bischoff, T.; Tallent, S.M.; Seifert, H.; Wenzel, R.; Edmond, M.B. Nosocomial bloodstream infections in US hospitals: Analysis of 24,179 cases from a prospective nationwide surveillance study. Clin. Infect. Dis. 2004, 39, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Van Harten, R.M.; Willems, R.J.; Martin, N.I.; Hendrickx, A.P. Multidrug-resistant enterococcal infections: New compounds, novel antimicrobial therapies? Trends Microbiol. 2017, 25, 467–479. [Google Scholar] [CrossRef]
- Chlebicki, M.P.; Kurup, A. Vancomycin-resistant enterococcus: A review from a Singapore perspective. Ann. Acad. Med. Singap. 2008, 37, 861–869. [Google Scholar]
- Raza, T.; Ullah, S.R.; Mehmood, K.; Andleeb, S. Vancomycin resistant Enterococci: A brief review. J. Pak. Med. Assoc. 2018, 68, 768–772. [Google Scholar]
- Tan, C.A.Z.; Antypas, H.; Kline, K.A. Overcoming the challenge of establishing biofilms in vivo: A roadmap for Enterococci. Curr. Opin. Microbiol. 2020, 53, 9–18. [Google Scholar] [CrossRef]
- Górski, A.; Międzybrodzki, R.; Lobocka, M.; Głowacka-Rutkowska, A.; Bednarek, A.; Borysowski, J.; Jończyk-Matysiak, E.; Łusiak-Szelachowska, M.; Weber-Dąbrowska, B.; Bagińska, N.; et al. Phage therapy: What have we learned? Viruses 2018, 10, 288. [Google Scholar] [CrossRef] [Green Version]
- Ladero, V.; Gómez-Sordo, C.; Sanchez-Llana, E.; Del Rio, B.; Redruello, B.; Fernández, M.; Martín, M.C.; Alvarez, M.A. Q69 (an E. faecalis-infecting bacteriophage) as a biocontrol agent for reducing tyramine in dairy products. Front. Microbiol. 2016, 7, 445. [Google Scholar] [CrossRef]
- Del Rio, B.; Sánchez-Llana, E.; Redruello, B.; Magadan, A.; Fernández, M.; Martin, M.C.; Ladero, V.; Alvarez, M.A. Enterococcus faecalis bacteriophage 156 is an effective biotechnological tool for reducing the presence of tyramine and putrescine in an experimental cheese model. Front. Microbiol. 2019, 10, 566. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Upadrasta, A.; Bettio, P.H.D.A.; Clooney, A.G.; Draper, L.A.; Ross, R.P.; Hill, C. Evaluation of phage therapy in the context of Enterococcus faecalis and its associated diseases. Viruses 2019, 11, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.; Xing, S.; Zhang, X.; Pei, G.; An, X.; Mi, Z.; Huang, Y.; Tong, Y.-G. Complete genome sequence of a new Enterococcus faecalis bacteriophage, vB_EfaS_IME197. Genome Announc. 2016, 4, 00827-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, R.H.; Zhang, H.; Hsiao, C.; Kachlany, S.; Tinoco, E.M.B.; DePew, J.; Fouts, D.E. Structural proteins of Enterococcus faecalis bacteriophage ϕEf11. Bacteriophage 2016, 6, e1251381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Rodriguez, T.M.; Ly, M.; Pride, D.T.; Toranzos, G.A. Draft genome sequence of an Enterococcus faecalis ATCC 19433 Siphovirus isolated from raw domestic sewage. Genome Announc. 2017, 5, e01179-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, S.R.; Andleeb, S.; Raza, T.; Jamal, M.; Mehmood, K. Effectiveness of a lytic phage SRG1 against vancomycin-resistant Enterococcus faecalis in compost and soil. BioMed Res. Int. