
The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa


Abstract
:1. Introduction
2. Results
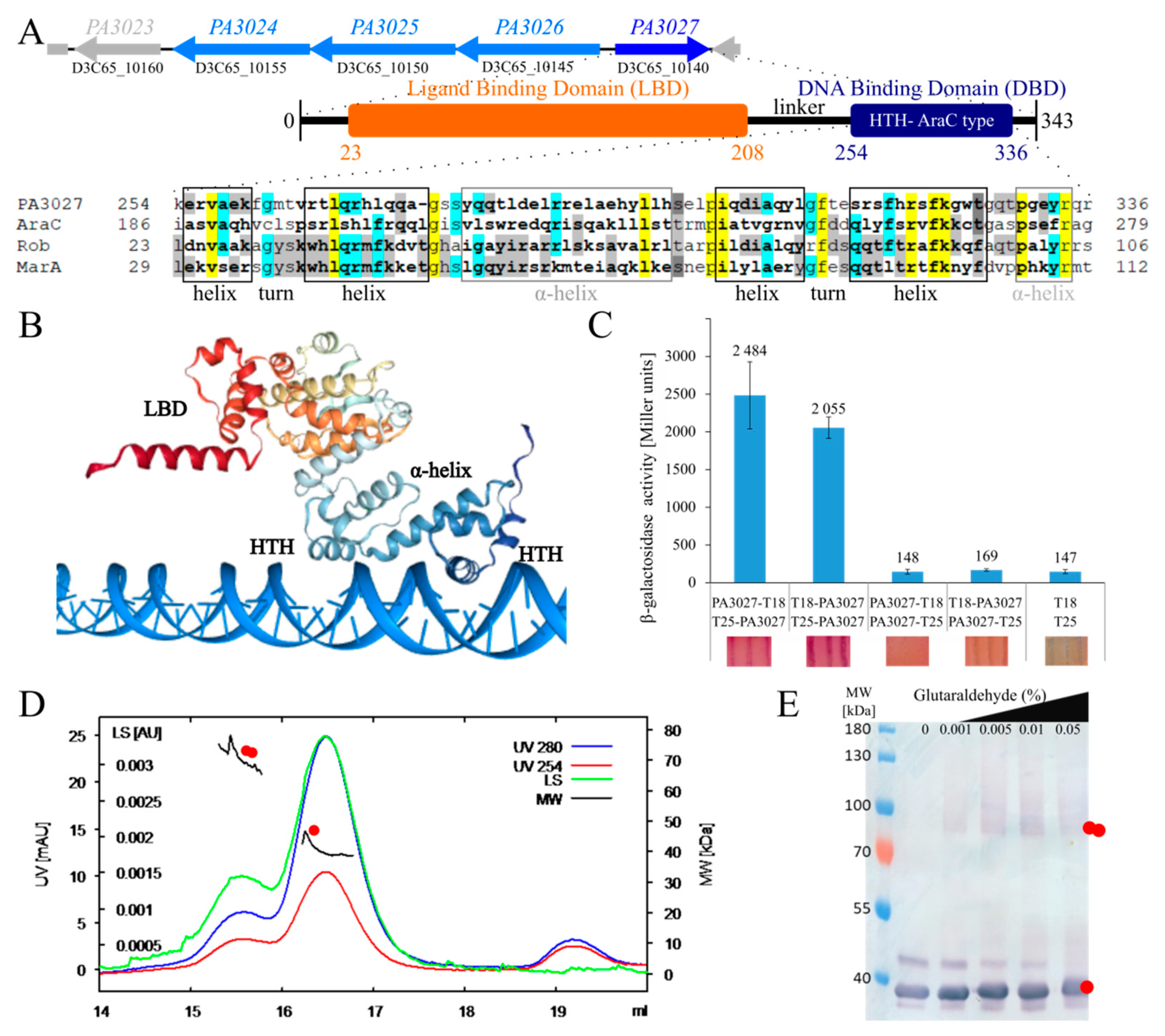
2.1. Overview of the PA3027 from P. aeruginosa PAO1161
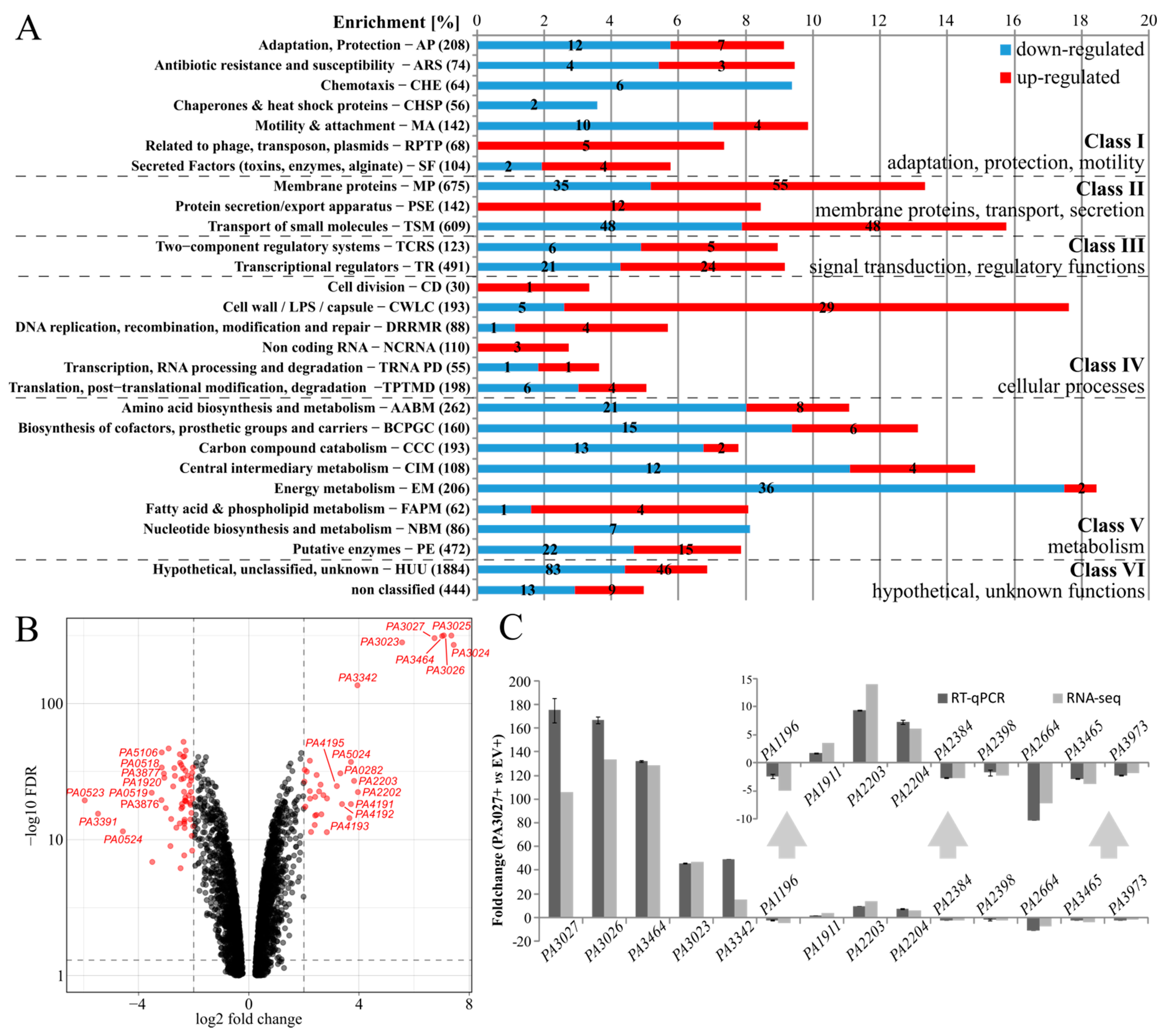
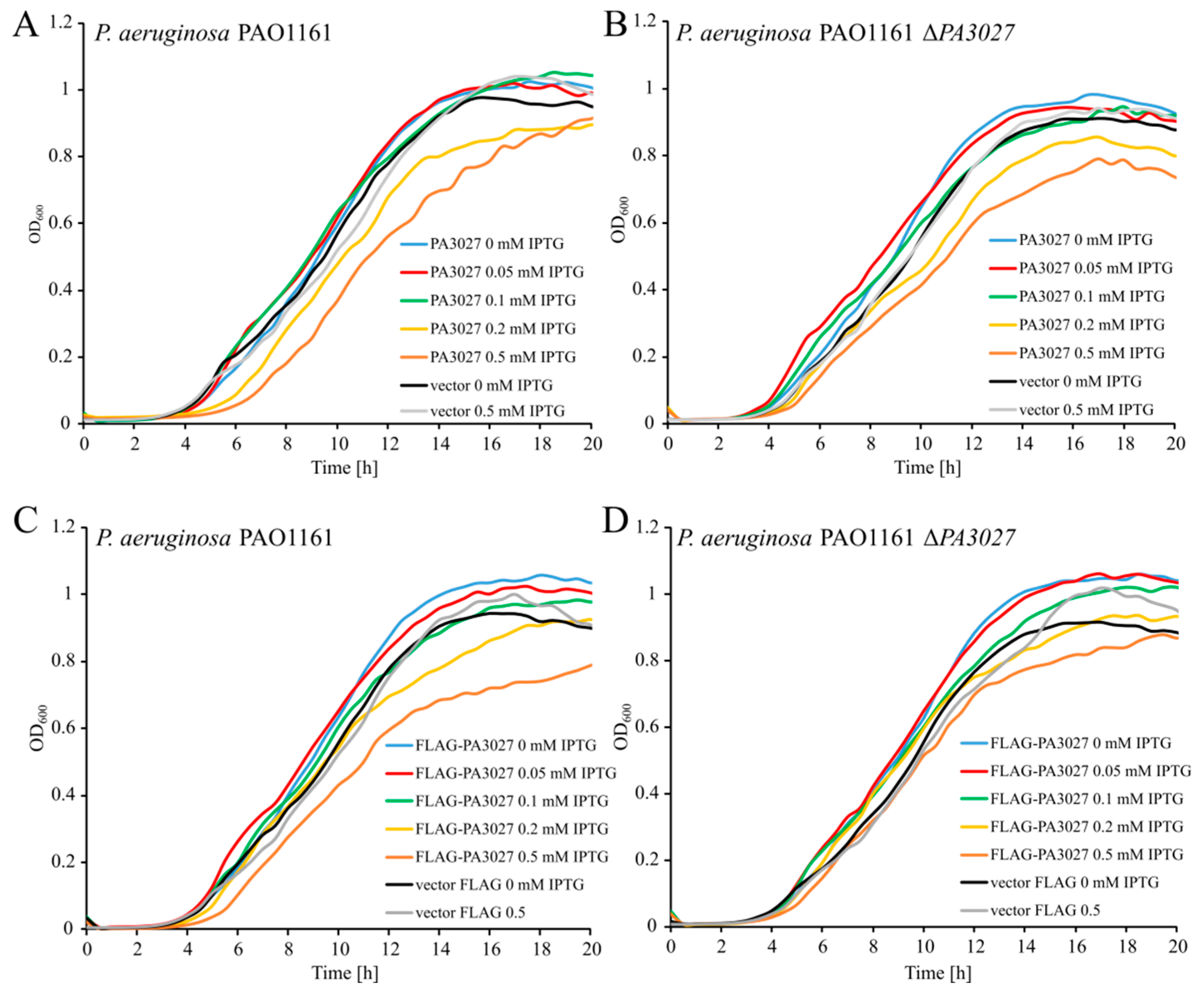
2.2. Effect of Increased PA3027 Level on Gene Expression
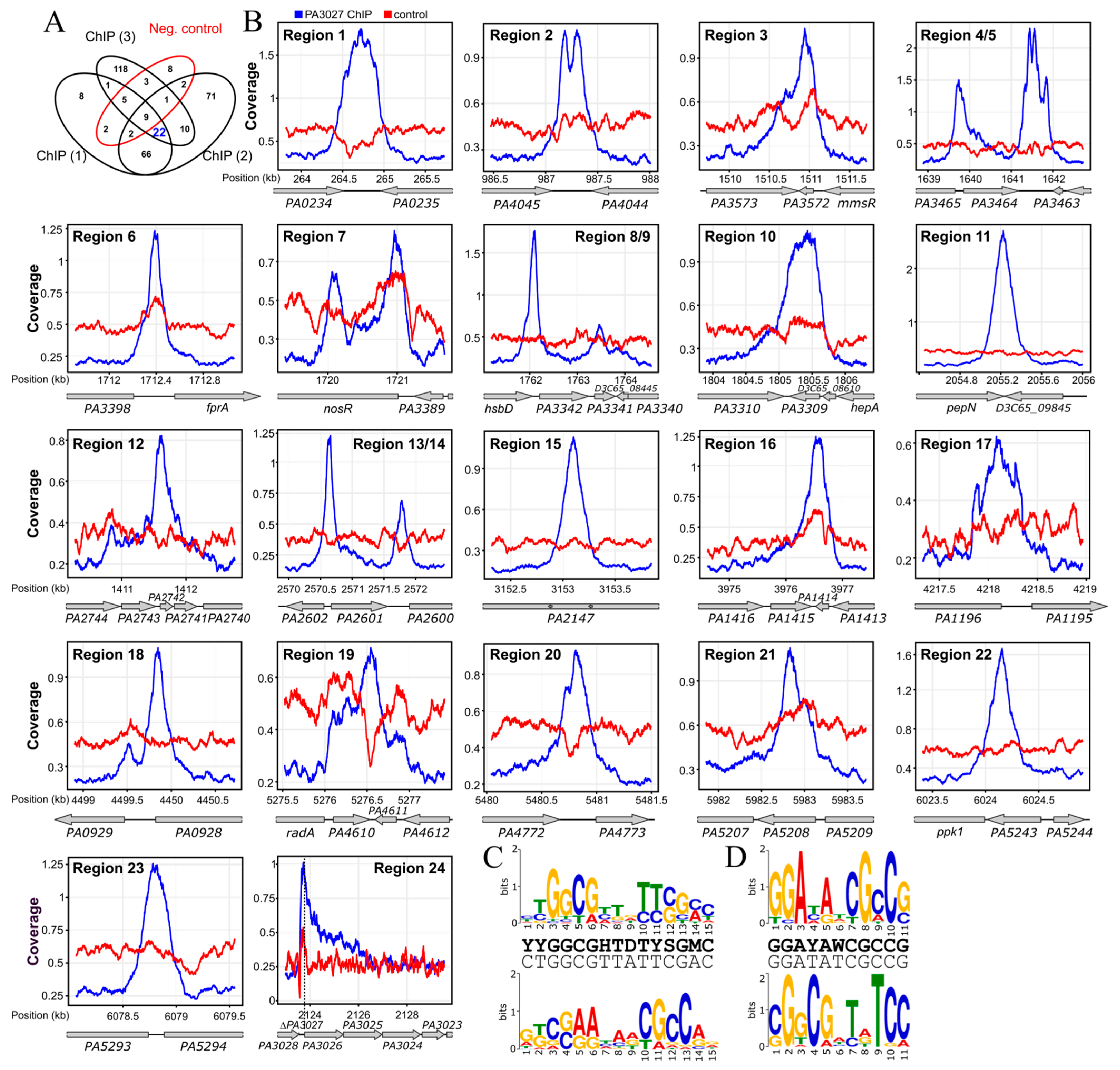
2.3. Identification of PA3027 Binding Sites in P. aeruginosa
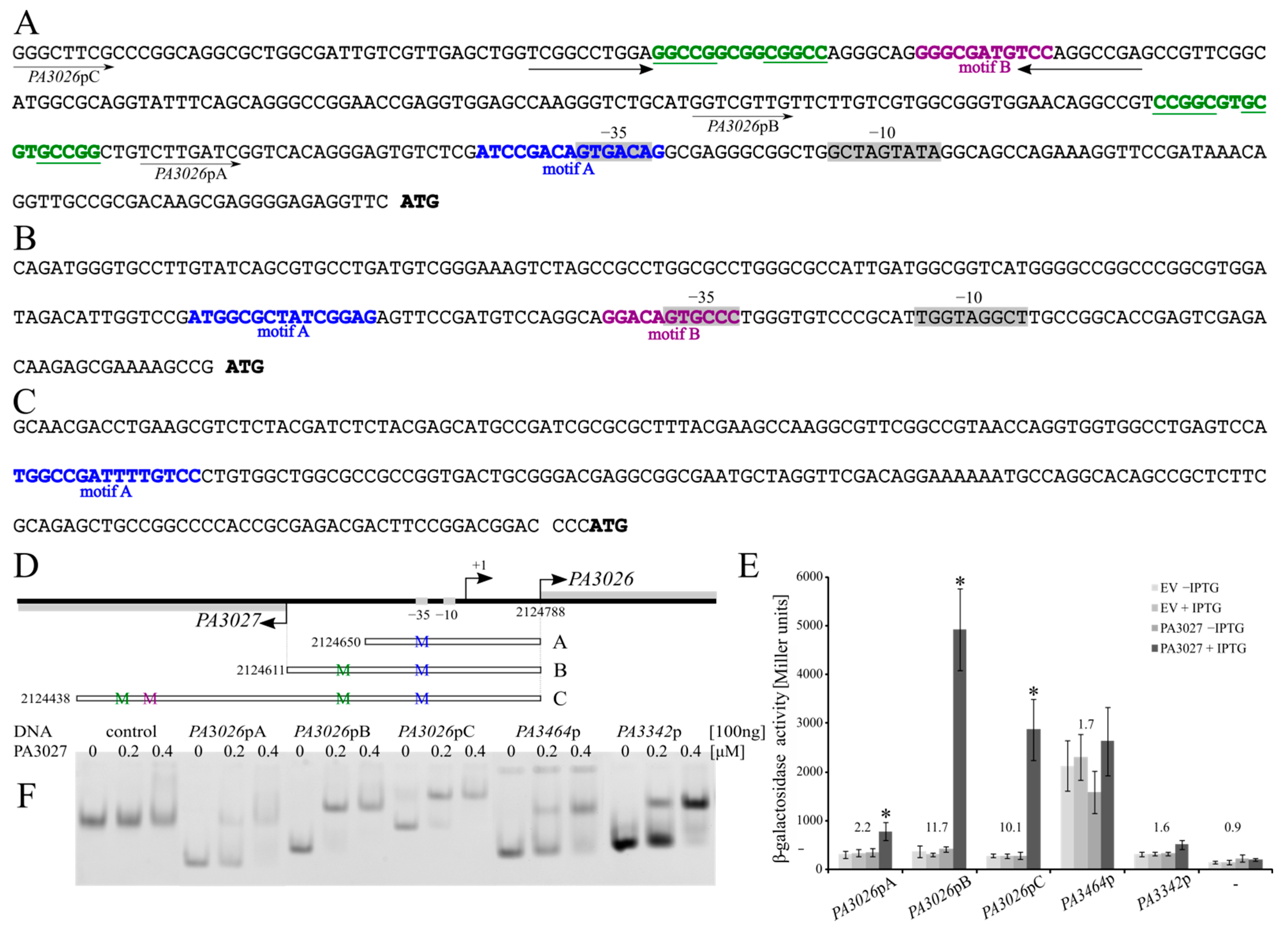
2.4. Regulatory Properties of PA3027
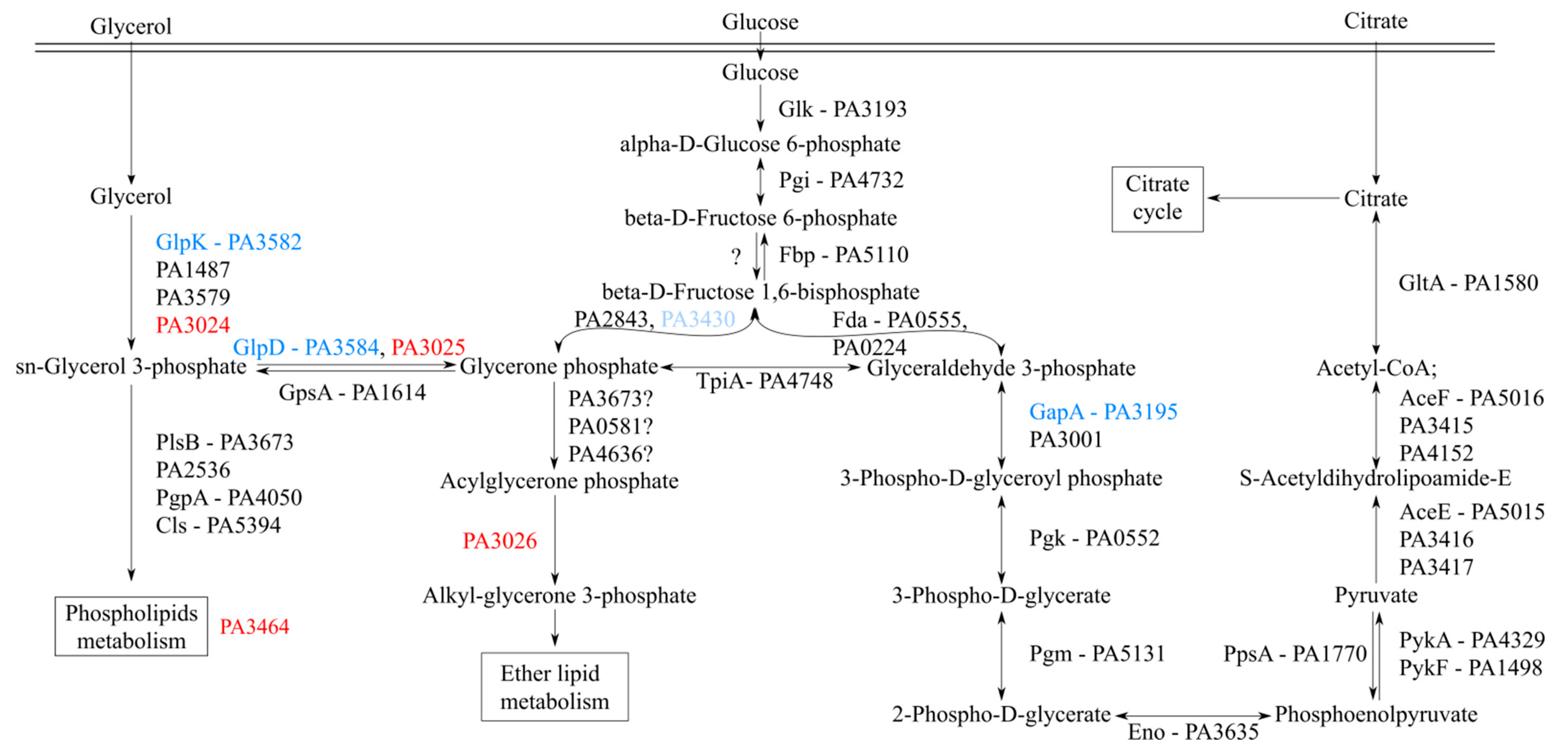
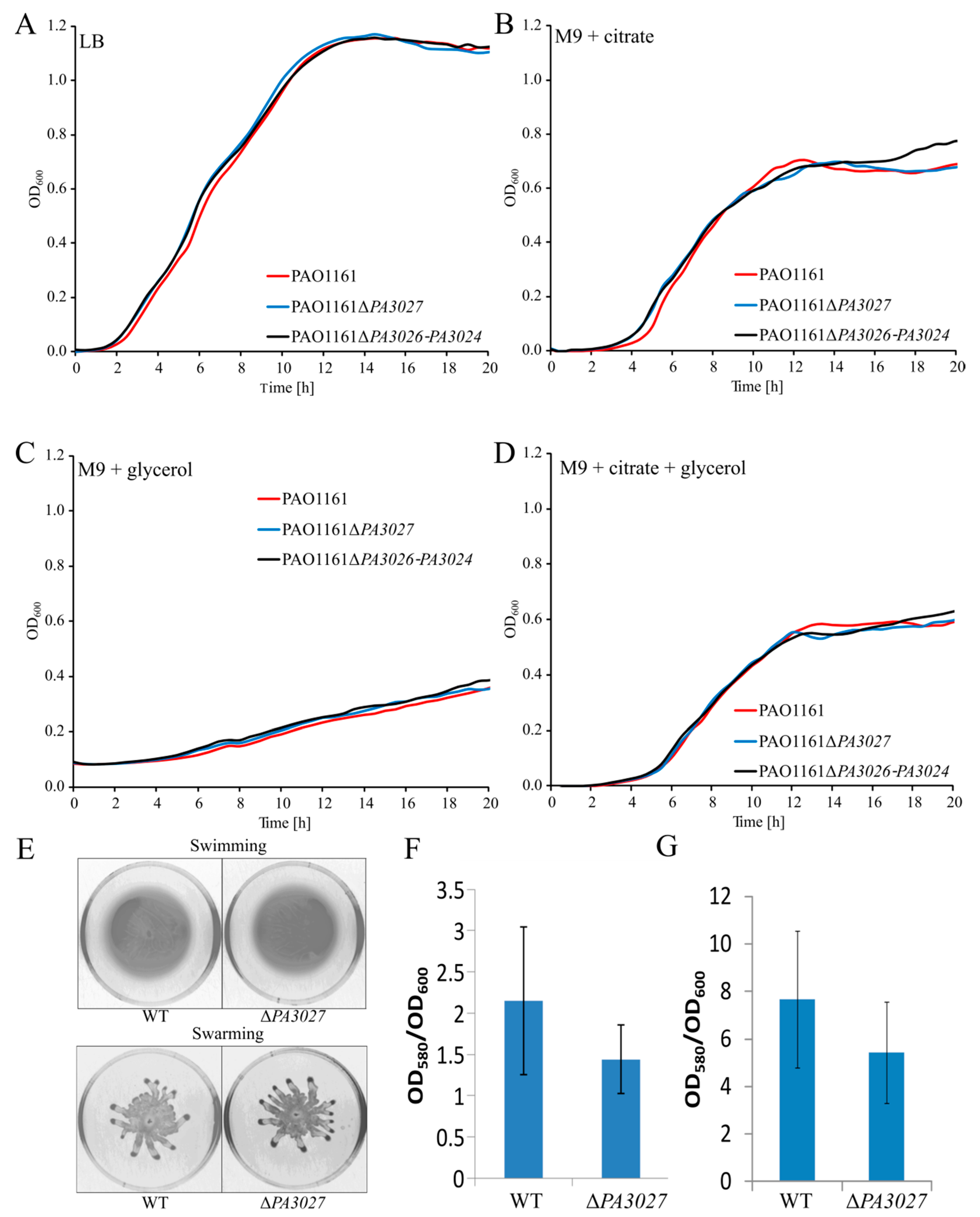
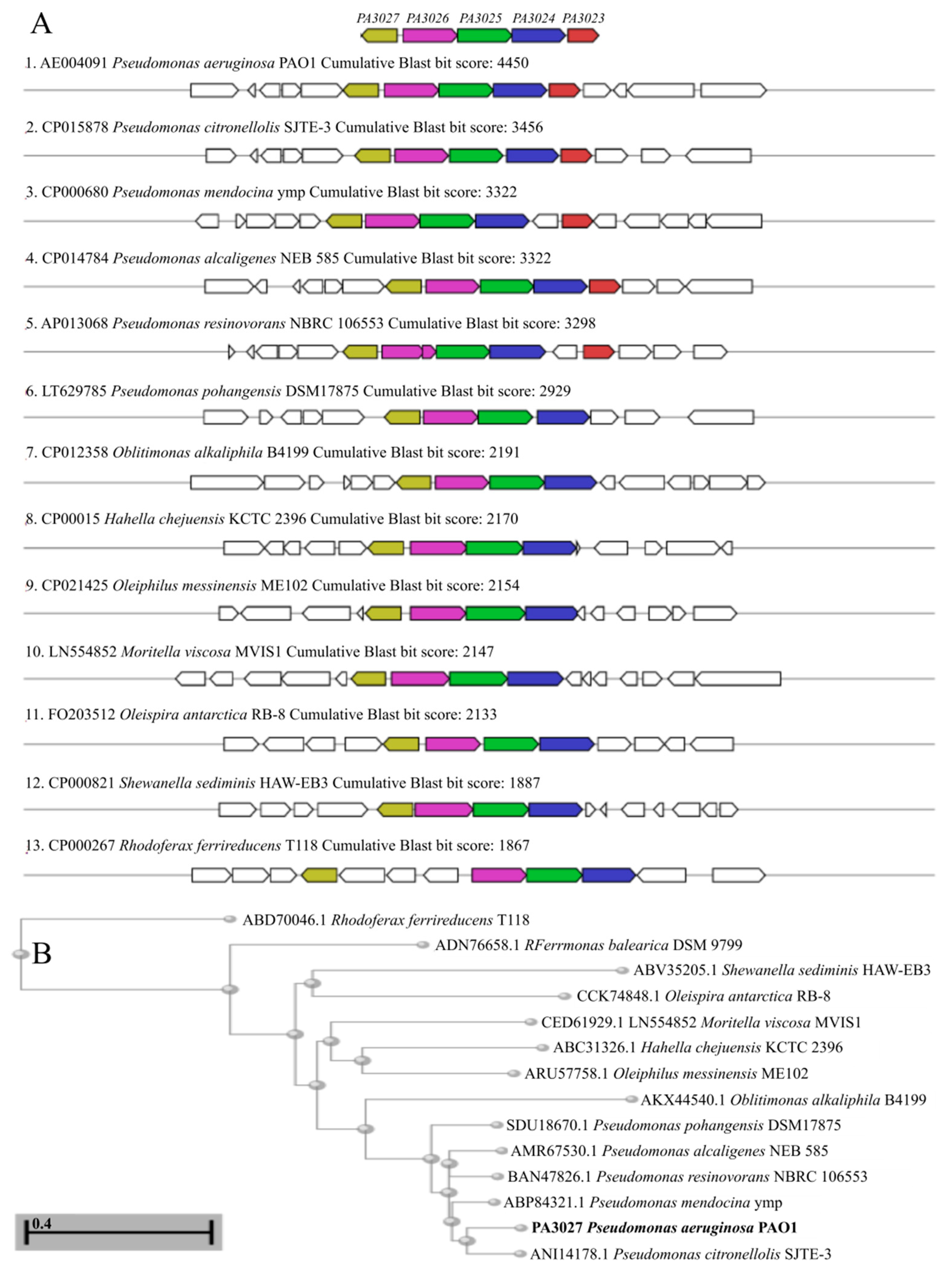
2.5. Toward the Biological Function of PA3027-PA3026-PA3023 Gene Cluster
3. Discussion
4. Materials and Methods
4.1. Growth Conditions Bacterial Strains and Plasmids
4.2. Vectors and Strains Construction
4.3. Protein Overproduction and Purification
4.4. SEC-MALS Analysis
4.5. Glutaraldehyde Crosslinking
4.6. RNA Isolation, RNA-Seq, and RT-qPCR Analysis
4.7. Chromatin Immunoprecipitation with Sequencing
4.8. Regulatory Experiments
4.9. Electrophoretic Mobility Shift Assay (EMSA)
4.10. Bacterial Two-Hybrid Analysis (BATCH)
4.11. Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Number | Strand | Start | p-Value | Site |
|---|---|---|---|---|
| Motif A | ||||
| 20 | + | 71 | 9.60 × 10−8 | CTGGCGCTATTCGCC |
| 2 | + | 72 | 5.89 × 10−7 | GTGGCGTTATTCGCC |
| 1 | + | 10 | 2.01 × 10−6 | CCGGCGTTATCCGCC |
| 9 | + | 47 | 5.80 × 10−6 | TTGGTGTTGTTGCAT |
| 7 | + | 37 | 5.80 × 10−6 | CTGGCGCAGTTGGAC |
| 5 | + | 2 | 7.42 × 10−6 | CGGGCGTTATCCGCC |
| 15 | + | 34 | 1.32 × 10−5 | ATGGCGCGATCGGAC |
| 11 | + | 40 | 1.32 × 10−5 | CCGGCGAAATTCGCC |
| 12 | − | 28 | 1.48 × 10−5 | TTGGCATGTTCGGAT |
| 21 | + | 60 | 2.78 × 10−5 | TTGGCGACTTTGTCC |
| 16 | + | 22 | 2.78 × 10−5 | CCGGCGTTGCTCGTC |
| 4 | + | 36 | 4.53 × 10−5 | ATGGCGCTATCGGAG |
| 17 | − | 23 | 7.14 × 10−5 | GCGCCGATTTTCGAC |
| 10 | + | 4 | 7.14 × 10−5 | TCGACGTTGCTCGCC |
| 23 | − | 62 | 7.79 × 10−5 | CTGGCATGGCTCCAC |
| 18 | + | 14 | 7.79 × 10−5 | CTGGCGCCGCTGGCC |
| 24 | − | 17 | 7.79 × 10−5 | CTGTCACTGTCGGAT |
| 8 | + | 39 | 1.62 × 10−4 | TGGCCGATTTTGTCC |
| 13 | − | 44 | 2.03 × 10−4 | ATTGCATTTCTCGCA |
| 19 | + | 54 | 2.18 × 10−4 | TCGTCGCGGTCGGAC |
| 3 | − | 83 | 2.35 × 10−4 | CTGGCAAGTCTGCCT |
| 22 | − | 16 | 3.31 × 10−4 | TCGGTGATCCTCGAA |
| 6 | + | 9 | 4.03 × 10−4 | ATCGTGTTATCCCAT |
| 14 | − | 70 | 4.29 × 10−4 | TGGGTGCTTTTCTTA |
| Motif B | ||||
| 2 | + | 61 | 7.20 × 10−7 | GGATAACGCCG |
| 1 | − | 11 | 7.20 × 10−7 | GGATAACGCCG |
| 21 | − | 40 | 1.90 × 10−6 | GGACATCGCCG |
| 20 | + | 60 | 3.50 × 10−6 | GGATAGCGCCG |
| 5 | − | 3 | 4.83 × 10−6 | GGATAACGCCC |
| 24 | − | 43 | 6.79 × 10−6 | GGACATCGCCC |
| 16 | − | 33 | 1.22 × 10−5 | GGAAAACGACG |
| 18 | + | 49 | 1.48 × 10−5 | GGATACCGCCG |
| 17 | + | 77 | 1.84 × 10−5 | GGAAATCGTCG |
| 13 | + | 83 | 2.31 × 10−5 | TGACATCGACG |
| 7 | + | 24 | 3.02 × 10−5 | TGATGTCGCCG |
| 19 | + | 19 | 3.88 × 10−5 | GCACATCGCCG |
| 22 | + | 51 | 7.02 × 10−5 | GGATGGCGCCC |
| 4 | + | 69 | 8.23 × 10−5 | GGACAGTGCCC |
| 3 | + | 60 | 9.79 × 10−5 | GGAAGTTGACG |
| Strains | ||
|---|---|---|
| E. coli DH5α | F− (φ80d lacZΔM15) recA1 endA1 gyrA96 thi-1 hsdR17 (rk−mk+) supE44 relA1 deoR Δ(lacZYA–argF)U196 | [100] |
| E. coli BL21 | F− ompT hsdS (rB−mB−) gal dcm (λ DE3) | [100] |
| E. coli S17-1 | pro hsdR hsdM recA TpR SmRΩRP4-Tc::Mu-Km::Tn7 | [100] |
| E. coli BTH101 | F−cya−99 araD139 galE15 galK16 rpsL1 (Strr) hsdR2 mcrA1 mcrB1 | [83] |
| E. coli DH5α Δlac | F− (φ80d lacZΔM15) recA1 endA1 gyrA96 thi-1 hsdR17 (rk-mk+) supE44 relA1 deoR Δ(lacZYA–argF)U196 | lab collection |
| P. aeruginosa PAO1161 | leu r−m+ RifR | [101] |
| P. aeruginosa PAO1161 ΔPA3027 | deletion of 1015 bp fragment encompassing PA3027 gene | this work |
| P. aeruginosa PAO1161 ΔPA3026–PA3024 | deletion of 4711 bp fragment encompassing PA3026-PA3024 operon | this work |
| Plasmids | ||
| pAKE600 | ApR; oriMB1, oriTRK2, sacB | [80] |
| pKKB1.61 | ApR; pAKE600 derivative with 431 bp fragment encompassing up- and down- sequence of PA3027, cloned using BamHI-HindIII-EcoRI and 1#/2# and 3#/4# primers | this work |
| pKKB1.62 | ApR; pAKE600 derivative with 472bp fragment encompassing up- and down- sequence of PA3026–PA3024, cloned using BamHI, HindIII/HindIII, EcoRI EcoRI and 4#/5# and 6#/7# primers | this work |
| pET28a(+) | KmR; oripBR322; orif1; expression vector | Novagen |
