Resident Memory T Cells and Their Role within the Liver
by
 , , , and
, , , and
Sonia Ghilas
1,2,* ,
,
Ana-Maria Valencia-Hernandez
1,
Matthias H. Enders
1,2,
William R. Heath
1,2 and
Daniel Fernandez-Ruiz
1 
1
Department of Microbiology and Immunology, University of Melbourne, at the Peter Doherty Institute for Infection and Immunity, Melbourne, VIC 3000, Australia
2
Australian Research Council Centre of Excellence in Advanced Molecular Imaging, University of Melbourne, Parkville, VIC 3010, Australia
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2020, 21(22), 8565; https://doi.org/10.3390/ijms21228565
Submission received: 8 October 2020
/
Revised: 29 October 2020
/
Accepted: 11 November 2020
/
Published: 13 November 2020
(This article belongs to the Special Issue Recent Advances in T Cell Immunity)
Abstract
:Immunological memory is fundamental to maintain immunity against re-invading pathogens. It is the basis for prolonged protection induced by vaccines and can be mediated by humoral or cellular responses—the latter largely mediated by T cells. Memory T cells belong to different subsets with specialized functions and distributions within the body. They can be broadly separated into circulating memory cells, which pace the entire body through the lymphatics and blood, and tissue-resident memory T (TRM) cells, which are constrained to peripheral tissues. Retained in the tissues where they form, TRM cells provide a frontline defense against reinfection. Here, we review this population of cells with specific attention to the liver, where TRM cells have been found to protect against infections, in particular those by Plasmodium species that cause malaria.
1. Introduction
The successful containment of infections relies on the speed with which immune responses of sufficient intensity are mounted. Immunological memory enables the long-term maintenance of a small fraction of those cells that responded to and resolved an earlier infection. The number of specific memory T cells generated after an infection, while declining over time, is generally larger than that of naïve T cells of the same specificities [1]. In addition, memory T cells display an enhanced antigen sensitivity, requiring lower levels of antigen for activation [2]. Memory T cells thus respond more rapidly and potently to pathogen invasion, and can exert efficient protection, potentially lifelong, against previously encountered infections. Different subsets of memory CD8+ T cells have been identified on the basis of their migratory properties, e.g., circulatory memory T cells and resident memory T cells (TRM cells). The latter have recently emerged as important mediators of protection in peripheral organs, a common point of entrance of pathogens, by inducing rapid and local responses upon antigen recall [3]. By combining transcriptional and phenotypic features with different approaches to investigate residency, studies have identified TRM cells in various disease models and within several tissue settings, including the liver. Importantly, strategies have been devised to favour the formation of TRM cells through vaccination, achieving promising results, for example, in the case of herpes virus infection in the mucosa of the female genital tract [4] and Plasmodium infection of the liver [5,6,7,8,9].
The liver is essential for the maintenance of homeostasis and is central to many metabolic and immunological processes. Hepatic functions are tightly regulated; and disturbances that lead to liver diseases such as microbial infections, chronic inflammation or cancer can result in death. The liver is also the target of certain pathogens, such as Plasmodium, Leishmania, or Listeria, which infect and develop in this organ during stages of their life cycles. Given the highly protective capacity of memory T cells, and in particular of TRM cells, studying the biology of these cells may aid the development of prophylactic and therapeutic strategies against life-threatening conditions associated with organ damage or infection. In this review, we will focus on recent advances in understanding memory T cell and TRM cell biology, focusing on liver TRM cells. Indeed, knowledge on this cell subset has been successfully implemented in the development of novel, highly effective immunization strategies against infectious diseases.
2. Memory T Cells
Shortly after activation, T cells generally differentiate into either short-lived effector cells (SLECs), expressing KLRG1, CX3CR1, and S1PR5, or memory precursor effector cells (MPECs), which are KLRG1- CX3CR1- and IL-7R+ [10,11,12,13]. T cell activation results in the formation of large numbers of SLECs, but these cells rapidly decline in numbers upon clearance of the infection. MPECs, however, are less numerous but become long lived memory cells and show a greater ability to generate recall responses [11]. IL-7Rhi cells comprise most of the memory cells at late time points (>8 months) after infection [14]. Importantly, this general classification is not exhaustive. Thus, while certain memory T cells (KLRG1hi, described in detail below) mainly arise from IL-7Rhi cells [11,12], a small proportion of IL-7Rlo cells can persist for prolonged periods [11], and display low expression of this marker in the spleen 60 days after lymphocytic choriomeningitis virus (LCMV) infection [12]. Indeed, splenic CX3CR1- (KLRG1lo) effector T cells can give rise to all circulating and non-circulating memory T cell population, while CX3CR1+ (KLRG1hi) effector cells mainly differentiate into effector memory T cells after LCMV infection [13]. However, a peptide immunization model revealed that not all IL-7R+ cells in the spleen are long-lived [15] and, conversely, some KLRG1hi T cells can persist for long periods of time, providing control against Listeria infection [16].
The establishment and long-term survival of MPECs and the memory T cells they give rise to, requires the cytokines IL-7 and IL-15 [11,17]. Downstream signaling after IL-15 and IL-7 recognition results in the expression of anti-apoptotic molecules, such as Bcl2 and Mcl1, shown to prevent the death of activated effector T cells and therefore to promote memory formation [18,19,20,21].
IL-15 signaling induces a metabolic switch from glycolysis, typical of effector T cells, to fatty acid oxidation [22], which comparably generates about 6 times more energy per unit of weight of substrate [23] and is essential to sustain memory T cell survival. Indeed, TRAF6 deficient T cells, presenting defective mitochondrial fatty acid oxidation, display an enhanced contraction phase after activation. In turn, stimulation of fatty acid metabolism in these cells with a drug that promotes AMP-activated kinases and circumvents the deficiency in TRAF6, prevents this decline in the number of activated T cells [24]. The expression of the chemokine receptor CCR7 on MPECs facilitates migration to T cell areas in secondary lymphoid organs along a CCL19 and CCL21 gradient. In these organs, T cells are exposed to IL-7 predominantly produced by stromal cells [25]. IL-7 is also produced by epithelial cells in organs such as the skin and the intestine [25]. As mentioned above, most memory T cells arise from the subpopulation of effector T cells that express IL-7R [11]; and IL-7 signaling has been linked with elevated fatty acid uptake and oxidation in CD8+ T cells through the induction of aquaporin 9 expression, a glycerol transporter that supports fatty acid uptake [26].
In the absence of IL-15, basal CD8+ T cell memory proliferation is impaired and leads to a progressive decline in memory T cell numbers [27,28]. In addition, under steady state conditions or after infection, mice lacking IL-15 display low numbers TRM cells in liver and skin [29,30,31], suggesting this cytokine provides an important maintenance and/or developmental signal for resident memory T cells. However, more recent studies suggest that IL-15 dependency might not be absolute for CD8+ memory T cells or tissue-resident T cell populations in some organs, such as the mucosa and central nervous system, after viral infection [32,33,34].
Memory T cells were initially separated into two subsets based on the expression of the lymph node homing molecules CCR7 and CD62L, with CCR7+ CD62L+ cells being termed central memory (TCM), and CCR7- CD62L- cells, effector memory T (TEM) cells [35]. TCM cells were found to migrate through lymphoid tissues, whereas TEM cells were thought to traffic through peripheral tissues and the blood [35,36]. However, recent work has shown that T cell memory populations display a higher degree of complexity. Based on the expression of the chemokine receptor CX3CR1, CX3CR1int peripheral memory (TPM) cells can be discriminated from CX3CR1- TCM and CX3CR1hi TEM cells [13]. Gerlach et al. showed that TPM cells can also express CCR7 and CD62L, reflecting a TCM phenotype. However, contradicting previous descriptions [35,36], this study found CX3CR1int TPM cells in tissues and the thoracic duct lymph, while CX3CR1high TEM cells were predominantly found in the blood. Gerlach et al. therefore concluded that TPM cells and not TEM cells embody the major migratory memory subsets in peripheral tissues [13]. Another memory T cell subpopulation described, in humans and mice, are termed memory T stem cells (TSCM) [37,38]. These cells are CD44low CD62Lhi, similarly to naive T cells, but can be further distinguished by the expression of Bcl2 and CD122 and, in mice, of Sca-1. Transcriptome analyses showed that TSCM cells are the least differentiated memory subset population. TSCM, as their name suggests, can give rise to a variety of different T cell populations such as SLECs, TEM, and TCM cells. Furthermore, the capacity of TSCM cells for self-renewal, survival, and proliferation exceeds that of TCM and TEM cells. They are also of major interest in cancer research due to their superior anti-tumor response and resistance to chemotherapy [37,38,39].
