Preparative Scale Production of Recombinant Human Transthyretin for Biophysical Studies of Protein-Ligand and Protein-Protein Interactions


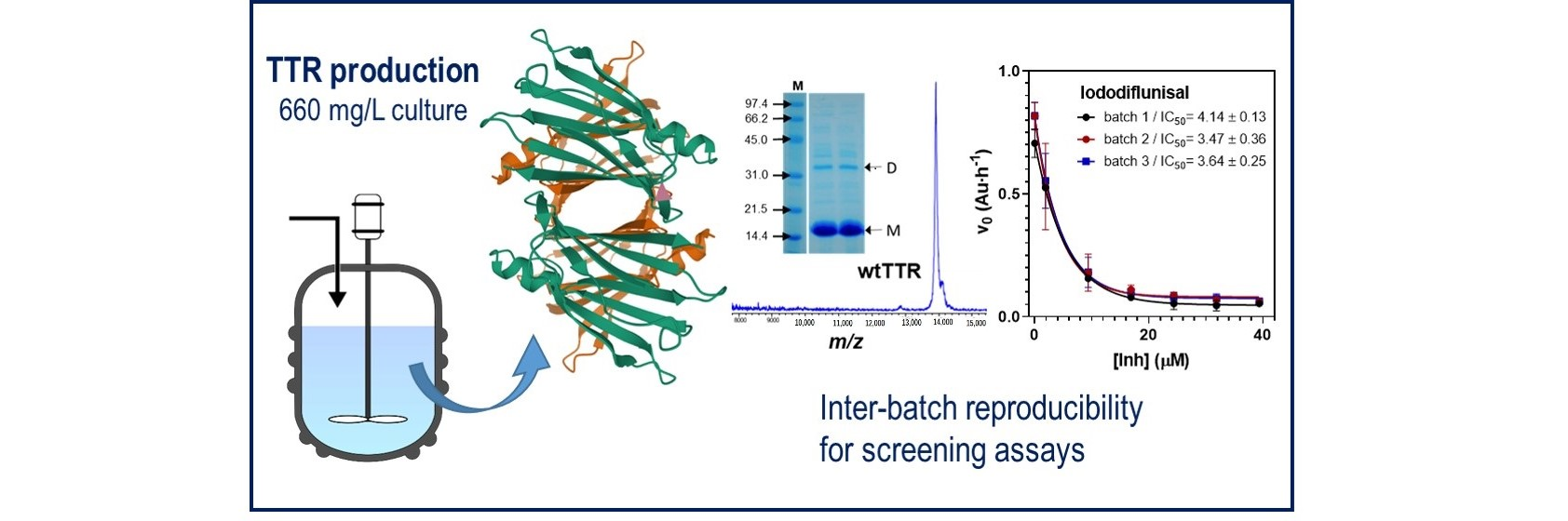
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. Culture Medium and Induction Conditions
2.2. Production in Fed-Batch Bioreactor
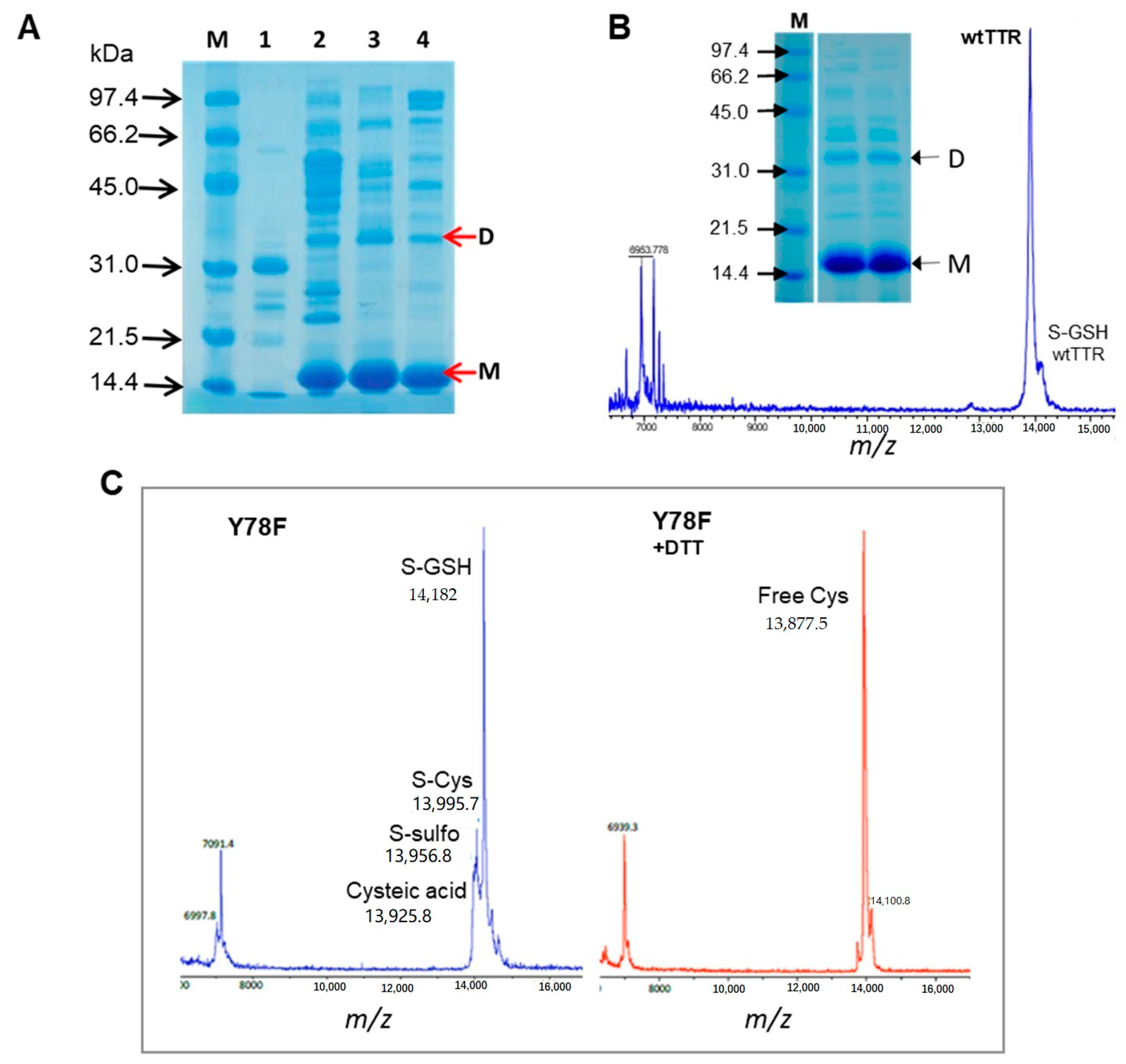
2.3. Protein Purification
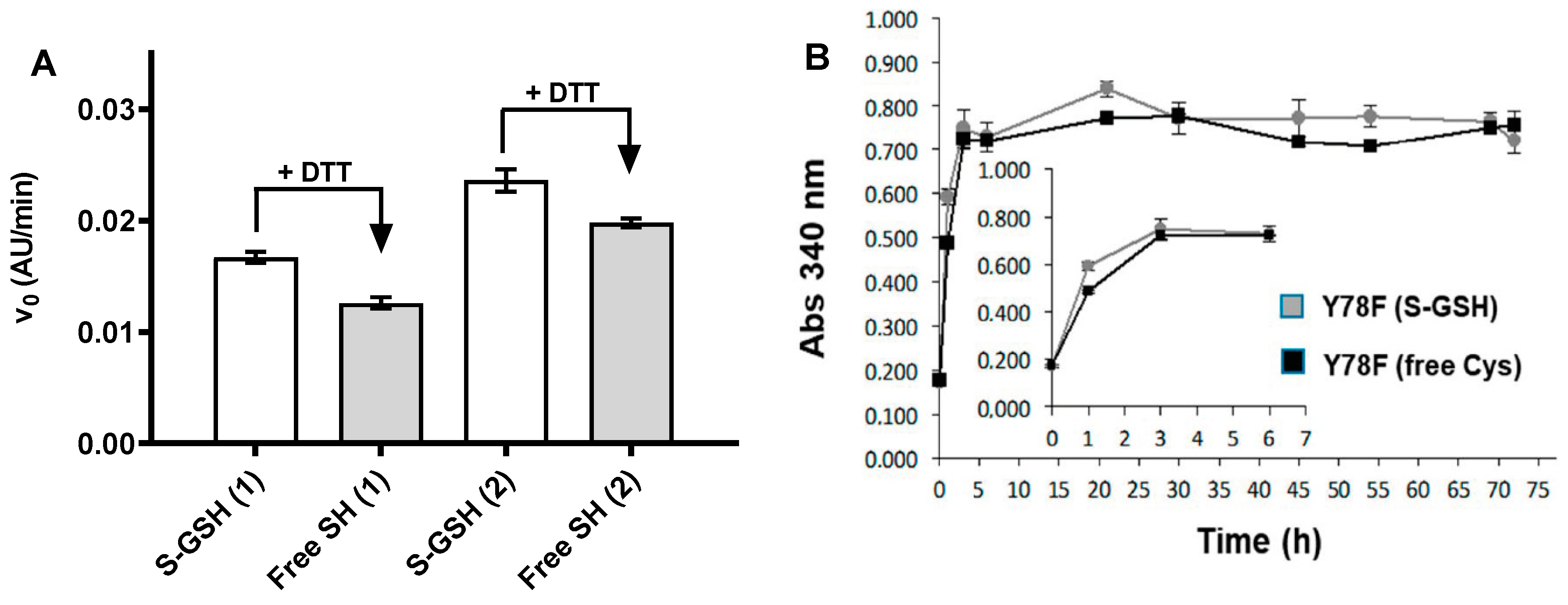
2.4. Protein Quality Assessment
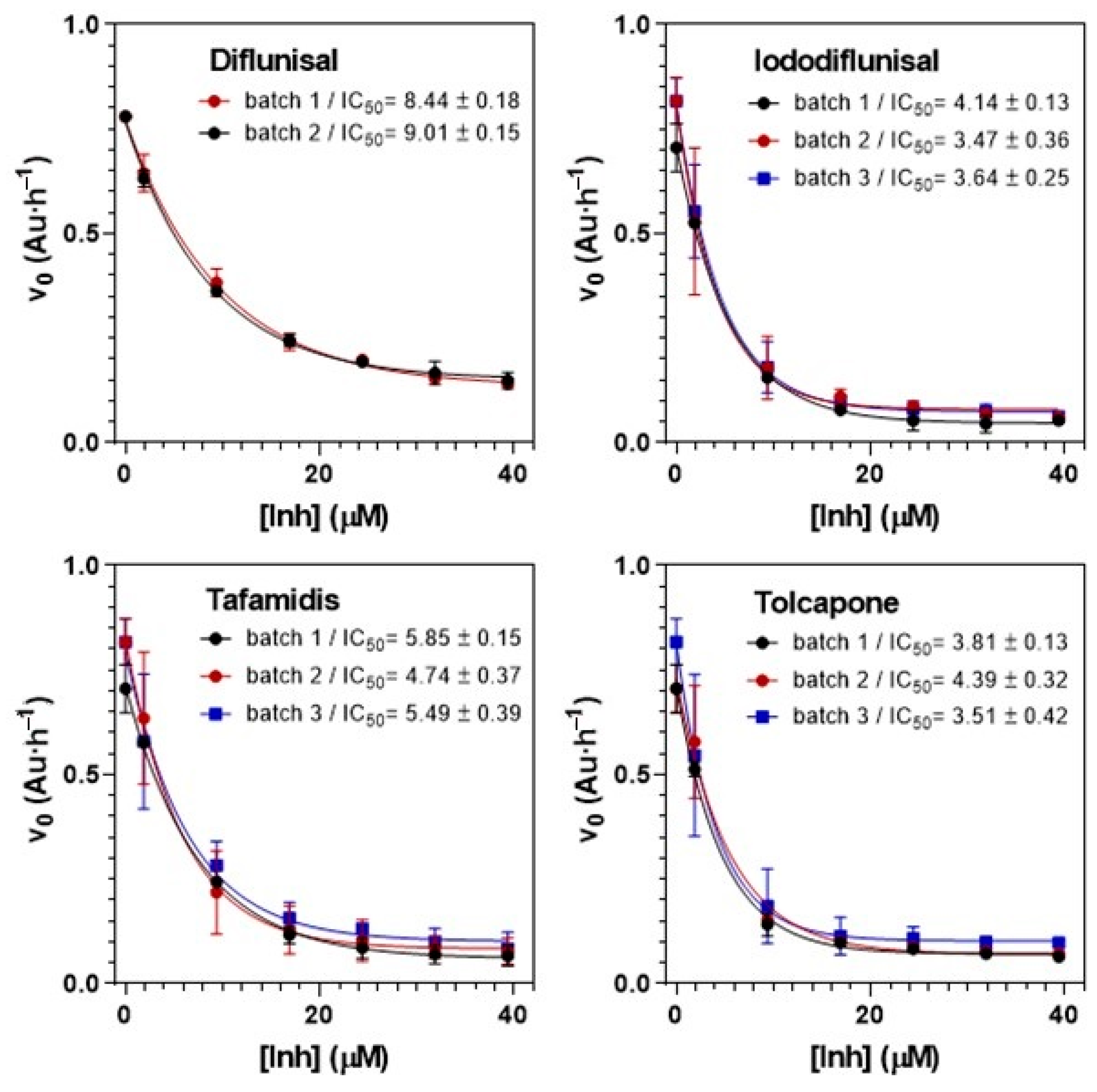
2.5. Applications to Screening Assays
2.5.1. Kinetic Turbidimetry Assay for Screening TTR-Amyloidogenesis Inhibitors
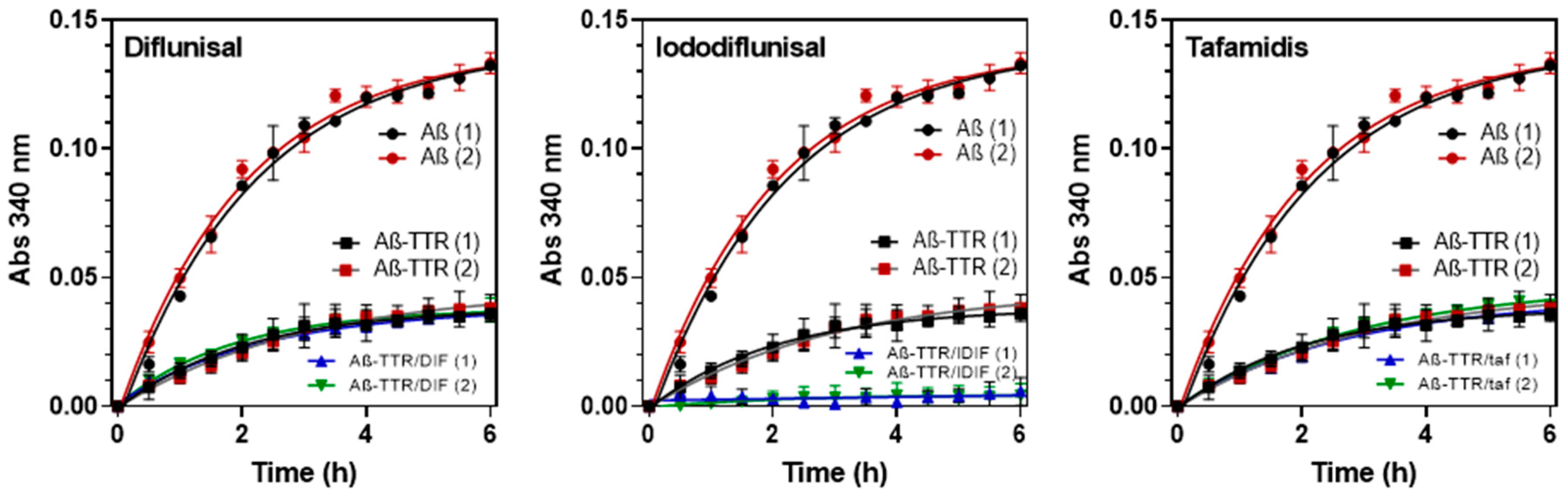
2.5.2. Turbidity-Based Assay for Screening Small-Molecule Chaperones Enhancing the TTR:Aβ Interaction
3. Materials and Methods
3.1. Protein Expression
3.1.1. Batch Expression in Flask Cultures
3.1.2. Expression in Fed-Batch Bioreactor
3.2. Protein Purification
3.3. Reduction with 1,4-Dithiothreitol (DTT)
3.4. MALDI-TOF Mass Spectrometry of Purified Proteins
3.5. Acid-Induced TTR Fibrillogenesis Assay
3.6. Kinetic Turbidimetry Assay for Screening TTR Fibrillogenesis Inhibitors
3.7. Turbidimetry Assay for Screening Small-Molecule Chaperones of TTR:Aβ Interaction
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Manning, M.C.; Chou, D.K.; Murphy, B.M.; Payne, R.W.; Katayama, D.S. Stability of Protein Pharmaceuticals: An Update. Pharm. Res. 2010, 27, 544–575. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Garcia, L.; Martín, L.; Mangues, R.; Ferrer-Miralles, N.; Vazquez, E.; Villaverde, A. Recombinant pharmaceuticals from microbial cells: A 2015 update. Microb. Cell Factories 2016, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef] [PubMed]
- Blake, C.; Geisow, M.; Oatley, S.; Rérat, B.; Rérat, C. Structure of prealbumin: Secondary, tertiary and quaternary interactions determined by Fourier refinement at 1.8 Å. J. Mol. Biol. 1978, 121, 339–356. [Google Scholar] [CrossRef]
