1. Introduction
Oral squamous cell carcinoma (OSCC) is a major global health problem with high incidence and low survival rates. Chronic bacterial infections have been shown to be associated with chronic inflammation, anti-apoptotic activity and pro-proliferative activities, and the Gram-negative, anaerobic bacteria,
Porphyromonas gingivalis and
Fusobacterium nucleatum contribute to oral carcinogenesis and progression of oral cancer [
1].
Early oral colonisers include Gram-positive bacterial species from the Streptococcus and Actinomyces families which produce lipoteichoic acid (LTA). Late oral colonisers include Gram-negative bacterial species
F. nucleatum,
P. gingivalis and
Prevotella spp., which produce lipopolysaccharide (LPS). Both Gram-positive and Gram-negative bacteria form oral biofilms. Bacterial infection has been shown to be associated with chronic inflammation, anti-apoptotic activity, pro-proliferative activities, which play a role in cancer development [
2].
LPS is found in the cell wall of Gram-negative bacteria. Previous studies have shown that
P. gingivalis LPS tend to increase the viability of oral cancer cells [
3]. Studies of the tumour microenvironment have shown that macrophages demonstrate increased IL-6 and CD14 expression and increased nitric oxide secretion after the stimulation of LPS [
2]. In macrophages, LPS upregulates tumour necrosis factor alpha (TNFα), that induces suppression of cytokine signalling 3 (
SOCS3) mRNA expression and inhibits IL-6-induced activation of signal transducer and activator of transcription 3 (
STAT3) [
4]. In contrast, the LPS–squamous cell carcinoma–monocyte interaction leads to rapid STAT3 protein activation and cancer progression [
5].
LTA from
Staphylococcus aureus has been shown to stimulate proliferation of human non-small cell lung cancer cells [
6]. LTA from
Streptococcus mutans,
Streptococcus pyogenes,
Streptococcus faecalis,
Streptococcus sanguis and
S. aureus upregulate Human hepatocyte growth factor (HuHGF) [
7], which has been shown to be associated with the progression of oral cancer by stimulating stromal fibroblast-induced invasion (epithelial mesenchymal cells) [
8].
Recently, we demonstrated that combined stimulation of oral cancer cells Cal 27 by LPS and LTA resulted in greater proliferation of cells than stimulation with either LPS or LTA alone [
9].
Capsaicin induces apoptosis in gastric cancer cells, various types of breast cancer as well as in pancreatic, oesophageal, colonic, prostrate, skin, endothelial cell and lung cancer [
10]. Capsaicin is reported to reduce the proliferation of an oral squamous cancer cell line of Asian origin, ORL-48 [
11], and induces apoptosis in SCC4 and SCC25 oral cancer cells [
12].
In the clinical settings, the oral cavity contains oral biofilms as dental plaques that harbour both Gram-negative and Gram-positive bacteria. These bacteria release antigens, which may present prior to and/or during oral cancer chemotherapies given the presence of dental plaque in cancer patients. The aim of this study is to investigate whether the bacterial antigens in the oral cavity affect the efficacy of chemotherapy treatment of oral cancer cells by measuring cell metabolism, viability, and apoptotic cell death. We also investigated the effect of capsaicin on the expression of oral cancer cell proliferation factors SOCS3, EGFR (epidermal growth factor receptor), STAT3 and PI3K (phosphoinositide 3-kinase) in the presence or absence of bacterial antigens.
The various capsaicin and LPS/LPS + LTA combination tests represent various clinical scenarios, such as the presence or absence of dental plaque containing Gram-negative and Gram-positive bacterial biofilms providing antigenic stimulation, which is demonstrated in
Figure 1.
