Distinct Preflowering Drought Tolerance Strategies of Sorghum bicolor Genotype RTx430 Revealed by Subcellular Protein Profiling

 ,
,
Abstract
:1. Introduction
2. Results
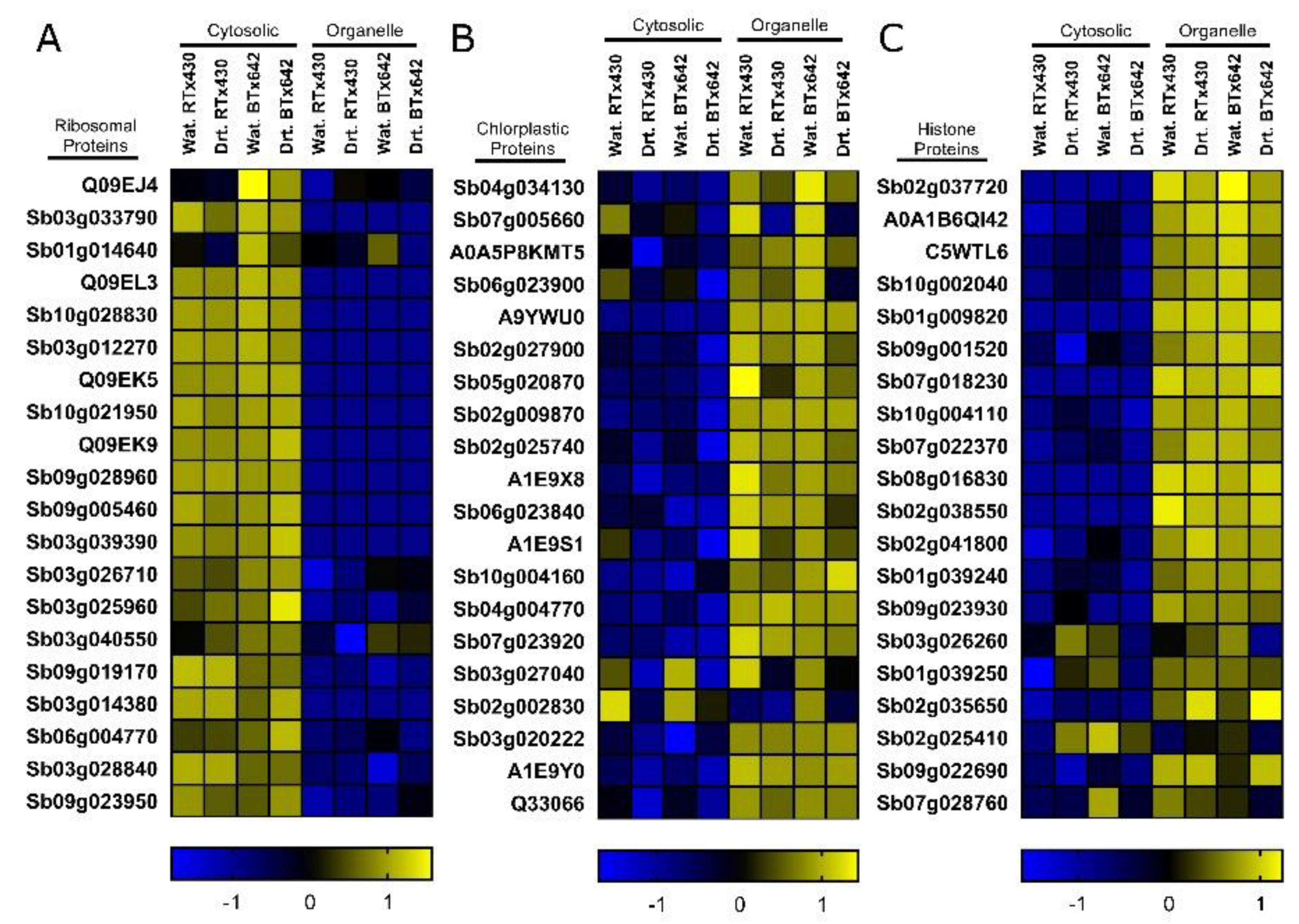
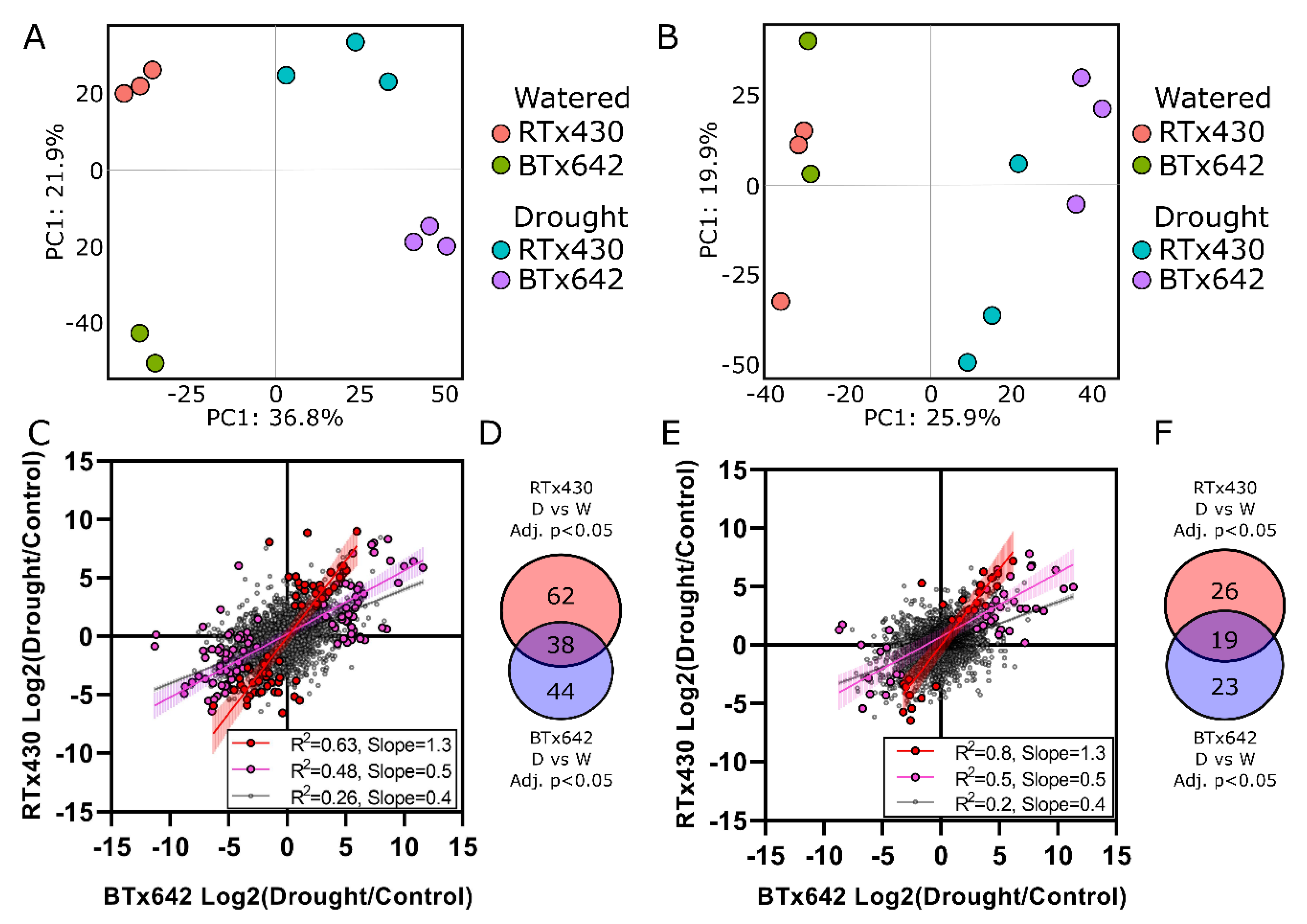
2.1. Protein Profiles Are Enriched for Organelle and Cytoplasmic Compartments
2.2. Organelle-Enriched Protein Profiles Comprise Genotype-Specific Drought Signatures
2.3. Protein Profiles in Response to Drought Support Previous Findings
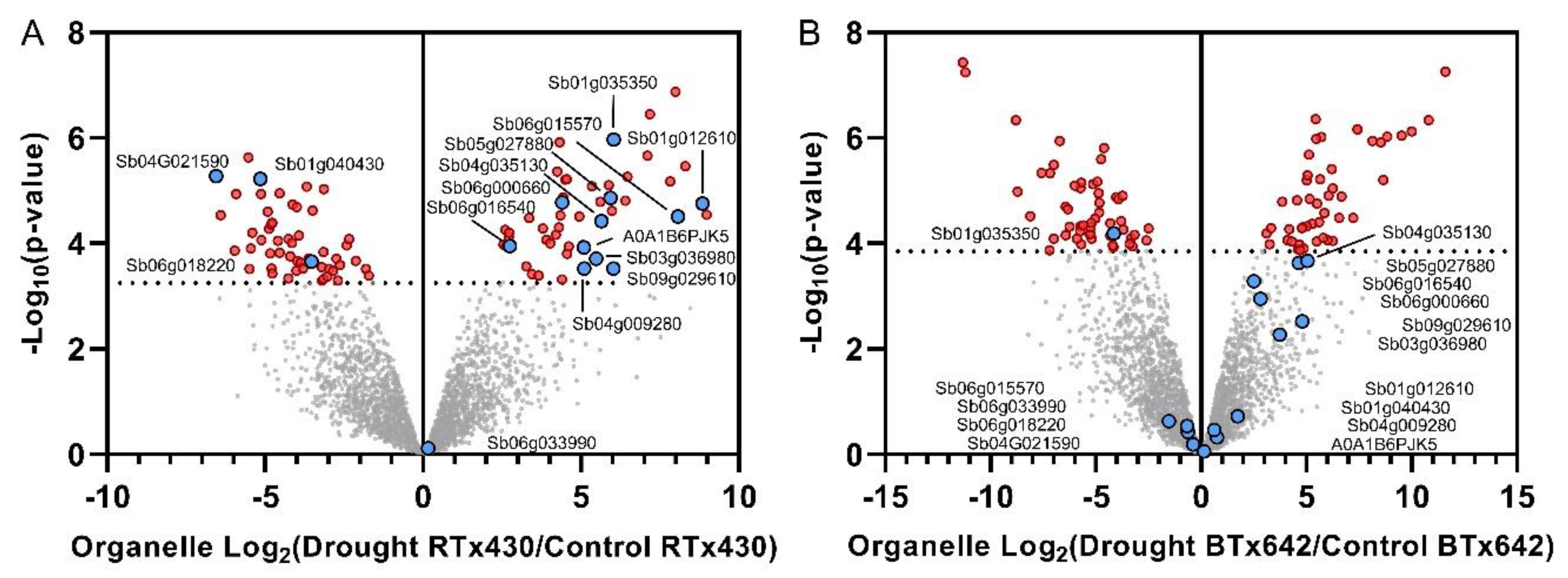
2.4. Genotype-Specific Drought Responses Involved in Flowering Time Control, Starch Biosynthesis, and Rubisco Activation May Explain RTx430′s Drought Tolerance
2.5. Genotype-Specific Drought Responses Involved in Abscisic Acid Signaling May Explain RTx430′s Drought Tolerance
2.6. Other RTx430-Specific Preflowering Drought Responses Include Proteins Involved in ROS Scavenging, HSPs, Epicuticular Wax Production, and Phospholipid Metabolism
3. Discussion
4. Materials and Methods
4.1. Plant Growth and Drought Stress
4.2. Organelle and Cytosolic Enrichment
4.3. Protein Digestion
4.4. LC-MS/MS Analysis
4.5. Protein Search and Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DEP | Differentially Expressed Proteins |
| ABA | Abscisic Acid |
| GO | Gene Ontology |
| FLC | FLOWERING LOCUS C |
| RCA | Rubisco Activase |
| ASR | Abscisic Acid Stress-Ripening |
| sHSP | Small Heat Shock Protein |
| ZEP | Zeaxanthin Epoxidase |
| SOD | Superoxide Dismutase |
