
Hypoxic Culture Maintains Cell Growth of the Primary Human Valve Interstitial Cells with Stemness

 ,
,
Abstract
:1. Introduction
2. Results
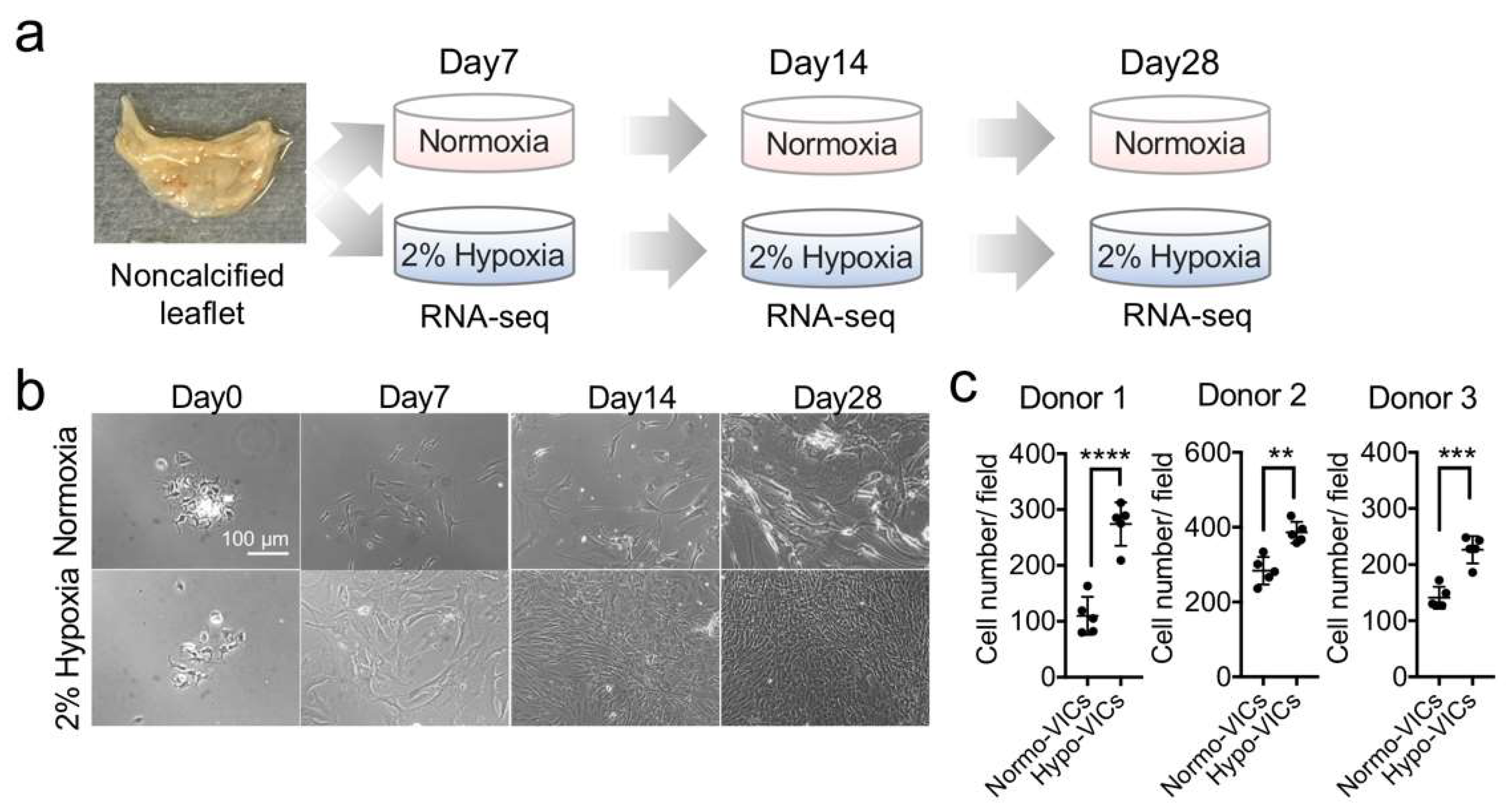
2.1. Low Oxygen Tension Promotes VIC Proliferation
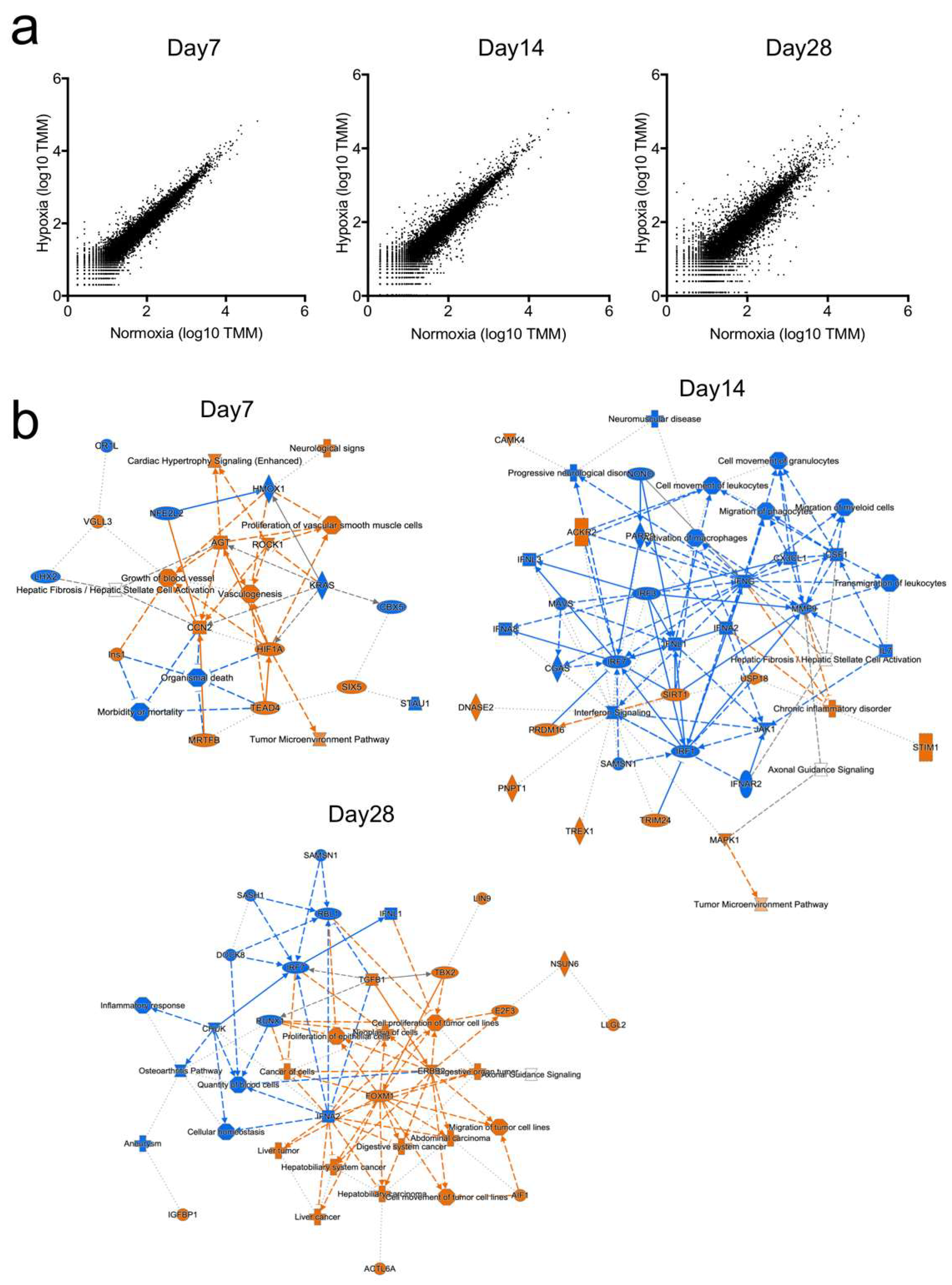
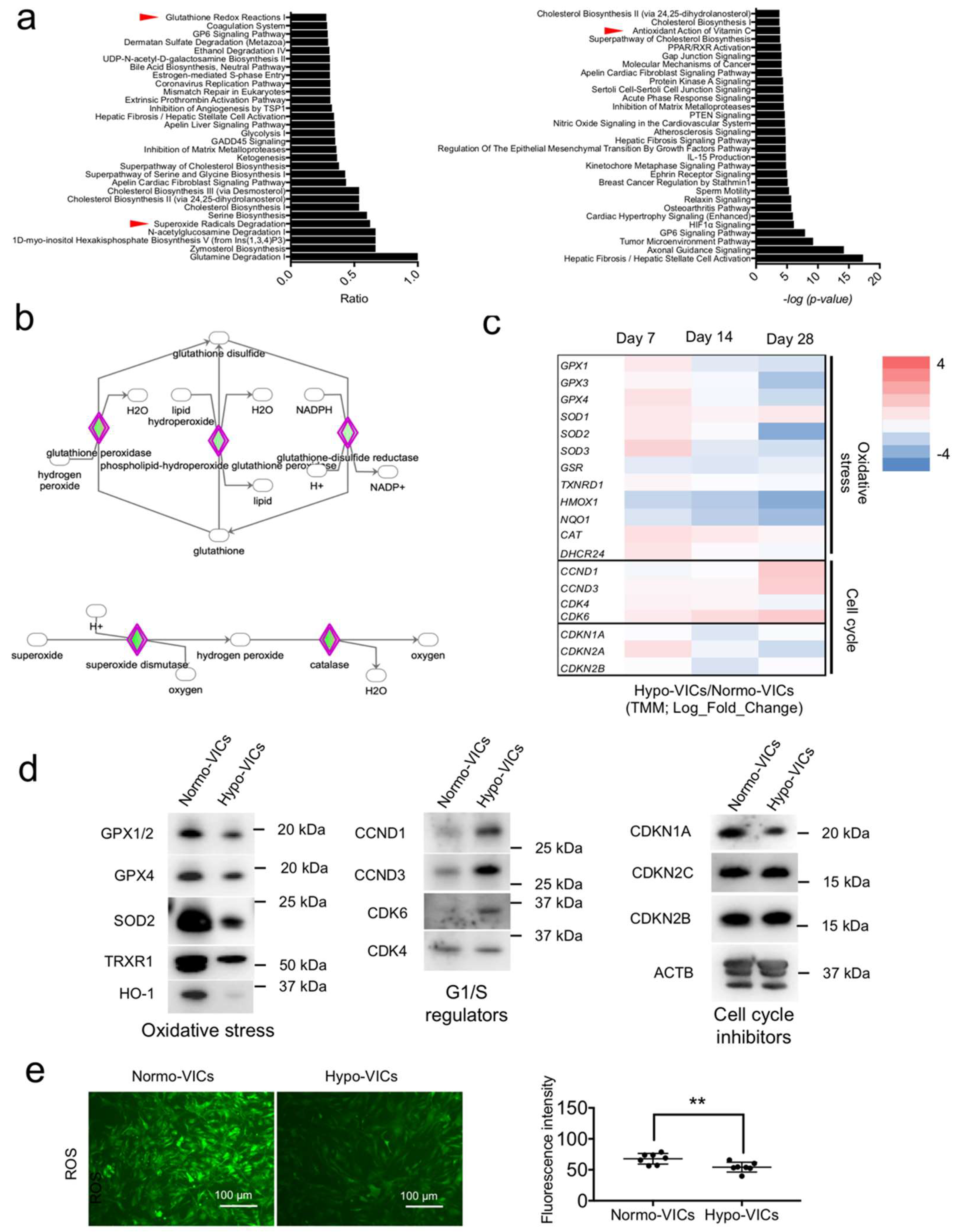
2.2. Gene Expression Profiling
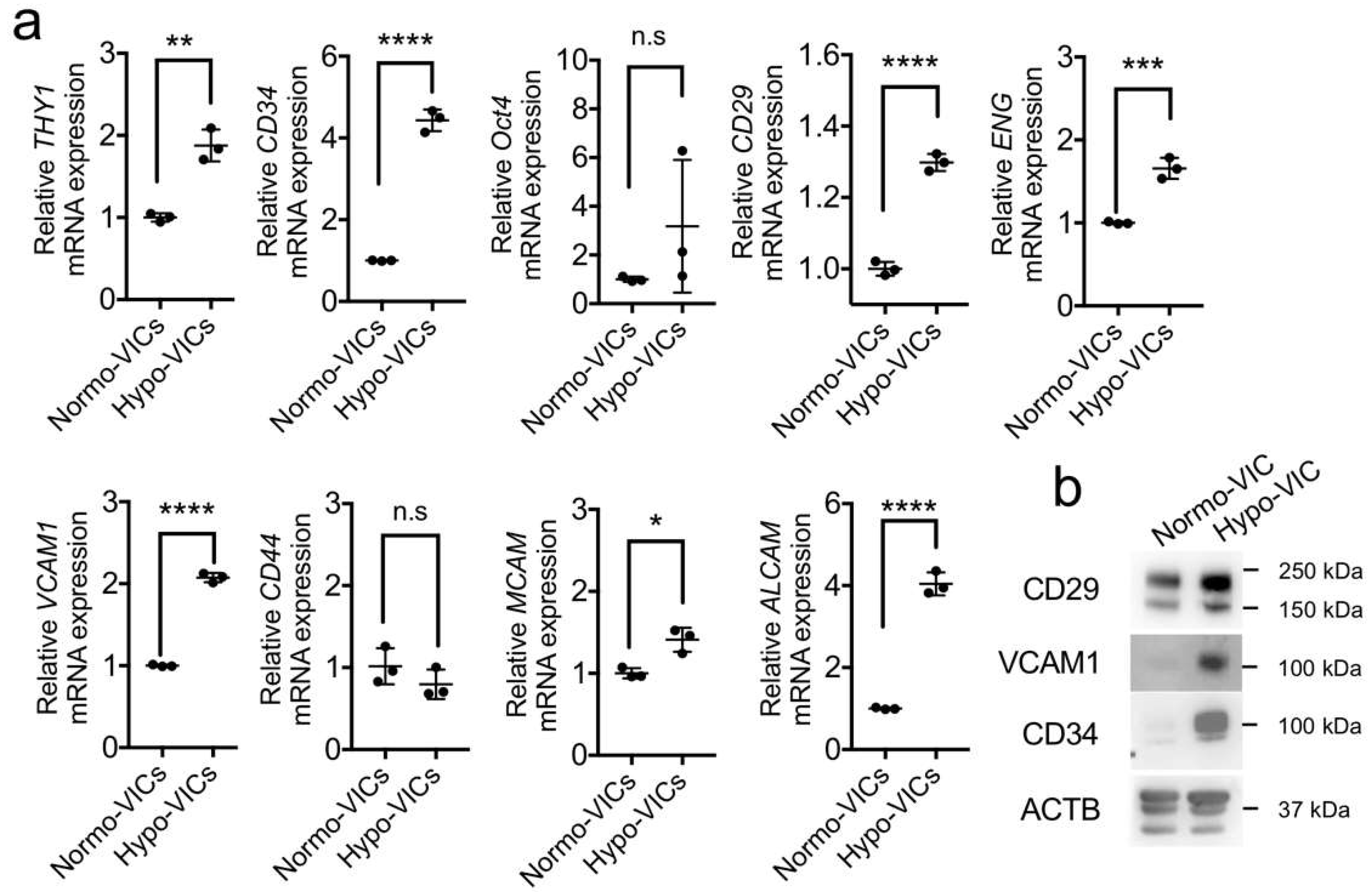
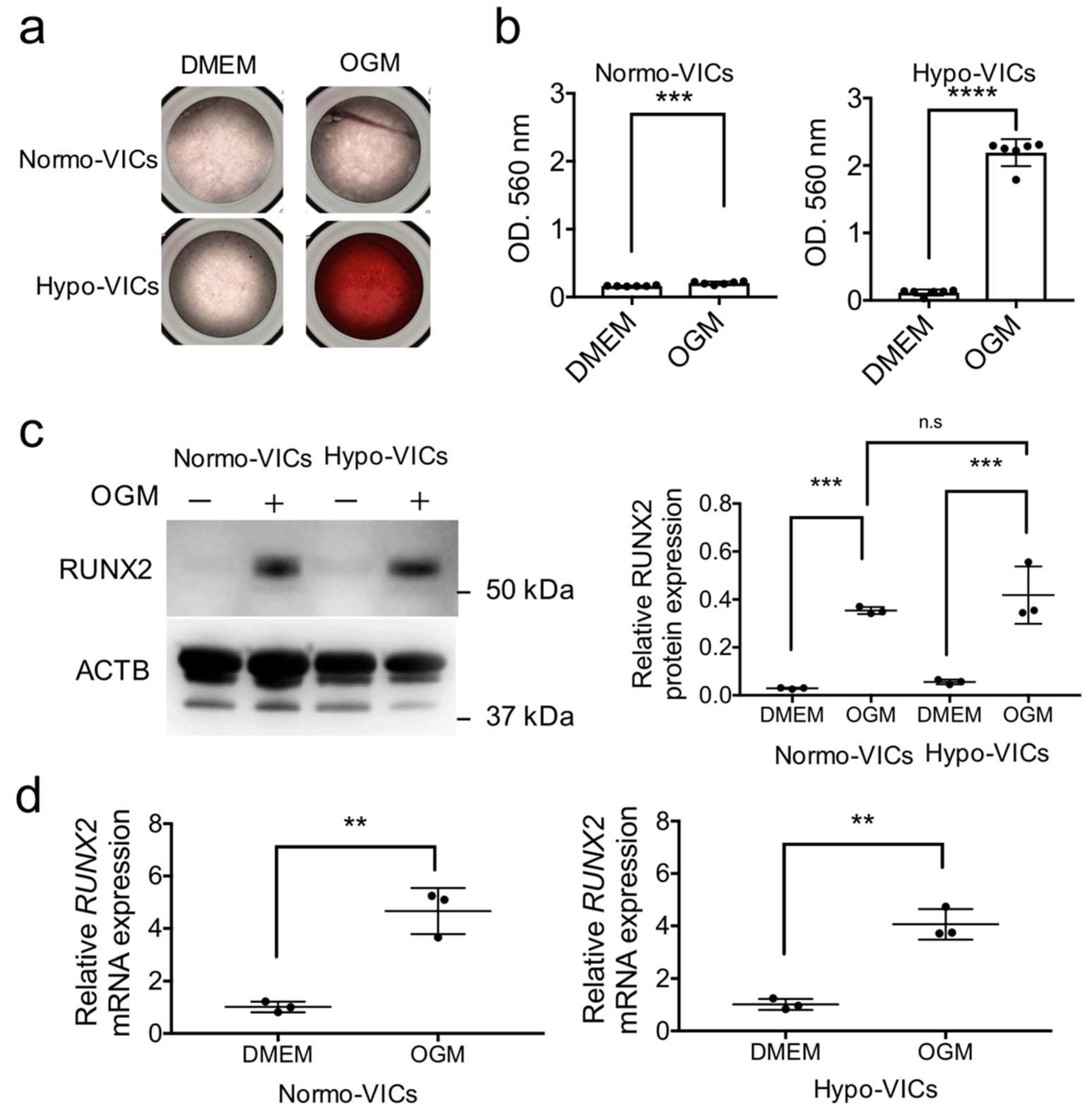
2.3. Differentiation of Hypo-VICs into Osteoblasts
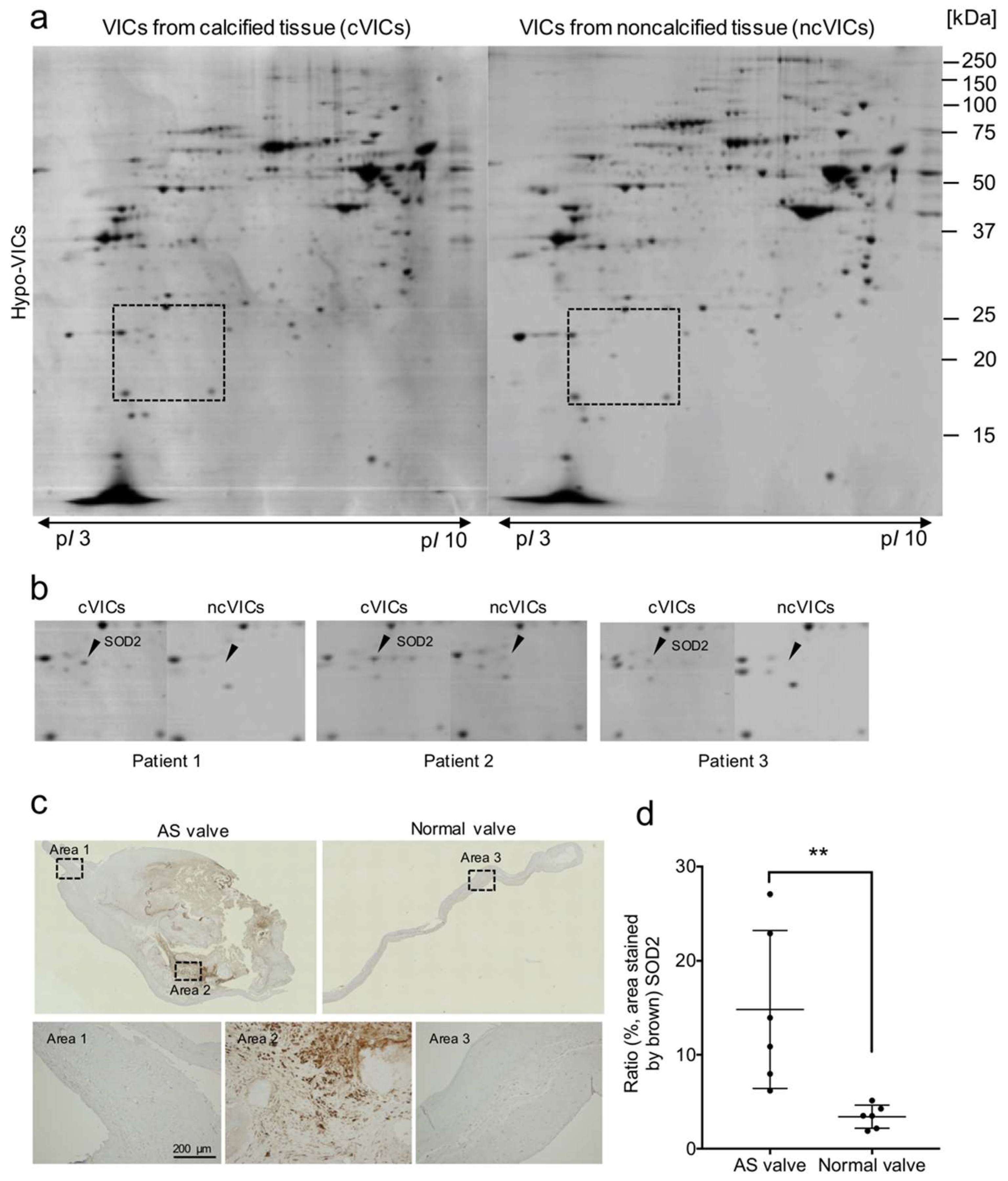
2.4. Expression of the Oxidative Stress Marker SOD2 Is Increased in VICs from Calcified AS Tissues
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Patients and Ethical Approval
4.3. VICs Isolation and Culture
4.4. VIC Proliferation
4.5. In Vitro Osteoblast Differentiation
4.6. ROS Assays
4.7. RNA Isolation and Sequencing
4.8. Real-Time RT-qPCR
4.9. Western Blotting
4.10. Two-Dimensional Gel Electrophoresis (2D-GE) and LC-MS/MS
4.11. Immunohistochemistry
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nkomo, V.T.; Gardin, J.M.; Skelton, T.N.; Gottdiener, J.S.; Scott, C.G.; Enriquez-Sarano, M. Burden of valvular heart diseases: A population-based study. Lancet 2006, 368, 1005–1011. [Google Scholar] [CrossRef]
- Yutzey, K.E.; Demer, L.L.; Body, S.C.; Huggins, G.S.; Towler, D.A.; Giachelli, C.M.; Hofmann-Bowman, M.A.; Mortlock, D.P.; Rogers, M.B.; Sadeghi, M.M.; et al. Calcific aortic valve disease: A consensus summary from the Alliance of Investigators on Calcific Aortic Valve Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2387–2393. [Google Scholar] [CrossRef] [Green Version]