3. Discussion
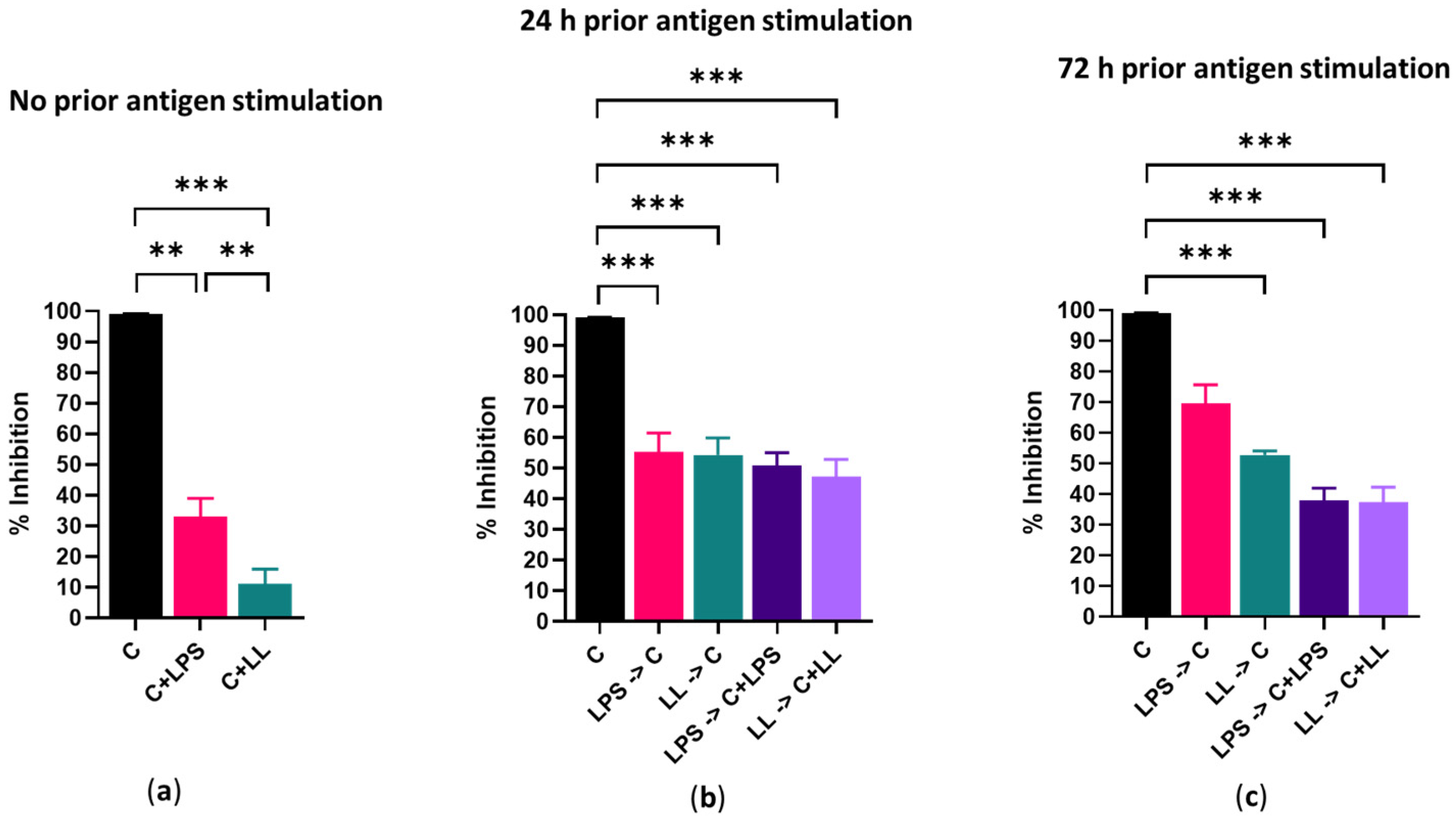
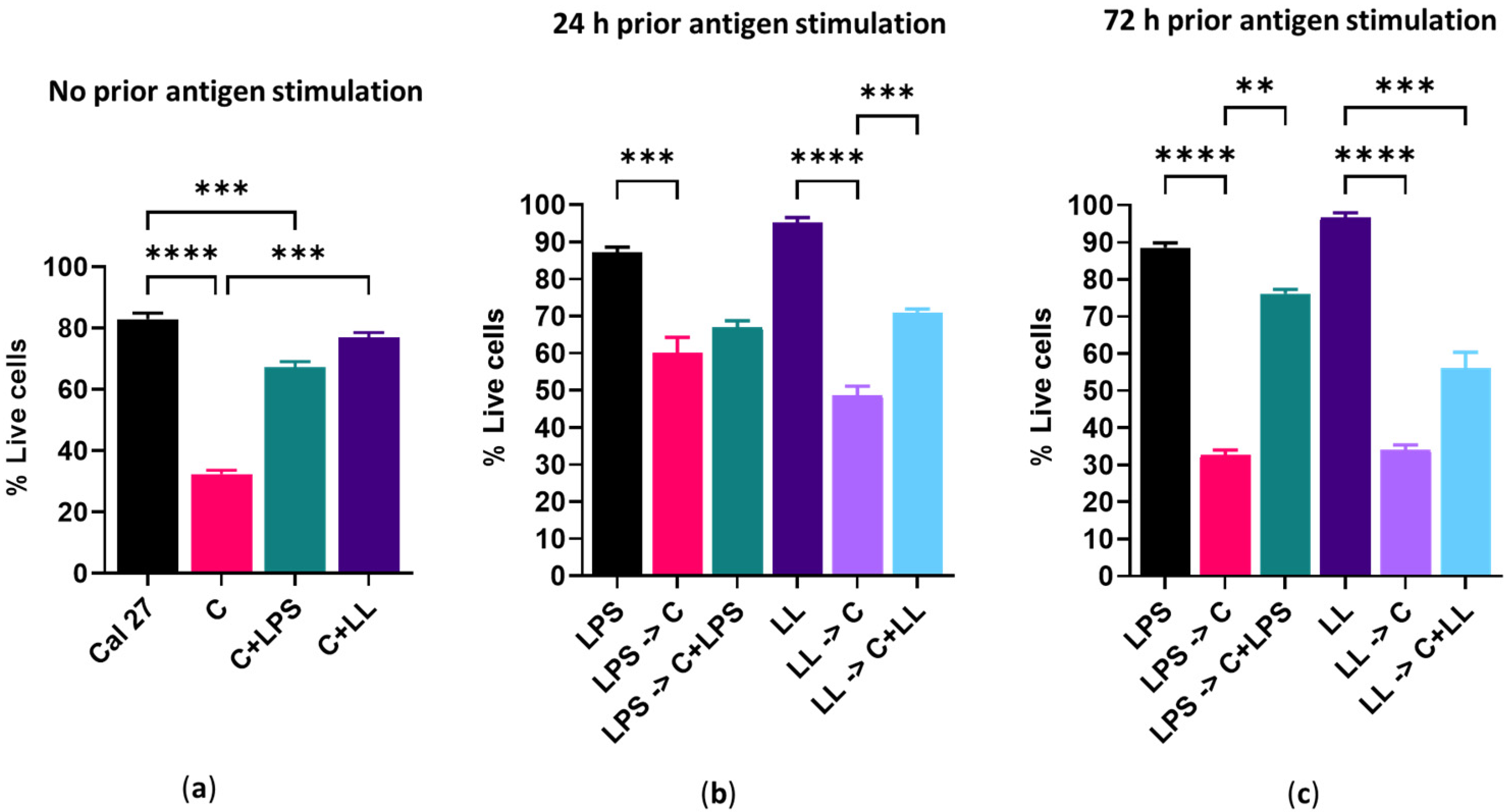
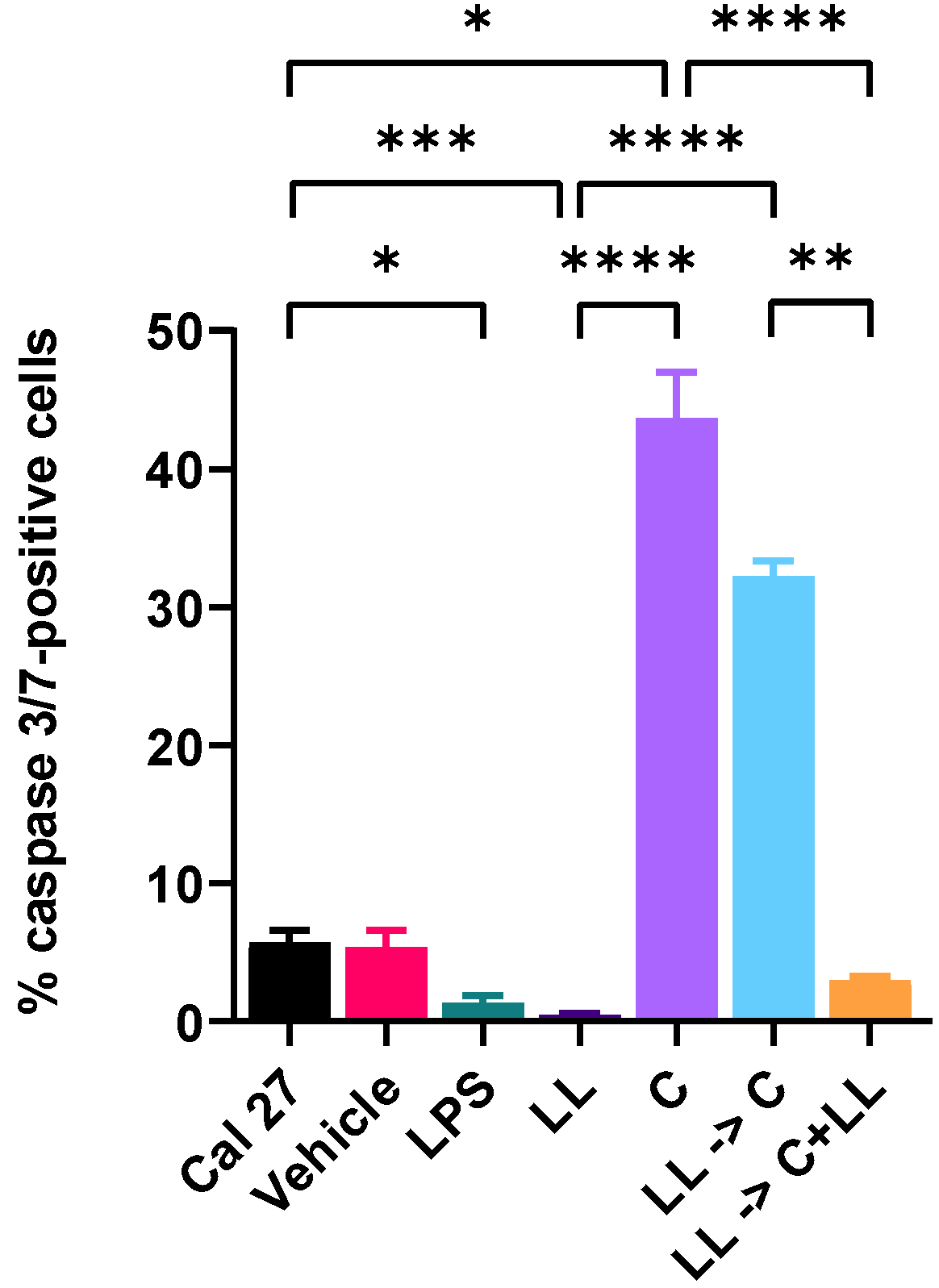
In the absence of antigenic stimulation, 150 µM capsaicin reduced the metabolism of Cal 27 cells by 100% and killed 68% of the cells. However, the addition of bacterial antigens prior to drug treatment reduced the efficacy of capsaicin treatment. Simultaneous addition of bacterial antigens with drug treatment with and without prior bacterial antigen stimulation also significantly reduced capsaicin efficacy, resulting in decreased cell killing and decreased inhibition of metabolism. In general, the addition of both LPS and LTA resulted in greater inhibition of drug efficacy than the addition of LPS only. We believe the mechanism for this decreased efficacy is that bacterial antigens increase cancer cell proliferation. Other researchers using a co-culture system found that cancer drug efficacy was significantly changed due to the presence of bacteria [
13].
The most challenging aspect of the project was to design a drug combination treatment that could mimic the real-world challenge of anti-cancer drugs in the presence of bacterial antigens that themselves mimic the presence of Gram-negative and Gram-positive bacteria in dental plaques containing oral biofilms. Oral biofilm formation is a continuous process. Even after proper extirpation/elimination