
Partial Inhibition of Calcineurin Activity by Rcn2 as a Potential Remedy for Vps13 Deficiency



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The RCN2 Gene is a Multicopy Suppressor of vps13Δ SDS Hypersensitivity
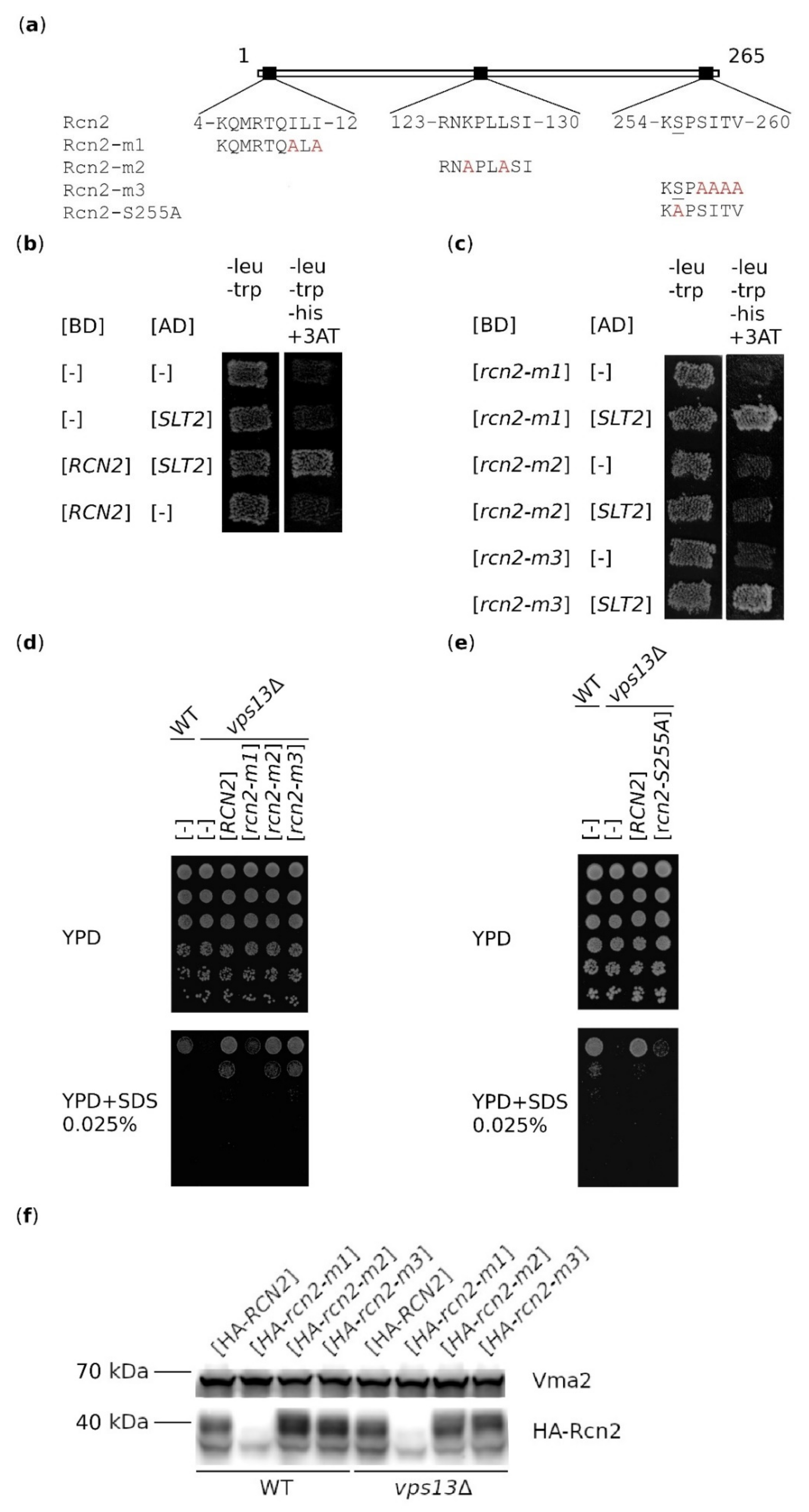
2.2. Binding of Rcn2 to Slt2 Kinase is Not Important for Suppression
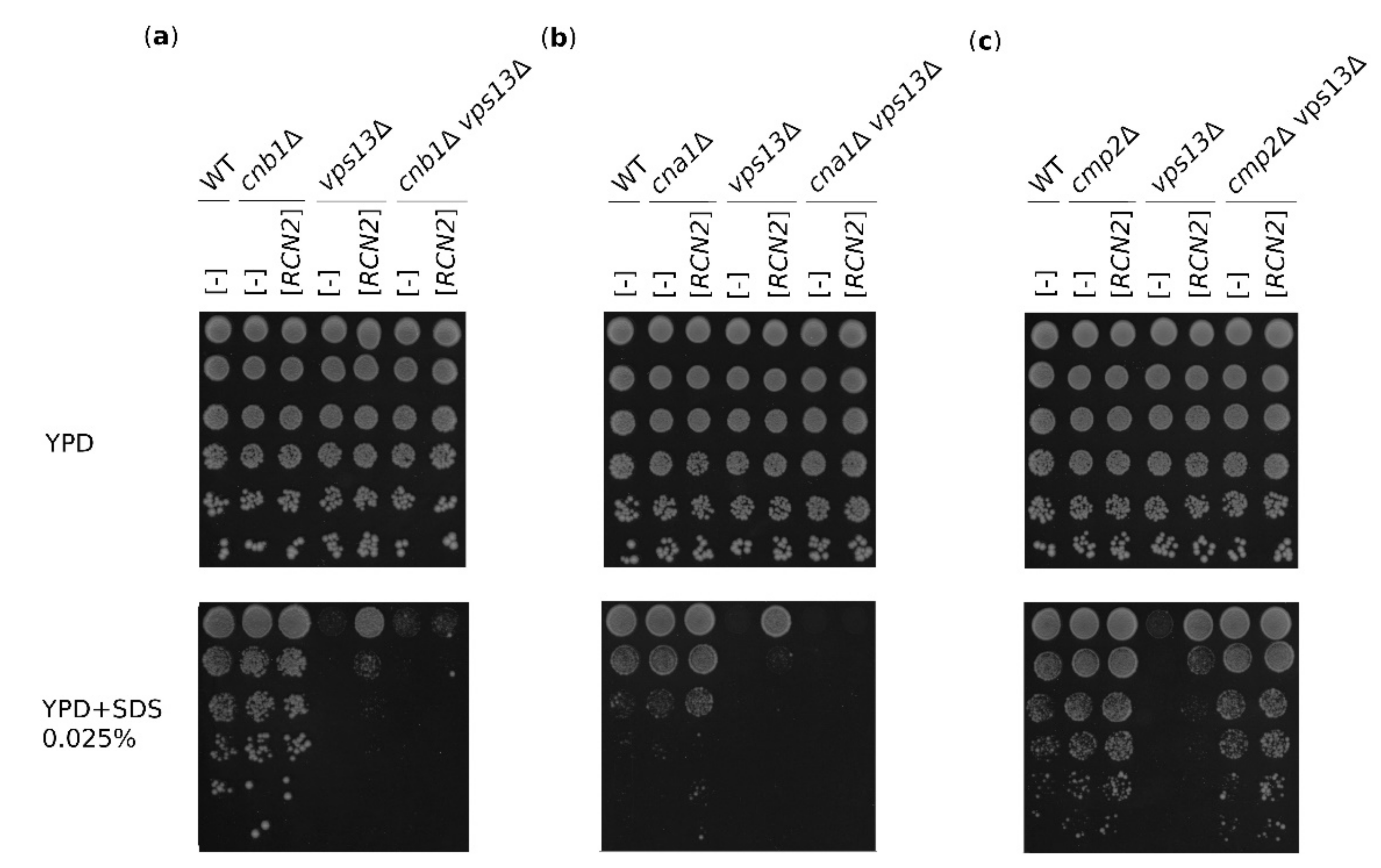
2.3. The Suppression of vps13Δ by RCN2 Requires Calcineurin
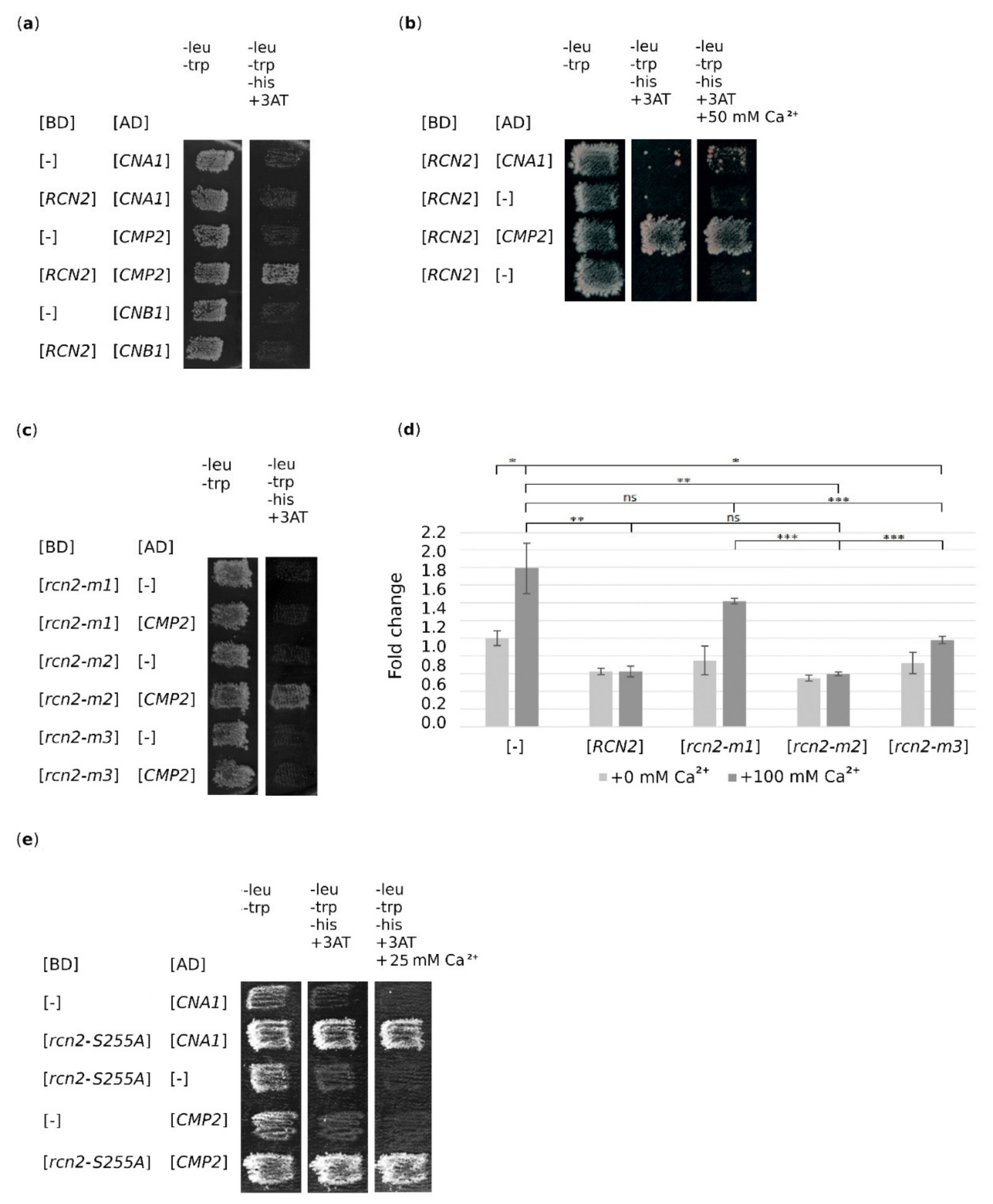
2.4. Both rcn2-m1 and rcn2-m3 Mutations Abolish the Binding of Rcn2 to Catalytic Subunit of Calcineurin, Cmp2
2.5. Partial Inhibition of Calcineurin Activity is Necessary for Suppression
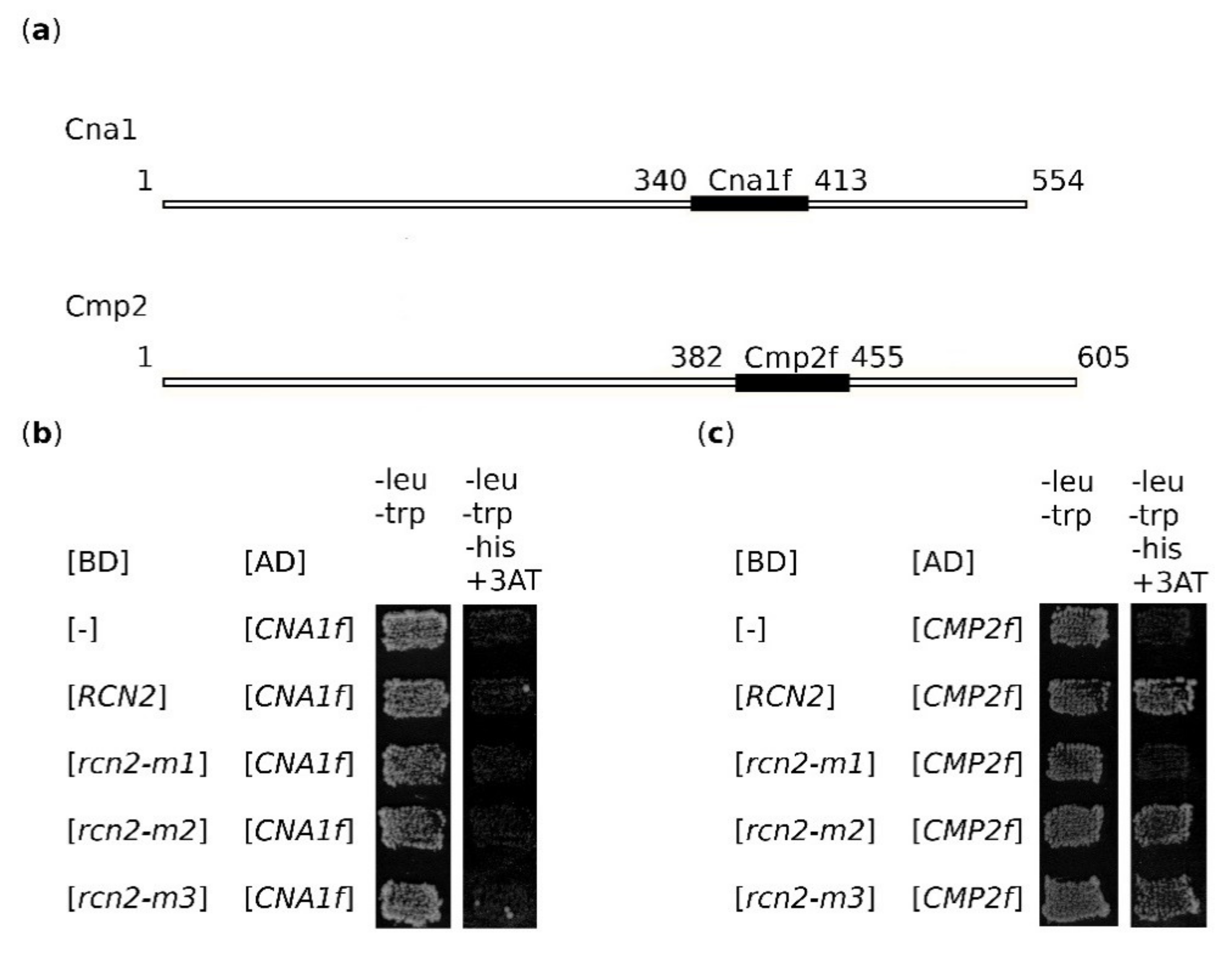
2.6. Rcn2 Binds to the Cmp2 Fragment Corresponding to aa 382–455
3. Discussion
4. Materials and Methods
4.1. Strains, Media, and Growth Conditions
4.2. Plasmids, Mutagenesis, and Screen for Multicopy Suppressors of vps13 Mutations
4.3. β-Galactosidase Activity Assay
4.4. Protein Extracts and Western Blot Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rao, A.; Luo, C.; Hogan, P.G. Transcription factors of the nfat family: Regulation and Function. Annu. Rev. Immunol. 1997, 15, 707–747. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yi, H.; Guerini, D.; Klee, C.; McBride, O. Calcineurin A alpha (PPP3CA), calcineurin A beta (PPP3CB) and calcineurin B (PPP3R1) are located on human chromosomes 4, 10q21→q22 and 2p16→p15 respectively. Cytogenet. Genome Res. 1996, 72, 236–241. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Zhang, J.; Yuan, J.; Dang, Y.; Yang, C.; Chen, X.; Xu, J.; Yu, L. Characterization of a human regulatory subunit of protein phosphatase 3 gene (PPP3RL) expressed specifically in testis. Mol. Biol. Rep. 2005, 32, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Saraf, J.; Bhattacharya, P.; Kalia, K.; Borah, A.; Sarmah, D.; Kaur, H.; Dave, K.R.; Yavagal, D.R. A Friend or Foe: Calcineurin across the Gamut of Neurological Disorders. ACS Central Sci. 2018, 4, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Klee, C.B.; Ren, H.; Wang, X. Regulation of the Calmodulin-stimulated Protein Phosphatase, Calcineurin. J. Biol. Chem. 1998, 273, 13367–13370. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, A.; Roy, J.; Martínez-Martínez, S.; López-Maderuelo, M.D.; Niño-Moreno, P.; Ortí, L.; Pantoja-Uceda, D.; Pineda-Lucena, A.; Cyert, M.S.; Redondo, J.M. A Conserved Docking Surface on Calcineurin Mediates Interaction with Substrates and Immunosuppressants. Mol. Cell 2009, 33, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Liu, J. FK506 and cyclosporin, molecular probes for studying intracellular signal transduction. Immunol. Today 1993, 14, 290–295. [Google Scholar] [CrossRef]