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Dbattisti, D.S.; Naylor, R.L. Historical Warnings of Future Food Insecurity with Unprecedented Seasonal Heat. Science 2009, 323, 240–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwayama, Y.; Thompson, A.; Bernknopf, R.; Zaitchik, B.; Vail, P. Estimating the Impact of Drought on Agriculture Using the U.S. Drought Monitor. Am. J. Agric. Econ. 2018, 101, 193–210. [Google Scholar] [CrossRef]
- United Nations, Department of Economic and Social Affairs. World Population Prospects Highlights; 2019; ISBN 978-92-1-148316-1. Available online: https://www.un.org/development/desa/publications/world-population-prospects-2019-highlights.html (accessed on 15 October 2020).
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000Research 2016, 5, 1554. [Google Scholar] [CrossRef]
- Okamoto, M.; Peterson, F.C.; DeFries, A.; Park, S.-Y.; Endo, A.; Nambara, E.; Volkman, B.F.; Cutler, S.R. Activation of dimeric ABA receptors elicits guard cell closure, ABA-regulated gene expression, and drought tolerance. Proc. Natl. Acad. Sci. USA 2013, 110, 12132–12137. [Google Scholar] [CrossRef] [Green Version]
- Fàbregas, N.; Lozano-Elena, F.; Blasco-Escámez, D.; Tohge, T.; Martínez-Andújar, C.; Albacete, A.; Osorio, S.; Bustamante, M.; Riechmann, J.L.; Nomura, T.; et al. Overexpression of the vascular brassinosteroid receptor BRL3 confers drought resistance without penalizing plant growth. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.; Inzé, D.; Nelissen, H.; Saibo, N.J.M. Source–Sink Regulation in Crops under Water Deficit. Trends Plant Sci. 2019, 24, 652–663. [Google Scholar] [CrossRef]