- Hamilton, J.; Benson, M. Transthyretin: A review from a structural perspective. Cell. Mol. Life Sci. 2001, 58, 1491–1521. [Google Scholar] [CrossRef]
- Andrade, C. A Peculiar Form of Peripheral Neuropathy. Brain 1952, 75, 408–427. [Google Scholar] [CrossRef]
- Saraiva, M.J.; Magalhaes, J.; Ferreira, N.; Almeida, M.R. Transthyretin Deposition in Familial Amyloidotic Polyneuropathy. Curr. Med. Chem. 2012, 19, 2304–2311. [Google Scholar] [CrossRef] [PubMed]
- Koike, H.; Katsuno, M. Transthyretin Amyloidosis: Update on the Clinical Spectrum, Pathogenesis, and Disease-Modifying Therapies. Neurol. Ther. 2020, 9, 317–333. [Google Scholar] [CrossRef]
- Grodin, J.L.; Maurer, M.S. The Truth Is Unfolding About Transthyretin Cardiac Amyloidosis. Circulation 2019, 140, 27–30. [Google Scholar] [CrossRef]
- Sekijima, Y.; Hammarström, P.; Matsumura, M.; Shimizu, Y.; Iwata, M.; Tokuda, T.; Ikeda, S.-I.; Kelly, J.W. Energetic Characteristics of the New Transthyretin Variant A25T May Explain Its Atypical Central Nervous System Pathology. Lab. Investig. 2003, 83, 409–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, M.R.; Gales, L.; Damas, A.M.; Cardoso, I.; Saraiva, M.J. Small Transthyretin (TTR) Ligands as Possible Therapeutic Agents in TTR Amyloidoses. Curr. Drug Target CNS Neurol. Disord. 2005, 4, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Connelly, S.; Choi, S.; Johnson, S.M.; Kelly, J.W.; Wilson, I.A. Structure-based design of kinetic stabilizers that ameliorate the transthyretin amyloidoses. Curr. Opin. Struct. Biol. 2010, 20, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevone, A.; Merlini, G.; Nuvolone, M. Treating Protein Misfolding Diseases: Therapeutic Successes Against Systemic Amyloidoses. Front. Pharmacol. 2020, 11, 1024. [Google Scholar] [CrossRef]