| pKKB1.21 | pET28a(+) derivative with his6–PA3027, PA3027 cloned 1078 bp as EcoRI, SacI | this work |
| pBBR1-MCS1 | CmR; IncA/C broad-host-range cloning vector, lacZα–MCS, mob, T7p, T3p | [81] |
| pAMB9.37 | CmR, oriCIncA/C, pBBR1-MCS-1 derivative with lacIQ–tacp, expression vector | [82] |
| pABB28.1 | CmR, oriCIncA/C, pBBR1-MCS-1 derivative with lacIQ–tacp–flag expression vector | this work |
| pKKB1.11 | pAMB9.37 (lacIQ–tacp) derivative with 1097 bp fragment, encoding PA3027 gene, cloned using EcoRI, SalI | this work |
| pKKB1.12 | pABB28.1 (lacIQ–tacp–flag) derivative with 1097 bp flag–PA3027 fragment cloned using EcoRI, SalI | this work |
| pLKB2 | KmR, orip15, pKT25 modified with lacp–cyaT25–MCS | [83,102] |
| pKNT25 | KmR; orip15, lacp–MCS–cyaT25, | [83] |
| pUT18 | ApR; oriColE1, lacp–MCS–cyaT18, | [83] |
| pLKB4 | ApR, oriColE1, pUT18C modified withlacp–cyaT18–MCS | [83,102] |
| pKKB1.51 | pKNT25 derivative with PA3027–cyaT25; 1042 bp PA3027 fragment cloned using EcoRI, SacI enzymes | this work |
| pKKB1.52 | pLKB2 derivative with cyaT25–PA3027; 1092 bp PA3027 fragment cloned using EcoRI, Ecl136II/SmaI enzymes | this work |
| pKKB1.81 | pUT18 derivative with PA3027–cyaT18; 1042 bp PA3027 fragment cloned using EcoRI, SacI enzymes | this work |
| pKKB1.82 | pLKB4 derivative with cyaT18–PA3027; 1092 bp PA3027 fragment cloned using EcoRI, SacI enzymes | this work |
| pCM132 | KmR; oriVRK2; oriTRK2; promoter-less lacZ reporter gene | [47] |
| pKKB1.305 | pCM132 derivative with PA3026pA, 120 bp fragment amplified with primers 14#/13# cloned using EcoRI, BamHI/BglII | this work |
| pKKB1.303 | pCM132 derivative with PA3026pB, 176 bp fragment amplified with primers 12#/13# cloned using EcoRI, BamHI/BglII | this work |
| pKKB1.304 | pCM132 derivative with PA3026pC, 330 bp fragment amplified with primers 15#/13# cloned using EcoRI, BamHI/BglII | this work |
| pKKB1.307 | pCM132 derivative with PA3464p, 228 bp fragment amplified with primers 16#/17# cloned using BglII | this work |
| pKKB1.309 | pCM132 derivative with PA3342p, 167 bp fragment amplified with primers 18#/19# cloned using EcoRI, BamHI/BglII | this work |
| Nr | Starter | Used for: | Sequence |
|---|---|---|---|
| #1 | 3027mLF | PA3027 gene deletion | gcggatcCAATTCGACCACGGTGCTTTC |
| #2 | 3027mLR | gcaagcttGGTCTGCATGGTCGTTGTTC | |
| #3 | 3027mPF | gcaagctttagtAATGAGAACGGCGGCCATCCG | |
| #4 | 3027mPR | gcgaattcCGGTGCTCTATCCGAACCAGAGTTCC | |
| #5 | 3026-4mLF | PA3026–PA3024 operon deletion | gcggatccCATGGCGCAGGTATTTCAGC |
| #6 | 3026-4mLR | gcaagcttACGACGCATGAACCTCTCC | |
| #7 | 3026-4mPF | gcaagctttagtaaTGAACCGGGCGCCGCACTTCC | |
| #8 | 3026-4mPR | gcgaattcTCGCCGTCCTCCCAGGTTAC | |
| #9 | 3027eF | PA3027 expression | gcgaattcATGCAGACCCTTGGCTCCAC |
| #10 | 3027eR | gcgaGcTCAGCGAACTGCTCGATTG | |
| #11 | 3027eR2 | gcgagctcgTTGCGCCGCCGGCTCCTTGC | |
| #12 | 3027pdF | PA3026 promoter | cagaattcgcatgcGGTCGTTGTTCTTGTCGTGGCGGG |
| #13 | p3026R2 | caggatccGAACCTCTCCCCTCGCTTGT | |
| #14 | p3026F2 | cagaattcgcatgcTCTTGATCGGTCACAGGGAG | |
| #15 | p3026F3 | cagaattcgcatgcGGGCTTCGCCCGGCAGGCGC | |
| #16 | p3464R | PA3464 promoter | gcagatctCGGCTTTTCGCTCTTGTCTC |
| #17 | p3464F | gcagatctCAGATGGGTGCCTTGTATCA | |
| #18 | p3342F | PA3342 promoter | cagaattcgcatgcGCAACGACCTGAAGCGTCTC |
| #19 | p3342R | caggatccGTCCGTCCGGAAGTCGTCTC | |
| #20 | CM132RCy5 | EMSA analysis | Cy5—CTTCCACAGTAGTTCACCACC |
| #21 | CM132pF | GTGAACGCTCTCCTGAGTAG | |
| #22 | 3027qF | RT-qPCR | CTGGATCGCCGACCTGGAAG |
| #23 | 3027qR | CCGGACAGCCGAAGAAGGTC | |
| #24 | 3026qF | AATCGCTACCTTCCCGGCATCC | |
| #25 | 3026qR | ATGATCCCGAAGCGTCCCTCAG | |
| #26 | 3464qF | CAACCTGTTCAGCGACAACC | |
| #27 | 3464qR | TAGAAGCCCATGTGGAACGG | |
| #28 | 3023qF | CTACCTGTTGACCGGGTTGA | |
| #29 | 3023qR | CCTTCCCACTGGAAGTCCG | |
| #30 | 3342qF | GAGAACCGTATGGCTCACCG | |
| #31 | 3342qR | TACACAGGCACATCGGCTG | |
| #32 | 2203qF | TCTTCTGGTACTTCGGCGTT | |
| #33 | 2203qR | GAGGAACTCGAAGGACGGC | |
| #34 | 2204qF | CCCCTGCCAAACTCCCTTC | |
| #35 | 2204qR | GATCAGGGCCTTGCAATGTG | |
| #36 | 1911qF | AGATCTTCGTCGATAGCCGC | |
| #37 | 1911qR | CATCTCCTGGCGTACGTTGA | |
| #38 | 2398qF | CGACCAACCCCGCCATCACC | |
| #39 | 2398qR | CCAGCCTGAACCTGCCAGCC | |
| #40 | 3973qF | GGATCCTGAAGTCGACGAGC | |
| #41 | 3973qR | GAAAGCTGGAATGCGCCAC | |
| #42 | 2384qF | CGTCTCTCCGAAACCGGTAC | |
| #43 | 2384qR | TGTAGACATCCTGTTCGTCGAG | |
| #44 | 3465qF | CTGGTGATGATGCGCTCCTT | |
| #45 | 3465qR | AGCATGTAGCCGTAGATCGC | |