- Bechstein, W.O. Neurotoxicity of calcineurin inhibitors: Impact and clinical management. Transpl. Int. 2000, 13, 313–326. [Google Scholar] [CrossRef]
- Mehta, S.; Li, H.; Hogan, P.G.; Cunningham, K.W. Domain Architecture of the Regulators of Calcineurin (RCANs) and Identification of a Divergent RCAN in Yeast. Mol. Cell. Biol. 2009, 29, 2777–2793. [Google Scholar] [CrossRef] [Green Version]
- Strippoli, P.; Lenzi, L.; Petrini, M.; Carinci, P.; Zannotti, M. A New Gene Family Including DSCR1 (Down Syndrome Candidate Region 1) and ZAKI-4: Characterization from Yeast to Human and Identification of DSCR1-like 2, a Novel Human Member (DSCR1L2). Genomics 2000, 64, 252–263. [Google Scholar] [CrossRef]
- Candelas, E.S.; Farré, D.; Aranguren-Ibáñez, Á.; Martínez-Høyer, S.; Pérez-Riba, M. The Vertebrate RCAN Gene Family: Novel Insights into Evolution, Structure and Regulation. PLoS ONE 2014, 9, e85539. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.; Zhang, Y.; Zhang, C.; Huang, L.; Tan, S.; Wang, P.; Vilariño-Güell, C.; Song, W.; Sun, X. Regulator of calcineurin 1 is a novel RNA-binding protein to regulate neuronal apoptosis. Mol. Psychiatry 2019, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Ahnn, J. Regulator of Calcineurin (RCAN): Beyond down Syndrome Critical Region. Mol. Cells 2020, 43, 671–685. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, J.-J.; A Pritchard, M.; Planas, A.M.; Bosch, A.; Ferrer, I.; Estivill, X. A new human gene from the Down syndrome critical region encodes a proline-rich protein highly expressed in fetal brain and heart. Hum. Mol. Genet. 1995, 4, 1935–1944. [Google Scholar] [CrossRef] [PubMed]
- Sanna, B.; Brandt, E.B.; Kaiser, R.A.; Pfluger, P.T.; Witt, S.A.; Kimball, T.R.; Van Rooij, E.; De Windt, L.J.; Rothenberg, M.E.; Tschöp, M.; et al. Modulatory calcineurin-interacting proteins 1 and 2 function as calcineurin facilitators in vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 7327–7332. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, R.-B.; Cao, Q.; Fan, K.-Q.; Huang, L.-J.; Yu, J.-S.; Gao, Z.-J.; Huang, T.; Zhong, J.-Y.; Mao, X.-T.; et al. USP16-mediated deubiquitination of calcineurin A controls peripheral T cell maintenance. J. Clin. Investig. 2019, 129, 2856–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafadar, K.A.; Zhu, H.; Snyder, M.; Cyert, M.S. Negative regulation of calcineurin signaling by Hrr25p, a yeast homolog of casein kinase I. Genes Dev. 2003, 17, 2698–2708. [Google Scholar] [CrossRef] [Green Version]