- Fracasso, A.; Trindade, L.M.; Amaducci, S. Drought stress tolerance strategies revealed by RNA-Seq in two sorghum genotypes with contrasting WUE. BMC Plant Biol. 2016, 16, 1–18. [Google Scholar] [CrossRef]
- Ananda, G.K.S.; Myrans, H.; Norton, S.L.; Gleadow, R.; Furtado, A.; Henry, R.J. Wild Sorghum as a Promising Resource for Crop Improvement. Front. Plant Sci. 2020, 11, 1108. [Google Scholar] [CrossRef]
- Kebede, H.; Subudhi, P.K.; Rosenow, D.T.; Nguyen, H.T. Quantitative trait loci influencing drought tolerance in grain sorghum (Sorghum bicolor L. Moench). Theor. Appl. Genet. 2001, 103, 266–276. [Google Scholar] [CrossRef]
- Sabadin, P.K.; Malosetti, M.; Boer, M.P.; Tardin, F.D.; Santos, F.G.; Guimaraes, C.T.; Gomide, R.L.; Andrade, C.L.T.; Albuquerque, P.E.P.; Caniato, F.F.; et al. Studying the genetic basis of drought tolerance in sorghum by managed stress trials and adjustments for phenological and plant height differences. Theor. Appl. Genet. 2012, 124, 1389–1402. [Google Scholar] [CrossRef] [PubMed]
- Ogbaga, C.C.; Stępień, P.; Dyson, B.C.; Rattray, N.J.W.; Ellis, D.I.; Goodacre, R.; Johnson, G.N. Biochemical Analyses of Sorghum Varieties Reveal Differential Responses to Drought. PLoS ONE 2016, 11, e0154423. [Google Scholar] [CrossRef] [PubMed]
- Jedmowski, C.; Ashoub, A.; Beckhaus, T.; Berberich, T.; Karas, M.; Brüggemann, W. Comparative Analysis of Sorghum bicolor Proteome in Response to Drought Stress and following Recovery. Available online: https://www.hindawi.com/journals/ijpro/2014/395905/ (accessed on 28 September 2020).