- Rajamannan, N.M.; Subramaniam, M.; Rickard, D.; Stock, S.R.; Donovan, J.; Springett, M.; Orszulak, T.; Fullerton, D.A.; Tajik, A.J.; Bonow, R.O.; et al. Human aortic valve calcification is associated with an osteoblast phenotype. Circulation 2003, 107, 2181–2184. [Google Scholar] [CrossRef]
- Mohler, E.R., 3rd; Gannon, F.; Reynolds, C.; Zimmerman, R.; Keane, M.G.; Kaplan, F.S. Bone formation and inflammation in cardiac valves. Circulation 2001, 103, 1522–1528. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.H.; Yip, C.Y.; Sone, E.D.; Simmons, C.A. Identification and characterization of aortic valve mesenchymal progenitor cells with robust osteogenic calcification potential. Am. J. Pathol. 2009, 174, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Sun, W.; Wu, S.M.; Xiao, J.; Kong, X. Telocytes in human heart valves. J. Cell. Mol. Med. 2014, 18, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Chow, D.C.; Wenning, L.A.; Miller, W.M.; Papoutsakis, E.T. Modeling pO(2) distributions in the bone marrow hematopoietic compartment. I. Krogh’s model. Biophys. J. 2001, 81, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Chow, D.C.; Wenning, L.A.; Miller, W.M.; Papoutsakis, E.T. Modeling pO(2) distributions in the bone marrow hematopoietic compartment. II. Modified Kroghian models. Biophys. J. 2001, 81, 685–696. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.C.; Chen, Y.J.; Yew, T.L.; Chen, L.L.; Wang, J.Y.; Chiu, C.H.; Hung, S.C. Hypoxia inhibits senescence and maintains mesenchymal stem cell properties through down-regulation of E2A-p21 by HIF-TWIST. Blood 2011, 117, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Valorani, M.G.; Montelatici, E.; Germani, A.; Biddle, A.; D’Alessandro, D.; Strollo, R.; Patrizi, M.P.; Lazzari, L.; Nye, E.; Otto, W.R.; et al. Pre-culturing human adipose tissue mesenchymal stem cells under hypoxia increases their adipogenic and osteogenic differentiation potentials. Cell Prolif. 