| #46 | 1196qF | TGGAAAGTACCCTGTTCGGC | |
| #47 | 1196qR | ATCTCGTCGAGGAACAAGCTG | |
| #48 | 2664qF | CCGAGGGATTGCTGAGCCGC | |
| #49 | 2664qR | GCCATGAAAGGCCGGGGTCC | |
| #50 | nadBF | CTACCTGGACATCAGCCACA | |
| #51 | nadBR | GGTAATGTCGATGCCGAAGT |
References
- Perez-Rueda, E.; Hernandez-Guerrero, R.; Martinez-Nuñez, M.A.; Armenta-Medina, D.; Sanchez, I.; Ibarra, J.A. Abundance, diversity and domain architecture variability in prokaryotic DNA-binding transcription factors. PLoS ONE 2018, 13, e0195332. [Google Scholar] [CrossRef] [Green Version]
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef] [PubMed]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete genome sequence of Pseudomonas aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Shao, X.; Xie, Y.; Wang, T.; Zhang, Y.; Wang, X.; Deng, X. An integrated genomic regulatory network of virulence-related transcriptional factors in Pseudomonas aeruginosa. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, M.T.; Schleif, R.; Bairoch, A.; Hofmann, K.; Ramos, J.L. Arac/XylS family of transcriptional regulators. Microbiol. Mol. Biol. Rev. 1997, 61, 393–410. [Google Scholar] [PubMed]
- Lee, N.; Wilcox, G.; Gielow, W.; Arnold, J.; Cleary, P.; Englesberg, E. In Vitro activation of the transcription of araBAD operon by AraC activator. Proc. Natl. Acad. Sci. USA 1974, 71, 634–638. [Google Scholar] [CrossRef] [Green Version]
- Schleif, R. Regulation of the L-arabinose operon of Escherichia coli. Trends Genet. 2000, 16, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Schleif, R. AraC protein, regulation of the L-arabinose operon in Escherichia coli, and the light switch mechanism of AraC action. FEMS Microbiol. Rev. 2010, 34, 779–796. [Google Scholar] [CrossRef] [Green Version]
- Caron, K.; Trowell, S.C. Highly sensitive and selective biosensor for a disaccharide based on an AraC-like transcriptional regulator transduced with bioluminescence resonance energy transfer. Anal. Chem. 2018, 90, 12986–12993. [Google Scholar] [CrossRef] [Green Version]
- Willsey, G.G.; Wargo, M.J. Sarcosine catabolism in Pseudomonas aeruginosa is transcriptionally regulated by SouR. J. Bacteriol. 2016, 198, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Gendlina, I.; Gutman, D.M.; Thomas, V.; Collins, C.M. Urea-dependent signal transduction by the virulence regulator UreR. J. Biol. Chem. 2002, 277, 37349–37358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprušanský, O.; Řežuchová, B.; Homerová, D.; Kormanec, J. Expression of the gap gene encoding glyceraldehyde-3-phosphate dehydrogenase of streptomyces aureofaciens requires GapR, a member of the AraC/XylS family of transcriptional activators. Microbiology 2001, 147, 1291–1301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francis, M.S.; Wolf-Watz, H.; Forsberg, A. Regulation of type III secretion systems. Curr. Opin. Microbiol. 2002, 5, 166–172. [Google Scholar] [CrossRef]
- Pletzer, D.; Schweizer, G.; Weingart, H. AraC/XylS family stress response regulators Rob, SoxS, PliA, and OpiA in the fire blight pathogen Erwinia amylovora. J. Bacteriol. 2014, 196, 3098–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hu, L.; Qiu, Y.; Osei-Adjei, G.; Tang, H.; Zhang, Y.; Zhang, R.; Sheng, X.; Xu, S.; Yang, W.; et al. QsvR integrates into quorum sensing circuit to control Vibrio parahaemolyticus virulence. Environ. Microbiol. 2019, 21, 1054–1067. [Google Scholar] [CrossRef]
- Hebert, M.D.; Houghton, J.E. Regulation of ornithine utilization in Pseudomonas aeruginosa (PAO1) is mediated by a transcriptional regulator, OruR. J. Bacteriol. 1997, 179, 7834–7842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.M.; Lu, C.D.; Abdelal, A.T. Cloning and characterization of argR, a gene that participates in regulation of arginine biosynthesis and catabolism in Pseudomonas aeruginosa PAO1. J. Bacteriol. 1997, 179, 5300–5308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.-K.; Im, S.-J.; Yeom, D.-H.; Lee, J.-H. AntR-mediated bidirectional activation of antA and antR, anthranilate degradative genes in Pseudomonas aeruginosa. Gene 2012, 505, 146–152. [Google Scholar] [CrossRef]
- Steele, M.I.; Lorenz, D.; Hatter, K.; Park, A.; Sokatch, J.R. Characterization of the mmsAB operon of Pseudomonas aeruginosa PAO encoding methylmalonate-semialdehyde dehydrogenase and 3-hydroxyisobutyrate dehydrogenase. J. Biol. Chem. 1992, 267, 13585–13592. [Google Scholar] [CrossRef]
