
The Lactococcal dgkB (yecE) and dxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials


Abstract
:1. Introduction
2. Results
2.1. Accumulation of L. lactis BHT-B Resistant Mutants
2.2. Identification of Genes Altered by Mutations in the BHT-B Resistant Mutants
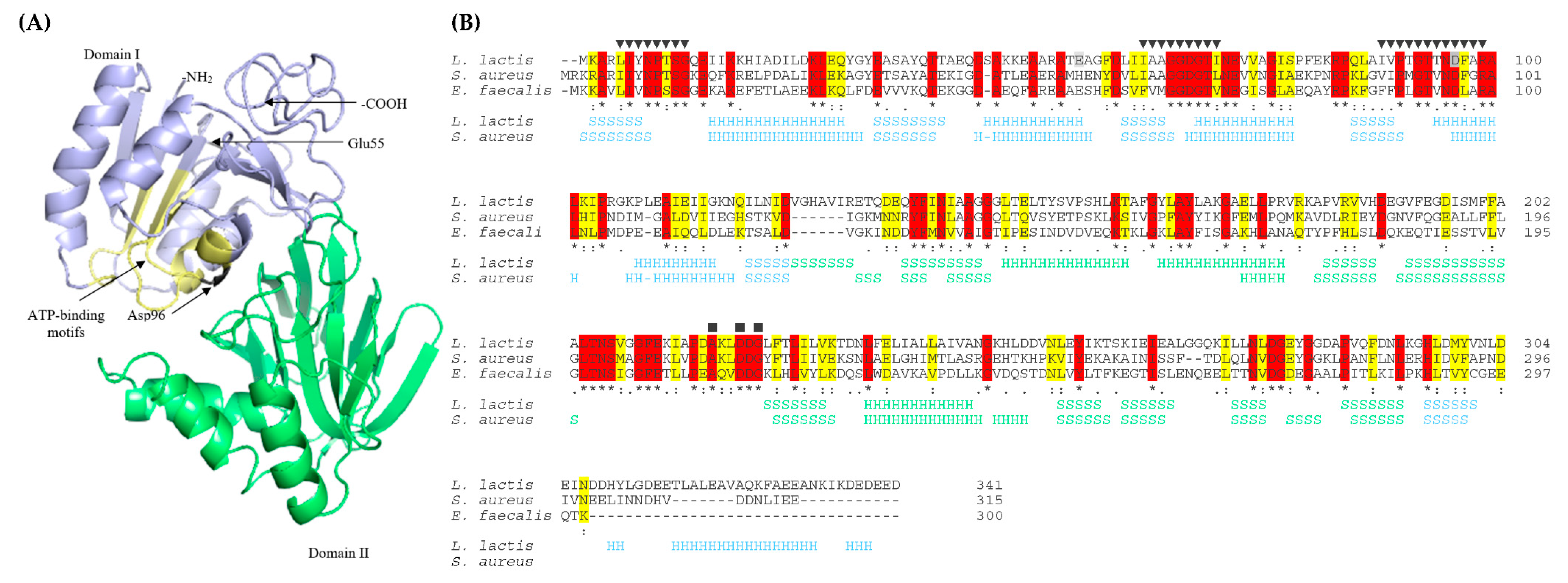
2.3. Analysis of Mutations in the yecE and dxsA Genes
2.4. Effect of Mutations on the Sensitivity to Bacteriocins
2.5. Effect of Mutations on the Sensitivity to Antibiotics
2.6. The Effect of Deletion or Overexpression of the dxsA Gene
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, and Culture Conditions
4.2. Bacteriocin Preparation
4.3. Selection of BHT-B Resistant Mutants
4.4. DNA Isolation and Manipulation
4.5. Construction of the Deletion Mutant
4.6. Construction of the Expression Mutants
4.7. Antibiotic Susceptibility Testing
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jordan, S.; Hutchings, M.I.; Mascher, T. Cell envelope stress response in Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 107–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta 2016, 1858, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Athanasaki, F.; Voulgaris, G.L.; Triarides, N.A.; Vardakas, K.Z. Resistance to fosfomycin: Mechanisms, frequency and clinical consequences. Int. J. Antimicrob. Agents 2019, 53, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, M.; Walker, S. Envelope Structures of Gram-Positive Bacteria. Curr. Top. Microbiol. Immunol. 2017, 404, 1–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cudic, P.; Kranz, J.K.; Behenna, D.C.; Kruger, R.G.; Tadesse, H.; Wand, A.J.; Veklich, Y.I.; Weisel, J.W.; McCafferty, D.G. Complexation of peptidoglycan intermediates by the lipoglycodepsipeptide antibiotic ramoplanin: Minimal structural requirements for intermolecular complexation and fibril formation. Proc. Natl. Acad. Sci. USA 2002, 99, 7384–7389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, D.; Debabov, D.; Hartsell, T.L.; Cano, R.J.; Adams, S.; Schuyler, J.A.; McMillan, R.; Pace, J.L. Approved glycopeptide antibacterial drugs: Mechanism of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a026989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economou, N.J.; Cocklin, S.; Loll, P.J. High-resolution crystal structure reveals molecular details of target recognition by bacitracin. Proc. Natl. Acad. Sci. USA 2013, 110, 14207–14212. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.S.; Apisarnthanarak, A.; Hsu, L.Y. Mechanisms of β-lactam antimicrobial resistance and epidemiology of major community- and healthcare-associated multidrug-resistant bacteria. Adv. Drug Deliv. Rev. 2014, 78, 3–13. [Google Scholar] [CrossRef]
- Dias, C.; Rauter, A.P. Membrane-targeting antibiotics: Recent developments outside the peptide space. Future Med. Chem. 2019, 11, 211–228. [Google Scholar] [CrossRef]
- Miller, W.R.; Bayer, A.S.; Arias, C.A. Mechanism of action and resistance to daptomycin in Staphylococcus aureus and enterococci. Cold Spring Harb. Perspect. Med. 2016, 6, a026997. [Google Scholar] [CrossRef] [Green Version]