2012, 45, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, C.; Toki, F.; Shiraishi, K.; Sayama, K.; Nishimura, E.K.; Miura, H.; Higashiyama, S.; Nanba, D. Two clonal types of human skin fibroblasts with different potentials for proliferation and tissue remodeling ability. J. Dermatol. Sci. 2016, 82, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Sakaue, T.; Hamaguchi, M.; Aono, J.; Nakashiro, K.I.; Shikata, F.; Kawakami, N.; Oshima, Y.; Kurata, M.; Nanba, D.; Masumoto, J.; et al. Valve Interstitial Cell-Specific Cyclooxygenase-1 Associated With Calcification of Aortic Valves. Ann. Thorac. Surg. 2020, 110, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Brandl, A.; Meyer, M.; Bechmann, V.; Nerlich, M.; Angele, P. Oxidative stress induces senescence in human mesenchymal stem cells. Exp. Cell Res. 2011, 317, 1541–1547. [Google Scholar] [CrossRef]
- Sun, Y.; Coppe, J.P.; Lam, E.W. Cellular Senescence: The Sought or the Unwanted? Trends Mol. Med. 2018, 24, 871–885. [Google Scholar] [CrossRef]
- Kakudo, N.; Morimoto, N.; Ogawa, T.; Taketani, S.; Kusumoto, K. Hypoxia Enhances Proliferation of Human Adipose-Derived Stem Cells via HIF-1a Activation. PLoS ONE 2015, 10, e0139890. [Google Scholar]
- Lv, B.; Li, F.; Fang, J.; Xu, L.; Sun, C.; Han, J.; Hua, T.; Zhang, Z.; Feng, Z.; Jiang, X. Hypoxia inducible factor 1alpha promotes survival of mesenchymal stem cells under hypoxia. Am. J. Transl. Res. 2017, 9, 1521–1529. [Google Scholar]