- Cyert, M.S.; Kunisawa, R.; Kaim, D.; Thorner, J. Yeast has homologs (CNA1 and CNA2 gene products) of mammalian calcineurin, a calmodulin-regulated phosphoprotein phosphatase. Proc. Natl. Acad. Sci. USA 1991, 88, 7376–7380. [Google Scholar] [CrossRef] [Green Version]
- Cyert, M.S.; Thorner, J. Regulatory subunit (CNB1 gene product) of yeast Ca2+/calmodulin-dependent phosphoprotein phosphatases is required for adaptation to pheromone. Mol. Cell. Biol. 1992, 12, 3460–3469. [Google Scholar] [CrossRef] [Green Version]
- Kingsbury, T.J.; Cunningham, K.W. A conserved family of calcineurin regulators. Genes Dev. 2000, 14, 1595–1604. [Google Scholar]
- Stathopoulos, A.M.; Cyert, M.S. Calcineurin acts through the CRZ1/TCN1-encoded transcription factor to regulate gene expression in yeast. Genes Dev. 1997, 11, 3432–3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, J.; Li, H.; Hogan, P.G.; Cyert, M.S. A Conserved Docking Site Modulates Substrate Affinity for Calcineurin, Signaling Output, and In Vivo Function. Mol. Cell 2007, 25, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Espeso, E.A. The CRaZy Calcium Cycle. Adv. Exp. Med. Biol. 2016, 169–186. [Google Scholar]
- Batiza, A.F.; Schulz, T.; Masson, P.H. Yeast Respond to Hypotonic Shock with a Calcium Pulse. J. Biol. Chem. 1996, 271, 23357–23362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denis, V.; Cyert, M.S. Internal Ca2+ release in yeast is triggered by hypertonic shock and mediated by a TRP channel homologue. J. Cell Biol. 2002, 156, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, W.E.; Vlasek, C.; Klukovich, R.; Coffee, S. Ethanol induces calcium influx via the Cch1-Mid1 transporter in Saccharomyces cerevisiae. Arch. Microbiol. 2011, 193, 323–334. [Google Scholar] [CrossRef]
- Bonilla, M.; Cunningham, K.W. Mitogen-activated Protein Kinase Stimulation of Ca2+ Signaling Is Required for Survival of Endoplasmic Reticulum Stress in Yeast. Mol. Biol. Cell 2003, 14, 4296–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett-Engele, P.; Moilanen, B.; Cyert, M.S. Calcineurin, the Ca2+/calmodulin-dependent protein phosphatase, is essential in yeast mutants with cell integrity defects and in mutants that lack a functional vacuolar H(+)-ATPase. Mol. Cell. Biol. 1995, 15, 4103–4114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caraveo, G.; Soste, M.; Cappelleti, V.; Fanning, S.; Van Rossum, D.B.; Whitesell, L.; Huang, Y.; Chung, C.Y.; Baru, V.; Zaichick, S.; et al. FKBP12 contributes to α-synuclein toxicity by regulating the calcineurin-dependent phosphoproteome. Proc. Natl. Acad. Sci. USA 2017, 114, E11313–E11322. [Google Scholar] [CrossRef] [Green Version]