3. Resident Memory T Cells
In addition to the aforementioned memory T cell subtypes, which all circulate throughout lymphoid and/or non-lymphoid organs, another subtype of memory T cells that reside in peripheral tissues, termed tissue-resident memory T (TRM) cells, became evident in the skin after infection with herpes simplex virus (HSV) type 1 [3]. These skin-resident CD8+ T cells were found to be in disequilibrium with circulating T cells, and efficiently controlled re-infection in a herpes simplex virus model [3]. TRM cells have now been identified in virtually all organs in mice [40,41] and humans [42] including lymphoid and non-lymphoid tissues (Table 1). Recent evidence suggests that, upon restimulation, a small portion of these cells may seed back into the circulation [43,44]. However, the veracity of this conclusion is questioned by other studies that indicate TRM cells remain localized to their niche even when exposed to antigen [45]. While we will focus on CD8+ TRM cells in this review, TRM cells can derive from both CD4+ or CD8+ T cells. TRM cells have become a major focus of T cell research throughout the last decade as they are an essential first line of defense against pathogen invasion in most tissues.
3.1. TRM Cell Development and General Features
Identification of cell surface markers that can clearly distinguish TRM cells from other memory T cell subsets in both mouse and human tissues is complicated by the fact that no single marker associated with TRM cells is exclusive to this cell subset. Different TRM cell populations are known to share a common transcriptional signature [31]. However, they can adapt to their local microenvironment resulting in marker and cell feature variations from tissue to tissue [41]. Examples of this phenomenon will be given in the following paragraphs.
The cell surface molecule CD69 is a canonical marker of TRM cells. This molecule promotes tissue retention by complexing with and antagonizing sphingosine-1 phosphate receptor 1 (S1PR1), a receptor that is required for tissue egress [46]. In mice, the majority, but not all of TRM cells retained in tissues during parabiosis studies express CD69 [41]. In humans, sorting of CD69+ memory T cells from different tissues demonstrated a conserved transcriptional profile distinct from blood memory T cells and similar to that of mouse TRM cells [47,48]. However, expression of CD69 is not sufficient to distinguish TRM cells from other T cell subsets. One major issue is that T cells express CD69 upon TCR engagement, and hence local exposure to antigen may prevent distinction of TRM cells from activated T cells. Exposure to type I IFN can also cause upregulation of this molecule on T cells [46], complicating TRM cell identification during ongoing inflammation. Finally, CD69 has been shown to be dispensable for the generation and maintenance of TRM cells in various tissues, such as liver, salivary gland, or lymph nodes [49]. Other markers are therefore necessary for TRM cell identification.
Another marker widely used to identify TRM cells is the molecule CD103 (the α subunit of the αEβ7 integrin), which binds E-cadherin expressed on epithelial and thus retains cells on the epithelium. This molecule is broadly expressed by murine TRM cells from mucosa and barrier tissues [31,50,51,52]. However, murine TRM cells from lymphoid organs and some non-barrier tissues such as the kidney and the liver do not express CD103 [6,53,54,55]. Similar observations have been made in humans where TRM cells express CD103 in mucosa and barrier tissues but not in lymphoid organs [47,48,56,57]. Interestingly, unlike mice, some human liver TRM cells do express CD103 [58]. This is thought to be related to the broad expression of E-cadherin by human hepatocytes [58], which may promote the retention of human TRM cells within the liver. On the contrary, the retention of murine liver TRM cells within the liver is achieved through the interaction of lymphocyte function-associated antigen-1 (LFA-1) with the intercellular adhesion molecule-1 (ICAM-1) expressed by the liver sinusoidal endothelial cells [55]. Thus, while CD69 and CD103 are useful markers to define TRM cells from several tissues, they are not sufficient, and the context of expression must be considered when interpreting analyses.
More recent studies in both mice and humans have demonstrated the importance of the molecule CD49a in the biology of some TRM cell subsets. This protein, also known as integrin α1, pairs with CD29 (integrin β1) to form the very late antigen (VLA-1), which binds to extracellular collagen and laminin and promotes the retention of T cells in tissues [59]. In peripheral tissues, like skin or liver, the majority, but not all, of murine and human TRM cells express CD49a [3,48,60,61,62,63]. Importantly, in human skin, CD49a expression has been shown to discriminate two functionally different populations of TRM cells, with CD49a+ TRM cells producing IFN-γ and CD49a- TRM cells producing IL-17 [64]. CD49a may play a role in adhesion of TRM cells to basement membranes of the epithelium. In support of this view, depletion of CD49a results in a decrease of memory T cells within the lung [59]. However, a recent study has shown that CD49a expression on T cells facilitates locomotion of virus specific CD8+ T cells in the trachea, suggesting that CD49a supports TRM motility in this organ [63].
Other molecules have been identified as signature markers of TRM cells in different tissues. For instance, the chemokine receptor CXCR6 is expressed by TRM cells in several mouse organs like the liver and the lung, where it promotes respectively their maintenance and airway localization [6,65,66]. Likewise, human TRM cells express CXCR6 across multiple tissues [48]. The molecules PD-1 and CD101 are also commonly expressed by TRM cells from different tissues [45,48,67,68]. In contrast, most TRM cells are negative for the chemokine receptor CX3CR1 [6], which is found on some circulating memory T cells in mice and humans [13,48]. Similarly, murine and human TRM cells do not express KLRG1, nor lymph node homing molecules, such as CD62L, CCR7, or S1PR1 [40,48,69].
Environmental factors particular to each tissue, such as the expression of differential cytokines, can shape the formation and maintenance of TRM cells. For example, tumor necrosis factor (TNF), IL-33, IL-15, IL-21, as well as transforming growth factor-β (TGF-β) have been shown to influence generation of TRM cells in various non lymphoid tissues, such as the skin, salivary glands, or intestine [31,69,70,71]. As TGF-β is known to promote CD103 upregulation, and some TRM cells such as those in the liver are CD103-, these cells are suggested to be maintained in a TGF-β independent manner. However, these cells are not unresponsive to TGF-β, as a recent RNA-seq based study revealed that TGF-β stimulation in vitro induced the upregulation of core signature TRM cell genes in CD8+ T cells from several tissues, including the liver [72].
Transcription profiling has also highlighted a broad range of transcription factors associated with TRM cell formation and/or maintenance. For instance, the development of several murine TRM cell populations, including liver resident cells, requires cooperation of the transcription factors Hobit and Blimp1 [73]. Nonetheless, in humans, different observations have been made. For instance, while Pallett et al. found that human liver TRM cells are Hobitlow Blimp1high and suggested that Blimp1 compensates for the lack of Hobit upregulation [58], Stelma et al. showed that human liver TRM cells express low levels of both molecules indicating that an alternative molecular mechanism could be involved in their differentiation process [74]. Indeed, it is possible that these studies looked at different subsets of TRM cells: a recent study on memory CD8+ T cells in the murine intestine suggests that Blimp1 expression identifies functionally and transcriptionally distinct TRM cell subsets [75]. Blimp1high TRM cells display strong effector capabilities and govern the early phase of acute infections whereas Blimp1low TRM cells are described as a memory population that persists long after infection [75].
3.2. Function of TRM Cells
Upon re-exposure to a pathogen, TRM cells provide a first line of adaptive cellular defense in peripheral non-lymphoid tissues. Mouse TRM cells from various organs have been shown to mediate rapid protection against diverse bacterial, viral, and parasitic infections with more effective and rapid pathogen clearance compared with other subsets of memory T cells [3,6,53,85,91]. TRM cells have also been associated with improved solid cancer prognosis (reviewed in [92]).
Upon antigen encounter TRM cells rapidly produce different effector molecules including cytotoxic factors like granzyme B (GzmB) or perforin, and inflammatory cytokines such as Interferon-γ (IFN-γ) and Tumor Necrosis Factor (TNF) as observed in different organs and upon various infection model [6,93,94]. Hence, TRM cells likely exert their protective function by either direct killing of infected cells or by attracting other immune cells to the site of infection. TRM cells in the skin have been found able to clear HSV infection in the absence of circulating cells [95], and WT, but not IFN-γ or perforin-deficient TRM cells in the brain were able to control intracerebral LCMV infection in mice depleted of circulating cells [94]. These findings suggest that TRM cells can mediate direct killing of pathogens. Additionally, the chemokines and inflammatory cytokines produced by TRM cells upon recall infection can trigger the recruitment and the activation of other inflammatory cells in particular circulating memory T cells [53,96,97]. As a consequence of their recruiting capacity, a small number of pathogen specific TRM cells can trigger very rapid and efficient local immunity.
As a result of their remarkable protective capacities, TRM cells have emerged as a promising means to combat infection and cancer. Indeed, recent studies on liver TRM cells provide a clear example of the protective potential of these cells, as well as the opportunities to promote their formation through vaccination for effective immunity against infection.