- Fadoul, H.E.; El Siddig, M.A.; Abdalla, A.W.H.; Hussien, A.A. Physiological and proteomic analysis of two contrasting Sorghum bicolor genotypes in response to drought stress. Aust. J. Crop. Sci. 2018, 12, 1543–1551. [Google Scholar] [CrossRef]
- Thomas, H.; Howarth, C.J. Five ways to stay green. J. Exp. Bot. 2000, 51, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.H.; Bhaskaran, S.; Miller, F.R. Screening for drought tolerance in Sorghum using cell culture. Vitr. Cell. Dev. Biol. Anim. 1985, 21, 541–545. [Google Scholar] [CrossRef]
- Varoquaux, N.; Cole, B.; Gao, C.; Pierroz, G.; Baker, C.R.; Patel, D.; Madera, M.; Jeffers, T.; Hollingsworth, J.; Sievert, J.; et al. Transcriptomic analysis of field-droughted sorghum from seedling to maturity reveals biotic and metabolic responses. Proc. Natl. Acad. Sci. USA 2019, 116, 27124–27132. [Google Scholar] [CrossRef] [Green Version]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Malhan, N.; Ahkami, A.H.; Engbrecht, K.; Myers, G.; Dahlberg, J.; Hollingsworth, J.; Sievert, J.A.; Hutmacher, R.; Madera, M.; et al. Top-down mass spectrometry of histone modifications in sorghum reveals potential epigenetic markers for drought acclimation. Methods 2019. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Zhang, X.; Smits, A.H.; van Tilburg, G.B.; Ovaa, H.; Huber, W.; Vermeulen, M. Proteome-wide identification of ubiquitin interactions using UbIA-MS. Nat. Protoc. 2018, 13, 530. [Google Scholar] [CrossRef]
- Thomas, P.D.; Muruganujan, A.; Ebert, D.; Huang, X.; Thomas, P.D. PANTHER version 14: More genomes, a new PANTHER GO-slim and improvements in enrichment analysis tools. Nucleic Acids Res. 2019, 47, D419–D426. [Google Scholar] [CrossRef]
- Nagaraju, M.; Reddy, P.S.; Kumar, S.A.; Kumar, A.; Rajasheker, G.; Rao, D.M.; Kishor, P.B.K. Genome-wide identification and transcriptional profiling of small heat shock protein gene family under diverse abiotic stress conditions in Sorghum bicolor (L.). Int. J. Biol. Macromol. 2020, 142, 822–834. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.M.; Lim, F.-L.; Finkler, A.; Fromm, H.; Slabas, A.R.; Knight, M.R. Transcriptomic analysis of Sorghum bicolor responding to combined heat and drought stress. BMC Genom. 2014, 15, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Ghany, S.E.; Ullah, F.; Ben-Hur, A.; Reddy, A.S. Transcriptome Analysis of Drought-Resistant and Drought-Sensitive Sorghum (Sorghum bicolor) Genotypes in Response to PEG-Induced Drought Stress. Int. J. Mol. Sci. 2020, 21, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.; Michaels, S.D. The Arabidopsis Paf1c Complex Component CDC73 Participates in the Modification of FLOWERING LOCUS C Chromatin. Plant Physiol. 2010, 153, 1074–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Oh, S.; Ek-Ramos, J.; Van Nocker, S. Plant Homologous to Parafibromin is a Component of the PAF1 Complex and Assists in Regulating Expression of Genes within H3K27ME3-Enriched Chromatin. Plant Physiol. 2010, 153, 821–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventriglia, T.; Kuhn, M.L.; Ruiz, M.T.; Ribeiro-Pedro, M.; Valverde, F.; Ballicora, M.A.; Preiss, J.; Romero, J.M. Two Arabidopsis ADP-Glucose Pyrophosphorylase Large Subunits (APL1 and APL2) Are Catalytic. Plant Physiol. 2008, 148, 65–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AbdelGawad, H.; Avramova, V.; Baggerman, G.; Van Raemdonck, G.; Valkenborg, D.; Van Ostade, X.; Guisez, Y.; Prinsen, E.; Asard, H.; Ende, W.V.D.; et al. Starch biosynthesis contributes to the maintenance of photosynthesis and leaf growth under drought stress in maize. Plant Cell Environ. 2020, 43, 2254–2271. [Google Scholar] [CrossRef]
- Perdomo, J.A.; Capó-Bauçà, S.; Carmo-Silva, A.E.; Galmés, J. Rubisco and Rubisco Activase Play an Important Role in the Biochemical Limitations of Photosynthesis in Rice, Wheat, and Maize under High Temperature and Water Deficit. Front. Plant Sci. 2017, 8, 490. [Google Scholar] [CrossRef] [Green Version]
- Dalal, M.; Inupakutika, M. Transcriptional regulation of ABA core signaling component genes in sorghum (Sorghum bicolor L. Moench). Mol. Breed. 2014, 34, 1517–1525. [Google Scholar] [CrossRef]