- Soczewka, P.; Kolakowski, D.; Rooij, I.S.-D.; Rzepnikowska, W.; Ayscough, K.R.; Kaminska, J.; Żołądek, T. Yeast-model-based study identified myosin- and calcium-dependent calmodulin signalling as a potential target for drug intervention in chorea-acanthocytosis. Dis. Model. Mech. 2019, 12, dmm036830. [Google Scholar] [CrossRef] [Green Version]
- Rampoldi, L.; Dobson-Stone, C.; Rubio, J.P.; Danek, A.; Chalmers, R.M.; Wood, N.W.; Verellen, C.; Ferrer, X.; Malandrini, A.; Fabrizi, G.M.; et al. A conserved sorting-associated protein is mutant in chorea-acanthocytosis. Nat. Genet. 2001, 28, 119–120. [Google Scholar] [CrossRef] [PubMed]
- Kolehmainen, J.; Black, G.C.; Saarinen, A.; Chandler, K.; Clayton-Smith, J.; Träskelin, A.L.; Perveen, R.; Kivitie-Kallio, S.; Norio, R.; Warburg, M.; et al. Cohen syndrome is caused by mutations in a novel gene, COH1, encoding a transmembrane protein with a presumed role in vesicle-mediated sorting and intracellular protein transport. Am. J. Hum. Genet 2003, 72, 1359–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage, S.; Drouet, V.; Majounie, E.; Deramecourt, V.; Jacoupy, M.; Nicolas, A.; Cormier-Dequaire, F.; Hassoun, S.M.; Pujol, C.; Ciura, S.; et al. Loss of VPS13C Function in Autosomal-Recessive Parkinsonism Causes Mitochondrial Dysfunction and Increases PINK1/Parkin-Dependent Mitophagy. Am. J. Hum. Genet. 2016, 98, 500–513. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, J.; Meijer, I.A.; Lessel, D.; Mencacci, N.E.; Krainc, D.; Hempel, M.; Tsiakas, K.; Prokisch, H.; Rossignol, E.; Helm, M.H.; et al. Recessive mutations in VPS13D cause childhood onset movement disorders. Ann. Neurol. 2018, 83, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Seong, E.; Insolera, R.; Dulovic, M.; Kamsteeg, E.; Trinh, J.; Brüggemann, N.; Sandford, E.; Li, S.; Ozel, A.B.; Li, J.; et al. Mutations in VPS13D lead to a new recessive ataxia with spasticity and mitochondrial defects. Ann. Neurol. 2018, 83, 1075–1088. [Google Scholar] [CrossRef]
- Velayos-Baeza, A.; Vettori, A.; Copley, R.R.; Dobson-Stone, C.; Monaco, A.P. Analysis of the human VPS13 gene family. Genomics 2004, 84, 536–549. [Google Scholar] [CrossRef]
- Kumar, N.; Leonzino, M.; Hancock-Cerutti, W.; Horenkamp, F.A.; Li, P.; Lees, J.A.; Wheeler, H.; Reinisch, K.M.; De Camilli, P. VPS13A and VPS13C are lipid transport proteins differentially localized at ER contact sites. J. Cell Biol. 2018, 217, 3625–3639. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Lees, J.A.; Lusk, C.P.; Reinisch, K.M. Cryo-EM reconstruction of a VPS13 fragment reveals a long groove to channel lipids between membranes. J. Cell Biol. 2020, 219, e202001161. [Google Scholar] [CrossRef]