4. Liver TRM Cell Location
The liver is the recipient of both arterial and venous blood. The portal vein delivers large volumes of blood from the gastrointestinal tract and spleen to the liver [98]. Once there, the blood flows through narrow vascular capillaries known as hepatic sinusoids, which reduce the flow rate and allow resident cells to interact with a vast variety of antigens and circulating cells [99]. The hepatic sinusoids are lined with liver sinusoidal endothelial cells that form a fenestrated thin layer that separates hepatocytes from circulating cells. These fenestrae grant lymphocytes in the blood direct access to the surface of hepatocytes for antigen recognition and effector function [100,101]. In contrast to TRM cells in most tissues, which are anatomically separated from the circulation, liver TRM cells are present within the sinusoids and are constantly exposed to the blood stream but are able to access antigen on tissue stroma through the fenestrated endothelium [6]. Intravital images shows that liver TRM cells, which display an ameboid shape, are uniquely located in the vasculature where they patrol the hepatic sinusoids at migration speeds more rapid than seen for skin TRM cells (Figure 1) [6,41,73].
4.1. Identification of Liver TRM Cells
Malaria is a major infectious disease caused by Plasmodium parasites. In their vertebrate host, parasites first develop in the liver for a short period of time, where they infect hepatocytes, before being released into the bloodstream to cause blood-stage infection, which leads to disease symptoms. Early evidence supporting the existence of resident memory T cells in the liver came from studies investigating the role of CD8+ T cells against the liver-stage of Plasmodium. These studies identified a long-lasting population of memory CD8+ T cells present in the liver and absent in the spleen of mice vaccinated with radiation-attenuated Plasmodium sporozoites (the infectious stage transmitted by the mosquito) [102]. Vaccinated mice were protected against Plasmodium sporozoite challenge for more than 6 months [102]. Later reports revealed that a subpopulation of memory CD8+ T cells associated with the liver, but absent from the circulation, expressed high levels of CXCR6, CXCR3, and CD69 [5,65], markers commonly displayed by TRM cells [103].
The presence of bona fide memory cells permanently residing in the liver was confirmed by parabiosis studies in mice systemically infected with LCMV or Plasmodium sporozoites [6,41]. Parabiosis requires the surgical union of the flank skin of two animals. This enables the mixing of blood between the parabionts, and thus evaluation of T cell migration from one animal to the other. Unlike circulating cells, which equilibrate between both animals, resident populations remain in the parabiont in which they originally formed. This technique has been extensively used to identify TRM cells in different murine tissues [41]. Although liver TRM cells are in constant contact with circulating blood [6], parabiosis studies have confirmed that these cells, counterintuitively, do not recirculate and can only be found in the livers of the immunized parabiont partner [6,41].
Liver TRM cells were found to express a similar phenotypic and transcriptional signature to that of TRM cells previously identified in the lung, skin, and gut [6,31]. Maintenance of liver TRM cells in mice relies on the expression of the transcription factor Hobit, and on basal levels of expression of Blimp1 [73]. These TRM cell signatures have been found in T cells from grafted or isolated human tissues, enabling the unequivocal identification of TRM cells in several human organs [48], including the liver [58,74]. As mentioned earlier, contrary to liver TRM cells in mice which express high levels of Hobit and low to intermediate levels of Blimp1 [73], human liver TRM cells are Hobitlow Blimp1high [58]. In a recent publication, a small proportion of donor cells were found in HLA-mismatched liver and allografts 11 years after transplant, demonstrating the resident nature and remarkable longevity of these cells [104].
4.2. Liver TRM Cell Immune Responses to Infection
Murine studies have shown that liver TRM cells can confer efficient protection against liver-stage Plasmodium infection [6,9]. These studies have also demonstrated that substantial numbers of liver TRM cells are associated with higher levels of immunity to malaria, and depletion of these cells ablates protection [6,9]. Based on these results, several complex vaccinations strategies, aimed at trapping activated CD8+ T cells in the liver, have now successfully induced the formation of liver TRM cells in mice [6,7,8,9]. One vaccination strategy, prime-and-trap, is a single injection of a 3-component vaccine designed to prime Plasmodium-specific CD8+ T cells in the spleen and recruit them to the liver to form TRM cells via locally expressed antigen recognition and adjuvant-induced inflammation [6,9]. Another strategy, termed prime and target requires the administration of two components injected two weeks apart and uses a modified adenovirus for priming and either nanoparticles or a modified viral vector to target cells to the liver [7]. More recently, we have also used a glycoprotein-peptide vaccination strategy that utilizes NKT cell help to induce the formation of liver TRM cells [8]. In mice, vaccine-induced TRM cells patrol the liver sinusoids, form aggregates around infected hepatocytes and, based on expression of molecules such as GzmB, IFN-γ and TNF-α (Figure 1) [6,7], potentially exert infection control through direct lysis and/or cytokine-mediated mechanisms. Moreover, vaccination studies with attenuated Plasmodium sporozoites in non-human primates have found high frequencies of intrahepatic memory CD8+ T cells in protected subjects [105].
Importantly, in humans, liver TRM cells have been associated with disease control. For example, recent studies have investigated paired blood and liver samples from patients with chronic hepatitis B and hepatitis C virus infection and healthy volunteers to determine the role of liver TRM cells during viral infections [58,74]. Researchers found that human TRM cells in the liver express high levels of IL-2 and accumulate in larger numbers in the livers of infected patients compared to healthy patients. These studies also determined higher expression of GzmB and IFN-γ in HBV infected patients. Importantly, an inverse correlation between liver TRM frequencies and viral titers was observed, indicating that high numbers of specific liver TRM cells were associated with viral control [58]. However, accumulation of intrahepatic CD8+ CD103+ perforin+ T cells has been observed in cases of autoimmune hepatitis, particularly in indetermined pediatric acute liver failure [106]. These findings suggest that liver TRM cells could also have a pathogenic function.
5. Conclusions
TRM cells are pivotal mediators of protective immune responses within tissues and have been identified in nearly all organs, including lymphoid, non-lymphoid and barrier tissues. They are loaded with effector molecules, including GzmB, perforin, IFN-γ, and TNF, and likely exert their function by the direct killing of targets, or by recruiting other immune cells. Several infection models have correlated the presence of TRM cells with pathogen and tumour control in tissues. Notably, in the liver, CD8+ TRM cells can mediate efficient control of liver-stage Plasmodium parasites, and likely, HBV and HCV infections. For this reason, TRM cells appear of particular interest in the course of vaccine development, especially for liver TRM cells for malaria vaccines. Further research unveiling the mechanisms for the formation and maintenance of TRM cells will facilitate the design of next generation TRM-based vaccines that realize the protective potential of these cells for unprecedented immunity against infections.