- Bulawa, C.; Connelly, S.; DeVit, M.; Wang, L.; Weigel, C.; Fleming, J.A.; Packman, J.; Powers, E.T.; Wiseman, R.L.; Foss, T.R.; et al. Tafamidis, a potent and selective transthyretin kinetic stabilizer that inhibits the amyloid cascade. Proc. Natl. Acad. Sci. USA 2012, 109, 9629–9634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berk, J.L.; Suhr, O.B.; Obici, L.; Sekijima, Y.; Zeldenrust, S.R.; Yamashita, T.; Heneghan, M.A.; Gorevic, P.D.; Litchy, W.J.; Wiesman, J.F.; et al. Repurposing Diflunisal for Familial Amyloid Polyneuropathy: A randomized clinical trial. JAMA 2013, 310, 2658–2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wixner, J.; Westermark, P.; Ihse, E.; Pilebro, B.; Lundgren, H.-E.; Anan, I. The Swedish open-label diflunisal trial (DFNS01) on hereditary transthyretin amyloidosis and the impact of amyloid fibril composition. Amyloid 2019, 26, 39–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, T.; Mizuguchi, M. Transthyretin Amyloidogenesis Inhibitors: From Discovery to Current Developments. J. Med. Chem. 2020. [Google Scholar] [CrossRef]
- Cruz, M.W.; Benson, M.D. A Review of Tafamidis for the Treatment of Transthyretin-Related Amyloidosis. Neurol. Ther. 2015, 4, 61–79. [Google Scholar] [CrossRef] [Green Version]
- Maurer, M.S.; Schwartz, J.H.; Gundapaneni, B.; Elliott, P.M.; Merlini, G.; Waddington-Cruz, M.; Kristen, A.V.; Grogan, M.; Witteles, R.; Damy, T.; et al. Tafamidis Treatment for Patients with Transthyretin Amyloid Cardiomyopathy. N. Engl. J. Med. 2018, 379, 1007–1016. [Google Scholar] [CrossRef]
- Park, J.; Egolum, U.; Parker, S.; Andrews, E.; Ombengi, D.; Ling, H. Tafamidis: A First-in-Class Transthyretin Stabilizer for Transthyretin Amyloid Cardiomyopathy. Ann. Pharmacother. 2019, 54, 470–477. [Google Scholar] [CrossRef]
