Internalization of the Aspergillus nidulans AstA Transporter into Mitochondria Depends on Growth Conditions, and Affects ATP Levels and Sulfite Oxidase Activity

 ,
,
Abstract
:
1. Introduction
2. Results
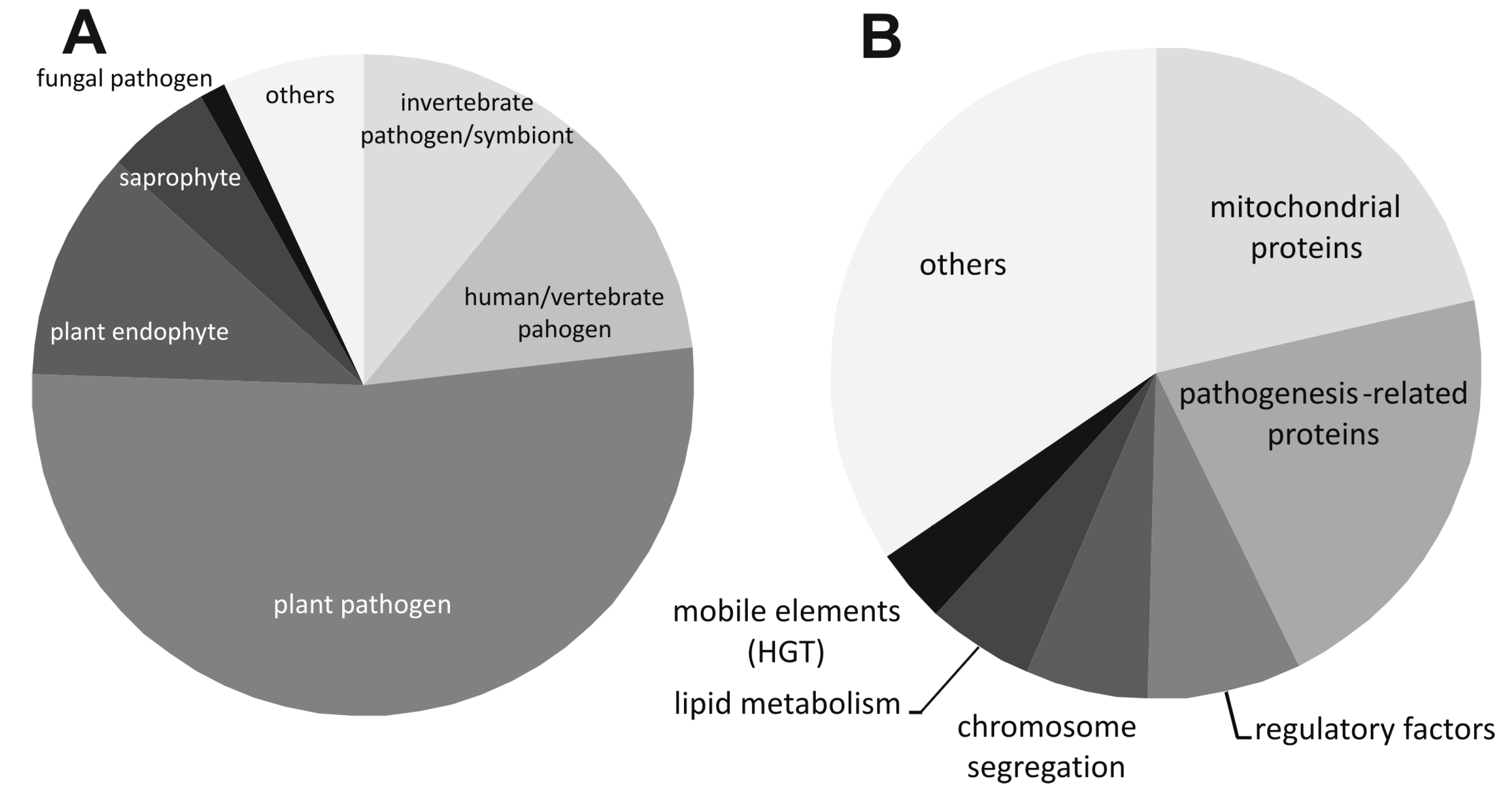
2.1. Conservation of the astA Gene in Pathogenic or Endophytic Fungi of Dikarya Phylum
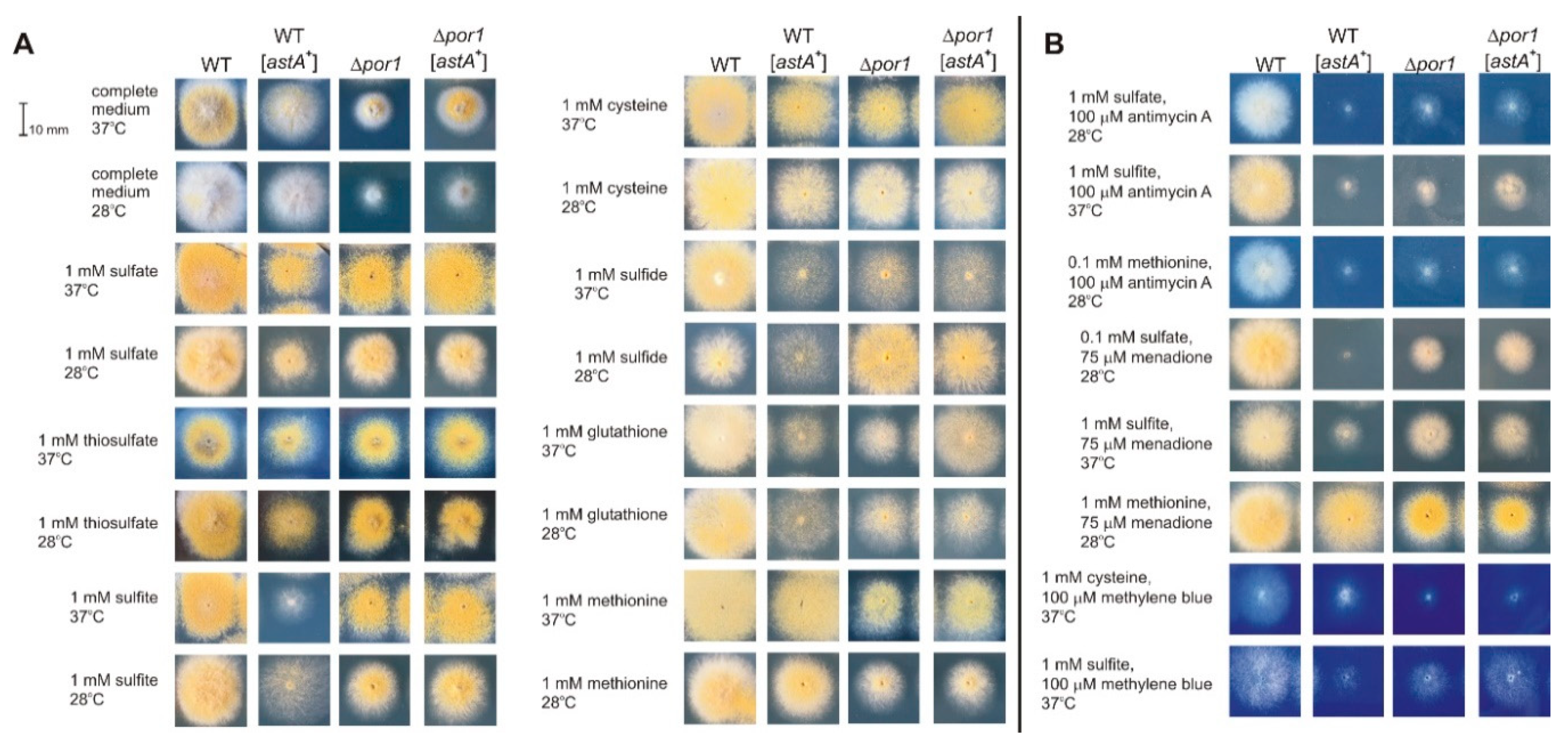
2.2. Redox-Reactive Sulfur Sources or Altered Redox Conditions Are Detrimental for Growth of an astA-Overexpressing Strain
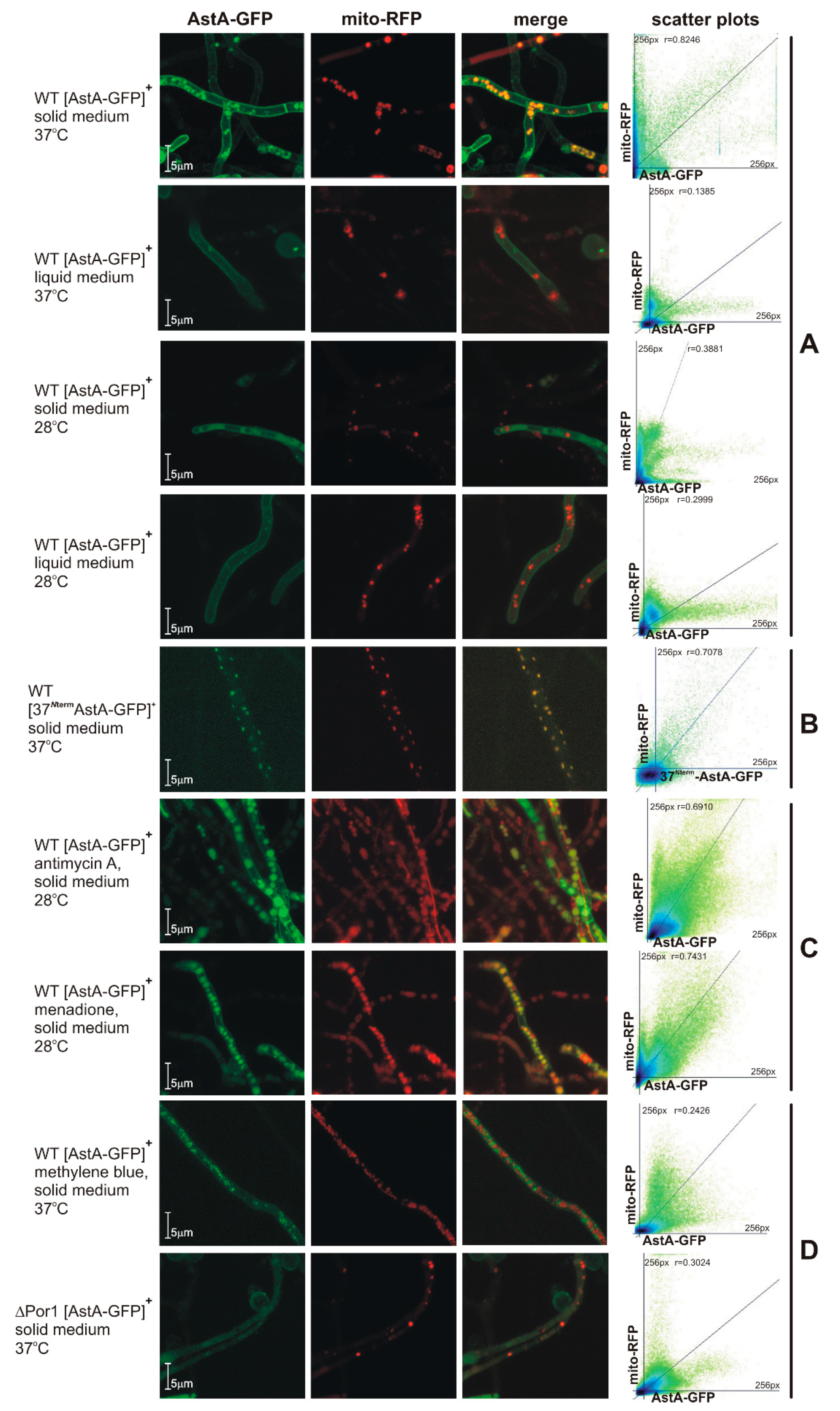
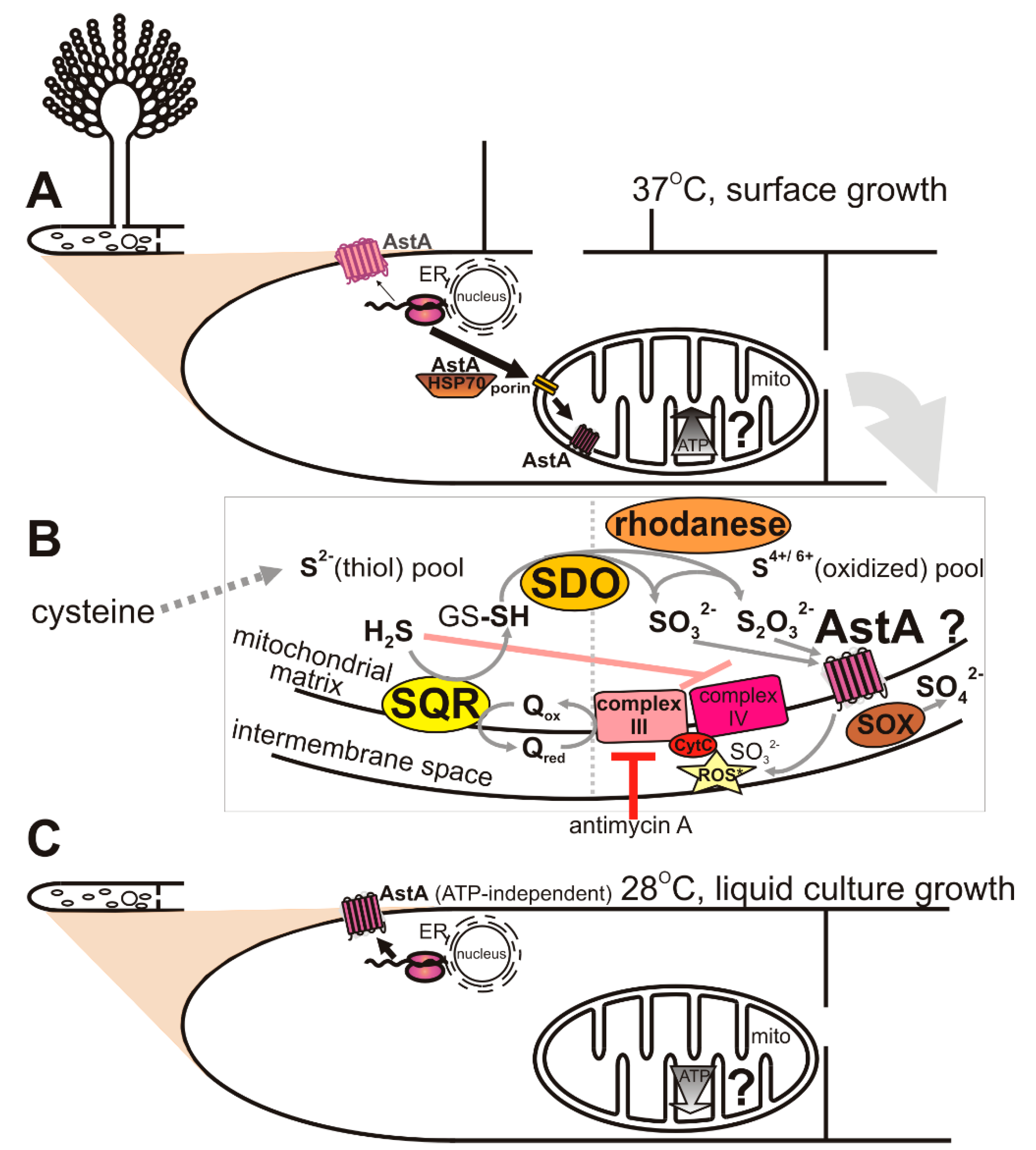
2.3. Dual-Cell-Membrane and Mitochondrial Localization of AstA Depends on Culture Conditions and Is Achieved via N-Terminal Cytoplasmic Targeting Signal
2.4. AstA Internalization into the Mitochondria via Hsp70–Porin1 Import System
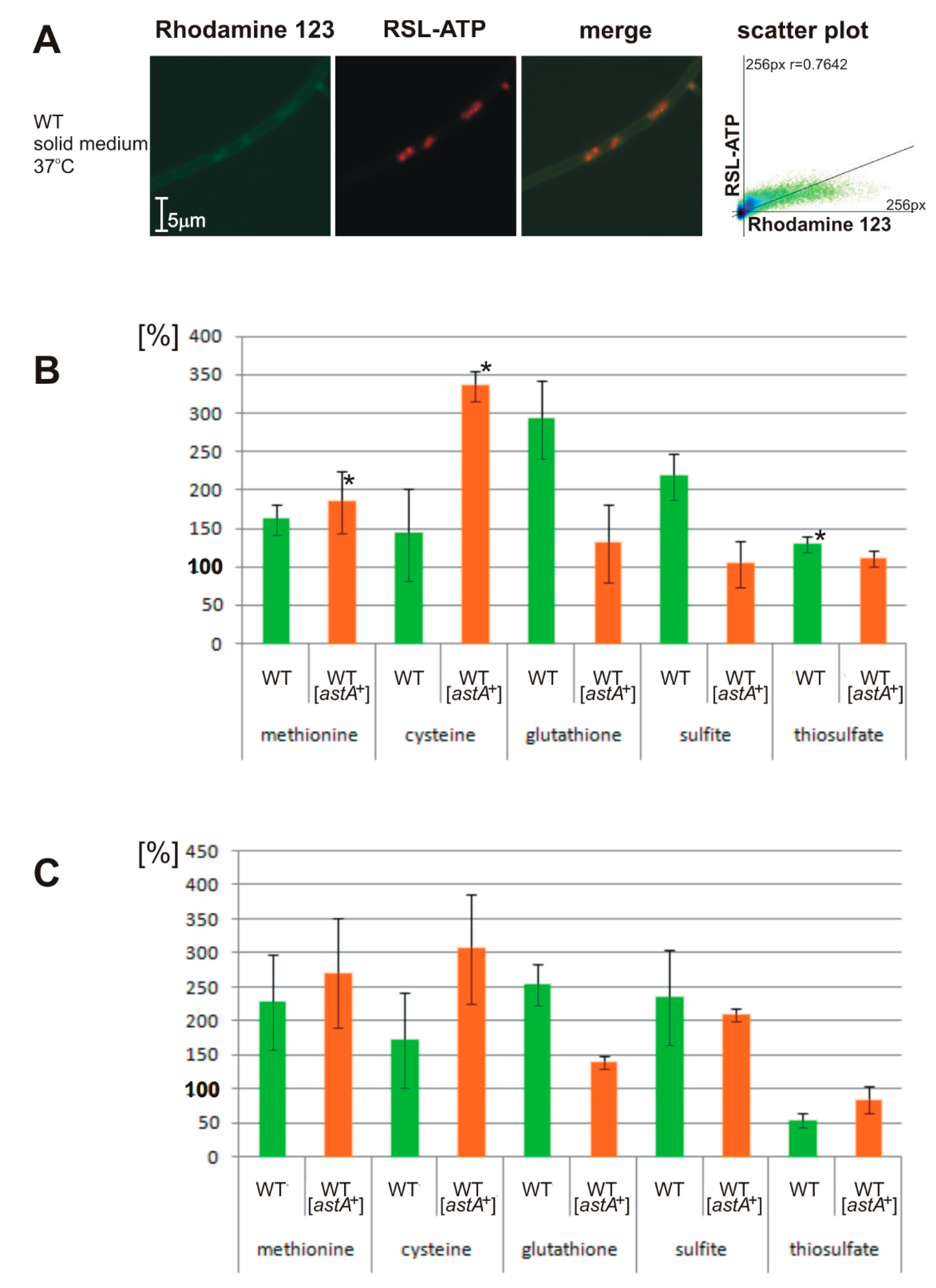
2.5. Altered ATP Levels and Sulfite Oxidase (SOX) Activity in astA-Overexpressing Strain Dependent on Sulfur Sources
3. Discussion
4. Materials and Methods
4.1. Strains and Media
4.2. Growth Conditions
4.3. Nucleic Acid Manipulation and Plasmid Construction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Genotype or Relevant Features | Reference or Source |
|---|---|---|
| E. coli strain XL1 Blue | recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F’ proAB lacIq ZΔM15 Tn10 (Tetr)]c | Stratagene |
| A. nidulans strains | ||
| W1 (WT) | pyroA4 yA2 | [34] |
| W13 | pyrG89 pyroA4 yA2 | [39] |
| W13 [astA+] (WT [astA+]) | pyrG89 [astA+ Ncpyr-4+] pyroA4 yA2 | [3] |
| WT [37NtermastA-GFP+, mitoRFPff+] | pyroA4 yA2 [37NtermastA-GFP+::gen, mitoRFPff+] | This work |
| WT ΔPorin | pyrG89 [por1::Ncpyr-4+] pyroA4 yA2 | This work |