of a bacterial plaque via surgical manoeuvre or antibiotic treatment, oral cancer patients frequently experience unhygienic oral conditions, leading to continuous formation of bacterial biofilm. The elimination of a bacterial plaque during treatment does not necessarily remove the existing bacterial antigen stimulatory effect on oral cancer cells. Therefore, combination drug treatment may be an appropriate approach to elucidate the confounding effects of bacterial antigens on chemotherapeutic drugs.
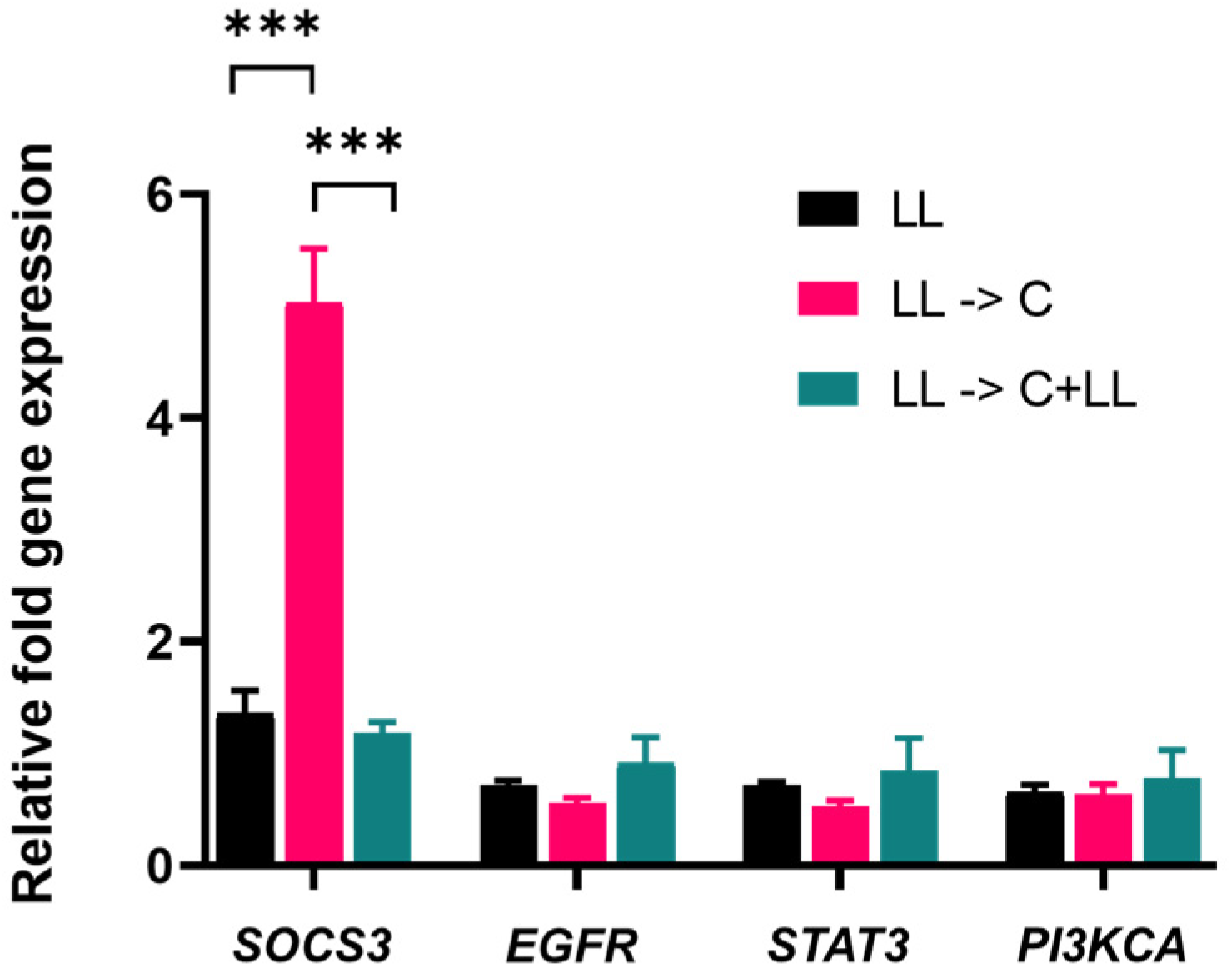
Capsaicin treatment significantly increased gene expression of
SOCS3 of oral cancer cells. However, the increased
SOCS3 gene expression was reversed in the presence of bacterial antigens during treatment. SOCS3, considered to be a tumour suppressor protein, primarily acts on the JAK/STAT pathway [
14] and regulates proliferation and activation of T-helper cells [
15]. Loss of
SOCS3 gene expression turns STAT3 from anti-apoptotic to pro-apoptotic [
16]. Hyperactivation of EGFR mediates apoptosis via STAT3 [
17], and EGFR hyperactivation may result in lowering of cell viability, independent of the PI3K/Akt pathway [
18]. However, there was no significant difference between
EGFR and
STAT3 gene and protein expression in capsaicin-treated pre-stimulated cells. Despite a significant increase in