- Lang, A.B.; Peter, A.T.J.; Walter, P.; Kornmann, B. ER–mitochondrial junctions can be bypassed by dominant mutations in the endosomal protein Vps13. J. Cell Biol. 2015, 210, 883–890. [Google Scholar] [CrossRef] [Green Version]
- Park, J.-S.; Thorsness, M.K.; Policastro, R.; McGoldrick, L.L.; Hollingsworth, N.M.; Thorsness, P.E.; Neiman, A.M. Yeast Vps13 promotes mitochondrial function and is localized at membrane contact sites. Mol. Biol. Cell 2016, 27, 2435–2449. [Google Scholar] [CrossRef] [Green Version]
- Peter, A.T.J.; Herrmann, B.; Antunes, D.; Rapaport, D.; Dimmer, K.S.; Kornmann, B. Vps13-Mcp1 interact at vacuole–mitochondria interfaces and bypass ER–mitochondria contact sites. J. Cell Biol. 2017, 216, 3219–3229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolakowski, D.; Kaminska, J.; Zoladek, T. The binding of the APT1 domains to phosphoinositides is regulated by metal ions in vitro. Biochim. Biophys. Acta (BBA) Biomembr. 2020, 1862, 183349. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Honisch, S.; Schmidt, S.; Yan, J.; Schmid, E.; Alkahtani, S.; AlKahtane, A.A.; Alarifi, S.; Stournaras, C.; Lang, F. Chorein Sensitive Orai1 Expression and Store Operated Ca2+ Entry in Rhabdomyosarcoma Cells. Cell. Physiol. Biochem. 2016, 40, 1141–1152. [Google Scholar] [CrossRef]
- Pelzl, L.; Elsir, B.; Sahu, I.; Bissinger, R.; Singh, Y.; Sukkar, B.; Honisch, S.; Schoels, L.; Jemaà, M.; Lang, E.; et al. Lithium Sensitivity of Store Operated Ca2+ Entry and Survival of Fibroblasts Isolated from Chorea-Acanthocytosis Patients. Cell. Physiol. Biochem. 2017, 42, 2066–2077. [Google Scholar] [CrossRef] [Green Version]
- Stanslowsky, N.; Reinhardt, P.; Glass, H.; Kalmbach, N.; Naujock, M.; Hensel, N.; Lübben, V.; Pal, A.; Venneri, A.; Lupo, F.; et al. Neuronal Dysfunction in iPSC-Derived Medium Spiny Neurons from Chorea-Acanthocytosis Patients Is Reversed by Src Kinase Inhibition and F-Actin Stabilization. J. Neurosci. 2016, 36, 12027–12043. [Google Scholar] [CrossRef] [Green Version]
- Rzepnikowska, W.; Flis, K.; Muñoz-Braceras, S.; Menezes, R.; Escalante, R.; Żołądek, T. Yeast and other lower eukaryotic organisms for studies of Vps13 proteins in health and disease. Traffic 2017, 18, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Rzepnikowska, W.; Flis, K.; Kaminska, J.; Grynberg, M.; Urbanek, A.; Ayscough, K.R.; Żołądek, T. Amino acid substitution equivalent to human chorea-acanthocytosis I2771R in yeast Vps13 protein affects its binding to phosphatidylinositol 3-phosphate. Hum. Mol. Genet. 2017, 26, 1497–1510. [Google Scholar] [CrossRef] [Green Version]