- Michel, L.; González, N.; Jagdeep, S.; Nguyen-Ngoc, T.; Reimmann, C. PchR-box recognition by the AraC-type regulator PchR of Pseudomonas aeruginosa requires the siderophore pyochelin as an effector. Mol. Microbiol. 2005, 58, 495–509. [Google Scholar] [CrossRef]
- Sánchez, D.G.; Primo, E.D.; Damiani, M.T.; Lisa, A.T. Pseudomonas aeruginosa gbdR gene is transcribed from a σ54-dependent promoter under the control of NtrC/CbrB, IHF and BetI. Microbiology 2017, 163, 1343–1354. [Google Scholar] [CrossRef] [PubMed]
- Meadows, J.A.; Wargo, M.J. Transcriptional regulation of carnitine catabolism in Pseudomonas aeruginosa by CdhR. mSphere 2018, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, R.; Feng, X.; Wei, X.; Pan, X.; Liu, C.; Song, R.; Jin, Y.; Bai, F.; Jin, S.; Wu, W.; et al. PutA is required for virulence and regulated by PruR in Pseudomonas aeruginosa. Front. Microbiol 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Whitchurch, C.B.; Leech, A.J.; Young, M.D.; Kennedy, D.; Sargent, J.L.; Bertrand, J.J.; Semmler, A.B.T.; Mellick, A.S.; Martin, P.R.; Alm, R.A.; et al. Characterization of a complex chemosensory signal transduction system which controls twitching motility in Pseudomonas aeruginosa. Mol. Microbiol. 2004, 52, 873–893. [Google Scholar] [CrossRef]
- Lee, E.J.; Evans, D.J.; Fleiszig, S.M.J. Role of Pseudomonas aeruginosa ExsA in penetration through corneal epithelium in a novel in vivo model. Investig. Ophthalmol. Vis. Sci. 2003, 44, 5220–5227. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.-H.; Zhang, X.-F.; Xu, J.-L.; Tan, A.-T.; Zhang, L.-H. VqsM, a novel AraC-type global regulator of quorum-sensing signalling and virulence in Pseudomonas aeruginosa. Mol. Microbiol. 2005, 58, 552–564. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Deng, X.; Li, X.; Ye, Y.; Wu, M. Molecular mechanisms of master regulator VqsM mediating quorum-sensing and antibiotic resistance in Pseudomonas aeruginosa. Nucleic Acids Res. 2014, 42, 10307–10320. [Google Scholar] [CrossRef]
- Zhao, J.; Yu, X.; Zhu, M.; Kang, H.; Ma, J.; Wu, M.; Gan, J.; Deng, X.; Liang, H. Structural and molecular mechanism of CdpR involved in quorum-sensing and bacterial virulence in Pseudomonas aeruginosa. PLoS Biol. 2016, 14. [Google Scholar] [CrossRef] [Green Version]
- LaBauve, A.E.; Wargo, M.J. Detection of host-derived sphingosine by Pseudomonas aeruginosa is important for survival in the murine lung. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Juarez, P.; Jeannot, K.; Plésiat, P.; Llanes, C. Toxic electrophiles induce expression of the multidrug efflux pump MexEF-OprN in Pseudomonas aeruginosa through a novel transcriptional regulator, CmrA. Antimicrob Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Bartosik, A.A.; Glabski, K.; Jecz, P.; Mikulska, S.; Fogtman, A.; Koblowska, M.; Jagura-Burdzy, G. Transcriptional profiling of parA and parB mutants in actively dividing cells of an opportunistic human pathogen Pseudomonas aeruginosa. PLoS ONE 2014, 9, e87276. [Google Scholar] [CrossRef] [PubMed]
- Lasocki, K.; Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. Deletion of the parA (soj) homologue in Pseudomonas aeruginosa causes ParB instability and affects growth rate, chromosome segregation, and motility. J. Bacteriol. 2007, 189, 5762–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosik, A.A.; Mierzejewska, J.; Thomas, C.M.; Jagura-Burdzy, G. ParB deficiency in Pseudomonas aeruginosa destabilizes the partner protein ParA and affects a variety of physiological parameters. Microbiololy 2009, 155, 1080–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawalek, A.; Bartosik, A.A.; Glabski, K.; Jagura-Burdzy, G. Pseudomonas aeruginosa partitioning protein ParB acts as a nucleoid-associated protein binding to multiple copies of a ParS-related motif. Nucleic Acids Res. 2018, 46, 4592–4606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.; Martin, R.G.; Rosner, J.L.; Davies, D.R. A novel DNA-binding motif in MarA: The first structure for an AraC family transcriptional activator. Proc. Natl. Acad. Sci. USA 1998, 95, 10413–10418. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein–ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef]