Author Contributions
S.G., A.-M.V.-H., and M.H.E. contributed equally to the conception, drafting, and writing of the manuscript. D.F.-R. and W.R.H. reviewed, edited, and supervised the work. S.G. prepared the final draft and did the revisions. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Australian Research Council (CE140100011), National Health and Medical Research Council (NHMRC; 1113293 and 1105817) and W-R.H is an NHMRC Senior Principal Research Fellow (1154457).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Ag | Antigen |
| GzmB | Granzyme B |
| HBV | Hepatitis B Virus |
| HCV | Hepatitis C Virus |
| HSV | Herpes simplex virus |
| ICAM-1 | Intercellular adhesion molecule-1 |
| IFN-γ | Interferon-γ |
| IL | Interleukin |
| LCMV | Lymphocytic choriomeningitis virus |
| LFA-1 | Lymphocyte function-associated antigen-1 |
| LSEC | Liver sinusoidal endothelial cell |
| MPEC | Memory precursor effector cells |
| NKT | Natural Killer T cells |
| RNA-seq | RNA-sequencing |
| S1PR1 | Sphingosine-1 phosphate receptor 1 |
| SLEC | Short-lived effector cells |
| TCM | Central memory T cells |
| TEM | Effector memory T cells |
| TPM | Peripheral memory T cells |
| TRM | Resident memory T cells |
| TSCM | Memory T stem cells |
| TCR | T cell receptor |
| TNF | Tumor necrosis factor |
| TGF-β | Transforming Growth Factor-β |
| VLA-1 | Very late antigen |
References
- Kalia, V.; Sarkar, S.; Ahmed, R. CD8 T-cell memory differentiation during acute and chronic viral infections. Adv. Exp. Med. Biol. 2010, 684, 79–95. [Google Scholar] [CrossRef]
- Kumar, R.; Ferez, M.; Swamy, M.; Arechaga, I.; Rejas, M.T.; Valpuesta, J.M.; Schamel, W.W.; Alarcon, B.; van Santen, H.M. Increased sensitivity of antigen-experienced T cells through the enrichment of oligomeric T cell receptor complexes. Immunity 2011, 35, 375–387. [Google Scholar] [CrossRef] [Green Version]
- Gebhardt, T.; Wakim, L.M.; Eidsmo, L.; Reading, P.C.; Heath, W.R.; Carbone, F.R. Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat. Immunol. 2009, 10, 524–530. [Google Scholar] [CrossRef]
- Shin, H.; Iwasaki, A. A vaccine strategy that protects against genital herpes by establishing local memory T cells. Nature 2012, 491, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Tse, S.W.; Cockburn, I.A.; Zhang, H.; Scott, A.L.; Zavala, F. Unique transcriptional profile of liver-resident memory CD8+ T cells induced by immunization with malaria sporozoites. Genes Immun. 2013, 14, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ruiz, D.; Ng, W.Y.; Holz, L.E.; Ma, J.Z.; Zaid, A.; Wong, Y.C.; Lau, L.S.; Mollard, V.; Cozijnsen, A.; Collins, N.; et al. Liver-Resident Memory CD8+ T Cells Form a Front-Line Defense against Malaria Liver-Stage Infection. Immunity 2016, 45, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gola, A.; Silman, D.; Walters, A.A.; Sridhar, S.; Uderhardt, S.; Salman, A.M.; Halbroth, B.R.; Bellamy, D.; Bowyer, G.; Powlson, J.; et al. Prime and target immunization protects against liver-stage malaria in mice. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holz, L.E.; Chua, Y.C.; de Menezes, M.N.; Anderson, R.J.; Draper, S.L.; Compton, B.J.; Chan, S.T.S.; Mathew, J.; Li, J.; Kedzierski, L.; et al. Glycolipid-peptide vaccination induces liver-resident memory CD8+ T cells that protect against rodent malaria. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Valencia-Hernandez, A.M.; Ng, W.Y.; Ghazanfari, N.; Ghilas, S.; de Menezes, M.N.; Holz, L.E.; Huang, C.; English, K.; Naung, M.; Tan, P.S.; et al. A Natural Peptide Antigen within the Plasmodium Ribosomal Protein RPL6 Confers Liver TRM Cell-Mediated Immunity against Malaria in Mice. Cell Host Microbe 2020, 27, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.S.; Cui, W.; Chandele, A.; Lee, H.K.; Urso, D.R.; Hagman, J.; Gapin, L.; Kaech, S.M. Inflammation directs memory precursor and short-lived effector CD8+ T cell fates via the graded expression of T-bet transcription factor. Immunity 2007, 27, 281–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaech, S.M.; Tan, J.T.; Wherry, E.J.; Konieczny, B.T.; Surh, C.D.; Ahmed, R. Selective expression of the interleukin 7 receptor identifies effector CD8 T cells that give rise to long-lived memory cells. Nat. Immunol. 2003, 4, 1191–1198. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.W.; Rutishauser, R.L.; Joshi, N.S.; Haberman, A.M.; Kaech, S.M. Differential localization of effector and memory CD8 T cell subsets in lymphoid organs during acute viral infection. J. Immunol. 2010, 185, 5315–5325. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, C.; Moseman, E.A.; Loughhead, S.M.; Alvarez, D.; Zwijnenburg, A.J.; Waanders, L.; Garg, R.; de la Torre, J.C.; von Andrian, U.H. The Chemokine Receptor CX3CR1 Defines Three Antigen-Experienced CD8 T Cell Subsets with Distinct Roles in Immune Surveillance and Homeostasis. Immunity 2016, 45, 1270–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M.D.; Kim, M.T.; Shan, Q.; Sompallae, R.; Xue, H.H.; Harty, J.T.; Badovinac, V.P. Phenotypic and Functional Alterations in Circulating Memory CD8 T Cells with Time after Primary Infection. PLoS Pathog. 2015, 11, e1005219. [Google Scholar] [CrossRef] [Green Version]
- Lacombe, M.H.; Hardy, M.P.; Rooney, J.; Labrecque, N. IL-7 receptor expression levels do not identify CD8+ memory T lymphocyte precursors following peptide immunization. J. Immunol. 2005, 175, 4400–4407. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.A.; McDonald-Hyman, C.; Jameson, S.C.; Hamilton, S.E. Effector-like CD8+ T cells in the memory population mediate potent protective immunity. Immunity 2013, 38, 1250–1260. [Google Scholar] [CrossRef] [Green Version]
- Surh, C.D.; Sprent, J. Homeostasis of naive and memory T cells. Immunity 2008, 29, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Schluns, K.S.; Kieper, W.C.; Jameson, S.C.; Lefrancois, L. Interleukin-7 mediates the homeostasis of naive and memory CD8 T cells in vivo. Nat. Immunol. 2000, 1, 426–432. [Google Scholar] [CrossRef]
- Hildeman, D.A.; Zhu, Y.; Mitchell, T.C.; Bouillet, P.; Strasser, A.; Kappler, J.; Marrack, P. Activated T cell death in vivo mediated by proapoptotic bcl-2 family member bim. Immunity 2002, 16, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Opferman, J.T.; Letai, A.; Beard, C.; Sorcinelli, M.D.; Ong, C.C.; Korsmeyer, S.J. Development and maintenance of B and T lymphocytes requires antiapoptotic MCL-1. Nature 2003, 426, 671–676. [Google Scholar] [CrossRef]
- Yajima, T.; Yoshihara, K.; Nakazato, K.; Kumabe, S.; Koyasu, S.; Sad, S.; Shen, H.; Kuwano, H.; Yoshikai, Y. IL-15 regulates CD8+ T cell contraction during primary infection. J. Immunol. 2006, 176, 507–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Windt, G.J.; Everts, B.; Chang, C.H.; Curtis, J.D.; Freitas, T.C.; Amiel, E.; Pearce, E.J.; Pearce, E.L. Mitochondrial Respiratory Capacity is a Critical Regulator of CD8+ T Cell Memory Development. Immunity 2012, 36, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodish, H.B.A.; Zipursky, S.L. Oxidation of Glucose and Fatty Acids to CO2. In Molecular Cell Biology, 4th ed.; W. H. Freeman: New York, NY, USA, 2000. [Google Scholar]