- David, J.M.; Rajasekaran, A.K. Gramicidin A: A new mission for an old antibiotic. J. Kidney Cancer VHL 2015, 2, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bierbaum, G.; Sahl, H.-G. Lantibiotics: Mode of action, biosynthesis and bioengineering. Curr. Pharm. Biotechnol. 2009, 10, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Martínez, B.; Böttiger, T.; Schneider, T.; Rodríguez, A.; Sahl, H.-G.; Wiedemann, I. Specific interaction of the unmodified bacteriocin lactococcin 972 with the cell wall precursor lipid II. Appl. Environ. Microbiol. 2008, 74, 4666–4670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, Y.; Zendo, T.; Sonomoto, K. New type non-lantibiotic bacteriocins: Circular and leaderless bacteriocins. Benef. Microbes 2012, 3, 3–12. [Google Scholar] [CrossRef]
- Perez, R.H.; Zendo, T.; Sonomoto, K. Circular and leaderless bacteriocins: Biosynthesis, mode of action, applications, and prospects. Front. Microbiol. 2018, 9, 2085. [Google Scholar] [CrossRef]
- Netz, D.J.A.; Pohl, R.; Beck-Sickinger, A.G.; Selmer, T.; Pierik, A.J.; de Freire Bastos, M.D.; Sahl, H.G. Biochemical characterisation and genetic analysis of aureocin A53, a new, atypical bacteriocin from Staphylococcus aureus. J. Mol. Biol. 2002, 319, 745–756. [Google Scholar] [CrossRef]
- Hyink, O.; Balakrishnan, M.; Tagg, J.R. Streptococcus rattus strain BHT produces both a class I two-component lantibiotic and a class II bacteriocin. FEMS Microbiol. Lett. 2005, 252, 235–241. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Ichimasa, S.; Zendo, T.; Koga, S.; Yoneyama, F.; Nakayama, J.; Sonomoto, K. Structural analysis and characterization of lacticin Q, a novel bacteriocin belonging to a new family of unmodified bacteriocins of Gram-positive bacteria. Appl. Environ. Microbiol. 2007, 73, 2871–2877. [Google Scholar] [CrossRef] [Green Version]
- Iwatani, S.; Zendo, T.; Yoneyama, F.; Nakayama, J.; Sonomoto, K. Characterization and structure analysis of a novel bacteriocin, lacticin Z, produced by Lactococcus lactis QU 14. Biosci. Biotechnol. Biochem. 2007, 71, 1984–1992. [Google Scholar] [CrossRef] [Green Version]
- Sandiford, S.; Upton, M. Identification, characterization, and recombinant expression of epidermicin NI01, a novel unmodified bacteriocin produced by Staphylococcus epidermidis that displays potent activity against staphylococci. Antimicrob. Agents Chemother. 2012, 56, 1539–1547. [Google Scholar] [CrossRef] [Green Version]
- Lozo, J.; Mirkovic, N.; O’Connor, P.M.; Malesevic, M.; Miljkovic, M.; Polovic, N.; Jovcic, B.; Cotter, P.D.; Kojic, M. Lactolisterin BU, a novel class II broad-spectrum bacteriocin from Lactococcus lactis subsp. lactis bv. diacetylactis BGBU1-4. Appl. Environ. Microbiol. 2017, 83, e01519-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cintas, L.M.; Casaus, P.; Holo, H.; Hernandez, P.E.; Nes, I.F.; Håvarstein, L.S. Enterocins L50A and L50B, two novel bacteriocins from Enterococcus faecium L50, are related to staphylococcal hemolysins. J. Bacteriol. 