- Yang, J.; Roy, A.; Zhang, Y. BioLiP: A Semi-manually curated database for biologically relevant ligand–protein interactions. Nucleic. Acids Res. 2013, 41, D1096–D1103. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Tao, H.; He, J.; Huang, S.-Y. The HDOCK server for integrated protein–protein docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Winsor, G.L.; Lo, R.; Sui, S.J.H.; Ung, K.S.E.; Huang, S.; Cheng, D.; Ching, W.-K.H.; Hancock, R.E.W.; Brinkman, F.S.L. Pseudomonas aeruginosa genome database and PseudoCAP: Facilitating community-based, continually updated, genome annotation. Nucleic Acids Res. 2005, 33, D338–D343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawalek, A.; Glabski, K.; Bartosik, A.A.; Fogtman, A.; Jagura-Burdzy, G. Increased ParB level affects expression of stress response, adaptation and virulence operons and potentiates repression of promoters adjacent to the high affinity binding sites parS3 and parS4 in Pseudomonas aeruginosa. PLoS ONE 2017, 12, e0181726. [Google Scholar] [CrossRef] [Green Version]
- Cuypers, H.; Viebrock-Sambale, A.; Zumft, W.G. NosR, a membrane-bound regulatory component necessary for expression of nitrous oxide reductase in denitrifying Pseudomonas stutzeri. J. Bacteriol. 1992, 174, 5332–5339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machanick, P.; Bailey, T.L. MEME-ChIP: Motif analysis of large DNA datasets. Bioinformatics 2011, 27, 1696–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Martin, R.G.; Jair, K.W.; Wolf, R.E.; Rosner, J.L. Autoactivation of the marRAB multiple antibiotic resistance operon by the MarA transcriptional activator in Escherichia coli. J. Bacteriol. 1996, 178, 2216–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marx, C.J.; Lidstrom, M.E. Development of improved versatile broad-host-range vectors for use in methylotrophs and other gram-negative bacteria. Microbiology 2001, 147, 2065–2075. [Google Scholar] [CrossRef] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S.L. Enhanced annotations and features for comparing thousands of Pseudomonas genomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Feng, B.; Wen, G.; Xue, W.; Ma, G.; Zhang, H.; Wu, S. Bacterial lipoprotein biosynthetic pathway as a potential target for structure-based design of antibacterial agents. Curr. Med. Chem. 2020, 27, 1132–1150. [Google Scholar] [CrossRef] [PubMed]
- Corona, F.; Martínez, J.L.; Nikel, P.I. The global regulator Crc orchestrates the metabolic robustness underlying oxidative stress resistance in Pseudomonas aeruginosa. Environ. Microbiol. 2019, 21, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Bartell, J.A.; Blazier, A.S.; Yen, P.; Thøgersen, J.C.; Jelsbak, L.; Goldberg, J.B.; Papin, J.A. Reconstruction of the metabolic network of Pseudomonas aeruginosa to interrogate virulence factor synthesis. Nat. Commun. 2017, 8, 14631. [Google Scholar] [CrossRef] [Green Version]
- Shea, A.; Wolcott, M.; Daefler, S.; Rozak, D.A. Biolog phenotype microarrays. In Microbial Systems Biology: Methods and Protocols; Navid, A., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2012; pp. 331–373. ISBN 978-1-61779-827-6. [Google Scholar]
- Remus-Emsermann, M.N.P.; Schmid, M.; Gekenidis, M.-T.; Pelludat, C.; Frey, J.E.; Ahrens, C.H.; Drissner, D. Complete genome sequence of Pseudomonas citronellolis P3B5, a candidate for microbial phyllo-remediation of hydrocarbon-contaminated sites. Stand. Genom. Sci. 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Egan, S.M. Growing repertoire of AraC/XylS activators. J. Bacteriol. 2002, 184, 5529–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basturea, G.N.; Bodero, M.D.; Moreno, M.E.; Munson, G.P. Residues near the amino terminus of Rns are essential for positive autoregulation and dna binding. J. Bacteriol. 2008, 190, 2279–2285. [Google Scholar] [CrossRef] [Green Version]
- Marsden, A.E.; Schubot, F.D.; Yahr, T.L. Self-association is required for occupation of adjacent binding sites in Pseudomonas aeruginosa type III secretion system promoters. J. Bacteriol. 2014, 196, 3546–3555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra, M.C.; Collins, C.M. Mutational analysis of the N-terminal domain of UreR, the positive transcriptional regulator of urease gene expression. Microbiol. Res. 2012, 167, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Soisson, S.M.; MacDougall-Shackleton, B.; Schleif, R.; Wolberger, C. Structural basis for ligand-regulated oligomerization of AraC. Science 1997, 276, 421–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, E.P.; Lee, N. Three binding sites for AraC protein are required for autoregulation of AraC in Escherichia coli. Proc. Natl. Acad. Sci. USA 1988, 85, 1749–1753. [Google Scholar] [CrossRef] [Green Version]