SOCS3 mRNA expression, a corresponding increase in protein expression was not detected. The gene expression found in mRNA levels may not necessarily correspond to protein levels.
Recent evidence supports the role of microbiota in cancer development and carcinogenesis, especially oral cancer [
2,
19] and colon cancer [
20]. To the best of our knowledge, this is the first study to investigate the effect of bacterial antigens on oral cancer treatment, mimicking the real-world scenario.
However, there are some limitations in this study. The limitations and possible future studies are discussed below.
First, this project was not carried out in a co-culture model setup (i.e., oral bacteria and oral cancer cells were cultured together) that incorporated immune cells. Bacterial antigens have been found to mostly target immune cells for cancer progression. The project found that bacterial antigens can stimulate oral cancer cells without the presence of immune cells. However, in the oral cavity, there are confounding factors such as immune cells and cytokines, and future studies using an immune cell–cancer cell co-culture model, or a tissue study on T-cell phenotypic changes, should be undertaken to elucidate the role of bacterial antigens and capsaicin more thoroughly.
Second, the project only used one oral cancer cell line (Cal 27), which is an aggressive cancer cell line and has been shown to respond to bacterial antigen stimulation more than SSC4, SSC9 and SSC25 cell lines [
9]. The effect of bacteria antigens on cancer drug efficacy needs to be determined in additional cancer cell lines. Cancer tissues are heterogeneous in nature, and the effect of bacterial antigens and capsaicin based on one oral cancer cell line may not be generalised to other cancer cell lines.
Third, this project used LTA and LPS to simulate Gram-positive and Gram-negative bacterial infection, respectively. However, components of bacteria other than their bacterial cell wall antigens may also be responsible for the progression of oral squamous cell carcinomas. The project could not establish a temporal relationship in the effect of different bacterial antigens on the proliferation of the oral cancer cells. During the formation of a biofilm, Gram-positive bacteria are the initial colonisers. Gram-positive bacterial antigen may result in the upregulation or downregulation of various proliferating, apoptotic or anti apoptotic factors before Gram-negative bacterial antigens have any effect on oral cancer cells.
Fourth, the project has only investigated the effect of bacterial antigens on one anticancer agent—capsaicin. Cancer development and progression is a complex process, involves many pathways and there are many other cancer target and anticancer agents. Future studies should explore the effect of bacterial antigens on other anticancer agents.