- Zhang, B.; Li, Y.L.; Zhao, J.L.; Zhen, O.; Yu, C.; Yang, B.H.; Yu, X.R. Hypoxia-inducible factor-1 promotes cancer progression through activating AKT/Cyclin D1 signaling pathway in osteosarcoma. Biomed. Pharm. 2018, 105, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bhanja, A.; Bhattacharyya, J.; Jaganathan, B.G. Multiple roles of CD90 in cancer. Tumour Biol. 2016, 37, 11611–11622. [Google Scholar] [CrossRef] [PubMed]
- Harting, M.; Jimenez, F.; Pati, S.; Baumgartner, J.; Cox, C., Jr. Immunophenotype characterization of rat mesenchymal stromal cells. Cytotherapy 2008, 10, 243–253. [Google Scholar] [CrossRef]
- Wieczorek, G.; Steinhoff, C.; Schulz, R.; Scheller, M.; Vingron, M.; Ropers, H.H.; Nuber, U.A. Gene expression profile of mouse bone marrow stromal cells determined by cDNA microarray analysis. Cell Tissue Res. 2003, 311, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Vieira, N.M.; Brandalise, V.; Zucconi, E.; Secco, M.; Strauss, B.E.; Zatz, M. Isolation, characterization, and differentiation potential of canine adipose-derived stem cells. Cell Transplant. 2010, 19, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Haynesworth, S.E.; Goshima, J.; Goldberg, V.M.; Caplan, A.I. Characterization of cells with osteogenic potential from human marrow. Bone 1992, 13, 81–88. [Google Scholar] [CrossRef]
- Yang, Z.X.; Han, Z.B.; Ji, Y.R.; Wang, Y.W.; Liang, L.; Chi, Y.; Yang, S.G.; Li, L.N.; Luo, W.F.; Li, J.P.; et al. CD106 identifies a subpopulation of mesenchymal stem cells with unique immunomodulatory properties. PLoS ONE 2013, 8, e59354. [Google Scholar] [CrossRef] [Green Version]