- Gamez, J.; Salvadó, M.; Reig, N.; Suñé, P.; Casasnovas, C.; Rojas-García, R.; Boronat, R.I. Transthyretin stabilization activity of the catechol-O-methyltransferase inhibitor tolcapone (SOM0226) in hereditary ATTR amyloidosis patients and asymptomatic carriers: Proof-of-concept study. Amyloid 2019, 26, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Sant’Anna, R.; Gallego, P.; Robinson, L.Z.; Pereira-Henriques, A.; Ferreira, N.; Pinheiro, F.; Esperante, S.; Pallares, I.; Huertas, O.; Almeida, M.R.; et al. Repositioning tolcapone as a potent inhibitor of transthyretin amyloidogenesis and associated cellular toxicity. Nat. Commun. 2016, 7, 10787. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, F.; Varejão, N.; Esperante, S.; Santos, J.; Velazquez-Campoy, A.; Reverter, D.; Pallarès, I.; Ventura, S. Tolcapone, a potent aggregation inhibitor for the treatment of familial leptomeningeal amyloidosis. FEBS J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Pal, A.; Albusairi, W.; Joo, H.; Pappas, B.; Tuhin, T.H.; Liang, D.; Jampala, R.; Liu, F.; Khan, J.; et al. Enthalpy-Driven Stabilization of Transthyretin by AG10 Mimics a Naturally Occurring Genetic Variant That Protects from Transthyretin Amyloidosis. J. Med. Chem. 2018, 61, 7862–7876. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.C.; Hellawell, J.L.; Rao, S.; O’Reilly, T.; Lumpkin, R.; Jernelius, J.; Gretler, D.; Sinha, U. First-in-Human Study of AG10, a Novel, Oral, Specific, Selective, and Potent Transthyretin Stabilizer for the Treatment of Transthyretin Amyloidosis: A Phase 1 Safety, Tolerability, Pharmacokinetic, and Pharmacodynamic Study in Healthy Adult Volunteers. Clin. Pharmacol. Drug Dev. 2020, 9, 115–129. [Google Scholar] [CrossRef] [PubMed]
- Corazza, A.; Verona, G.; Waudby, C.A.; Mangione, P.P.; Bingham, R.; Uings, I.; Canetti, D.; Nocerino, P.; Taylor, G.W.; Pepys, M.B.; et al. Binding of Monovalent and Bivalent Ligands by Transthyretin Causes Different Short- and Long-Distance Conformational Changes. J. Med. Chem. 2019, 62, 8274–8283. [Google Scholar] [CrossRef] [Green Version]