- Schäper, S.; Steinchen, W.; Krol, E.; Altegoer, F.; Skotnicka, D.; Søgaard-Andersen, L.; Bange, G.; Becker, A. AraC-like transcriptional activator CuxR binds c-di-GMP by a PilZ-like mechanism to regulate extracellular polysaccharide production. Proc. Natl. Acad. Sci. USA 2017, 114, E4822–E4831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasil, M.L. Pseudomonas aeruginosa phospholipases and phospholipids. In Pseudomonas: Volume 4 Molecular Biology of Emerging Issues; Ramos, J.-L., Levesque, R.C., Eds.; Springer US: Boston, MA, USA, 2006; pp. 69–97. ISBN 978-0-387-28881-9. [Google Scholar]
- Munson, G.P.; Scott, J.R. Rns, a virulence regulator within the AraC family, requires binding sites upstream and downstream of its own promoter to function as an activator. Mol. Microbiol. 2000, 36, 1391–1402. [Google Scholar] [CrossRef]
- Munson, G.P.; Holcomb, L.G.; Scott, J.R. Novel group of virulence activators within the AraC family that are not restricted to upstream binding sites. Infect. Immun. 2001, 69, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Schweizer, H.P.; Jump, R.; Po, C. Structure and gene-polypeptide relationships of the region encoding glycerol diffusion facilitator (GlpF) and glycerol kinase (Glpk) of Pseudomonas aeruginosa. Microbiology 1997, 143, 1287–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, S.T.; Eiglmeier, K.; Ahmed, S.; Honore, N.; Elmes, L.; Anderson, W.F.; Weiner, J.H. Nucleotide sequence and gene-polypeptide relationships of the glpABC operon encoding the anaerobic sn-glycerol-3-phosphate dehydrogenase of Escherichia coli K-12. J. Bacteriol. 1988, 170, 2448. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.C.C. Glycerol dissimilation and its regulation in bacteria. Annu. Rev. Microbiol. 1976, 30, 535–578. [Google Scholar] [CrossRef]
- Nichols, C.E.; Lamb, H.K.; Lockyer, M.; Charles, I.G.; Pyne, S.; Hawkins, A.R.; Stammers, D.K. Characterization of Salmonella typhimurium YegS, a putative lipid kinase homologous to eukaryotic sphingosine and diacylglycerol kinases. Proteins 2007, 68, 13–25. [Google Scholar] [CrossRef]
- Schleif, R. AraC protein: A love–hate relationship. BioEssays 2003, 25, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, H.P.; Po, C. Cloning and nucleotide sequence of the glpD gene encoding sn-glycerol-3-phosphate dehydrogenase of Pseudomonas aeruginosa. J. Bacteriol. 1994, 176, 2184–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Titball, R.W. Bacterial phospholipases C. Microbiol. Rev. 1993, 57, 347–366. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.N.R.L.; Farias, C.B.B.; Rufino, R.D.; Luna, J.M.; Sarubbo, L.A. Glycerol as substrate for the production of biosurfactant by Pseudomonas aeruginosa UCP0992. Colloids Surf. B Biointerfaces 2010, 79, 174–183. [Google Scholar] [CrossRef]
- Scoffield, J.; Silo-Suh, L. Glycerol metabolism promotes biofilm formation by Pseudomonas aeruginosa. Can. J. Microbiol. 2016, 62, 704–710. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.P.; Mack, M.; Contiero, J. Glycerol: A promising and abundant carbon source for industrial microbiology. Biotechnol. Adv. 2009, 27, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Cronan, J.E.; Vagelos, P.R. Metabolism and function of the membrane phospholipids of Escherichia coli. Biochim. Biophys. Acta (BBA) Rev. Biomembr. 1972, 265, 25–60. [Google Scholar] [CrossRef]
- Xue, L.-L.; Chen, H.-H.; Jiang, J.-G. Implications of glycerol metabolism for lipid production. Prog. Lipid Res. 2017, 68, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Kahn, M.; Kolter, R.; Thomas, C.; Figurski, D.; Meyer, R.; Remaut, E.; Helinski, D.R. Plasmid cloning vehicles derived from plasmids ColE1, F, R6K, and RK2. Meth. Enzymol. 1979, 68, 268–280. [Google Scholar]
- Sambrook, J.; Maniatis, T.; Fritsch, E.F.; Laboratory, C.S.H. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1987; ISBN 978-0-87969-309-1. [Google Scholar]
- Rashid, M.H.; Kornberg, A. Inorganic polyphosphate is needed for swimming, swarming, and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 4885–4890. [Google Scholar] [CrossRef] [Green Version]
- Irani, V.R.; Rowe, J.J. Enhancement of transformation in Pseudomonas aeruginosa PAO1 by Mg2+ and heat. BioTechniques 1997, 22, 54–56. [Google Scholar] [CrossRef]