1998, 180, 1988–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Vederas, J.C.; Whittal, R.M.; Zheng, J.; Stiles, M.E.; Carlson, D.; Franz, C.M.A.P.; McMullen, L.M.; van Belkum, M.J. Identification of an N-terminal formylated, two-peptide bacteriocin from Enterococcus faecalis 710C. J. Agric. Food Chem. 2011, 59, 5602–5608. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Zendo, T.; Sawa, N.; Perez, R.H.; Nakayama, J.; Sonomoto, K. Characterization and identification of weissellicin Y and weissellicin M, novel bacteriocins produced by Weissella hellenica QU 13. J. Appl. Microbiol. 2012, 112, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, A.M.; Salvioli, R.; Tatti, M.; Ciaffoni, F. Saposins and their interaction with lipids. Neurochem. Res. 1999, 24, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Towle, K.M.; Vederas, J.C. Structural features of many circular and leaderless bacteriocins are similar to those in saposins and saposin-like peptides. MedChemComm 2017, 8, 276–285. [Google Scholar] [CrossRef] [Green Version]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef]
- Netz, D.J.A.; de Freire Bastos, M.D.; Sahl, H.G. Mode of action of the antimicrobial peptide aureocin A53 from Staphylococcus aureus. Appl. Environ. Microbiol. 2002, 68, 5274–5280. [Google Scholar] [CrossRef] [Green Version]
- Yoneyama, F.; Imura, Y.; Ohno, K.; Zendo, T.; Nakayama, J.; Matsuzaki, K.; Sonomoto, K. Peptide-lipid huge toroidal pore, a new antimicrobial mechanism mediated by a lactococcal bacteriocin, lacticin Q. Antimicrob. Agents Chemother. 2009, 53, 3211–3217. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Yoneyama, F.; Toshimitsu, N.; Zendo, T.; Nakayama, J.; Sonomoto, K. Lethal hydroxyl radical accumulation by a lactococcal bacteriocin, lacticin Q. Antimicrob. Agents Chemother. 2013, 57, 3897–3902. [Google Scholar] [CrossRef] [Green Version]
- Schrecke, K.; Staroń, A.; Mascher, T. Two-component signalling in the Gram-positive envelope stress response: Intramembrane-sensing histidine kinases and accessory membrane proteins. In Two-Component Systems in Bacteria; Horizon Scientific Press: Poole, UK, 2012; p. 450. ISBN 978-1-908230-08-9. [Google Scholar]
- Kramer, N.E.; van Hijum, S.A.F.T.; Knol, J.; Kok, J.; Kuipers, O.P. Transcriptome analysis reveals mechanisms by which Lactococcus lactis acquires nisin resistance. Antimicrob. Agents Chemother. 2006, 50, 1753–1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheul, A.; Russell, N.J.; Van’T Hof, R.; Rombouts, F.M.; Abee, T. Modifications of membrane phospholipid composition in nisin-resistant Listeria monocytogenes Scott A. Appl. Environ. Microbiol. 1997, 63, 3451–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crandall, A.D.; Montville, T.J. Nisin resistance in Listeria monocytogenes ATCC 700302 is a complex phenotype. Appl. Environ. Microbiol. 1998, 64, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhardwaj, P.; Hans, A.; Ruikar, K.; Guan, Z.; Palmer, K.L. Reduced chlorhexidine and daptomycin susceptibility in vancomycin-resistant Enterococcus faecium after serial chlorhexidine exposure. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [Green Version]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef]