- Espagnolle, N.; Guilloton, F.; Deschaseaux, F.; Gadelorge, M.; Sensebe, L.; Bourin, P. CD146 expression on mesenchymal stem cells is associated with their vascular smooth muscle commitment. J. Cell. Mol. Med. 2014, 18, 104–114. [Google Scholar] [CrossRef]
- Zhu, H.; Mitsuhashi, N.; Klein, A.; Barsky, L.W.; Weinberg, K.; Barr, M.L.; Demetriou, A.; Wu, G.D. The role of the hyaluronan receptor CD44 in mesenchymal stem cell migration in the extracellular matrix. Stem Cells 2006, 24, 928–935. [Google Scholar] [CrossRef]
- Traktuev, D.O.; Merfeld-Clauss, S.; Li, J.; Kolonin, M.; Arap, W.; Pasqualini, R.; Johnstone, B.H.; March, K.L. A population of multipotent CD34-positive adipose stromal cells share pericyte and mesenchymal surface markers, reside in a periendothelial location, and stabilize endothelial networks. Circ. Res. 2008, 102, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, K.; Shigeura, T.; Matsumoto, D.; Sato, T.; Takaki, Y.; Aiba-Kojima, E.; Sato, K.; Inoue, K.; Nagase, T.; Koshima, I.; et al. Characterization of freshly isolated and cultured cells derived from the fatty and fluid portions of liposuction aspirates. J. Cell Physiol. 2006, 208, 64–76. [Google Scholar] [CrossRef]
- Lis, G.J.; Dubrowski, A.; Lis, M.; Solewski, B.; Witkowska, K.; Aleksandrovych, V.; Jasek-Gajda, E.; Holda, M.K.; Gil, K.; Litwin, J.A. Identification of CD34+/PGDFRalpha+ Valve Interstitial Cells (VICs) in Human Aortic Valves: Association of Their Abundance, Morphology and Spatial Organization with Early Calcific Remodeling. Int. J. Mol. Sci. 2020, 21, 6330. [Google Scholar] [CrossRef] [PubMed]