| WTΔPorin [ astA-GFP+, mitoRFPff+] | pyrG89 [por1::Ncpyr-4+] pyroA4 yA2 [astA-GFP+, mitoRFPff+] | This work |
| Plasmids | Description | |
| pBluescript KS(–) | Cloning vector, Ampr | Stratagene |
| pGEM-T Easy | Linear vector for cloning of PCR products | Promega |
| HELp1 | Increases A. nidulans transformation efficiency about 200 fold | [40] |
| pEGFP-N1 | Vector bearing GFP-encoding ORF and gentamycin selectable cassette | Clontech |
| pEastA37NtermGFP-N1 | pEGFP-N1 plasmid bearing the astA ORF 5′ sequence fragment encoding first 37 amino acids | This work |
| pXY142-mtRFPff | Bearing nucleotide sequence encoding N. crassa H+ mitochondrial ATPase leader fused to fast folding RFP-encoding sequence | Prof. Shaw gift |
| kPG-26B | Bearing entire N. crassa pyr-4 in pBluescript II KS(−) | laboratory collection |
| kPMS11-52 | BamHI-SalI insert bearing the astA gene in pBluescript KS(–) vector | [1] |
| kPMS11-5243 (astA-GFP) | kPMS11-524-derived astA gene fused with GFP-encoding gene with N. crassa pyr-4 cassette | [3] |
| pGEMPor1 | pGEM-T Easy bearing PCR fragment of entire AN4402 locus (porin) | This work |
| pGEMPor1::pyr-4 | pGEMPor1 bearing disrupted AN4402 locus with N. crassa pyr-4 | This work |
4.4. Transformation of A. nidulans
4.5. ATP Measurements and Sulfite Oxidase Assay
4.6. Confocal Microscopy
4.7. Bioinformatic Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| SOX | sulfite oxidase |
| GSH | glutathione |
| GS-SH | persulfide glutathione |
| RSL | rhodamine-based spirolactam |
References
- Piłsyk, S.; Natorff, R.; Sieńko, M.; Paszewski, A. Sulfate transport in Aspergillus nidulans A novel gene encoding alternative sulfate transporter. Fungal Genet. Biol. 2007, 44, 715–725. [Google Scholar] [CrossRef]