- Pearce, E.L.; Walsh, M.C.; Cejas, P.J.; Harms, G.M.; Shen, H.; Wang, L.S.; Jones, R.G.; Choi, Y. Enhancing CD8 T-cell memory by modulating fatty acid metabolism. Nature 2009, 460, 103–107. [Google Scholar] [CrossRef]
- Hara, T.; Shitara, S.; Imai, K.; Miyachi, H.; Kitano, S.; Yao, H.; Tani-ichi, S.; Ikuta, K. Identification of IL-7-producing cells in primary and secondary lymphoid organs using IL-7-GFP knock-in mice. J. Immunol. 2012, 189, 1577–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.; Staron, M.M.; Gray, S.M.; Ho, P.C.; Amezquita, R.A.; Wu, J.; Kaech, S.M. IL-7-Induced Glycerol Transport and TAG Synthesis Promotes Memory CD8+ T Cell Longevity. Cell 2015, 161, 750–761. [Google Scholar] [CrossRef] [Green Version]
- Becker, T.C.; Wherry, E.J.; Boone, D.; Murali-Krishna, K.; Antia, R.; Ma, A.; Ahmed, R. Interleukin 15 is required for proliferative renewal of virus-specific memory CD8 T cells. J. Exp. Med. 2002, 195, 1541–1548. [Google Scholar] [CrossRef] [Green Version]
- Goldrath, A.W.; Sivakumar, P.V.; Glaccum, M.; Kennedy, M.K.; Bevan, M.J.; Benoist, C.; Mathis, D.; Butz, E.A. Cytokine requirements for acute and Basal homeostatic proliferation of naive and memory CD8+ T cells. J. Exp. Med. 2002, 195, 1515–1522. [Google Scholar] [CrossRef]
- Herndler-Brandstetter, D.; Ishigame, H.; Shinnakasu, R.; Plajer, V.; Stecher, C.; Zhao, J.; Lietzenmayer, M.; Kroehling, L.; Takumi, A.; Kometani, K.; et al. KLRG1+ Effector CD8+ T Cells Lose KLRG1, Differentiate into All Memory T Cell Lineages, and Convey Enhanced Protective Immunity. Immunity 2018, 48, 716–729 e718. [Google Scholar] [CrossRef] [Green Version]
- Holz, L.E.; Prier, J.E.; Freestone, D.; Steiner, T.M.; English, K.; Johnson, D.N.; Mollard, V.; Cozijnsen, A.; Davey, G.M.; Godfrey, D.I.; et al. CD8+ T Cell Activation Leads to Constitutive Formation of Liver Tissue-Resident Memory T Cells that Seed a Large and Flexible Niche in the Liver. Cell Rep. 2018, 25, 68–79e64. [Google Scholar] [CrossRef] [Green Version]
- Mackay, L.K.; Rahimpour, A.; Ma, J.Z.; Collins, N.; Stock, A.T.; Hafon, M.L.; Vega-Ramos, J.; Lauzurica, P.; Mueller, S.N.; Stefanovic, T.; et al. The developmental pathway for CD103+ CD8+ tissue-resident memory T cells of skin. Nat. Immunol. 2013, 14, 1294–1301. [Google Scholar] [CrossRef]
- Verbist, K.C.; Field, M.B.; Klonowski, K.D. Cutting edge: IL-15-independent maintenance of mucosally generated memory CD8 T cells. J. Immunol. 2011, 186, 6667–6671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenkel, J.M.; Fraser, K.A.; Casey, K.A.; Beura, L.K.; Pauken, K.E.; Vezys, V.; Masopust, D. IL-15-Independent Maintenance of Tissue-Resident and Boosted Effector Memory CD8 T Cells. J. Immunol. 2016, 196, 3920–3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, J.; Stohlman, S.A.; Parra, G.I.; Bergmann, C.C. IL-15 independent maintenance of virus-specific CD8+ T cells in the CNS during chronic infection. J. Neuroimmunol. 2009, 207, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Masopust, D.; Vezys, V.; Marzo, A.L.; Lefrancois, L. Preferential localization of effector memory cells in nonlymphoid tissue. Science 2001, 291, 2413–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Joe, G.; Hexner, E.; Zhu, J.; Emerson, S.G. Host-reactive CD8+ memory stem cells in graft-versus-host disease. Nat. Med. 2005, 11, 1299–1305. [Google Scholar] [CrossRef]
- Gattinoni, L.; Lugli, E.; Ji, Y.; Pos, Z.; Paulos, C.M.; Quigley, M.F.; Almeida, J.R.; Gostick, E.; Yu, Z.; Carpenito, C.; et al. A human memory T cell subset with stem cell-like properties. Nat. Med. 2011, 17, 1290–1297. [Google Scholar] [CrossRef]
- Gattinoni, L.; Zhong, X.S.; Palmer, D.C.; Ji, Y.; Hinrichs, C.S.; Yu, Z.; Wrzesinski, C.; Boni, A.; Cassard, L.; Garvin, L.M.; et al. Wnt signaling arrests effector T cell differentiation and generates CD8+ memory stem cells. Nat. Med. 2009, 15, 808–813. [Google Scholar] [CrossRef]
- Casey, K.A.; Fraser, K.A.; Schenkel, J.M.; Moran, A.; Abt, M.C.; Beura, L.K.; Lucas, P.J.; Artis, D.; Wherry, E.J.; Hogquist, K.; et al. Antigen-independent differentiation and maintenance of effector-like resident memory T cells in tissues. J. Immunol. 2012, 188, 4866–4875. [Google Scholar] [CrossRef] [Green Version]
- Steinert, E.M.; Schenkel, J.M.; Fraser, K.A.; Beura, L.K.; Manlove, L.S.; Igyarto, B.Z.; Southern, P.J.; Masopust, D. Quantifying Memory CD8 T Cells Reveals Regionalization of Immunosurveillance. Cell 2015, 161, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Sathaliyawala, T.; Kubota, M.; Yudanin, N.; Turner, D.; Camp, P.; Thome, J.J.; Bickham, K.L.; Lerner, H.; Goldstein, M.; Sykes, M.; et al. Distribution and compartmentalization of human circulating and tissue-resident memory T cell subsets. Immunity 2013, 38, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, R.; Beura, L.K.; Quarnstrom, C.F.; Ghoneim, H.E.; Fan, Y.; Zebley, C.C.; Scott, M.C.; Fares-Frederickson, N.J.; Wijeyesinghe, S.; Thompson, E.A.; et al. Developmental plasticity allows outside-in immune responses by resident memory T cells. Nat. Immunol. 2020, 21, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Stolley, J.M.; Johnston, T.S.; Soerens, A.G.; Beura, L.K.; Rosato, P.C.; Joag, V.; Wijeyesinghe, S.P.; Langlois, R.A.; Osum, K.C.; Mitchell, J.S.; et al. Retrograde migration supplies resident memory T cells to lung-draining LN after influenza infection. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Park, S.L.; Zaid, A.; Hor, J.L.; Christo, S.N.; Prier, J.E.; Davies, B.; Alexandre, Y.O.; Gregory, J.L.; Russell, T.A.; Gebhardt, T.; et al. Local proliferation maintains a stable pool of tissue-resident memory T cells after antiviral recall responses. Nat. Immunol. 2018, 19, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Shiow, L.R.; Rosen, D.B.; Brdickova, N.; Xu, Y.; An, J.; Lanier, L.L.; Cyster, J.G.; Matloubian, M. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature 2006, 440, 540–544. [Google Scholar] [CrossRef]
- Hombrink, P.; Helbig, C.; Backer, R.A.; Piet, B.; Oja, A.E.; Stark, R.; Brasser, G.; Jongejan, A.; Jonkers, R.E.; Nota, B.; et al. Programs for the persistence, vigilance and control of human CD8+ lung-resident memory T cells. Nat. Immunol. 2016, 17, 1467–1478. [Google Scholar] [CrossRef]
- Kumar, B.V.; Ma, W.; Miron, M.; Granot, T.; Guyer, R.S.; Carpenter, D.J.; Senda, T.; Sun, X.; Ho, S.H.; Lerner, H.; et al. Human Tissue-Resident Memory T Cells Are Defined by Core Transcriptional and Functional Signatures in Lymphoid and Mucosal Sites. Cell Rep. 2017, 20, 2921–2934. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.A.; Borges da Silva, H.; Beura, L.K.; Peng, C.; Hamilton, S.E.; Masopust, D.; Jameson, S.C. The Functional Requirement for CD69 in Establishment of Resident Memory CD8+ T Cells Varies with Tissue Location. J. Immunol. 2019, 203, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Woodberry, T.; Suscovich, T.J.; Henry, L.M.; August, M.; Waring, M.T.; Kaur, A.; Hess, C.; Kutok, J.L.; Aster, J.C.; Wang, F.; et al. Alpha E beta 7 (CD103) expression identifies a highly active, tonsil-resident effector-memory CTL population. J. Immunol. 2005, 175, 4355–4362. [Google Scholar] [CrossRef] [Green Version]