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Khalifa, L.; Gelman, D.; Shlezinger, M.; Dessal, A.L.; Coppenhagen-Glazer, S.; Beyth, N.; Hazan, R. Defeating antibiotic- and phage-resistant Enterococcus faecalis using a phage cocktail in vitro and in a clot model. Front. Microbiol. 2018, 9, 326. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Xing, S.; Zhao, F.; Li, P.; Mi, Z.; Shi, T.-X.; Liu, H.; Tong, Y.-G. Characterization and genome analysis of novel phage vB_EfaP_IME195 infecting Enterococcus faecalis. Virus Genes 2018, 54, 804–811. [Google Scholar] [CrossRef]
- Melo, L.D.R.; Ferreira, R.; Costa, A.R.; Oliveira, H.; Azeredo, J. Efficacy and safety assessment of two enterococci phages in an in vitro biofilm wound model. Sci. Rep. 2019, 9, 6643. [Google Scholar] [CrossRef] [Green Version]
- Barros, J.; Melo, L.D.R.; Poeta, P.; Igrejas, G.; Ferraz, M.P.; Azeredo, J.; Monteiro, F.J. Lytic bacteriophages against multidrug-resistant Staphylococcus aureus, Enterococcus faecalis and Escherichia coli isolates from orthopaedic implant-associated infections. Int. J. Antimicrob. Agents 2019, 54, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Zhao, F.; Wang, L.; Tan, D.; Cong, C.; Li, X.; Xu, Y. Complete genome analysis of the novel Enterococcus faecalis phage vB_EfaS_AL3. Arch. Virol. 2019, 164, 2599–2603. [Google Scholar] [CrossRef] [PubMed]
- Al-Zubidi, M.; Widziolek, M.; Court, E.K.; Gains, A.F.; Smith, R.E.; Ansbro, K.; Alrafaie, A.; Evans, C.; Murdoch, C.; Mesnage, S.; et al. Identification of novel bacteriophages with therapeutic potential that target Enterococcus faecalis. Infect. Immun. 2019, 87, e00512-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Andrea, M.M.; Frezza, D.; Romano, E.; Marmo, P.; De Angelis, L.H.; Perini, N.; Thaller, M.; Di Lallo, G. The lytic bacteriophage vB_EfaH_EF1TV, a new member of the Herelleviridae family, disrupts biofilm produced by Enterococcus faecalis clinical strains. J. Glob. Antimicrob. Resist. 2020, 21, 68–75. [Google Scholar] [CrossRef]
- Bhardwaj, S.B.; Mehta, M.; Sood, S.; Sharma, J. Isolation of a novel phage and targeting biofilms of drug-resistant oral enterococci. J. Glob. Infect. Dis. 2020, 12, 11–15. [Google Scholar] [CrossRef]
- Tinoco, J.M.; Buttaro, B.; Zhang, H.; Liss, N.; Sassone, L.M.; Stevens, R.H. Effect of a genetically engineered bacteriophage on Enterococcus faecalis biofilms. Arch. Oral Boil. 2016, 71, 80–86. [Google Scholar] [CrossRef] [Green Version]
- Tinoco, J.M.; Liss, N.; Zhang, H.; Nissan, R.; Gordon, W.; Tinoco, E.M.B.; Sassone, L.M.; Stevens, R.H. Antibacterial effect of genetically-engineered bacteriophage ϕEf11/ϕFL1C(Δ36)P nisA on dentin infected with antibiotic-resistant Enterococcus faecalis. Arch. Oral Boil. 2017, 82, 166–170. [Google Scholar] [CrossRef]
- Cheng, M.; Liang, J.; Zhang, Y.; Hu, L.; Gong, P.; Cai, R.; Zhang, L.; Zhang, H.; Ge, J.; Ji, Y.; et al. The bacteriophage EF-P29 efficiently protects against lethal vancomycin-resistant Enterococcus faecalis and alleviates gut microbiota imbalance in a murine bacteremia model. Front. Microbiol. 2017, 8, 837. [Google Scholar] [CrossRef]