- Dziurdzik, S.K.; Bean, B.D.M.; Davey, M.; Conibear, E. A VPS13D spastic ataxia mutation disrupts the conserved adaptor-binding site in yeast Vps13. Hum. Mol. Genet. 2020, 29, 635–648. [Google Scholar] [CrossRef] [Green Version]
- De, M.; Oleskie, A.N.; Ayyash, M.; Dutta, S.; Mancour, L.; Abazeed, M.; Brace, E.J.; Skiniotis, G.; Fuller, R.S. The Vps13p–Cdc31p complex is directly required for TGN late endosome transport and TGN homotypic fusion. J. Cell Biol. 2017, 216, 425–439. [Google Scholar] [CrossRef]
- Brickner, J.H.; Fuller, R.S. SOI1 encodes a novel, conserved protein that promotes TGN-endosomal cycling of Kex2p and other membrane proteins by modulating the function of two TGN localization signals. J. Cell Biol. 1997, 139, 23–36. [Google Scholar] [CrossRef]
- Redding, K.; Brickner, J.H.; Marschall, L.G.; Nichols, J.W.; Fuller, R.S. Allele-specific suppression of a defective trans-Golgi network (TGN) localization signal in Kex2p identifies three genes involved in localization of TGN transmembrane proteins. Mol. Cell. Biol. 1996, 16, 6208–6217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-S.; Neiman, A.M. VPS13 regulates membrane morphogenesis during sporulation in Saccharomyces cerevisiae. J. Cell Sci. 2012, 125, 3004–3011. [Google Scholar] [CrossRef] [Green Version]
- Soczewka, P.; Flis, K.; Tribouillard-Tanvier, D.; Di Rago, J.-P.; Santos, C.N.; Menezes, R.; Kaminska, J.; Żołądek, T. Flavonoids as Potential Drugs for VPS13-Dependent Rare Neurodegenerative Diseases. Genes 2020, 11, 828. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, R.; Dupre, S.; Avaro, S.; Haguenauer-Tsapis, R.; Słonimski, P.P.; Rytka, J. The novel protein Ccz1p required for vacuolar assembly in Saccharomyces cerevisiae functions in the same transport pathway as Ypt7p. J. Cell Sci. 2000, 113, 4301–4311. [Google Scholar]
- Alonso-Rodríguez, E.; Fernández-Piñar, P.; Sacristán-Reviriego, A.; Molina, M.; Martín, H. An Analog-sensitive Version of the Protein Kinase Slt2 Allows Identification of Novel Targets of the Yeast Cell Wall Integrity Pathway. J. Biol. Chem. 2016, 291, 5461–5472. [Google Scholar] [CrossRef] [Green Version]
- Levin, D.E. Regulation of Cell Wall Biogenesis in Saccharomyces cerevisiae: The Cell Wall Integrity Signaling Pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, J.; Okawa, Y.; Oyama, T.; Shimoda, N.; Uesugi, S.; Takagi, H.; Ito, Y.; Kimura, K. Inhibition of Calcineurin and Glycogen Synthase Kinase-3β by Ricinoleic Acid Derived from Castor Oil. Lipids 2020, 55, 89–99. [Google Scholar] [CrossRef]