- Takamura, S.; Yagi, H.; Hakata, Y.; Motozono, C.; McMaster, S.R.; Masumoto, T.; Fujisawa, M.; Chikaishi, T.; Komeda, J.; Itoh, J.; et al. Specific niches for lung-resident memory CD8+ T cells at the site of tissue regeneration enable CD69-independent maintenance. J. Exp. Med. 2016, 213, 3057–3073. [Google Scholar] [CrossRef]
- Bergsbaken, T.; Bevan, M.J. Proinflammatory microenvironments within the intestine regulate the differentiation of tissue-resident CD8+ T cells responding to infection. Nat. Immunol. 2015, 16, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, J.M.; Fraser, K.A.; Masopust, D. Cutting edge: Resident memory CD8 T cells occupy frontline niches in secondary lymphoid organs. J. Immunol. 2014, 192, 2961–2964. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Mishra, S.; Demel, E.L.; Liu, Y.; Zhang, N. TGF-beta Controls the Formation of Kidney-Resident T Cells via Promoting Effector T Cell Extravasation. J. Immunol. 2017, 198, 749–756. [Google Scholar] [CrossRef] [Green Version]
- McNamara, H.A.; Cai, Y.; Wagle, M.V.; Sontani, Y.; Roots, C.M.; Miosge, L.A.; O’Connor, J.H.; Sutton, H.J.; Ganusov, V.V.; Heath, W.R.; et al. Up-regulation of LFA-1 allows liver-resident memory T cells to patrol and remain in the hepatic sinusoids. Sci. Immunol. 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thome, J.J.; Yudanin, N.; Ohmura, Y.; Kubota, M.; Grinshpun, B.; Sathaliyawala, T.; Kato, T.; Lerner, H.; Shen, Y.; Farber, D.L. Spatial map of human T cell compartmentalization and maintenance over decades of life. Cell 2014, 159, 814–828. [Google Scholar] [CrossRef] [Green Version]
- Bartolome-Casado, R.; Landsverk, O.J.B.; Chauhan, S.K.; Richter, L.; Phung, D.; Greiff, V.; Risnes, L.F.; Yao, Y.; Neumann, R.S.; Yaqub, S.; et al. Resident memory CD8 T cells persist for years in human small intestine. J. Exp. Med. 2019, 216, 2412–2426. [Google Scholar] [CrossRef] [PubMed]
- Pallett, L.J.; Davies, J.; Colbeck, E.J.; Robertson, F.; Hansi, N.; Easom, N.J.W.; Burton, A.R.; Stegmann, K.A.; Schurich, A.; Swadling, L.; et al. IL-2(high) tissue-resident T cells in the human liver: Sentinels for hepatotropic infection. J. Exp. Med. 2017, 214, 1567–1580. [Google Scholar] [CrossRef]
- Ray, S.J.; Franki, S.N.; Pierce, R.H.; Dimitrova, S.; Koteliansky, V.; Sprague, A.G.; Doherty, P.C.; de Fougerolles, A.R.; Topham, D.J. The collagen binding alpha1beta1 integrin VLA-1 regulates CD8 T cell-mediated immune protection against heterologous influenza infection. Immunity 2004, 20, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Snyder, M.E.; Finlayson, M.O.; Connors, T.J.; Dogra, P.; Senda, T.; Bush, E.; Carpenter, D.; Marboe, C.; Benvenuto, L.; Shah, L.; et al. Generation and persistence of human tissue-resident memory T cells in lung transplantation. Sci. Immunol. 2019, 4. [Google Scholar] [CrossRef]
- Smolders, J.; Heutinck, K.M.; Fransen, N.L.; Remmerswaal, E.B.M.; Hombrink, P.; Ten Berge, I.J.M.; van Lier, R.A.W.; Huitinga, I.; Hamann, J. Tissue-resident memory T cells populate the human brain. Nat. Commun. 2018, 9, 4593. [Google Scholar] [CrossRef] [Green Version]
- Park S, P.J.; Kim, E.; Lee, Y. The Capicua/ETS Translocation Variant 5 Axis Regulates Liver-Resident Memory CD8+ T-Cell Development and the Pathogenesis of Liver Injury. Hepatology 2019, 70, 358–371. [Google Scholar] [CrossRef] [PubMed]
- Reilly, E.C.; Lambert Emo, K.; Buckley, P.M.; Reilly, N.S.; Smith, I.; Chaves, F.A.; Yang, H.; Oakes, P.W.; Topham, D.J. TRM integrins CD103 and CD49a differentially support adherence and motility after resolution of influenza virus infection. Proc. Natl. Acad Sci. USA 2020, 117, 12306–12314. [Google Scholar] [CrossRef] [PubMed]
- Cheuk, S.; Schlums, H.; Gallais Serezal, I.; Martini, E.; Chiang, S.C.; Marquardt, N.; Gibbs, A.; Detlofsson, E.; Introini, A.; Forkel, M.; et al. CD49a Expression Defines Tissue-Resident CD8+ T Cells Poised for Cytotoxic Function in Human Skin. Immunity 2017, 46, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Tse, S.W.; Radtke, A.J.; Espinosa, D.A.; Cockburn, I.A.; Zavala, F. The chemokine receptor CXCR6 is required for the maintenance of liver memory CD8+ T cells specific for infectious pathogens. J. Infect Dis. 2014, 210, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Wein, A.N.; McMaster, S.R.; Takamura, S.; Dunbar, P.R.; Cartwright, E.K.; Hayward, S.L.; McManus, D.T.; Shimaoka, T.; Ueha, S.; Tsukui, T.; et al. CXCR6 regulates localization of tissue-resident memory CD8 T cells to the airways. J. Exp. Med. 2019, 216, 2748–2762. [Google Scholar] [CrossRef] [Green Version]
- Weisberg, S.P.; Carpenter, D.J.; Chait, M.; Dogra, P.; Gartrell-Corrado, R.D.; Chen, A.X.; Campbell, S.; Liu, W.; Saraf, P.; Snyder, M.E.; et al. Tissue-Resident Memory T Cells Mediate Immune Homeostasis in the Human Pancreas through the PD-1/PD-L1 Pathway. Cell Rep. 2019, 29, 3916–3932 e3915. [Google Scholar] [CrossRef] [Green Version]
- Abdelsamed, H.A.; Frost, E.L.; Schmitz, H.M.; Mockus, T.E.; Youngblood, B.A.; Lukacher, A.E. Maintenance of PD-1 on brain-resident memory CD8 T cells is antigen independent. Immunol. Cell Biol. 2017, 95, 953–959. [Google Scholar] [CrossRef]
- Skon, C.N.; Lee, J.Y.; Anderson, K.G.; Masopust, D.; Hogquist, K.A.; Jameson, S.C. Transcriptional downregulation of S1pr1 is required for the establishment of resident memory CD8+ T cells. Nat. Immunol. 2013, 14, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Cox, M.A.; Kahan, S.M.; Ingram, J.T.; Bakshi, R.K.; Zajac, A.J. A Context-Dependent Role for IL-21 in Modulating the Differentiation, Distribution, and Abundance of Effector and Memory CD8 T Cell Subsets. J. Immunol. 2016, 196, 2153–2166. [Google Scholar] [CrossRef] [Green Version]
- Ren, H.M.; Kolawole, E.M.; Ren, M.; Jin, G.; Netherby-Winslow, C.S.; Wade, Q.; Rahman, Z.S.; Evavold, B.D.; Lukacher, A.E. IL-21 from high-affinity CD4 T cells drives differentiation of brain-resident CD8 T cells during persistent viral infection. Sci. Immunol. 2020, 5. [Google Scholar] [CrossRef]
- Nath, A.P.; Braun, A.; Ritchie, S.C.; Carbone, F.R.; Mackay, L.K.; Gebhardt, T.; Inouye, M. Comparative analysis reveals a role for TGF-beta in shaping the residency-related transcriptional signature in tissue-resident memory CD8+ T cells. PLoS ONE 2019, 14, e0210495. [Google Scholar] [CrossRef] [PubMed]
- Mackay, L.K.; Minnich, M.; Kragten, N.A.; Liao, Y.; Nota, B.; Seillet, C.; Zaid, A.; Man, K.; Preston, S.; Freestone, D.; et al. Hobit and Blimp1 instruct a universal transcriptional program of tissue residency in lymphocytes. Science 2016, 352, 459–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stelma, F.; de Niet, A.; Sinnige, M.J.; van Dort, K.A.; van Gisbergen, K.; Verheij, J.; van Leeuwen, E.M.M.; Kootstra, N.A.; Reesink, H.W. Human intrahepatic CD69+ CD8+ T cells have a tissue resident memory T cell phenotype with reduced cytolytic capacity. Sci. Rep. 2017, 7, 6172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milner, J.J.; Toma, C.; He, Z.; Kurd, N.S.; Nguyen, Q.P.; McDonald, B.; Quezada, L.; Widjaja, C.E.; Witherden, D.A.; Crowl, J.T.; et al. Heterogenous Populations of Tissue-Resident CD8+ T Cells Are Generated in Response to Infection and Malignancy. Immunity 2020, 52, 808–824 e807. [Google Scholar] [CrossRef]