- El-Sayed, A.K.; Hothersall, J.; Thomas, C.M. Quorum-sensing-dependent regulation of biosynthesis of the polyketide antibiotic mupirocin in Pseudomonas fluorescens NCIMB 10586. Microbiology 2001, 147, 2127–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovach, M.E.; Phillips, R.W.; Elzer, P.H.; Roop, R.M.; Peterson, K.M. pBBR1MCS: A broad-host-range cloning vector. BioTechniques 1994, 16, 800–802. [Google Scholar] [PubMed]
- Ludwiczak, M.; Dolowy, P.; Markowska, A.; Szarlak, J.; Kulinska, A.; Jagura-Burdzy, G. Global transcriptional regulator KorC coordinates expression of three backbone modules of the broad-host-range RA3 plasmid from IncU incompatibility group. Plasmid 2013, 70, 131–145. [Google Scholar] [CrossRef] [Green Version]
- Karimova, G.; Pidoux, J.; Ullmann, A.; Ladant, D. A Bacterial two-hybrid system based on a reconstituted signal transduction pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 5752–5756. [Google Scholar] [CrossRef] [Green Version]
- Kotecka, K.; Kawalek, A.; Kobylecki, K.; Bartosik, A.A. The MarR-type regulator PA3458 is involved in osmoadaptation control in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 3982. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S.; Häussler, S. Chromatin immunoprecipitation for ChIP-chip and ChIP-seq. In Pseudomonas Methods and Protocols; Filloux, A., Ramos, J.-L., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2014; pp. 591–605. ISBN 978-1-4939-0473-0. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. 1000 Genome project data processing subgroup the sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, F.; Ryan, D.P.; Grüning, B.; Bhardwaj, V.; Kilpert, F.; Richter, A.S.; Heyne, S.; Dündar, F.; Manke, T. DeepTools2: A next generation web server for deep-sequencing data analysis. Nucleic Acids Res. 2016, 44, W160–W165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, T.; Meyer, C.A.; Eeckhoute, J.; Johnson, D.S.; Bernstein, B.E.; Nusbaum, C.; Myers, R.M.; Brown, M.; Li, W.; et al. Model-based analysis of ChIP-Seq (MACS). Genome Biol. 2008, 9, R137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.J.; Gazin, C.; Lawson, N.D.; Pagès, H.; Lin, S.M.; Lapointe, D.S.; Green, M.R. ChIPpeakAnno: A bioconductor package to annotate ChIP-Seq and ChIP-Chip data. BMC Bioinform. 2010, 11, 237. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative Genomics Viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [Green Version]
- Thibodeau, S.A.; Fang, R.; Joung, J.K. High-throughput β-galactosidase assay for bacterial cell-based reporter systems. BioTechniques 2004, 36, 410–415. [Google Scholar] [CrossRef]
- Medema, M.H.; Takano, E.; Breitling, R. Detecting sequence homology at the gene cluster level with MultiGeneBlast. Mol. Biol. Evol. 2013, 30, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference Sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [Green Version]
- Goudenège, D.; Avner, S.; Lucchetti-Miganeh, C.; Barloy-Hubler, F. CoBaltDB: Complete bacterial and archaeal orfeomes subcellular localization database and associated resources. BMC Microbiol. 2010, 10, 88. [Google Scholar] [CrossRef] [Green Version]
- Simon, R.; O’Connell, M.; Labes, M.; Pühler, A. Plasmid vectors for the genetic analysis and manipulation of Rhizobia and other gram-negative bacteria. Meth. Enzymol. 1986, 118, 640–659. [Google Scholar]
- Kawalek, A.; Kotecka, K.; Modrzejewska, M.; Gawor, J.; Jagura-Burdzy, G.; Bartosik, A.A. Genome sequence of Pseudomonas aeruginosa PAO1161, a PAO1 derivative with the ICEPae1161 integrative and conjugative element. BMC Genom. 2020, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Mierzejewska, J.; Bartosik, A.A.; Macioszek, M.; Płochocka, D.; Thomas, C.M.; Jagura-Burdzy, G. Identification of C-Terminal hydrophobic residues important for dimerization and all known functions of ParB of Pseudomonas aeruginosa. Microbiology 2012, 158, 1183–1195. [Google Scholar] [CrossRef]





| PAO1 ID | PAO1161 ID (D3C65_) | Fold Change in RNA-seq | PseudoCAP Category | Gene Product |
|---|---|---|---|---|
| PA3024 | 10155 | 173.40 | EM; PE | carbohydrate kinase |
| PA3025 | 10150 | 163.31 | CCC; EM | glycerol-3-phosphate dehydrogenase/oxidase |
| PA3026 | 10145 | 135.03 | HUU | FAD-binding oxidoreductase |
| PA3464 | 07800 | 129.57 | HUU | phospholipase |
| PA3027 | 10140 | 107.00 | TR | AraC family transcriptional regulator |
| PA3023 | 10160 | 47.32 | HUU | lipid kinase YegS |
| PA2202 | 14525 | 15.56 | MP; TSM | amino acid ABC transporter permease |