- 2020 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2020, 16, 391–460. [CrossRef]
- dos Santos, P.; Leide, C.; Ozela, P.F.; de Fatima de Brito Brito, M.; Pinheiro, A.A.; Padilha, E.C.; Braga, F.S.; de Paula, D.S.; Carlos, H.T.; dos Santos, C.B.R.; et al. Alzheimer’s Disease: A Review from the Pathophysiology to Diagnosis, New Perspectives for Pharmacological Treatment. Curr. Med. Chem. 2018, 25, 3141–3159. [Google Scholar] [CrossRef]
- DeTure, M.A.; Dickson, D.W. The neuropathological diagnosis of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Forner, S.; Baglietto-Vargas, D.; Martini, A.C.; Trujillo-Estrada, L.; LaFerla, F.M. Synaptic Impairment in Alzheimer’s Disease: A Dysregulated Symphony. Trends Neurosci. 2017, 40, 347–357. [Google Scholar] [CrossRef]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Pate, K.M.; Murphy, R.M. Cerebrospinal Fluid Proteins as Regulators of Beta-amyloid Aggregation and Toxicity. Isr. J. Chem. 2017, 57, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Schwarzman, A.L.; Gregori, L.; Vitek, M.P.; Lyubski, S.; Strittmatter, W.J.; Enghilde, J.J.; Bhasin, R.; Silverman, J.; Weisgraber, K.H.; Coyle, P.K. Transthyretin sequesters amyloid beta protein and prevents amyloid formation. Proc. Natl. Acad. Sci. USA 1994, 91, 8368–8372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, X.; Ladiwala, A.R.A.; Du, D.; Yadav, J.K.; Tessier, P.M.; Wright, P.E.; Kelly, J.W.; Buxbaum, J.N. Mechanisms of Transthyretin Inhibition of -Amyloid Aggregation In Vitro. J. Neurosci. 2013, 33, 19423–19433. [Google Scholar] [CrossRef]
- Alemi, M.; Silva, S.C.; Santana, I.; Cardoso, I. Transthyretin stability is critical in assisting beta amyloid clearance- Relevance of transthyretin stabilization in Alzheimer’s disease. CNS Neurosci. Ther. 2017, 23, 605–619. [Google Scholar] [CrossRef] [Green Version]