- Ma, S.; Lv, L.; Meng, C.; Zhou, C.; Fu, J.; Shen, X.; Zhang, S.C.; Li, Y. Genome-Wide Analysis of Abscisic Acid Biosynthesis, Catabolism, and Signaling in Sorghum Bicolor under Saline-Alkali Stress. Biomolecules 2019, 9, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Zhang, Z.; Gao, J.; Wang, P.; Hu, T.; Wang, Z.; Hou, Y.-J.; Wan, Y.; Liu, W.; Xie, S.; et al. Arabidopsis Duodecuple Mutant of PYL ABA Receptors Reveals PYL Repression of ABA-Independent SnRK2 Activity. Cell Rep. 2018, 23, 3340–3351.e5. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y.; et al. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.; Zhong, J.; Sun, X.; Wang, B.; Terzaghi, W.; Dai, M. The Maize ABA Receptors ZmPYL8, 9, and 12 Facilitate Plant Drought Resistance. Front. Plant Sci. 2018, 9, 422. [Google Scholar] [CrossRef]
- Shaw, A.K.; Bhardwaj, P.K.; Ghosh, S.; Roy, S.; Saha, S.; Sherpa, A.R.; Saha, S.K.; Hossain, Z. β-aminobutyric acid mediated drought stress alleviation in maize (Zea mays L.). Environ. Sci. Pollut. Res. 2015, 23, 2437–2453. [Google Scholar] [CrossRef]
- Liang, Y.; Jiang, Y.; Du, M.; Li, B.; Chen, L.; Chen, M.; Jin, D.; Wu, J. ZmASR3 from the Maize ASR Gene Family Positively Regulates Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2019, 20, 2278. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; et al. TaASR1, a transcription factor gene in wheat, confers drought stress tolerance in transgenic tobacco. Plant Cell Environ. 2013, 36, 1449–1464. [Google Scholar] [CrossRef]
- Xing, Y.; Cao, Q.; Zhang, Q.; Qin, L.; Jia, W.; Zhang, J. MKK5 Regulates High Light-Induced Gene Expression of Cu/Zn Superoxide Dismutase 1 and 2 in Arabidopsis. Plant Cell Physiol. 2013, 54, 1217–1227. [Google Scholar] [CrossRef] [Green Version]
- Azzouz-Olden, F.; Hunt, A.G.; Dinkins, R.D. Transcriptome analysis of drought-tolerant sorghum genotype SC56 in response to water stress reveals an oxidative stress defense strategy. Mol. Biol. Rep. 2020, 47, 3291–3303. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-H.; Huang, L.-F.; Hase, T.; Huang, H.-E.; Feng, T.-Y. Expression of plant ferredoxin-like protein (PFLP) enhances tolerance to heat stress in Arabidopsis thaliana. New Biotechnol. 2015, 32, 235–242. [Google Scholar] [CrossRef]
- Huang, H.-E.; Ho, M.-H.; Chang, H.; Chao, H.-Y.; Ger, M.-J. Overexpression of plant ferredoxin-like protein promotes salinity tolerance in rice (Oryza sativa). Plant Physiol. Biochem. 2020, 155, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Kneeshaw, S.; Keyani, R.; Delorme-Hinoux, V.; Imrie, L.H.; Loake, G.J.; Le Bihan, T.; Reichheld, J.-P.; Spoel, S.H. Nucleoredoxin guards against oxidative stress by protecting antioxidant enzymes. Proc. Natl. Acad. Sci. USA 2017, 114, 8414–8419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewelam, N.A.; Kazan, K.; Hüdig, M.; Maurino, V.G.; Schenk, P.M. The AtHSP17.4C1 Gene Expression Is Mediated by Diverse Signals that Link Biotic and Abiotic Stress Factors with ROS and Can Be a Useful Molecular Marker for Oxidative Stress. Int. J. Mol. Sci. 2019, 20, 3201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samakovli, D.; Tichá, T.; Vavrdová, T.; Ovečka, M.; Luptovčiak, I.; Zapletalová, V.; Kuchařová, A.; Křenek, P.; Krasylenko, Y.; Margaritopoulou, T.; et al. YODA-HSP90 Module Regulates Phosphorylation-Dependent Inactivation of SPEECHLESS to Control Stomatal Development under Acute Heat Stress in Arabidopsis. Mol. Plant 2020, 13, 612–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guan, Y.; Zhang, D.; Dong, X.; Tian, L.; Qu, L.Q. A β-Ketoacyl-CoA Synthase Is Involved in Rice Leaf Cuticular Wax Synthesis and Requires a CER2-LIKE Protein as a Cofactor. Plant Physiol. 2017, 173, 944–955. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Zhang, X.; Lu, X.; Chen, G.; Chen, Z. Molecular and Evolutionary Mechanisms of Cuticular Wax for Plant Drought Tolerance. Front. Plant Sci. 2017, 8, 621. [Google Scholar] [CrossRef]
- Dhanyalakshmi, K.H.; Soolanayakanahally, R.Y.; Rahman, T.; Tanino, K.K.; Nataraja, K.N. Leaf Cuticular Wax, a Trait for Multiple Stress Resistance in Crop Plants. Abiotic Biotic Stress Plants 2019. [Google Scholar] [CrossRef] [Green Version]