- Gelman, D.; Beyth, S.; Lerer, V.; Adler, K.; Poradosu-Cohen, R.; Coppenhagen-Glazer, S.; Hazan, R.; Lerrer, V. Combined bacteriophages and antibiotics as an efficient therapy against VRE Enterococcus faecalis in a mouse model. Res. Microbiol. 2018, 169, 531–539. [Google Scholar] [CrossRef]
- Shlezinger, M.; Friedman, M.; Houri-Haddad, Y.; Hazan, R.; Beyth, N. Phages in a thermoreversible sustained-release formulation targeting E. faecalis in vitro and in vivo. PLoS ONE 2019, 14, e0219599. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, T.; Ishida, W.; Fukuda, K.; Nakajima, I.; Suzuki, T.; Uchiyama, J.; Matsuzaki, S.; Todokoro, D.; Daibata, M.; Fukushima, A. Therapeutic effects of intravitreously administered bacteriophage in a mouse model of endophthalmitis caused by vancomycin-sensitive or -resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 2019, 63, 01088-19. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Llorente, C.; Lang, S.; Brandl, K.; Chu, H.; Jiang, L.; White, R.C.; Clarke, T.H.; Nguyen, K.; Torralba, M.; et al. Bacteriophage targeting of gut bacterium attenuates alcoholic liver disease. Nature 2019, 575, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Çolakoğlu, M.; Xue, J.; Trajkovski, M. Bacteriophage prevents alcoholic liver disease. Cell 2020, 180, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Swift, S.; Rowley, D.T.; Young, C.; Hyman, P.; Donovan, D.M.; Franks, A.; Fitzgerald, R. The endolysin from the Enterococcus faecalis bacteriophage VD13 and conditions stimulating its lytic activity. FEMS Microbiol. Lett. 2016, 363, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, M.; Zhang, Y.; Li, X.; Liang, J.; Hu, L.; Gong, P.; Zhang, L.; Cai, R.; Zhang, H.; Ge, J.; et al. Endolysin LysEF-P10 shows potential as an alternative treatment strategy for multidrug-resistant Enterococcus faecalis infections. Sci. Rep. 2017, 7, 10164. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.; Xi, H.; Cheng, M.; Han, W. Phage-derived lysins as therapeutic agents against multidrug-resistant Enterococcus faecalis. Futur. Microbiol. 2018, 13, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yang, H.; He, J.; Wang, S.; Li, Y.; Wei, H. Effects of a chimeric lysin against planktonic and sessile Enterococcus faecalis hint at potential application in endodontic therapy. Viruses 2018, 10, 290. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Buttaro, B.A.; Fouts, D.E.; Sanjari, S.; Evans, B.S.; Stevens, R.H. Bacteriophage φEf11 ORF28 endolysin, a multifunctional lytic enzyme with properties distinct from all other identified Enterococcus faecalis phage endolysins. Appl. Environ. Microbiol. 2019, 85, e00555-19. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Zhen, X.; Zhou, H.; Zhao, F.; Fan, C.; Perčulija, V.; Tong, Y.; Mi, Z.; Liu, Z.-J. Structural and functional insights into a novel two-component endolysin encoded by a single gene in Enterococcus faecalis phage. PLoS Pathog. 2020, 16, e1008394. [Google Scholar] [CrossRef]