- Cotrina, E.Y.; Gimeno, A.; Llop, J.; Jiménez-Barbero, J.; Quintana, J.; Prohens, R.; Cardoso, I.; Arsequell, G. An Assay for Screening Potential Drug Candidates for Alzheimer’s Disease That Act as Chaperones of the Transthyretin and Amyloid-β Peptides Interaction. Chemistry 2020, 21. [Google Scholar] [CrossRef]
- Furuya, H.; Nakazato, M.; Saraiva, M.J.; Costa, P.P.; Sasaki, H.; Matsuo, H.; Goto, I.; Sakaki, Y. Tetramer formation of a variant type human transthyretin (prealbumin) produced by Escherichia coli expression system. Biochem. Biophys. Res. Commun. 1989, 163, 851–859. [Google Scholar] [CrossRef]
- Furuya, H.; Saraiva, M.J.M.; Gawinowicz, M.A.; Alves, I.L.; Costa, P.P.; Sasaki, H.; Goto, I.; Sakaki, Y. Production of recombinant human transthyretin with biological activities toward the understanding of the molecular basis of familial amyloidotic polyneuropathy (FAP). Biochemistry 1991, 30, 2415–2421. [Google Scholar] [CrossRef]
- Matsubara, K.; Mizuguchi, M.; Kawano, K. Expression of a synthetic gene encoding human transthyretin in Escherichia coli. Protein Expr. Purif. 2003, 30, 55–61. [Google Scholar] [CrossRef]
- Kingsbury, J.S.; Klimtchuk, E.S.; Theberge, R.; Costello, C.E.; Connors, L.H. Expression, purification, and in vitro cysteine-10 modification of native sequence recombinant human transthyretin. Protein Expr. Purif. 2007, 53, 370–377. [Google Scholar] [CrossRef] [Green Version]
- Dolado, I.; Nieto, J.; Saraiva, M.J.M.; Arsequell, G.; Valencia, G.; Planas, A. Kinetic Assay for High-Throughput Screening of In Vitro Transthyretin Amyloid Fibrillogenesis Inhibitors. J. Comb. Chem. 2005, 7, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Vidal, L.; Durany, O.; Suau, T.; Ferrer, P.; Benaiges, M.D.; Caminal, G. High-level production of recombinant His-tagged rhamnulose 1-phosphate aldolase in Escherichia coli. J. Chem. Technol. Biotechnol. 2003, 78, 1171–1179. [Google Scholar] [CrossRef]