- Sheridan, B.S.; Pham, Q.M.; Lee, Y.T.; Cauley, L.S.; Puddington, L.; Lefrancois, L. Oral infection drives a distinct population of intestinal resident memory CD8+ T cells with enhanced protective function. Immunity 2014, 40, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Tomov, V.T.; Palko, O.; Lau, C.W.; Pattekar, A.; Sun, Y.; Tacheva, R.; Bengsch, B.; Manne, S.; Cosma, G.L.; Eisenlohr, L.C.; et al. Differentiation and Protective Capacity of Virus-Specific CD8+ T Cells Suggest Murine Norovirus Persistence in an Immune-Privileged Enteric Niche. Immunity 2017, 47, 723–738 e725. [Google Scholar] [CrossRef] [Green Version]
- Zuber, J.; Shonts, B.; Lau, S.P.; Obradovic, A.; Fu, J.; Yang, S.; Lambert, M.; Coley, S.; Weiner, J.; Thome, J.; et al. Bidirectional intragraft alloreactivity drives the repopulation of human intestinal allografts and correlates with clinical outcome. Sci. Immunol. 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Zaid, A.; Hor, J.L.; Christo, S.N.; Groom, J.R.; Heath, W.R.; Mackay, L.K.; Mueller, S.N. Chemokine Receptor-Dependent Control of Skin Tissue-Resident Memory T Cell Formation. J. Immunol. 2017, 199, 2451–2459. [Google Scholar] [CrossRef] [Green Version]
- Boddupalli, C.S.; Bar, N.; Kadaveru, K.; Krauthammer, M.; Pornputtapong, N.; Mai, Z.; Ariyan, S.; Narayan, D.; Kluger, H.; Deng, Y.; et al. Interlesional diversity of T cell receptors in melanoma with immune checkpoints enriched in tissue-resident memory T cells. JCI Insight 2016, 1, e88955. [Google Scholar] [CrossRef]
- 8Wu, T.; Hu, Y.; Lee, Y.T.; Bouchard, K.R.; Benechet, A.; Khanna, K.; Cauley, L.S. Lung-resident memory CD8 T cells (TRM) are indispensable for optimal cross-protection against pulmonary virus infection. J. Leukoc Biol. 2014, 95, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Beura, L.K.; Mitchell, J.S.; Thompson, E.A.; Schenkel, J.M.; Mohammed, J.; Wijeyesinghe, S.; Fonseca, R.; Burbach, B.J.; Hickman, H.D.; Vezys, V.; et al. Intravital mucosal imaging of CD8+ resident memory T cells shows tissue-autonomous recall responses that amplify secondary memory. Nat. Immunol. 2018, 19, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Moylan, D.C.; Goepfert, P.A.; Kempf, M.C.; Saag, M.S.; Richter, H.E.; Mestecky, J.; Sabbaj, S. Diminished CD103 (alphaEbeta7) Expression on Resident T Cells from the Female Genital Tract of HIV-Positive Women. Pathog. Immun. 2016, 1, 371–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Garcia, M.; Fortier, J.M.; Barr, F.D.; Wira, C.R. Aging impacts CD103+ CD8+ T cell presence and induction by dendritic cells in the genital tract. Aging Cell 2018, 17, e12733. [Google Scholar] [CrossRef] [Green Version]
- Thom, J.T.; Weber, T.C.; Walton, S.M.; Torti, N.; Oxenius, A. The Salivary Gland Acts as a Sink for Tissue-Resident Memory CD8+ T Cells, Facilitating Protection from Local Cytomegalovirus Infection. Cell Rep. 2015, 13, 1125–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beura, L.K.; Wijeyesinghe, S.; Thompson, E.A.; Macchietto, M.G.; Rosato, P.C.; Pierson, M.J.; Schenkel, J.M.; Mitchell, J.S.; Vezys, V.; Fife, B.T.; et al. T Cells in Nonlymphoid Tissues Give Rise to Lymph-Node-Resident Memory T Cells. Immunity 2018, 48, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Woon, H.G.; Braun, A.; Li, J.; Smith, C.; Edwards, J.; Sierro, F.; Feng, C.G.; Khanna, R.; Elliot, M.; Bell, A.; et al. Compartmentalization of Total and Virus-Specific Tissue-Resident Memory CD8+ T Cells in Human Lymphoid Organs. PLoS Pathog. 2016, 12, e1005799. [Google Scholar] [CrossRef]
- de Leur, K.; Dieterich, M.; Hesselink, D.A.; Corneth, O.B.J.; Dor, F.; de Graav, G.N.; Peeters, A.M.A.; Mulder, A.; Kimenai, H.; Claas, F.H.J.; et al. Characterization of donor and recipient CD8+ tissue-resident memory T cells in transplant nephrectomies. Sci. Rep. 2019, 9, 5984. [Google Scholar] [CrossRef]
- Wakim, L.M.; Woodward-Davis, A.; Bevan, M.J. Memory T cells persisting within the brain after local infection show functional adaptations to their tissue of residence. Proc. Natl. Acad. Sci. USA 2010, 107, 17872–17879. [Google Scholar] [CrossRef] [Green Version]
- Prasad, S.; Hu, S.; Sheng, W.S.; Chauhan, P.; Singh, A.; Lokensgard, J.R. The PD-1: PD-L1 pathway promotes development of brain-resident memory T cells following acute viral encephalitis. J. Neuroinflammation 2017, 14, 82. [Google Scholar] [CrossRef]
- Jiang, X.; Clark, R.A.; Liu, L.; Wagers, A.J.; Fuhlbrigge, R.C.; Kupper, T.S. Skin infection generates non-migratory memory CD8+ T(RM) cells providing global skin immunity. Nature 2012, 483, 227–231. [Google Scholar] [CrossRef]
- Park, S.L.; Gebhardt, T.; Mackay, L.K. Tissue-Resident Memory T Cells in Cancer Immunosurveillance. Trends Immunol. 2019, 40, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Hu, S.; Sheng, W.S.; Singh, A.; Lokensgard, J.R. Tregs Modulate Lymphocyte Proliferation, Activation, and Resident-Memory T-Cell Accumulation within the Brain during MCMV Infection. PLoS ONE 2015, 10, e0145457. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, K.; Vincenti, I.; Kreutzfeldt, M.; Page, N.; Muschaweckh, A.; Wagner, I.; Drexler, I.; Pinschewer, D.; Korn, T.; Merkler, D. Brain-resident memory T cells represent an autonomous cytotoxic barrier to viral infection. J. Exp. Med. 2016, 213, 1571–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackay, L.K.; Wynne-Jones, E.; Freestone, D.; Pellicci, D.G.; Mielke, L.A.; Newman, D.M.; Braun, A.; Masson, F.; Kallies, A.; Belz, G.T.; et al. T-box Transcription Factors Combine with the Cytokines TGF-beta and IL-15 to Control Tissue-Resident Memory T Cell Fate. Immunity 2015, 43, 1101–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenkel, J.M.; Fraser, K.A.; Vezys, V.; Masopust, D. Sensing and alarm function of resident memory CD8+ T cells. Nat. Immunol. 2013, 14, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Ariotti, S.; Hogenbirk, M.A.; Dijkgraaf, F.E.; Visser, L.L.; Hoekstra, M.E.; Song, J.Y.; Jacobs, H.; Haanen, J.B.; Schumacher, T.N. T cell memory. Skin-resident memory CD8+ T cells trigger a state of tissue-wide pathogen alert. Science 2014, 346, 101–105. [Google Scholar] [CrossRef]
- Bogdanos, D.P.; Gao, B.; Gershwin, M.E. Liver immunology. Compr. Physiol. 2013, 3, 567–598. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.W.; Harmon, C.; O’Farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell. Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Guidotti, L.G.; Inverso, D.; Sironi, L.; Di Lucia, P.; Fioravanti, J.; Ganzer, L.; Fiocchi, A.; Vacca, M.; Aiolfi, R.; Sammicheli, S.; et al. Immunosurveillance of the liver by intravascular effector CD8+ T cells. Cell 2015, 161, 486–500. [Google Scholar] [CrossRef] [Green Version]