- Piłsyk, S.; Paszewski, A. Sulfate permeases-hylogenetic diversity of sulfate transport. Acta Biochim. Pol. 2009, 56, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Piłsyk, S.; Sieńko, M.; Brzywczy, J.; Perlińska-Lenart, U.; Golan, M.P.; Kruszewska, J.S. Molecular characterization of central cytoplasmic loop in Aspergillus nidulans AstA transporter. Acta Biochim. Pol. 2018, 65, 545–554. [Google Scholar] [PubMed]
- Holt, S.; Kankipati, H.; De Graeve, S.; Van Zeebroeck, G.; Foulquié-Moreno, M.R.; Lindgreen, S.; Thevelein, J.M. Major sulfonate transporter Soa1 in Saccharomyces cerevisiae and considerable substrate diversity in its fungal family. Nat. Commun. 2017, 8, 14247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirayama, S.; Udagawa, S. Taxonomic studies of fungi on stored rice grains II. Aspergillus group. Bull. Fac Agric. Mie Univ. 1958, 16, 7–28. [Google Scholar]
- Pontecorvo, G.; Roper, J.A.; Hemmons, L.M.; MacDonald, K.D.; Bufton, A.W. The genetics of Aspergillus nidulans. Adv. Genet. 1953, 5, 141–238. [Google Scholar]
- Piłsyk, S.; Natorff, R.; Gawińska-Urbanowicz, H.; Kruszewska, J.S. Fusarium sambucinum astA gene expressed during potato infection is a functional orthologue of Aspergillus nidulans astA. Fungal Biol. 2015, 119, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Libiad, M.; Yadav, P.K.; Vitvitsky, V.; Martinov, M.; Banerjee, R. Organization of the human mitochondrial hydrogen sulfide oxidation pathway. J. Biol. Chem. 2014, 289, 30901–30910. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Banerjee, R. PLP-dependent H2S biogenesis. Biochim. Biophys. Acta. 2011, 1814, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Kabil, O.; Banerjee, R. Enzymology of H2S Biogenesis, Decay and Signaling. Antioxid. Redox Signal. 2014, 20, 770–782. [Google Scholar] [CrossRef] [Green Version]