- Vilà-Rico, M.; Colomé-Calls, N.; Martín-Castel, L.; Gay, M.; Azorín, S.; Vilaseca, M.; Planas, A.; Canals, F. Quantitative analysis of post-translational modifications in human serum transthyretin associated with familial amyloidotic polyneuropathy by targeted LC–MS and intact protein MS. J. Proteom. 2015, 127, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, K.; Bahl, J.M.; Tanassi, J.T.; Simonsen, A.H.; Heegaard, N.H. Characterization and stability of transthyretin isoforms in cerebrospinal fluid examined by immunoprecipitation and high-resolution mass spectrometry of intact protein. Methods 2012, 56, 284–292. [Google Scholar] [CrossRef] [PubMed]
- Pont, L.; Benavente, F.; Barbosa, J.; Sanz-Nebot, V. Analysis of transthyretin in human serum by capillary zone electrophoresis electrospray ionization time-of-flight mass spectrometry. Application to familial amyloidotic polyneuropathy type I. Electrophoresis 2015, 36, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Arsequell, G.; Planas, A. Methods to evaluate the inhibition of TTR fibrillogenesis induced by small ligands. Curr. Med. Chem. 2012, 19, 2343–2355. [Google Scholar] [CrossRef]
- Mairal, T.; Nieto, J.; Pinto, M.; Almeida, M.R.; Gales, L.; Ballesteros, A.; Barluenga, J.; Pérez, J.J.; Vázquez, J.T.; Centeno, N.B.; et al. Iodine Atoms: A New Molecular Feature for the Design of Potent Transthyretin Fibrillogenesis Inhibitors. PLoS ONE 2009, 4, e4124. [Google Scholar] [CrossRef] [Green Version]
- Cotrina, E.Y.; Pinto, M.; Bosch, L.; Vilà, M.; Blasi, D.; Quintana, J.; Centeno, N.B.; Arsequell, G.; Planas, A.; Valencia, G. Modulation of the Fibrillogenesis Inhibition Properties of Two Transthyretin Ligands by Halogenation. J. Med. Chem. 2013, 56, 9110–9121. [Google Scholar] [CrossRef]