- Jordan, W.R.; Shouse, P.J.; Blum, A.; Miller, F.R.; Monk, R.L. Environmental Physiology of Sorghum. II. Epicuticular Wax Load and Cuticular Transpiration. Crop. Sci. 1984, 24, 1168–1173. [Google Scholar] [CrossRef]
- Awika, H.O.; Hays, D.B.; Mullet, J.E.; Rooney, W.L.; Weers, B.D. QTL mapping and loci dissection for leaf epicuticular wax load and canopy temperature depression and their association with QTL for staygreen in Sorghum bicolor under stress. Euphytica 2017, 213, 207. [Google Scholar] [CrossRef] [Green Version]
- Matos, A.R.; Pham-Thi, A.-T. Lipid deacylating enzymes in plants: Old activities, new genes. Plant Physiol. Biochem. 2009, 47, 491–503. [Google Scholar] [CrossRef]
- Verlotta, A.; Liberatore, M.T.; Cattivelli, L.; Trono, D. Secretory Phospholipases A2 in Durum Wheat (Triticum durum Desf.): Gene Expression, Enzymatic Activity, and Relation to Drought Stress Adaptation. Int. J. Mol. Sci. 2013, 14, 5146–5169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, L.-X.; Dong, K.; Liu, S.-Y.; Zhang, Z.; Zhang, X.-P.; Tong, X.; Zhu, F.-F.; Zou, J.-Z.; Wang, X.-B. A putative nuclear copper chaperone promotes plant immunity in Arabidopsis. J. Exp. Bot. 2020, 71, 6684–6696. [Google Scholar] [CrossRef] [PubMed]
- Shin, L.-J.; Lo, J.-C.; Yeh, K. Copper Chaperone Antioxidant Protein1 Is Essential for Copper Homeostasis. Plant Physiol. 2012, 159, 1099–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkhead, J.L.; Reynolds, K.A.G.; Abdel-Ghany, S.E.; Cohu, C.M.; Pilon, M. Copper homeostasis. New Phytol. 2009, 182, 799–816. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Huang, B.; Bonos, S.A.; Meyer, W.A. Abscisic Acid Accumulation in Relation to Drought Tolerance in Kentucky Bluegrass. HortScience 2004, 39, 1133–1137. [Google Scholar] [CrossRef]
- Ji, X.; Dong, B.; Shiran, B.; Talbot, M.J.; Edlington, J.E.; Hughes, T.; White, R.G.; Gubler, F.; Dolferus, R. Control of Abscisic Acid Catabolism and Abscisic Acid Homeostasis Is Important for Reproductive Stage Stress Tolerance in Cereals. Plant Physiol. 2011, 156, 647–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Doyle, M.R.; Amasino, R.M. PAF1-complex-mediated histone methylation of FLOWERING LOCUS C chromatin is required for the vernalization-responsive, winter-annual habit in Arabidopsis. Genes Dev. 2004, 18, 2774–2784. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Zhao, Z.; Dong, A.; Soubigou-Taconnat, L.; Renou, J.-P.; Steinmetz, A.; Shen, W.-H. Di- and Tri- but Not Monomethylation on Histone H3 Lysine 36 Marks Active Transcription of Genes Involved in Flowering Time Regulation and Other Processes in Arabidopsis thaliana. Mol. Cell. Biol. 2007, 28, 1348–1360. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Park, S.; Van Nocker, S. Genic and Global Functions for Paf1C in Chromatin Modification and Gene Expression in Arabidopsis. PLoS Genet. 2008, 4, e1000077. [Google Scholar] [CrossRef] [Green Version]
- Archie, R.P. Rubisco activase–Rubisco’s catalytic chaperone. Photosynth. Res. 2003, 75, 11–27. [Google Scholar] [CrossRef]
- Eckardt, N.A.; Snyder, C.W.; Jr, A.R.P.; Ogren, W.L. Growth and Photosynthesis under High and Low Irradiance of Arabidopsis thaliana Antisense Mutants with Reduced Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Activase Content. Plant Physiol. 1997, 113, 575–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukayama, H.; Ueguchi, C.; Nishikawa, K.; Katoh, N.; Ishikawa, C.; Masumoto, C.; Hatanaka, T.; Misoo, S. Overexpression of Rubisco Activase Decreases the Photosynthetic CO2 Assimilation Rate by Reducing Rubisco Content in Rice Leaves. Plant Cell Physiol. 2012, 53, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Law, R.; Crafts-Brandner, S.J. High Temperature Stress Increases the Expression of Wheat Leaf Ribulose-1,5-Bisphosphate Carboxylase/oxygenase Activase Protein. Arch. Biochem. Biophys. 2001, 386, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Feller, U. Drought stress and carbon assimilation in a warming climate: Reversible and irreversible impacts. J. Plant Physiol. 2016, 203, 84–94. [Google Scholar] [CrossRef] [Green Version]