- Olson, K.R.; Whitfield, N.L. Hydrogen sulfide and oxygen sensing in the cardiovascular system. Antioxid. Redox Signal. 2010, 12, 1219–1234. [Google Scholar] [CrossRef] [PubMed]
- Shoshan-Barmatz, V.; De, S. Mitochondrial VDAC, the Na+/Ca2+ Exchanger, and the Ca2+ Uniporter in Ca2+ Dynamics and Signaling. Adv. Exp. Med. Biol. 2017, 981, 323–347. [Google Scholar] [PubMed]
- Shoshan-Barmatz, V.; Maldonado, E.N.; Krelin, Y. VDAC1 at the crossroads of cell metabolism; apoptosis and cell stress. Cell Stress 2017, 1, 11–36. [Google Scholar] [CrossRef]
- Young, J.C.; Hoogenraad, N.J.; Hartl, F.U. Molecular chaperones Hsp90 and Hsp70 deliver preproteins to the mitochondrial import receptor Tom70. Cell 2003, 112, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Zemanovic, S.; Ivanov, M.V.; Ivanova, L.V.; Bhatnagar, A.; Michalkiewicz, T.; Teng, R.J.; Kumar, S.; Rathore, R.; Pritchard, K.A., Jr.; Konduri, G.G.; et al. Dynamic Phosphorylation of the C Terminus of Hsp70 Regulates the Mitochondrial Import of SOD2 and Redox Balance. Cell Rep. 2018, 25, 2605–2616.e7. [Google Scholar] [CrossRef] [Green Version]
- Velasco, L.; Dublang, L.; Moro, F.; Muga, A. The Complex Phosphorylation Patterns that Regulate the Activity of Hsp70 and Its Cochaperones. Int. J. Mol. Sci. 2019, 20, 4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Y.Y.; Cao, Y.B.; Xu, Z.; Ying, K.; Li, Y.; Xie, Y.; Zhu, Z.Y.; Chen, W.S.; Jiang, Y.Y. cDNA microarray analysis of differential gene expression in Candida albicans biofilm exposed to farnesol. Antimicrob. Agents Chemother. 2005, 49, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Martineau, C.N.; Beckerich, J.M.; Kabani, M. Flo11p-independent control of “mat” formation by hsp70 molecular chaperones and nucleotide exchange factors in yeast. Genetics 2007, 177, 1679–1689. [Google Scholar] [CrossRef] [Green Version]