- Nomura, A.; Seya, K.; Yu, Z.; Daitoku, K.; Motomura, S.; Murakami, M.; Fukuda, I.; Furukawa, K. CD34-negative mesenchymal stem-like cells may act as the cellular origin of human aortic valve calcification. Biochem. Biophys. Res. Commun. 2013, 440, 780–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.A.; Guo, W.; Liao, M.J.; Eaton, E.N.; Ayyanan, A.; Zhou, A.Y.; Brooks, M.; Reinhard, F.; Zhang, C.C.; Shipitsin, M.; et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell 2008, 133, 704–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, X.C.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.Y.; Li, X.M.; Luo, S.Q. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB. Biochem. Biophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef]
- Mody, N.; Parhami, F.; Sarafian, T.A.; Demer, L.L. Oxidative stress modulates osteoblastic differentiation of vascular and bone cells. Free Radic. Biol. Med. 2001, 31, 509–519. [Google Scholar] [CrossRef]
- Martin-Rojas, T.; Gil-Dones, F.; Lopez-Almodovar, L.F.; Padial, L.R.; Vivanco, F.; Barderas, M.G. Proteomic profile of human aortic stenosis: Insights into the degenerative process. J. Proteome Res. 2012, 11, 1537–1550. [Google Scholar] [CrossRef]
- Suzuki, H.; Chikada, M.; Yokoyama, M.K.; Kurokawa, M.S.; Ando, T.; Furukawa, H.; Arito, M.; Miyairi, T.; Kato, T. Aberrant Glycosylation of Lumican in Aortic Valve Stenosis Revealed by Proteomic Analysis. Int. Heart J. 2016, 57, 104–111. [Google Scholar] [CrossRef] [Green Version]
- Torre, M.; Hwang, D.H.; Padera, R.F.; Mitchell, R.N.; VanderLaan, P.A. Osseous and chondromatous metaplasia in calcific aortic valve stenosis. Cardiovasc. Pathol. 2016, 25, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Nigam, V.; Srivastava, D. Notch1 represses osteogenic pathways in aortic valve cells. J. Mol. Cell. Cardiol. 