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
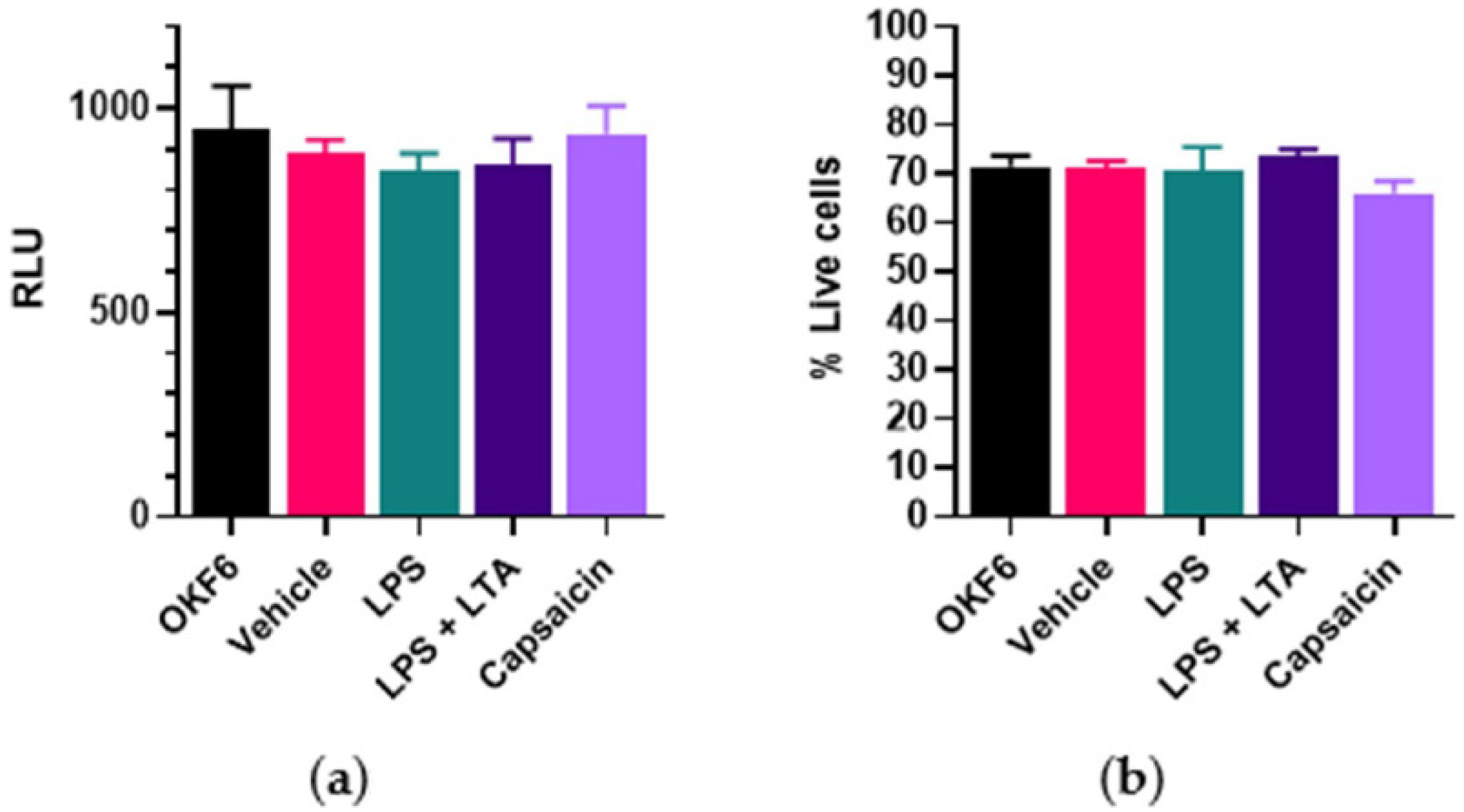
Oral cancer cells Cal 27 (American Type Culture Collection CRL-2095, Manassas, VA, USA) were used for this study, as we previously showed that LPS and LTA stimulation resulted in greater proliferation in Cal 27 cells than in other oral cancer cell lines (SSC4, SSC9 and SS25) or normal oral cell lines (
p ≤ 0.001) tested [
9]. Cal 27 cells were cultured using Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% foetal bovine serum and 1% penicillin/streptomycin, while OKF6 (normal oral cells) were cultured in keratinocyte serum-free medium (KSFM) plus growth factors, as previously described [
9]. Cells were seeded at 5000 cells per well in a 96-well plate as previously described [
9]. Each 75 cm
2 flask of cells acted as a biological replicate, and each flask was obtained from three different cell passages. Experiments were repeated at different times. Throughout this report,
n refers to the number of biological replicates.