- Faria-Oliveira, F.; Carvalho, J.; Ferreira, C.; Hernáez, M.L.; Gil, C.; Lucas, C. Quantitative differential proteomics of yeast extracellular matrix: There is more to it than meets the eye. BMC Microbiol. 2015, 15, 271. [Google Scholar] [CrossRef] [Green Version]
- Gil-Bona, A.; Amador-García, A.; Gil, C.; Monteoliva, L. The external face of Candida albicans: A proteomic view of the cell surface and the extracellular environment. J. Proteom. 2018, 180, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Sanchala, D.; Bhatt, L.K.; Pethe, P.; Shelat, R.; Kulkarni, Y.A. Anticancer activity of methylene blue via inhibition of heat shock protein 70. Biomed. Pharmacother. 2018, 107, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Loor, G.; Kondapalli, J.; Schriewer, J.M.; Chandel, N.S.; Vanden Hoek, T.L.; Schumacker, P.T. Menadione Triggers Cell Death Through ROS-dependent Mechanisms Involving PARP Activation Without Requiring Apoptosis. Free Radic. Biol. Med. 2010, 49, 1925–1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Barboza, G.D.; Guizzardi, S.; Moine, L.; de Talamoni, N.T. Oxidative Stress, Antioxidants and Intestinal Calcium Absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Esser, L.; Hossain, M.B.; Xia, D.; Yu, C.A.; Rizo, J.; van der Helm, D.; Deisenhofer, J. Structure of Antimycin A1, a Specific Electron Transfer Inhibitor of Ubiquinol–Cytochrome c Oxidoreductase. J. Am. Chem. Soc. 1999, 121, 4902–4903. [Google Scholar] [CrossRef]