- Campelo, A.B.; López-González, M.J.; Escobedo, S.; Janzen, T.; Neves, A.R.; Rodríguez, A.; Martínez, B. Mutations selected after exposure to bacteriocin Lcn972 activate a Bce-like bacitracin resistance module in Lactococcus lactis. Front. Microbiol. 2020, 11, 1805. [Google Scholar] [CrossRef]
- Facklam, R. What happened to the streptococci: Overview of taxonomic and nomenclature changes. Clin. Microbiol. Rev. 2002, 15, 613–630. [Google Scholar] [CrossRef] [Green Version]
- Miller, D.J.; Jerga, A.; Rock, C.O.; White, S.W. Analysis of the Staphylococcus aureus DgkB structure reveals a common catalytic mechanism for the soluble diacylglycerol kinases. Struct. Lond. Engl. 1993 2008, 16, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.; Usunow, G.; Lange, G.; Busch, M.; Tong, L. Crystal structure of 1-deoxy-D-xylulose 5-phosphate synthase, a crucial enzyme for isoprenoids biosynthesis. J. Biol. Chem. 2007, 282, 2676–2682. [Google Scholar] [CrossRef] [Green Version]
- Tymoszewska, A.; Diep, D.B.; Wirtek, P.; Aleksandrzak-Piekarczyk, T. The non-lantibiotic bacteriocin garvicin Q targets Man-PTS in a broad spectrum of sensitive bacterial genera. Sci. Rep. 2017, 7, 8359. [Google Scholar] [CrossRef] [Green Version]
- Tymoszewska, A.; Diep, D.B.; Aleksandrzak-Piekarczyk, T. The extracellular loop of Man-PTS subunit IID is responsible for the sensitivity of Lactococcus garvieae to garvicins A, B and C. Sci. Rep. 2018, 8, 15790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerga, A.; Lu, Y.-J.; Schujman, G.E.; de Mendoza, D.; Rock, C.O. Identification of a soluble diacylglycerol kinase required for lipoteichoic acid production in Bacillus subtilis. J. Biol. Chem. 2007, 282, 21738–21745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaspar, J.R.; Godwin, M.J.; Velsko, I.M.; Richards, V.P.; Burne, R.A. Spontaneously arising Streptococcus mutans variants with reduced susceptibility to chlorhexidine display genetic defects and diminished fitness. Antimicrob. Agents Chemother. 2019, 63, e00161-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, R.C.; Zeng, L.; Culp, D.J.; Burne, R.A. Genomewide Identification of Essential Genes and Fitness Determinants of Streptococcus mutans UA159. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, K.M.; Serio, A.W.; Kane, T.R.; Connolly, L.E. Aminoglycosides: An Overview. Cold Spring Harb. Perspect. Med. 2016, 6, a027029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grossman, T.H. Tetracycline Antibiotics and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meganathan, R.; Kwon, O. Biosynthesis of menaquinone (vitamin K2) and ubiquinone (coenzyme Q). EcoSal Plus 2009, 3. [Google Scholar] [CrossRef] [Green Version]
- Reusch, V.M. Lipopolymers, isoprenoids, and the assembly of the gram-positive cell wall. Crit. Rev. Microbiol. 1984, 11, 129–155. [Google Scholar] [CrossRef]
- Sáenz, J.P.; Grosser, D.; Bradley, A.S.; Lagny, T.J.; Lavrynenko, O.; Broda, M.; Simons, K. Hopanoids as functional analogues of cholesterol in bacterial membranes. Proc. Natl. Acad. Sci. USA 2015, 112, 11971–11976. [Google Scholar] [CrossRef] [Green Version]