- Yoong, P.; Schuch, R.; Nelson, D.; Fischetti, V.A. Identification of a broadly active phage lytic enzyme with lethal activity against antibiotic-resistant Enterococcus faecalis and Enterococcus faecium. J. Bacteriol. 2004, 186, 4808–4812. [Google Scholar] [CrossRef] [Green Version]
- Sonnhammer, E.L.L.; Eddy, S.R.; Durbin, R. Pfam: A comprehensive database of protein families based on seed alignments. Proteins 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Azadani, D.N.; Zhang, D.; Hatherill, J.R.; Silva, D.; Turner, J.W. Isolation, characterization, and comparative genomic analysis of a phage infecting high-level aminoglycoside-resistant (HLAR) Enterococcus faecalis. PeerJ 2020, 8, e9171. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Li, W.; Song, F.; Yang, X.; Zhou, J.; Yu, H.; Ji, X.; Wei, Y. Biological characteristics and whole-genome analysis of the Enterococcus faecalis phage PEf771. Can. J. Microbiol. 2020, 6, 1–16. [Google Scholar] [CrossRef]
- Chatterjee, A.; Willett, J.L.E.; Nguyen, U.T.; Monogue, B.; Palmer, K.L.; Dunny, G.M.; Duerkop, B.A. Parallel genomics uncover novel enterococcal-bacteriophage interactions. mBio 2020, 11, 03120-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Im, J.; Na, H.; Ryu, S.; Yun, C.-H.; Han, S.H. The Novel Enterococcus Phage vB_EfaS_HEf13 Has broad lytic activity against clinical isolates of Enterococcus faecalis. Front. Microbiol. 2019, 10, 2877. [Google Scholar] [CrossRef]
- Nowicki, D.; Maciąg-Dorszyńska, M.; Bogucka, K.; Szalewska-Pałasz, A.; Herman-Antosiewicz, A. Various modes of action of dietary phytochemicals, sulforaphane and phenethyl isothiocyanate, on pathogenic bacteria. Sci. Rep. 2019, 9, 13677. [Google Scholar] [CrossRef]
- Łuczkiewicz, A.; Felis, E.; Ziembińska, A.; Gnida, A.; Kotlarska, E.; Olanczuk-Neyman, K.; Surmacz-Górska, J. Resistance of Escherichia coli and Enterococcus spp. to selected antimicrobial agents present in municipal wastewater. J. Water Heal. 2013, 11, 600–612. [Google Scholar] [CrossRef]
- Sadowy, E.; Łuczkiewicz, A. Drug-resistant and hospital-associated Enterococcus faecium from wastewater, riverine estuary and anthropogenically impacted marine catchment basin. BMC Microbiol. 2014, 14, 66. [Google Scholar] [CrossRef] [Green Version]
- Murray, B.E.; Singh, K.V.; Ross, R.P.; Heath, J.D.; Dunny, G.M.; Weinstock, G. Generation of restriction map of Enterococcus faecalis OG1 and investigation of growth requirements and regions encoding biosynthetic function. J. Bacteriol. 1993, 175, 5216–5223. [Google Scholar] [CrossRef] [Green Version]
- Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Penicillin-resistant, ampicillin-susceptible Enterococcus faecalis in Polish hospitals. Microb. Drug Resist. 2020. [Google Scholar] [CrossRef]
- Kawalec, M.; Gniadkowski, M.; Zielińska, U.; Kłos, W.; Hryniewicz, W. Vancomycin-resistant Enterococcus faecium strain aarrying the vanB2 gene variant in a Polish hospital. J. Clin. Microbiol. 2001, 39, 811–815. [Google Scholar] [CrossRef] [Green Version]
- Sahm, D.F.; Kissinger, J.C.; Gilmore, M.S.; Murray, P.R.; Mulder, R.; Solliday, J.; Clarke, B. In vitro susceptibility studies of vancomycin-resistant Enterococcus faecalis. Antimicrob. Agents Chemother. 1989, 33, 1588–1591. [Google Scholar] [CrossRef] [Green Version]