2009, 47, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Hortells, L.; Sur, S.; St Hilaire, C. Cell Phenotype Transitions in Cardiovascular Calcification. Front. Cardiovasc. Med. 2018, 5, 27. [Google Scholar] [CrossRef]
- Jiang, W.; Zhang, Z.; Li, Y.; Chen, C.; Yang, H.; Lin, Q.; Hu, M.; Qin, X. The Cell Origin and Role of Osteoclastogenesis and Osteoblastogenesis in Vascular Calcification. Front. Cardiovasc. Med. 2021, 8, 639740. [Google Scholar] [CrossRef] [PubMed]
- Saeki, N.; Imai, Y. Reprogramming of synovial macrophage metabolism by synovial fibroblasts under inflammatory conditions. Cell Commun. Signal 2020, 18, 188. [Google Scholar] [CrossRef] [PubMed]
- Sakaue, T.; Shikata, F.; Utsunomiya, K.; Fukae, S.; Kurata, M.; Nakaoka, H.; Okazaki, M.; Kawanishi, Y.; Kojima, A.; Higashiyama, S.; et al. Proteomics-based analysis of lung injury-induced proteins in a mouse model of common bile duct ligation. Surgery 2017, 161, 1525–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession No. | Gene Name | Score | Sequence Coverage (%) | Matched Peptides | Matched Amino Acids | Nominal Mass [kDa] | Calculated pI |
|---|---|---|---|---|---|---|---|
| P04179 | Superoxide dismutase [Mn], mitochondrial (SOD2) | 2922 | 68.92 | 15 | 222 | 24.73 | 8.25 |
| Target Gene | Direction | Sequence (5′-3′) |
|---|---|---|
| Thy1 | forward | GAAGGTCCTCTACTTATCCGCC |
| reverse | TGATGCCCTCACACTTGACCAG | |
| CD34 | forward | CCTCAGTGTCTACTGCTGGTCT |
| reverse | GGAATAGCTCTGGTGGCTTGCA | |
| Oct4 | forward | CCTGAAGCAGAAGAGGATCACC |
| reverse | AAAGCGGCAGATGGTCGTTTGG | |
| CD29 | forward | GGATTCTCCAGAAGGTGGTTTCG |