- Goldman, A.; Roy, J.; Bodenmiller, B.; Wanka, S.; Landry, C.R.; Aebersold, R.; Cyert, M.S. The Calcineurin Signaling Network Evolves via Conserved Kinase-Phosphatase Modules that Transcend Substrate Identity. Mol. Cell 2014, 55, 422–435. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, T.; Nakamura, M.; Irie, K. Induction of Ptp2 and Cmp2 protein phosphatases is crucial for the adaptive response to ER stress in Saccharomyces cerevisiae. Sci. Rep. 2018, 8, 13078. [Google Scholar] [CrossRef]
- Francis, C.; Bai, Y. Differential expression of cyclosporine A-Induced calcineurin isoform-specific matrix metalloproteinase 9 (MMP-9) in renal fibroblasts. Biochem. Biophys. Res. Commun. 2018, 503, 2549–2554. [Google Scholar] [CrossRef]
- Takaishi, T.; Saito, N.; Kuno, T.; Tanaka, C. Differential distribution of the mRNA encoding two isoforms of the catalytic subunit of calcineurin in the rat brain. Biochem. Biophys. Res. Commun. 1991, 174, 393–398. [Google Scholar] [CrossRef]
- Kuno, T.; Mukai, H.; Ito, A.; Chang, C.-D.; Kishima, K.; Saito, N.; Tanaka, C. Distinct Cellular Expression of Calcineurin Aɑ and Aβ in Rat Brain. J. Neurochem. 1992, 58, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Usuda, N.; Arai, H.; Sasaki, H.; Hanai, T.; Nagata, T.; Muramatsu, T.; Kincaid, R.L.; Higuchi, S. Differential subcellular localization of neural isoforms of the catalytic subunit of calmodulin-dependent protein phosphatase (calcineurin) in central nervous system neurons: Immunohistochemistry on formalin-fixed paraffin sections employing antigen retrieval by microwave irradiation. J. Histochem. Cytochem. 1996, 44, 13–18. [Google Scholar] [CrossRef]
- Eastwood, S.L.; Salih, T.; Harrison, P.J. Differential expression of calcineurin A subunit mRNA isoforms during rat hippocampal and cerebellar development. Eur. J. Neurosci. 2005, 22, 3017–3024. [Google Scholar] [CrossRef]
- Porta, S.; Martí, E.; De La Luna, S.; Arbones, M.L. Differential expression of members of the RCAN family of calcineurin regulators suggests selective functions for these proteins in the brain. Eur. J. Neurosci. 2007, 26, 1213–1226. [Google Scholar] [CrossRef]
- Hilioti, Z.; Gallagher, D.A.; Low-Nam, S.T.; Ramaswamy, P.; Gajer, P.; Kingsbury, T.J.; Birchwood, C.J.; Levchenko, A.; Cunningham, K.W. GSK-3 kinases enhance calcineurin signaling by phosphorylation of RCNs. Genes Dev. 2004, 18, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Grigoriu, S.; Bond, R.; Cossio, P.; Chen, J.A.; Ly, N.; Hummer, G.; Page, R.; Cyert, M.S.; Peti, W. The Molecular Mechanism of Substrate Engagement and Immunosuppressant Inhibition of Calcineurin. PLoS Biol. 2013, 11, e1001492. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cheng, N.; Wang, P.; Li, J.; Jia, A.; Li, W.; Zhang, N.; Yin, Y.; Tong, L.; Wei, Q.; et al. A novel peptide exerts potent immunosuppression by blocking the two-site interaction of NFAT with calcineurin. J. Biol. Chem. 2020, 295, 2760–2770. [Google Scholar] [CrossRef]