- George, K.W.; Thompson, M.G.; Kim, J.; Baidoo, E.E.K.; Wang, G.; Benites, V.T.; Petzold, C.J.; Chan, L.J.G.; Yilmaz, S.; Turhanen, P.; et al. Integrated analysis of isopentenyl pyrophosphate (IPP) toxicity in isoprenoid-producing Escherichia coli. Metab. Eng. 2018, 47, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Sivy, T.L.; Fall, R.; Rosenstiel, T.N. Evidence of isoprenoid precursor toxicity in Bacillus subtilis. Biosci. Biotechnol. Biochem. 2011, 75, 2376–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belin, B.J.; Busset, N.; Giraud, E.; Molinaro, A.; Silipo, A.; Newman, D.K. Hopanoid lipids: From membranes to plant-bacteria interactions. Nat. Rev. Microbiol. 2018, 16, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmerk, C.L.; Bernards, M.A.; Valvano, M.A. Hopanoid production is required for low-pH tolerance, antimicrobial resistance, and motility in Burkholderia cenocepacia. J. Bacteriol. 2011, 193, 6712–6723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malott, R.J.; Steen-Kinnaird, B.R.; Lee, T.D.; Speert, D.P. Identification of hopanoid biosynthesis genes involved in polymyxin resistance in Burkholderia multivorans. Antimicrob. Agents Chemother. 2012, 56, 464–471. [Google Scholar] [CrossRef] [Green Version]
- Malott, R.J.; Wu, C.-H.; Lee, T.D.; Hird, T.J.; Dalleska, N.F.; Zlosnik, J.E.A.; Newman, D.K.; Speert, D.P. Fosmidomycin decreases membrane hopanoids and potentiates the effects of colistin on Burkholderia multivorans clinical isolates. Antimicrob. Agents Chemother. 2014, 58, 5211–5219. [Google Scholar] [CrossRef] [Green Version]
- Fritz, G.; Dintner, S.; Treichel, N.S.; Radeck, J.; Gerland, U.; Mascher, T.; Gebhard, S. A new way of sensing: Need-based activation of antibiotic resistance by a flux-sensing mechanism. mBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Kobras, C.M.; Piepenbreier, H.; Emenegger, J.; Sim, A.; Fritz, G.; Gebhard, S. BceAB-type antibiotic resistance transporters appear to act by target protection of cell wall synthesis. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef] [Green Version]
- López-González, M.J.; Escobedo, S.; Rodríguez, A.; Neves, A.R.; Janzen, T.; Martínez, B. Adaptive evolution of industrial Lactococcus lactis under cell envelope stress provides phenotypic diversity. Microbiology 2018, 9, 2654. [Google Scholar] [CrossRef] [Green Version]
- Kingston, A.W.; Liao, X.; Helmann, J.D. Contributions of the σ(W), σ(M) and σ(X) regulons to the lantibiotic resistome of Bacillus subtilis. Mol. Microbiol. 2013, 90, 502–518. [Google Scholar] [CrossRef] [Green Version]
- Sass, V.; Pag, U.; Tossi, A.; Bierbaum, G.; Sahl, H.-G. Mode of action of human β-defensin 3 against Staphylococcus aureus and transcriptional analysis of responses to defensin challenge. Int. J. Med. Microbiol. 2008, 298, 619–633. [Google Scholar] [CrossRef]