4.2. Determine the Capsaicin Treatment Time and Concentration
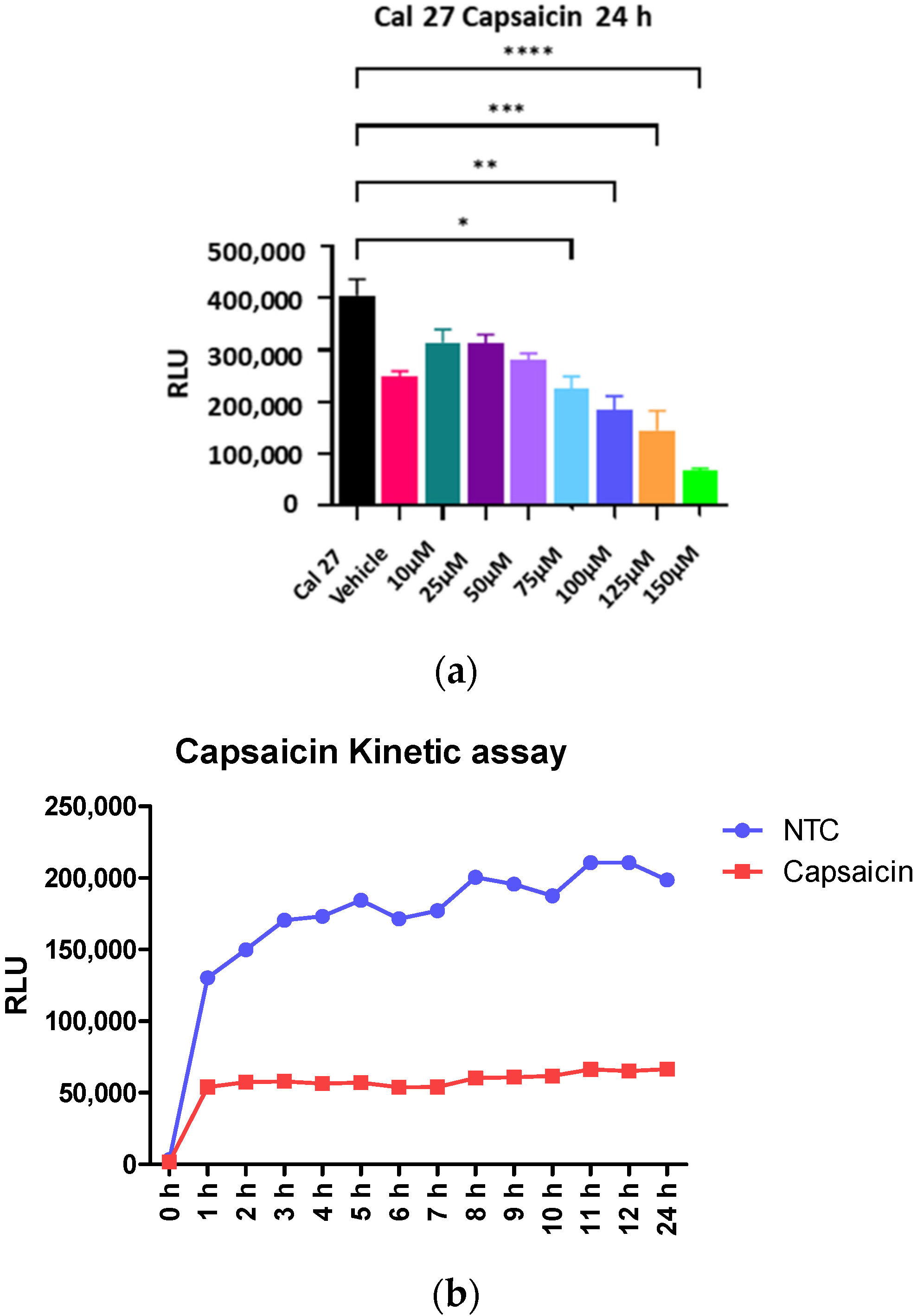
Cal 27 cells were incubated with capsaicin (Sigma-Aldrich, St. Louis, MO, USA) at varying concentrations from 0 to 150 µM for 24 h, and cellular metabolism was determined by the MT Glo assay. The level of inhibition remained steady for up to 24 h (
Figure 2b). In contrast, mitogens such as LPS or LTA require a longer time for activation of cells, and therefore, the capsaicin efficacy tests were conducted for 24 h.
4.3. Bacterial Antigens and Capsaicin Combination Tests
The clinical rationale of bacterial antigens’ stimulation and capsaicin treatment is explained in
Figure 1. Bacterial antigens, LPS (from
Escherichia coli O111:B4, Sigma Aldrich, St. Louis, MO, USA) and LTA (from Streptococcus pyogenes, Sigma Aldrich, St. Louis, MO, USA) were used at their previously found optimum concentration of 5 µg/mL [
9], and capsaicin at 150 µM.
Cal 27 cells were plated in a 96 well flat-bottomed plate at 5000 cells/well and stimulated with bacterial antigens LPS/LPS + LTA for 0, 24 and 72 h prior to treatment with capsaicin in the presence or absence of additional antigenic stimulation for 24 h. Then, cellular metabolism and viability were determined (
Figure 8). Cal 27 cells without capsaicin treatment were used as control cells for each condition.