| reverse | TGCCACCAAGTTTCCCATCTCC | |
| ENG | forward | CGGTGGTCAATATCCTGTCGAG |
| reverse | AGGAAGTGTGGGCTGAGGTAGA | |
| VCAM1 | forward | GATTCTGTGCCCACAGTAAGGC |
| reverse | TGGTCACAGAGCCACCTTCTTG | |
| CD44 | forward | CCAGAAGGAACAGTGGTTTGGC |
| reverse | ACTGTCCTCTGGGCTTGGTGTT | |
| MCAM | forward | ATCGCTGCTGAGTGAACCACAG |
| reverse | CTACTCTCTGCCTCACAGGTCA | |
| ALCAM | forward | TCCAGAACACGATGAGGCAGAC |
| reverse | GTAGACGACACCAGCAACAAGG | |
| RUNX2 | forward | TTCATCCCTCACTGAGAG |
| reverse | TCAGCGTCAACACCATCA | |
| GAPDH | forward | CATGAGAAGTATGACAACAGCCT |
| reverse | AGTCCTTCCACGATACCAAAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanno, K.; Sakaue, T.; Hamaguchi, M.; Namiguchi, K.; Nanba, D.; Aono, J.; Kurata, M.; Masumoto, J.; Higashiyama, S.; Izutani, H. Hypoxic Culture Maintains Cell Growth of the Primary Human Valve Interstitial Cells with Stemness. Int. J. Mol. Sci. 2021, 22, 10534. https://doi.org/10.3390/ijms221910534
Kanno K, Sakaue T, Hamaguchi M, Namiguchi K, Nanba D, Aono J, Kurata M, Masumoto J, Higashiyama S, Izutani H. Hypoxic Culture Maintains Cell Growth of the Primary Human Valve Interstitial Cells with Stemness. International Journal of Molecular Sciences. 2021; 22(19):10534. https://doi.org/10.3390/ijms221910534
Chicago/Turabian StyleKanno, Kaho, Tomohisa Sakaue, Mika Hamaguchi, Kenji Namiguchi, Daisuke Nanba, Jun Aono, Mie Kurata, Junya Masumoto, Shigeki Higashiyama, and Hironori Izutani. 2021. "Hypoxic Culture Maintains Cell Growth of the Primary Human Valve Interstitial Cells with Stemness" International Journal of Molecular Sciences 22, no. 19: 10534. https://doi.org/10.3390/ijms221910534