- Huang, L.S.; Cobessi, D.; Tung, E.Y.; Berry, E.A. Binding of the Respiratory Chain Inhibitor Antimycin to the Mitochondrial bc1 Complex: A New Crystal Structure Reveals an Altered Intramolecular Hydrogen-Bonding Pattern. J. Mol. Biol. 2005, 351, 573–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yogev, O.; Pines, O. Dual targeting of mitochondrial proteins: Mechanism, regulation and function. Biochim. Biophys. Acta 2011, 1808, 1012–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avadhani, N.G.; Sangar, M.C.; Bansal, S.; Bajpai, P. Bimodal targeting of cytochrome P450s to endoplasmic reticulum and mitochondria: The concept of chimeric signals. FEBS J. 2011, 278, 4218–4229. [Google Scholar] [CrossRef] [Green Version]
- Regev-Rudzki, N.; Yogev, O.; Pines, O. The mitochondrial targeting sequence tilts the balance between mitochondrial and cytosolic dual localization. J. Cell Sci. 2008, 121, 2423–2431. [Google Scholar] [CrossRef] [Green Version]
- Craig, E.A. Hsp70 at the membrane: Driving protein translocation. BMC Biol. 2018, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, R.O., Jr.; Bus, J.S.; Popp, J.A.; Boreiko, C.J.; Andjelkovich, D.A. A critical review of the literature on hydrogen sulfide toxicity. Crit. Rev. Toxicol. 1984, 13, 25–97. [Google Scholar] [CrossRef]
- Velayutham, M.; Hemann, C.F.; Cardounel, A.J.; Zweier, J.L. Sulfite Oxidase Activity of Cytochrome c: Role of Hydrogen Peroxide. Biochem. Biophys. Rep. 2016, 5, 96–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieńko, M.; Natorff, R.; Skoneczny, M.; Kruszewska, J.; Paszewski, A.; Brzywczy, J. Regulatory mutations affecting sulfur metabolism induce environmental stress response in Aspergillus nidulans. Fungal Genet. Biol. 2014, 65, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, S.; Chandra, T.S. Oxidative stress protection and glutathione metabolism in response to hydrogen peroxide and menadione in riboflavinogenic fungus Ashbya gossypii. Appl. Biochem. Biotechnol. 