- Tran, T.T.; Panesso, D.; Gao, H.; Roh, J.H.; Munita, J.M.; Reyes, J.; Diaz, L.; Lobos, E.A.; Shamoo, Y.; Mishra, N.N.; et al. Whole-genome analysis of a daptomycin-susceptible Enterococcus faecium strain and its daptomycin-resistant variant arising during therapy. Antimicrob. Agents Chemother. 2013, 57, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, B.; Suárez, J.E.; Rodríguez, A. Antimicrobials produced by wild lactococcal strains isolated from homemade cheeses. J. Food Prot. 1995, 58, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Gibson, T.J. Studies on the Epstein-Barr Virus Genome; Cambridge University: Cambridge, UK, 1984. [Google Scholar]
- Leenhouts, K.; Buist, G.; Bolhuis, A.; ten Berge, A.; Kiel, J.; Mierau, I.; Dabrowska, M.; Venema, G.; Kok, J. A general system for generating unlabelled gene replacements in bacterial chromosomes. Mol. Gen. Genet. MGG 1996, 253, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maguin, E.; Prévost, H.; Ehrlich, S.D.; Gruss, A. Efficient insertional mutagenesis in lactococci and other Gram-positive bacteria. J. Bacteriol. 1996, 178, 931–935. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Aleksandrzak-Piekarczyk, T.; Polak, J.; Jezierska, B.; Renault, P.; Bardowski, J. Genetic characterization of the CcpA-dependent, cellobiose-specific PTS system comprising CelB, PtcB and PtcA that transports lactose in Lactococcus lactis IL1403. Int. J. Food Microbiol. 2011, 145, 186–194. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mutant | Sensitivity Decrease * | Mutation in Nucleotide Sequence | Mutation in Amino Acid Sequence | Potential Function |
|---|---|---|---|---|
| MUT70 | 4× | 286G→T in dgkB (yecE) | Asp96→Tyr | Diacylglycerol kinase |
| MUT72 | 4× | 163G→T in dgkB (yecE) | Glu55→STOP | Diacylglycerol kinase |
| MUT71, MUT73, MUT78 | 4× | 1469C→G in dxsA | Ser490→STOP | 1-deoxy-D-xylulose 5-phosphate (DXP) synthase |
| MUT78.2 | 8× | 1469C→G in dxsA; 780T→G in ysaB; 328G→T in ynhD | Ser490→STOP;Tyr260→STOP; Glu110→STOP | DXP synthase; ABC transporter permease; hypothetical protein |
| Antimicrobial Agent | LMGT 3419 | MUT70 | MUT72 | MUT78 | MUT78.2 | MUT80 | MUT84 | MUT85 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Wild-Type | dgkB- (yecE-) | dgkB- (yecE-) | dxsA- | dxsA-ysaB- ynhD- | ∆dxsA | dxsA- pIBB-JZK | dxsA- pIBB-JZK:dxsA | ||||
| MIC50 [µg/mL] | |||||||||||
| K411 | 3.2 | 6.4 | 6.4 | 12.5 | >25 | 3.2 | 12.5 | 6.3 | |||
| BHT-B | 6.3 | 25 | 25 | 25 | >25 | 6.3 | 25 | 12.5 | |||
| Ent7 | 0.2 | 0.4 | 0.4 | 1.6 | 6.4 | 0.2 | 0.4 | 0.2 | |||
| EntL50 | 0.2 | 0.8 | 0.8 | 6.4 | 25 | 0.4 | 0.8 | 0.8 | |||
| WelM | 2.4 | 18.8 | 18.8 | 18.8 | >25 | 2.4 | >25 | 25 | |||
| SalC | 6.3 | 12.5 | 12.5 | >25 | >25 | 6.3 | 25 | 12.5 | |||
| Nisin | 0.47 | 1.9 | 1.9 | >15 | >15 | 0.47 | >15 | 15 | |||
| Lcn972 | 0.4 | 1.6 | 1.6 | 3.2 | >3.2 | 0.4 | 3.2 | 1.6 | |||
| MIC [µg/mL] | |||||||||||
| Bacitracin | 12 | 4 | 3 | 4–6 | 2 | 12 | 6 | 6 | |||
| Daptomycin | 0.094 | 1 | 0.5 | 1 | 2 | 0.064 | 0.5 | 0.5 | |||
| Gramicidin | 0.023 | 0.188 | 0.188 | >0.75 | 0.75 | 0.012 | 0.75 | 0.75 | |||
| Gentamicin | 2 | 12 | 8–12 | 8 | 12–16 | 2 | 6 | 6 | |||
| Kanamycin | 12 | 32 | 32 | 32 | 32 | 6 | 32 | 32 | |||
| Streptomycin | 24 | 48 | 48 | 48 | 64 | 12 | 64 | 64 | |||
| Vancomycin | 0.38 | 1 | 0.5–0.75 | 1 | 1.5 | 0.5 | 1 | 1 | |||