- Warren, A.; Le Couteur, D.G.; Fraser, R.; Bowen, D.G.; McCaughan, G.W.; Bertolino, P. T lymphocytes interact with hepatocytes through fenestrations in murine liver sinusoidal endothelial cells. Hepatology 2006, 44, 1182–1190. [Google Scholar] [CrossRef]
- Guebre-Xabier, M.; Schwenk, R.; Krzych, U. Memory phenotype CD8+ T cells persist in livers of mice protected against malaria by immunization with attenuated Plasmodium berghei sporozoites. Eur. J. Immunol. 1999, 29, 3978–3986. [Google Scholar] [CrossRef]
- Mackay, L.K.; Stock, A.T.; Ma, J.Z.; Jones, C.M.; Kent, S.J.; Mueller, S.N.; Heath, W.R.; Carbone, F.R.; Gebhardt, T. Long-lived epithelial immunity by tissue-resident memory T (TRM) cells in the absence of persisting local antigen presentation. Proc. Natl. Acad. Sci. USA 2012, 109, 7037–7042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallett, L.J.; Burton, A.R.; Amin, O.E.; Rodriguez-Tajes, S.; Patel, A.A.; Zakeri, N.; Jeffery-Smith, A.; Swadling, L.; Schmidt, N.M.; Baiges, A.; et al. Longevity and replenishment of human liver-resident memory T cells and mononuclear phagocytes. J. Exp. Med. 2020, 217. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, A.S.; Lyke, K.E.; DeZure, A.; Berry, A.A.; Richie, T.L.; Mendoza, F.H.; Enama, M.E.; Gordon, I.J.; Chang, L.J.; Sarwar, U.N.; et al. Corrigendum: Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat. Med. 2016, 22, 692. [Google Scholar] [CrossRef]
- Chapin, C.A.; Burn, T.; Meijome, T.; Loomes, K.M.; Melin-Aldana, H.; Kreiger, P.A.; Whitington, P.F.; Behrens, E.M.; Alonso, E.M. Indeterminate pediatric acute liver failure is uniquely characterized by a CD103+ CD8+ T-cell infiltrate. Hepatology 2018, 68, 1087–1100. [Google Scholar] [CrossRef] [Green Version]
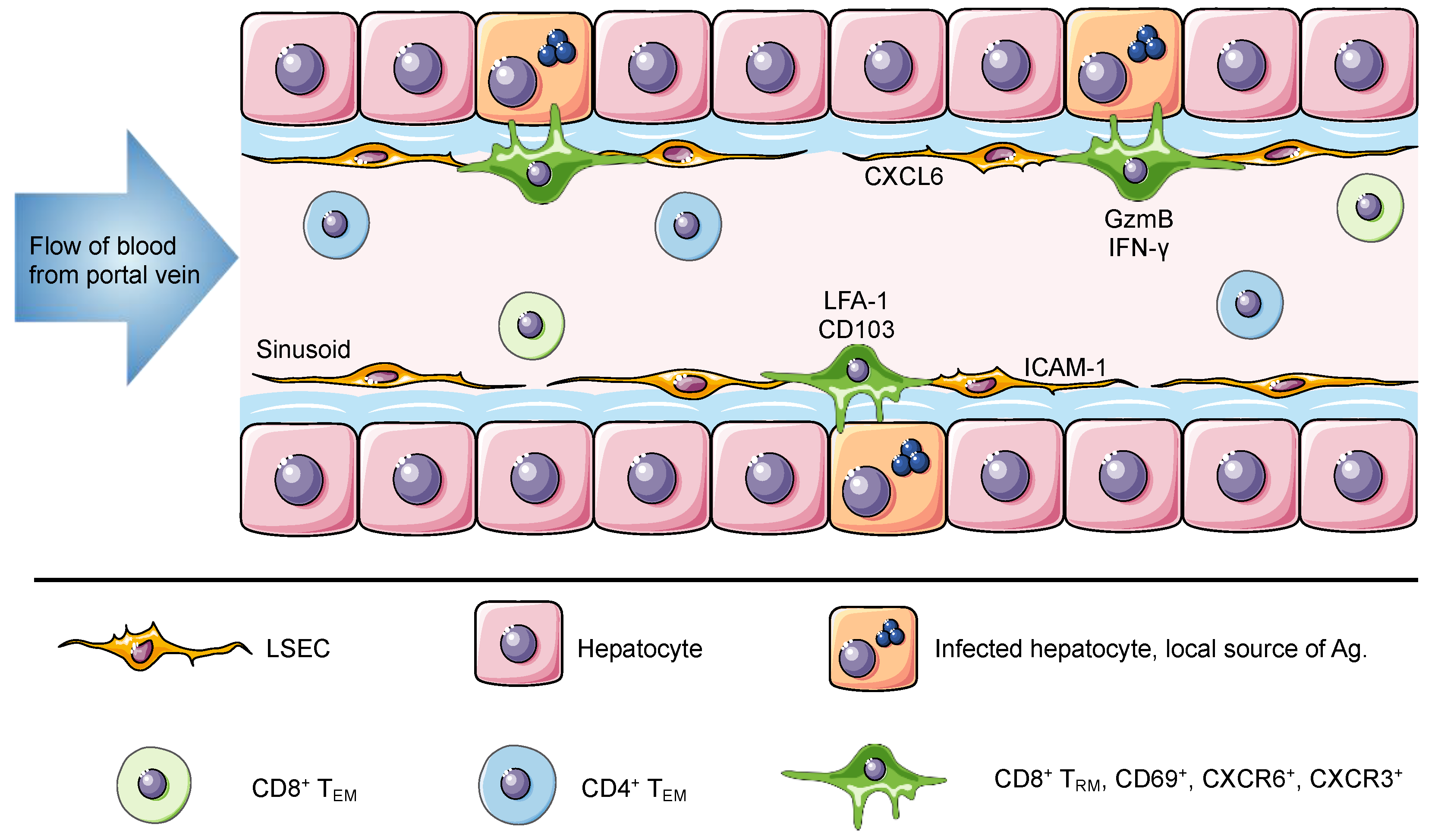
Figure 1.
The liver is a unique niche for tissue resident memory cells. The portal vein delivers antigen-rich blood from the gastrointestinal tract and spleen to the liver. This blood flows through the liver hepatic sinusoids lined with a thin layer of fenestrated liver sinusoidal endothelial cell (LSEC). Liver TRM cells are localized within the hepatic sinusoids, where they remain long-term and do not recirculate despite direct connection to the circulatory system and constant exposure to the blood. The expression of ICAM-1 and CXCL16 by LSEC can promote the retention of lymphocytes, through interactions with LFA-1 and CXCR6, respectively. Murine and human TRM cells in the liver express CD69, CXCR6, CXCR3 and high levels of LFA-1. Of note, human but not murine TRM cells express CD103. It has been suggested that this difference is associated with a broad versus a restricted expression of E-cadherin by human and murine hepatocytes, respectively. Intrahepatic lymphocytes including circulating and resident memory cells can access the surface of hepatocytes through LSEC fenestrae and exert effector functions. Using cytoplasmic protrusions, lymphocytes probe hepatocytes for the presence of antigen and can release factors such as GzmB and IFN-γ to promote hepatocyte killing. In murine studies, liver TRM cells can be generated through different vaccination strategies to confer protection against Plasmodium parasites and in humans they have been associated with disease control against HBV and HCV.
Figure 1.
The liver is a unique niche for tissue resident memory cells. The portal vein delivers antigen-rich blood from the gastrointestinal tract and spleen to the liver. This blood flows through the liver hepatic sinusoids lined with a thin layer of fenestrated liver sinusoidal endothelial cell (LSEC). Liver TRM cells are localized within the hepatic sinusoids, where they remain long-term and do not recirculate despite direct connection to the circulatory system and constant exposure to the blood. The expression of ICAM-1 and CXCL16 by LSEC can promote the retention of lymphocytes, through interactions with LFA-1 and CXCR6, respectively. Murine and human TRM cells in the liver express CD69, CXCR6, CXCR3 and high levels of LFA-1. Of note, human but not murine TRM cells express CD103. It has been suggested that this difference is associated with a broad versus a restricted expression of E-cadherin by human and murine hepatocytes, respectively. Intrahepatic lymphocytes including circulating and resident memory cells can access the surface of hepatocytes through LSEC fenestrae and exert effector functions. Using cytoplasmic protrusions, lymphocytes probe hepatocytes for the presence of antigen and can release factors such as GzmB and IFN-γ to promote hepatocyte killing. In murine studies, liver TRM cells can be generated through different vaccination strategies to confer protection against Plasmodium parasites and in humans they have been associated with disease control against HBV and HCV.


{kind=link}
Table 1.
Expression of the canonical markers used to define CD8+ TRM cells in diverse murine and human organs.
Table 1.
Expression of the canonical markers used to define CD8+ TRM cells in diverse murine and human organs.
| Organs | Expression of Canonical Markers (CD69, CD103, CD49a and CXCR6) | |||
|---|---|---|---|---|
| Mice | Humans | |||
| Intestine, Gut | CD69+ CD103+/− CD49a+ CXCR6+ | [40,41,52,76,77] | CD69+ CD103+ | [64,78] |
| Skin | CD69+ CD103+/− CD49a+ CXCR6+ | [31,79] | CD69+ CD103+/− CD49a+/− | [64,80] |
| Lungs | CD69+ CD103+ CD49a+ CXCR6+/- | [59,66,81] | CD69+ CD103+ CD49a+ CXCR6+ | [47,48] |
| Female reproductive tract | CD69+/− CD103+/− | [40,41,82] | CD69+ CD103+ (transcriptomic profiling is yet to be determined) | [83,84] |
| Salivary glands | CD69+/− CD103+/− CD49a+ | [41,85] | CD69+ CD103+/− | [48] |
| Lymphoid organs (Spleen, lymph nodes, tonsil) | CD69+ CD103− CD49a+ | [53,86] | CD69+ CD103+/− CD49a- | [87] |
| Liver | CD69+ CD103− CD49a+ CXCR6+ | [6,62,73] | CD69+ CD103+/− CXCR6+ | [58,74] |
| Kidneys | CD69+/− CD103- | [40,41,54] | CD69+ CD103+/− CD49a+/− CXCR6+/− | [88] |
| Pancreas | CD69+/− CD103+/− | [40,41] | CD69+ CD103+ CD49a+ CXCR6+ | [67] |
| Brain | CD69+ CD103+/− | [40,68,89,90] | CD69+ CD103+/− CD49a+ CXCR6+/− | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ghilas, S.; Valencia-Hernandez, A.-M.; Enders, M.H.; Heath, W.R.; Fernandez-Ruiz, D. Resident Memory T Cells and Their Role within the Liver. Int. J. Mol. Sci. 2020, 21, 8565. https://doi.org/10.3390/ijms21228565
AMA Style
Ghilas S, Valencia-Hernandez A-M, Enders MH, Heath WR, Fernandez-Ruiz D. Resident Memory T Cells and Their Role within the Liver. International Journal of Molecular Sciences. 2020; 21(22):8565. https://doi.org/10.3390/ijms21228565
Chicago/Turabian StyleGhilas, Sonia, Ana-Maria Valencia-Hernandez, Matthias H. Enders, William R. Heath, and Daniel Fernandez-Ruiz. 2020. "Resident Memory T Cells and Their Role within the Liver" International Journal of Molecular Sciences 21, no. 22: 8565. https://doi.org/10.3390/ijms21228565
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.