2014, 174, 2307–2325. [Google Scholar] [CrossRef] [PubMed]
- Clutterbuck, A.J. Linkage map and locus list. Prog. Ind. Microbiol. 1994, 29, 791–824. [Google Scholar]
- Martinelli, S.D. Gene symbols. Prog. Ind. Microbiol. 1994, 29, 825–827. [Google Scholar]
- Cove, D.J. The induction and repression of nitrate reductase in the fungus Aspergillus nidulans. Biochim. Biophys. Acta 1966, 113, 51–56. [Google Scholar] [CrossRef]
- Lukaszkiewicz, Z.; Paszewski, A. Hyper-repressible operator-type mutant in sulphate permease gene of Aspergillus nidulans. Nature 1976, 259, 337–338. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Sieńko, M.; Natorff, R.; Zieliński, Z.; Hejduk, A.; Paszewski, A. Two Aspergillus nidulans genes encoding methylenetetrahydrofolate reductases are up-regulated by homocysteine. Fungal Genet. Biol. 2007, 44, 691–700. [Google Scholar] [CrossRef]
- Gems, D.H.; Clutterbuck, A.J. Co-transformation with autonomously-replicating helper plasmids facilitates gene cloning from an Aspergillus nidulans gene library. Curr. Genet. 1993, 24, 520–524. [Google Scholar] [CrossRef]
- Westermann, B.; Neupert, W. Mitochondria-targeted green fluorescent proteins: Convenient tools for the study of organelle biogenesis in Saccharomyces cerevisiae. Yeast 2000, 16, 1421–1427. [Google Scholar] [CrossRef]