| Fosfomycin | 384 | >1024 | >1024 | 1024 | >1024 | >1024 | 384 | 384 | |||
| Diameter [mm] | |||||||||||
| Carbenicillin | 28 | 31 | 31 | 32 | 33–34 | 29 | 33 | 34 | |||
| Chlortetracycline | 21 | 29 | 32 | 33 | 32 | 24 | 14 * | 15 * | |||
| Sensitivity scale: | 6× | 4× | 3× | 2× | 2× | 4× | 8× | 16× | ≥32× | ||
| Fold of increased sensitivity← | → Fold of decreased sensitivity | ||||||||||
| Strains, Plasmids, Primers | Description, Primer Sequence | Source * (Reference) |
|---|---|---|
| Strains | ||
| Lactococcus lactis | ||
| IBB51 | Nisin producer | IBB PAS |
| IPLA 972 | Lcn972 producer | IPLA-CSIC [63] |
| LMGT 3419 | Indicator strain | LMGT |
| MUT70, MUT71, MUT72, MUT73, MUT78 | LMGT 3419 spontaneous first generation mutants resistant to AurA53- and EntL50-like bacteriocins | This study |
| MUT78.2 | LMGT 3419 spontaneous second-generation mutant resistant to AurA53- and EntL50-like bacteriocins | This study |
| MUT79c | LMGT 3419 strain carrying pGhost9:ΔdxsA | This study |
| MUT80 | LMGT 3419 strain with dxsA deletion | This study |
| MUT82 | LMGT 3419 strain carrying pIBB-JZK | This study |
| MUT83 | LMGT 3419 strain carrying pIBB-JZK:dxsA | This study |
| MUT84 | MUT78 strain carrying pIBB-JZK | This study |
| MUT85 | MUT78 strain carrying pIBB-JZK:dxsA | This study |
| Escherichia coli | ||
| TG1 | Host strain, Δ(hsdMS-mcrB) 5Δ(lac-proAB) supE thi-1 F’(traD36 proAB+lacIqZΔM15) | [64] |
| EC1000 | Host strain, Kmr, repA+ derivative of MC1000, carrying a single copy of the pWV01 repA gene in glgB | [65] |
| Plasmids | ||
| pGEMT | Ampr, M13ori, linear T-overhang vector | Promega |
| pGhost9 | Emr, repA(Ts) | IBB PAS [66] |
| pIBB-JZK pIBB-JZK:PptcB::dxsA | Ampr,Tetr, cellobiose-responsive promoter (PptcB) Ampr,Tetr, dxsA cloned under the control of PptcB | IBB PAS |
| Primers | DNA sequence (5′->3′), restriction site ** | |
| 1224/1233 | CGCCAGGGTTTTCCCAGTCACGA/AGCGGATAACAATTTCACACAGG | |
| pGhfor/rev | TGTAAAACGACGGCCAGTG/AGTACCGTTACTTATGAGC | |
| pJZK-IBBfor/rev | AGTCGCCTAAAGGTTGC/CGATGTTCTGTCCCTTG | |
| DxsAUPfor/rev | CAAGTATGCTCCAAGG/GAGGATCCCGAGCTCTTCTAGTTC | |
| DxsADNfor/rev | GAGGATCCGCAGAGCTTATCTCCTA/CCAGCAGAATGGAAAC | |
| DxsAOUTfor/rev | GAGGAGTCCTCCAAATG/TCTGTAGATGTCACGG | |
| DxsAfor/rev | CGATGGATCCCTAAATAGAACTAGGAAG/CGATCTCGAGGAGATAAGCTCTGCCTAC | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tymoszewska, A.; Aleksandrzak-Piekarczyk, T. The Lactococcal dgkB (yecE) and dxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials. Int. J. Mol. Sci. 2021, 22, 1014. https://doi.org/10.3390/ijms22031014
Tymoszewska A, Aleksandrzak-Piekarczyk T. The Lactococcal dgkB (yecE) and dxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials. International Journal of Molecular Sciences. 2021; 22(3):1014. https://doi.org/10.3390/ijms22031014
Chicago/Turabian StyleTymoszewska, Aleksandra, and Tamara Aleksandrzak-Piekarczyk. 2021. "The Lactococcal dgkB (yecE) and dxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials" International Journal of Molecular Sciences 22, no. 3: 1014. https://doi.org/10.3390/ijms22031014