- Jensen, K.F. The Escherichia coli K-12 “wild types” W3110 and MG1655 have an rph frameshift mutation that leads to pyrimidine starvation due to low pyrE expression levels. J. Bacteriol. 1993, 175, 3401–3407. [Google Scholar] [CrossRef] [Green Version]
- Appleyard, R.K. Segregation of new lysogenic types during growth of a doubly lysogenic strain derived from Escherichia coli K12. Genetics 1954, 39, 440–452. [Google Scholar]
- Patterson, T.A.; Dean, M. Preparation of high titer lambda phage lysates. Nucleic Acids Res. 1987, 15, 6298. [Google Scholar] [CrossRef]
- Bachmann, B.J. Pedigrees of some mutant strains of Escherichia coli K-12. Bacteriol. Rev. 1972, 36, 525–557. [Google Scholar] [CrossRef]
- Griffin, P.M.; Ostroff, S.M.; Tauxe, R.V.; Greene, K.D.; Wells, J.G.; Lewis, J.H.; Blake, P.A. Illnesses associated with Escherichia coli O157:H7 infections. Ann. Intern. Med. 1988, 109, 705–712. [Google Scholar] [CrossRef]
- Khalifa, L.; Brosh, Y.; Gelman, D.; Coppenhagen-Glazer, S.; Beyth, S.; Poradosu-Cohen, R.; Que, Y.-A.; Beyth, N.; Hazan, R. Targeting Enterococcus faecalis biofilms with phage therapy. Appl. Environ. Microbiol. 2015, 81, 2696–2705. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001; pp. 2.47–2.51. [Google Scholar]
- Necel, A.; Bloch, S.; Nejman-Faleńczyk, B.; Grabski, M.; Topka, G.; Dydecka, A.; Kosznik-Kwaśnicka, K.; Grabowski, Ł.; Jurczak-Kurek, A.; Wolkowicz, T.; et al. Characterization of a bacteriophage, vB_Eco4M-7, that effectively infects many Escherichia coli O157 strains. Sci. Rep. 2020, 10, 3743. [Google Scholar] [CrossRef]
- Bloch, S.; Nejman-Faleńczyk, B.; Łoś, J.M.; Barańska, S.; Łepek, K.; Felczykowska, A.; Łoś, M.; Węgrzyn, G.; Węgrzyn, A. Genes from the exo–xis region of λ and Shiga toxin-converting bacteriophages influence lysogenization and prophage induction. Arch. Microbiol. 2013, 195, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Topka, G.; Bloch, S.; Nejman-Faleńczyk, B.; Gąsior, T.; Jurczak-Kurek, A.; Necel, A.; Dydecka, A.; Richert, M.; Węgrzyn, G.; Węgrzyn, A. Characterization of, bBacteriophage vB-EcoS-95, isolated from urban sewage and revealing extremely rapid lytic development. Front. Microbiol. 2019, 9, 3326. [Google Scholar] [CrossRef]
- Mohamed, J.A.; Huang, W.; Nallapareddy, S.R.; Teng, F.; Murray, B.E. Influence of origin of isolates, especially Endocarditis isolates, and aarious genes on biofilm formation by Enterococcus faecalis. Infect. Immun. 2004, 72, 3658–3663. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, J.C.; Peabody, D.S. Stability and assembly in vitro of bacteriophage PP7 virus-like particles. J. Nanobiotechnol. 2007, 5, 10. [Google Scholar] [CrossRef] [Green Version]