4.4. Inhibition Effect of Capsaicin Treatment on Oral Cancer Cell Metabolism
Cell metabolism was determined using the real-time Glo MT cell viability assay (Promega) according to the manufacturer’s instructions.
The percentage of inhibition of metabolism of capsaicin treatment was calculated as:
4.5. Cell Viability Assay
Cell viability was determined by the Trypan blue exclusion assay. Cells were detached by the addition of 40 µL of 0.25% trypsin EDTA solution (Sigma, St. Louis, MO, USA), mixed in an equal volume of 0.4% Trypan blue (Invitrogen, Waltham, MA, USA), and live and dead cells were counted.
4.6. Apoptotic Assay
Caspase 3/7-positive cells are a marker of apoptosis. Cell Event Caspase-3/7 Green Detection Reagent (Invitrogen, Waltham, MA, USA) was used for live cell imaging and to estimate the total number of apoptotic cells, and 5000 cells (Cal 27) per 96-well plate (transparent, flat-bottomed) were plated and subjected to different treatments. After the incubation period for each treatment, the medium was carefully removed, and cells were washed once with 1× PBS, then 100 µL of 7.2 µM Cell Event Caspase-3/7 Green Detection Reagent in 1× PBS + 5% FBS was added to each well of the 96-well plate and incubated at 37 °C/5% CO
2 for 30 min. The cells were imaged using the FITC filter sets, as the maximum excitation/emission for the Cell Event Caspase-3/7 Green Detection Reagent is 502/530 nm. Caspase3/7 apoptotic analysis was carried out using ImageJ version 1.52a software (National Institutes of Health, Bethesda, MD, USA,
http://imagej.nih.gov/ij (accessed on 10 August 2021)).
4.7. Reverse Transcription Quantitative PCR for Proliferation Factors
Reverse transcription quantitative PCR was conducted for proliferation factors
SOCS3,
EGFR,
STAT3,
GAPDH and
PI3KCA. RNA extraction and RT-qPCR and analysis were performed as described previously [
9]. Briefly, total RNA was extracted with the Invitrogen Trizol Plus RNA purification kit. SSIV VILO Master Mix W/EzDNase (Invitrogen, Waltham, MA, USA) was used to reverse transcribe the RNA and qPCR conducted using Power-up SYBR Master Mix (Invitrogen, Waltham, MA, USA). Primer pair sequences are listed in
Supplementary Table S3. The house-keeping gene GAPDH (internal control) and the untreated Cal 27 cells were used to normalise the gene expression fold change in the treatment groups using the comparative Ct (ΔΔCt) method.
4.8. Western Blot
Ten μg of total protein from each bacterial antigen treatment condition was used for Western blotting. Primary antibodies were obtained from R&D Systems and used at the following concentrations: 0.1 μg/mL human/mouse SOCS3 antibody, 1 μg/mL human PI 3-kinase p110β antibody, 0.1 μg/mL STAT3 Mouse anti-human, mouse, rat, 1 μg/mL human EGFR antibody, 1:1000 human GAPDH. Blots were probed with the respective secondary antibodies from R&D Systems at the following concentrations: 1:1500 anti-mouse IgG HRP conjugate, 1:1500 anti-goat IgG HRP conjugate, 1:1500 anti-rabbit IgG HRP conjugate. High-resolution images were acquired using the Chemidoc MP Imaging system and processed using Image Lab version 6.0 (Bio-Rad Laboratories, Hercules, CA, USA). The band densities of the house-keeping gene GAPDH (internal control) and the untreated Cal 27 cells were used to normalise the protein expression in the treatments [
9].
4.9. Statistical Analysis
All statistical analyses were performed using GraphPad Prism version 9 (GraphPad Software, San Diego, CA, USA). The data were tested for normality of distribution using the D’Agostino and Pearson omnibus normality test. One-way or two-way analysis of variance (ANOVA) Kruskal–Wallis test multiple comparisons on mean ranks were performed in the oral cell proliferation and suppression, metabolism, viability and gene expression analyses. The probability threshold of p ≤ 0.05 was considered statistically significant.
5. Conclusions
This study demonstrated that bacterial antigens play a confounding role during the action of capsaicin on Cal 27 oral cancer cell proliferation. Capsaicin reduces bacterial antigen-induced oral cancer cell proliferation. The presence of bacterial antigens lowered the inhibition percentage of capsaicin, and combined LPS + LTA acted synergistically to lower the effect of capsaicin on Cal 27 oral cancer cells.
These findings may benefit other cancer researchers considering the presence of bacterial antigens when developing novel pharmacological strategies for cancer, not only oral cancer, head and neck cancer, but also colorectal cancer, where commensal bacteria may affect chemotherapeutic drugs. The importance of eliminating the bacterial antigens is also highlighted, such as thoroughly cleaning the dental plaque and maintaining oral hygiene prior to and during oral cancer chemotherapy.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