- Noguchi, H.; Sugimoto, K.; Miyagi-Shiohira, C.; Nakashima, Y.; Kobayashi, N.; Saitoh, I.; Watanabe, M.; Noguchi, Y. RCAN-11R peptide provides immunosuppression for fully mismatched islet allografts in mice. Sci. Rep. 2017, 7, 3043. [Google Scholar] [CrossRef] [Green Version]
- James, P.; Halladay, J.; Craig, E.A. Genomic libraries and a host strain designed for highly efficient two-hybrid selection in yeast. Genetics 1996, 144, 1425–1436. [Google Scholar] [CrossRef]
- Stettler, S.; Chiannilkulchai, N.; Denmat, S.H.-L.; Lalo, D.; Lacroute, F.; Sentenac, A.; Thuriaux, P. A general suppressor of RNA polymerase I, II and III mutations in Saccharomyces cerevisiae. Mol. Genet. Genom. 1993, 239, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Gietz, R.; Akio, S. New yeast-Escherichia coli shuttle vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction sites. Gene 1988, 74, 527–534. [Google Scholar] [CrossRef]
- Budhiraja, R.; Hermkes, R.; Müller, S.; Schmidt, J.; Colby, T.; Panigrahi, K.; Coupland, G.; Bachmair, A. Substrates Related to Chromatin and to RNA-Dependent Processes Are Modified by Arabidopsis SUMO Isoforms That Differ in a Conserved Residue with Influence on Desumoylation. Plant Physiol. 2009, 149, 1529–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminska, J.; Wysocka-Kapcińska, M.; Rooij, I.S.-D.; Rytka, J.; Żołądek, T. Pan1p, an actin cytoskeleton-associated protein, is required for growth of yeast on oleate medium. Exp. Cell Res. 2005, 310, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, M.P.; Schatz, G. Two nuclear mutations that block mitochondrial protein import in yeast. Proc. Natl. Acad. Sci. USA 1984, 81, 4819–4823. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wardaszka, P.; Soczewka, P.; Sienko, M.; Zoladek, T.; Kaminska, J. Partial Inhibition of Calcineurin Activity by Rcn2 as a Potential Remedy for Vps13 Deficiency. Int. J. Mol. Sci. 2021, 22, 1193. https://doi.org/10.3390/ijms22031193
Wardaszka P, Soczewka P, Sienko M, Zoladek T, Kaminska J. Partial Inhibition of Calcineurin Activity by Rcn2 as a Potential Remedy for Vps13 Deficiency. International Journal of Molecular Sciences. 2021; 22(3):1193. https://doi.org/10.3390/ijms22031193
Chicago/Turabian StyleWardaszka, Patrycja, Piotr Soczewka, Marzena Sienko, Teresa Zoladek, and Joanna Kaminska. 2021. "Partial Inhibition of Calcineurin Activity by Rcn2 as a Potential Remedy for Vps13 Deficiency" International Journal of Molecular Sciences 22, no. 3: 1193. https://doi.org/10.3390/ijms22031193