- Essoh, C.; Latino, L.; Midoux, C.; Blouin, Y.; Loukou, G.; Nguetta, S.-P.A.; Lathro, S.; Cablanmian, A.; Kouassi, A.K.; Vergnaud, G.; et al. Investigation of a large collection of Pseudomonas aeruginosa bacteriophages collected from a single environmental source in Abidjan, Côte d’Ivoire. PLoS ONE 2015, 10, e0130548. [Google Scholar] [CrossRef] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Marchler-Bauer, A.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; et al. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2016, 45, D200–D203. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2015, 44, D694–D697. [Google Scholar] [CrossRef]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing phage genomes for therapeutic applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef] [Green Version]
- Stothard, P.; Wishart, D.S. Circular genome visualization and exploration using CGView. Bioinformatics 2004, 21, 537–539. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Chan, P.P. tRNAscan-SE On-line: Integrating search and context for analysis of transfer RNA genes. Nucleic Acids Res. 2016, 44, W54–W57. [Google Scholar] [CrossRef]
- Chan, P.P.; Lowe, T.M. tRNAscan-SE: Searching for tRNA genes in genomic sequences. Methods Mol. Biol. 2019, 1962, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, M.; Rocha, M.; Oliveira, H.; Dias, O. Predicting promoters in phage genomes using PhagePromoter. Bioinformatics 2019, 35, 5301–5302. [Google Scholar] [CrossRef] [PubMed]
- Hofacker, I.L.; Fontana, W.; Stadler, P.F.; Bonhoeffer, S.; Tacker, M.; Schuster, P. Fast folding and comparison of RNA secondary structures. Mon. Chem. Chem. Mon. 1994, 125, 167–188. [Google Scholar] [CrossRef]
- Gautheret, D.; Lambert, A. Direct RNA motif definition and identification from multiple sequence alignments using secondary structure profiles 1 1Edited by J. Doudna. J. Mol. Boil. 2001, 313, 1003–1011. [Google Scholar] [CrossRef] [PubMed]
- Lesnik, E.A.; Sampath, R.; Levene, H.B.; Henderson, T.J.; McNeil, J.A.; Ecker, D.J. Prediction of rho-independent transcriptional terminators in Escherichia coli. Nucleic Acids Res. 2001, 29, 3583–3594. [Google Scholar] [CrossRef] [Green Version]
- Macke, T.J. RNAMotif, an RNA secondary structure definition and search algorithm. Nucleic Acids Res. 2001, 29, 4724–4735. [Google Scholar] [CrossRef]
- Galtier, N.; Gouy, M.; Gautier, C. Seaview and Phylo_Win: Two graphic tools for sequence alignment and molecular phylogeny. Comput. Appl. Biosci. 1996, 12, 543–548. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, R.; Nei, M. Efficiencies of the NJp, maximum likelihood, and Bayesian methods of phylogenetic construction for compositional and non-compositional genes. Mol. Boil. Evol. 2016, 33, 1618–1624. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Version 4; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Phage Sensitivity a |
|---|---|
| Enterococcus faecalis 230 | − |
| Enterococcus faecalis 271 | + |
| Enterococcus faecalis 272 | + |
| Enterococcus faecalis 273 | − |
| Enterococcus faecalis 274 | − |
| Enterococcus faecalis 275 | − |
| Enterococcus faecalis M2056 | − |
| Enterococcus faecalis OG1RF | − |
| Enterococcus faecalis V583 | + |
| Detected Protein | Predicted Function | Molecular Mass (kDa) | Number of Peptides | Sequence Coverage (%) | Protein Score |
|---|---|---|---|---|---|
| vB_EfaS-271_08 | portal protein | 43.0 | 18 | 83.03 | 144.10 |