- Li, C.Y.; Zou, C.X.; Li, Y.F.; Kong, X.F.; Zhou, Y.; Wu, Y.S.; Zhu, W.G. A colormetric and fluorescent chemosensor for adenosine-5′-triphosphate based on rhodamine derivative. Anal. Chim. Acta 2013, 795, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.J. The use of diaminobenzidine for spectrophotometric and acrylamide gel detection of sulfite oxidase and its applicability to hydrogen peroxide-generating enzymes. Anal. Biochem. 1973, 53, 208–222. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Altschul, S.F.; Lipman, D.J. Protein database searches for multiple alignments. Proc. Natl. Acad. Sci. USA 1990, 87, 5509–5513. [Google Scholar] [CrossRef] [Green Version]
- Nakai, K.; Horton, P. PSORT: A program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 1999, 24, 34–36. [Google Scholar] [CrossRef]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piłsyk, S.; Mieczkowski, A.; Golan, M.P.; Wawrzyniak, A.; Kruszewska, J.S. Internalization of the Aspergillus nidulans AstA Transporter into Mitochondria Depends on Growth Conditions, and Affects ATP Levels and Sulfite Oxidase Activity. Int. J. Mol. Sci. 2020, 21, 7727. https://doi.org/10.3390/ijms21207727
Piłsyk S, Mieczkowski A, Golan MP, Wawrzyniak A, Kruszewska JS. Internalization of the Aspergillus nidulans AstA Transporter into Mitochondria Depends on Growth Conditions, and Affects ATP Levels and Sulfite Oxidase Activity. International Journal of Molecular Sciences. 2020; 21(20):7727. https://doi.org/10.3390/ijms21207727
Chicago/Turabian StylePiłsyk, Sebastian, Adam Mieczkowski, Maciej P. Golan, Agata Wawrzyniak, and Joanna S. Kruszewska. 2020. "Internalization of the Aspergillus nidulans AstA Transporter into Mitochondria Depends on Growth Conditions, and Affects ATP Levels and Sulfite Oxidase Activity" International Journal of Molecular Sciences 21, no. 20: 7727. https://doi.org/10.3390/ijms21207727