- Djanaguiraman, M.; Prasad, P.V.V.; Murugan, M.; Perumal, R.; Reddy, U.K. Physiological differences among sorghum (Sorghum bicolor L. Moench) genotypes under high temperature stress. Environ. Exp. Bot. 2014, 100, 43–54. [Google Scholar] [CrossRef]
- Thalmann, M.; Santelia, D. Starch as a determinant of plant fitness under abiotic stress. New Phytol. 2017, 214, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Stitt, M.; Zeeman, S.C. Starch turnover: Pathways, regulation and role in growth. Curr. Opin. Plant Biol. 2012, 15, 282–292. [Google Scholar] [CrossRef]
- Roychoudhury, A.; Paul, S.; Basu, S. Cross-talk between abscisic acid-dependent and abscisic acid-independent pathways during abiotic stress. Plant Cell Rep. 2013, 32, 985–1006. [Google Scholar] [CrossRef]
- Yamamoto, H.; Nakayama, T.; Chichester, C. Studies on the light and dark interconversions of leaf xanthophylls. Arch. Biochem. Biophys. 1962, 97, 168–173. [Google Scholar] [CrossRef]
- Landi, P.; Sanguineti, M.; Conti, S.; Tuberosa, R. Direct and Correlated Responses to Divergent Selection for Leaf Abscisic Acid Concentration in Two Maize Populations. Crop. Sci. 2001, 41, 335–344. [Google Scholar] [CrossRef]
- Schwarz, N.; Armbruster, U.; Iven, T.; Brückle, L.; Melzer, M.; Petit-Houdenot, Y.; Jahns, P. Tissue-Specific Accumulation and Regulation of Zeaxanthin Epoxidase in Arabidopsis Reflect the Multiple Functions of the Enzyme in Plastids. Plant Cell Physiol. 2014, 56, 346–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- North, H.M.; Frey, A.; Boutin, J.-P.; Sotta, B.; Marion-Poll, A. Analysis of xanthophyll cycle gene expression during the adaptation of Arabidopsis to excess light and drought stress: Changes in RNA steady-state levels do not contribute to short-term responses. Plant Sci. 2005, 169, 115–124. [Google Scholar] [CrossRef]
- Park, S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.-F.F.; et al. Abscisic Acid Inhibits Type 2C Protein Phosphatases via the PYR/PYL Family of START Proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Szostkiewicz, I.; Korte, A.; Moes, D.; Yang, Y.; Christmann, A.; Grill, E. Regulators of PP2C Phosphatase Activity Function as Abscisic Acid Sensors. Science 2009, 324, 1064–1068. [Google Scholar] [CrossRef]
- Lee, S.C.; Lim, C.W.; Lan, W.; He, K.; Luan, S. ABA Signaling in Guard Cells Entails a Dynamic Protein–Protein Interaction Relay from the PYL-RCAR Family Receptors to Ion Channels. Mol. Plant 2013, 6, 528–538. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Wang, Z.Y.; Li, X.; Lv, T.; Liu, H.; Wang, L.; Niu, H.; Bu, Q. Characterization and Functional Analysis of Pyrabactin Resistance-Like Abscisic Acid Receptor Family in Rice. Rice 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kalifa, Y.; Perlson, E.; Gilad, A.; Konrad, Z.; Scolnik, P.A.; Bar-Zvi, D. Over-expression of the water and salt stress-regulated Asr1 gene confers an increased salt tolerance. Plant Cell Environ. 2004, 27, 1459–1468. [Google Scholar] [CrossRef]
- Yang, C.-Y.; Chen, Y.-C.; Jauh, G.Y.; Wang, C. A Lily ASR Protein Involves Abscisic Acid Signaling and Confers Drought and Salt Resistance in Arabidopsis. Plant Physiol. 2005, 139, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-J.; Lee, S.-C.; Hong, S.K.; An, K.; An, G.; Kim, S.-R. Ectopic expression of a cold-responsive OsAsr1 cDNA gives enhanced cold tolerance in transgenic rice plants. Mol. Cells 2009, 27, 449–458. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Dai, J.-R.; Feng, D.-R.; Liu, B.; Wang, H.; Wang, J.-F. Characterization of a Novel Plantain Asr Gene, MpAsr, that is Regulated in Response to Infection of Fusarium oxysporum f. sp. cubense and Abiotic Stresses. J. Integr. Plant Biol. 2010, 52, 315–323. [Google Scholar] [CrossRef]