| vB_EfaS-271_09 | prohead protease | 21.1 | 3 | 63.64 | 10.64 |
| vB_EfaS-271_10 | major capsid protein | 45.3 | 38 | 91.34 | 1412.60 |
| vB_EfaS-271_11 | major tail protein | 7.0 | 5 | 93.94 | 79.72 |
| vB_EfaS-271_14 | head-tail joining protein | 15.3 | 2 | 54.81 | 1.94 |
| vB_EfaS-271_15 | head-tail joining protein | 14.2 | 3 | 44.63 | 8.62 |
| vB_EfaS-271_16 | major tail protein | 19.9 | 7 | 65.96 | 4.38 |
| vB_EfaS-271_18 | tail tape measure protein | 156.0 | 44 | 78.64 | 131.99 |
| vB_EfaS-271_19 | tail protein | 78.9 | 34 | 77.13 | 168.36 |
| vB_EfaS-271_20 | minor tail protein | 88.4 | 44 | 88.65 | 161.67 |
| vB_EfaS-271_21 | tail fiber protein | 9.2 | 2 | 97.53 | 1.61 |
| vB_EfaS-271_23 | lysin | 39.9 | 8 | 45.21 | 4.04 |
| vB_EfaS-271_25 | DNA polymerase B-like protein | 87.5 | 11 | 73.00 | 11.70 |
| vB_EfaS-271_28 | unknown protein | 26.5 | 8 | 70.21 | 13.99 |
| vB_EfaS-271_49 | DNA primase | 60.7 | 31 | 89.58 | 149.50 |
| External Factors (Time and Conditions of Incubation) a | Percentage of Viability of vB_EfaS-271 under Certain Conditions ± SD |
|---|---|
| −80 °C (16 h) | 100.0 ± 0.0 |
| −20 °C (16 h) | 100.0 ± 1.9 |
| 20 °C (24 h) | 100.0 ± 0.0 |
| 30 °C (24 h) | 100.0 ± 0.5 |
| 37 °C (24 h) | 100.0 ± 0.0 |
| 40 °C (40 min) | 100.0 ± 7.9 |
| 42 °C (40 min) | 42.6 ± 11.3 |
| 62 °C (40 min) | 1.4 ± 0.5 |
| 95 °C (5 min) | 0.0 ± 0.0 |
| pH 2 (1 h; 37 °C) | 0.0 ± 0.0 |
| pH 4 (1 h; 37 °C) | 71.4 ± 2.8 |
| pH 10 (1 h; 37 °C) | 100.0 ± 0.0 |
| pH 12 (1 h; 37 °C) | 90.0 ± 12.3 |
| Osmotic shock (15 min; RT) | 78.8 ± 10.5 |
| 0.09% SDS (20 min; 45 °C) | 10.3 ± 2.6 |
| 0.1% CTAB (1 min; RT) | 0.0 ± 0.0 |
| 0.1% Sarkosyl (10 min; RT) | 100 ± 9.8 |
| 63% Ethanol (1 h; RT) | 0.2 ± 0.0 |
| 90% Acetone (1 h; RT) | 0.9 ± 0.4 |
| 50% DMSO (1 h; RT) | 82.4 ± 12.5 |
| Chloroform (1.5 h; 4 °C) | 55.6 ± 9.6 |
| 10% dish soap (5 min; RT) | 93.0 ± 6.4 |
| 10% soap (2 min; RT) | 77.8 ± 11.1 |
| Line-Antibacterial 70 (5 min; RT) | 81.5 ± 6.4 |
| 0.5% Virkon (30 min; RT) | 0.0 ± 0.0 |
| 5% Viruton Pulver (30 min; 30 °C) | 0.0 ± 0.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topka-Bielecka, G.; Bloch, S.; Nejman-Faleńczyk, B.; Grabski, M.; Jurczak-Kurek, A.; Górniak, M.; Dydecka, A.; Necel, A.; Węgrzyn, G.; Węgrzyn, A. Characterization of the Bacteriophage vB_EfaS-271 Infecting Enterococcus faecalis. Int. J. Mol. Sci. 2020, 21, 6345. https://doi.org/10.3390/ijms21176345
Topka-Bielecka G, Bloch S, Nejman-Faleńczyk B, Grabski M, Jurczak-Kurek A, Górniak M, Dydecka A, Necel A, Węgrzyn G, Węgrzyn A. Characterization of the Bacteriophage vB_EfaS-271 Infecting Enterococcus faecalis. International Journal of Molecular Sciences. 2020; 21(17):6345. https://doi.org/10.3390/ijms21176345
Chicago/Turabian StyleTopka-Bielecka, Gracja, Sylwia Bloch, Bożena Nejman-Faleńczyk, Michał Grabski, Agata Jurczak-Kurek, Marcin Górniak, Aleksandra Dydecka, Agnieszka Necel, Grzegorz Węgrzyn, and Alicja Węgrzyn. 2020. "Characterization of the Bacteriophage vB_EfaS-271 Infecting Enterococcus faecalis" International Journal of Molecular Sciences 21, no. 17: 6345. https://doi.org/10.3390/ijms21176345