| PA3342 | 08435 | 15.38 | MP | DUF2804 domain-containing protein |
| PA2203 | 14520 | 14.14 | MP; TSM | amino acid ABC transporter permease |
| PA5024 | 27340 | 13.07 | HUU | sulfite exporter TauE/SafE family protein |
| PA4191 | 04005 | 13.06 | PE | isopenicillin N synthase family oxygenase |
| PA4193 | 03995 | 12.57 | MP; TSM | amino acid ABC transporter permease |
| PA4192 | 04000 | 10.44 | TSM | amino acid ABC transporter ATP-binding protein |
| PA0282 | 01495 | 10.01 | MP; TSM | sulfate ABC transporter permease subunit CysT |
| PA3392 | 08180 | −11.34 | EM | TAT-dependent nitrous-oxide reductase |
| PA0519 | 02740 | −11.50 | EM | nitrite reductase |
| PA0524 | 02765 | −23.76 | EM | nitric-oxide reductase large subunit |
| PA3391 | 08185 | −44.43 | EM; MP | regulatory protein NosR |
| PA0523 | 02760 | −62.05 | EM | cytochrome c |
| Region ID | Binding Site Position | Gene in PAO1161 (D3C65_) | Gene in PAO1 | Fold Change (RNA-seq) | Fold Enrichment (ChIP-seq) | Position of Summit | PseudoCAP Category | Gene Product |
|---|---|---|---|---|---|---|---|---|
| 1 | term | 01245 | PA0234 | −1.71 | 5.73 | 264,723 | HUU | nucleoside-binding protein |
| 2 | term | 04760 | PA4045 | 1.54 | 3.76 | 987,296 | HUU | cobalamin-binding protein |
| 3 | intra | 07240 | PA3572 | −2.18 | 7.06 | 1,510,957 | HUU | hypothetical protein |
| 4 | prom | 07800 | PA3464 | 129.57 | HUU | phospholipase | ||
| 07795 | PA3465 | −3.80 | MP | conserved hypothetical protein | ||||
| 5 | term | 07800 | PA3464 | 129.57 | 11.84 | 1,641,558 | HUU | phospholipase |
| 6 | prom | 08155 | PA3397 | 1.38 | 3.72 | 1,712,386 | BCPGC; EM | ferredoxin-NADP reductase |
| 08150 | PA3398 | 1.17 | TR | probable transcriptional regulator | ||||
| 7 | intra | 08185 | PA3391 | −44.43 | 4.30 | 1,720,088 | EM; MP | regulatory protein NosR |
| 8 | prom | 08435 | PA3342 | 15.38 | 5.68 | 1,762,086 | MP | DUF2804 domain-containing protein |
| 9 | intra | 08440 | PA3341 | −1.16 | 2.42 | 1,763,471 | TR | MarR family transcriptional regulator |
| 10 | intra | 08605 | PA3309 | −3.36 | 9.83 | 1,805,406 | HUU | universal stress protein |
| 11 | intra | 09845 | PA3083 | 1.51 | 8.26 | 2,055,622 | TPTMD | aminopeptidase |
| 12 | intra | 11,685 | PA2742 | −1.20 | 3.10 | 2,411,617 | TPTMD | 50S ribosomal protein L35 |
| 13 | prom | 12,415 | PA2601 | 1.04 | 3.90 | 2,570,649 | TR | LysR family transcriptional regulator |
| 12,420 | PA2602 | 1.75 | HUU | 3-mercaptopropionate dioxygenase | ||||
| 14 | term | 12,415 | PA2601 | 1.04 | TR | LysR family transcriptional regulator | ||
| 15 | intra | 14,830 | PA2147 | −1.73 | 5.28 | 3,153,105 | AP | catalase HPII |
| 16 | intra | 18,720 | PA1414 | −2.19 | 9.03 | 3,976,577 | HUU | hypothetical protein |
| 17 | prom | 19,855 | PA1196 | −4.91 | 3.16 | 4,218,075 | TR | sigma-54-dependent Fis family transcriptional regulator |
| 19,860 | PA1195 | 1.35 | HUU | dimethylarginine dimethylaminohydrolase DdaH | ||||
| 18 | intra | 21,295 | PA0928 | −1.04 | 4.07 | 4,499,850 | TCRS | hybrid sensor histidine kinase/response regulator |
| 19 | intra | 25,030 | PA4610 | −3.03 | 2.56 | 5,276,490 | HUU | copper transporter |
| 20 | term | 26,025 | PA4772 | 1.83 | 3.09 | 5,480,812 | EM | FAD-binding oxidoreductase |
| 21 | intra | 28,310 | PA5208 | −3.54 | 3.28 | 5,982,827 | HUU | TIGR00153 family protein |
| 22 | intra | 28,490 | PA5243 | −1.56 | 3.49 | 6,024,153 | BCPGC | porphobilinogen synthase |
| 23 | prom | 28,760 | PA5294 | −1.60 | 3.02 | 6,078,786 | HUU | multidrug resistance protein NorM |
| 28,755 | PA5293 | −1.17 | TR | probable transcriptional regulator | ||||
| 24 | prom | 10,145 | PA3026 | 135.03 | 1.89 | 2,124,729 | HUU | FAD-binding oxidoreductase |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotecka, K.; Kawalek, A.; Kobylecki, K.; Bartosik, A.A. The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa. Int. J. Mol. Sci. 2021, 22, 5066. https://doi.org/10.3390/ijms22105066
Kotecka K, Kawalek A, Kobylecki K, Bartosik AA. The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa. International Journal of Molecular Sciences. 2021; 22(10):5066. https://doi.org/10.3390/ijms22105066
Chicago/Turabian StyleKotecka, Karolina, Adam Kawalek, Kamil Kobylecki, and Aneta Agnieszka Bartosik. 2021. "The AraC-Type Transcriptional Regulator GliR (PA3027) Activates Genes of Glycerolipid Metabolism in Pseudomonas aeruginosa" International Journal of Molecular Sciences 22, no. 10: 5066. https://doi.org/10.3390/ijms22105066