- Vilaró, M.; Nieto, J.; La Parra, J.R.; Almeida, M.R.; Ballesteros, A.; Planas, A.; Arsequell, G.; Valencia, G. Tuning Transthyretin Amyloidosis Inhibition Properties of Iododiflunisal by Combinatorial Engineering of the Nonsalicylic Ring Substitutions. ACS Comb. Sci. 2014, 17, 32–38. [Google Scholar] [CrossRef]
- Cotrina, E.Y.; Blasi, D.; Vilà, M.; Planas, A.; Abad-Zapatero, C.; Centeno, N.B.; Quintana, J.; Arsequell, G. Optimization of kinetic stabilizers of tetrameric transthyretin: A prospective ligand efficiency-guided approach. Bioorganic Med. Chem. 2020, 28, 115794. [Google Scholar] [CrossRef]
- Gonzalez, A.; Quirante, J.; Nieto, J.; Almeida, M.R.; Saraiva, M.J.; Planas, A.; Arsequell, G.; Valencia, G. Isatin derivatives, a novel class of transthyretin fibrillogenesis inhibitors. Bioorganic Med. Chem. Lett. 2009, 19, 5270–5273. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cotrina, E.Y.; Vilà, M.; Nieto, J.; Arsequell, G.; Planas, A. Preparative Scale Production of Recombinant Human Transthyretin for Biophysical Studies of Protein-Ligand and Protein-Protein Interactions. Int. J. Mol. Sci. 2020, 21, 9640. https://doi.org/10.3390/ijms21249640
Cotrina EY, Vilà M, Nieto J, Arsequell G, Planas A. Preparative Scale Production of Recombinant Human Transthyretin for Biophysical Studies of Protein-Ligand and Protein-Protein Interactions. International Journal of Molecular Sciences. 2020; 21(24):9640. https://doi.org/10.3390/ijms21249640
Chicago/Turabian StyleCotrina, Ellen Y., Marta Vilà, Joan Nieto, Gemma Arsequell, and Antoni Planas. 2020. "Preparative Scale Production of Recombinant Human Transthyretin for Biophysical Studies of Protein-Ligand and Protein-Protein Interactions" International Journal of Molecular Sciences 21, no. 24: 9640. https://doi.org/10.3390/ijms21249640