- Yu, D.; Ranathunge, K.; Huang, H.; Pei, Z.; Franke, R.; Schreiber, L.; He, C. Wax Crystal-Sparse Leaf1 encodes a β–ketoacyl CoA synthase involved in biosynthesis of cuticular waxes on rice leaf. Planta 2008, 228, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Burow, G.B.; Franks, C.D.; Xin, Z. Genetic and Physiological Analysis of an Irradiated Bloomless Mutant (Epicuticular Wax Mutant) of Sorghum. Crop. Sci. 2008, 48, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Naylor, D.; Dong, Z.; Simmons, T.; Pierroz, G.; Hixson, K.K.; Kim, Y.-M.; Zink, E.M.; Engbrecht, K.M.; Wang, Y.; et al. Drought delays development of the sorghum root microbiome and enriches for monoderm bacteria. Proc. Natl. Acad. Sci. USA 2018, 115, E4284–E4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Geen, H.; Echipare, L.; Farnham, P.J. Using ChIP-Seq Technology to Generate High-Resolution Profiles of Histone Modifications. In Epigenetics Protocols; Tollefsbol, T.O., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; pp. 265–286. ISBN 978-1-61779-316-5. [Google Scholar]
- Nakayasu, E.S.; Nicora, C.D.; Sims, A.C.; Burnum-Johnson, K.E.; Kim, Y.-M.; Kyle, J.E.; Matzke, M.M.; Shukla, A.K.; Chu, R.K.; Schepmoes, A.A.; et al. MPLEx: A Robust and Universal Protocol for Single-Sample Integrative Proteomic, Metabolomic, and Lipidomic Analyses. mSystems 2016, 1, e00043-16. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- The UniProt Consortium UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [CrossRef]
- Smyth, G.K. Linear Models and Empirical Bayes Methods for Assessing Differential Expression in Microarray Experiments. Stat. Appl. Genet. Mol. Biol. 2004, 3, 1–25. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Identifier | Protein | Abbr. | Ens. Ortholog | Function | Ref. |
|---|---|---|---|---|---|
| Sb01g012610 | Plant Homologous to Parafribomin | CDC73/PHP | At3g22590, Zm00001d013597, Os03g0664700 | Histone methylation, FLC-regulation | [27,28] |
| Sb09g029610 | ADP-Glc pyrophosphorylase | APL2 | At1G27680, Zm00001d039131, Os05g0580000 | Starch biosynthesis | [29,30] |
| Sb05g027880 | Rubisco activase alpha isoform | RCA | At2G39730, Zm00001d048592, | Photosynthesis | [31] |
| Sb04g009280 | Pyrobactin Resistance 1-like | PYL9 | At1G01360, Zm00001d016105, Os02g0255500 | ABA Signal Transduction | [32,33,34,35,36] |
| Sb06g018220 | Zeaxanthin epoxidase | ZEP | At5G67030, Os04g0448900 | ABA metabolism | [37] |
| Sb06g016540 | Abscisic Acid Stress Ripening | ASR2 | Zm00001d003712, Os04g0423400 | ABA metabolism | [38,39] |
| Sb01g035350 | Superoxide dismutase | SOD | At1G08830, Zm00001d047479, Os03g0351500 | ROS Scavenging | [40] |
| Sb06g015570 | Ferredoxin | Fd | At2G27510, Zm00001d003797, Os04g0412200 | ROS Scavenging | [41,42,43] |
| Sb03g036980 | NRX1-like Thioredoxin domain-containing | NRX | Zm00001d012591, Os01g0794400 | Oxidative Stress | [14,44] |
| Sb04g035130 | Small heatshock protein 17 | sHSP17 | At1G54050, Zm00001d018298, Os02g0782500 | Protein Folding | [45] |
| Sb06g000660 | Heatshock protein 90 | HSP90 | At5G52640, Zm00001d024903, Os04g0107900 | Protein Folding | [46] |
| A0A1B6PJK5 | 3-ketoacyl-CoA synthase | KCS | At1G25450, Os04g0116800 | Epicuticular Wax Production | [47,48,49,50,51] |
| Sb01g040430 | Phospholipase A2 | PLA2 | Zm00001d028505 | Fatty acid and membrane integrity | [52,53] |
| Sb04G021590 | HMA domain-containing protein | HMA | At3G56240, Zm00001d016691 | Copper Ion Homeostasis | [9,54,55,56] |
| Sb06g033990 | Sucrose non-fermenting 1-related prot. Kinase 2 | SnRK2 | At1G60940, Zm00001d026690, Os04g0691100 | ABA Signal Transduction | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogden, A.J.; Abdali, S.; Engbrecht, K.M.; Zhou, M.; Handakumbura, P.P. Distinct Preflowering Drought Tolerance Strategies of Sorghum bicolor Genotype RTx430 Revealed by Subcellular Protein Profiling. Int. J. Mol. Sci. 2020, 21, 9706. https://doi.org/10.3390/ijms21249706
Ogden AJ, Abdali S, Engbrecht KM, Zhou M, Handakumbura PP. Distinct Preflowering Drought Tolerance Strategies of Sorghum bicolor Genotype RTx430 Revealed by Subcellular Protein Profiling. International Journal of Molecular Sciences. 2020; 21(24):9706. https://doi.org/10.3390/ijms21249706
Chicago/Turabian StyleOgden, Aaron J., Shadan Abdali, Kristin M. Engbrecht, Mowei Zhou, and Pubudu P. Handakumbura. 2020. "Distinct Preflowering Drought Tolerance Strategies of Sorghum bicolor Genotype RTx430 Revealed by Subcellular Protein Profiling" International Journal of Molecular Sciences 21, no. 24: 9706. https://doi.org/10.3390/ijms21249706