
Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
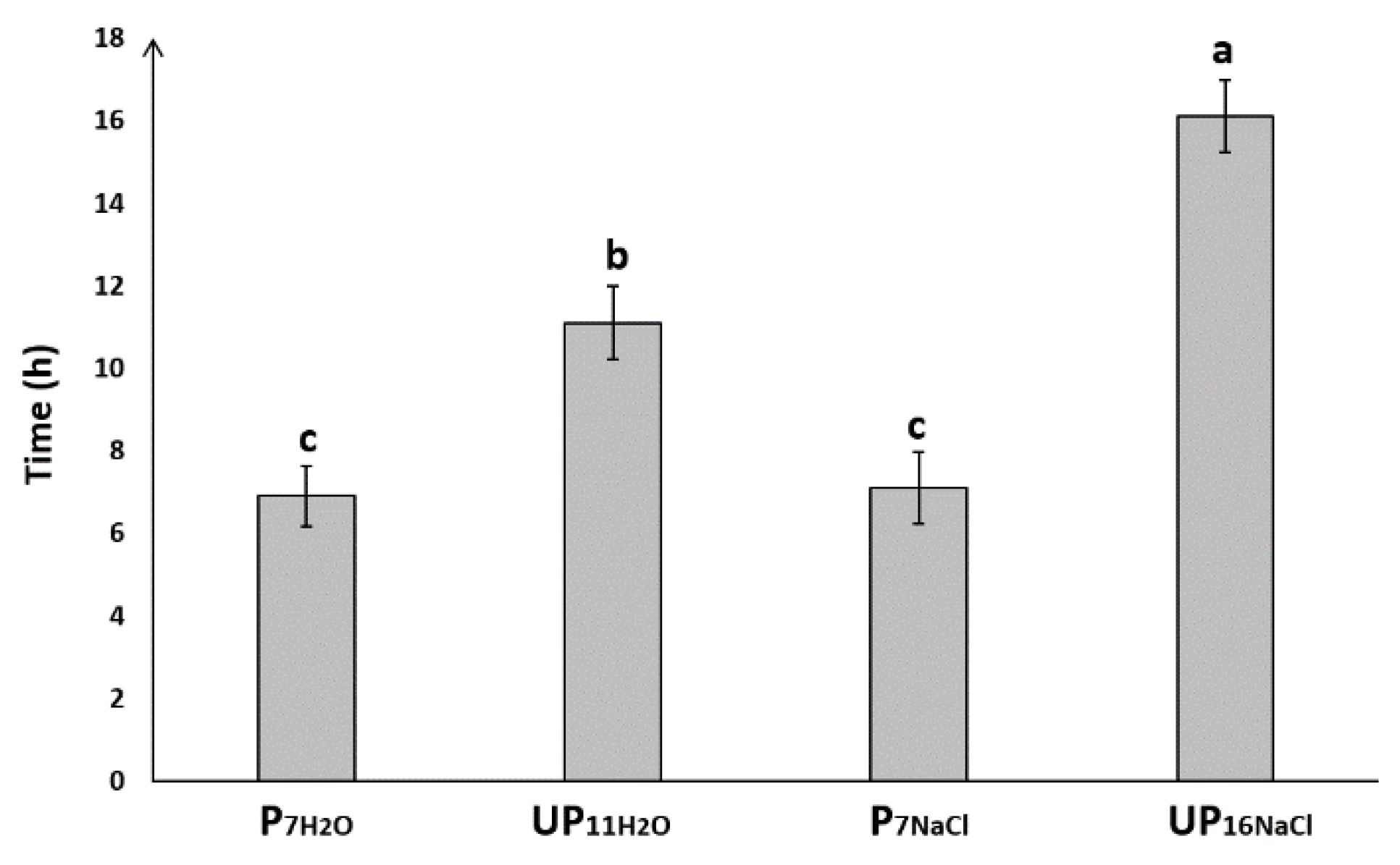
2.1. Priming Improves Seed Germination under Salt Stress
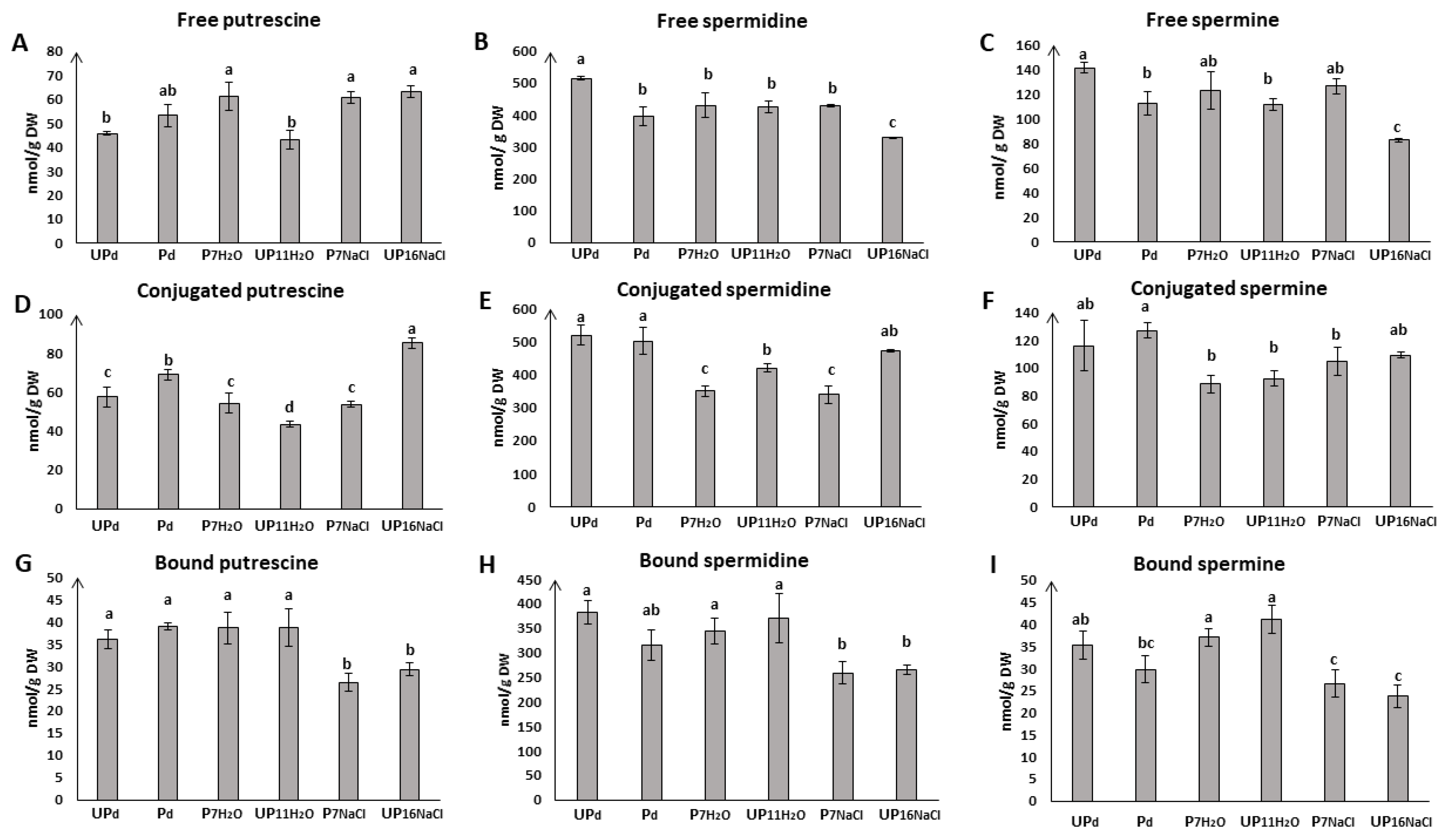
2.2. Seed Priming Modulates Pattern of Endogenous Polyamine Accumulation during Germination under Salt Stress
2.2.1. Seed Priming Modulates the Content of Free Polyamines
Free Putrescine
Free Spermidine and Spermine
2.2.2. Seed Priming Modulates the Content of Conjugated Polyamines
Conjugated Putrescine
Conjugated Spermidine
Conjugated Spermine
2.2.3. Seed Priming Modulates the Content of Bound Polyamines
2.3. Seed Priming Modulates PAs Ratios
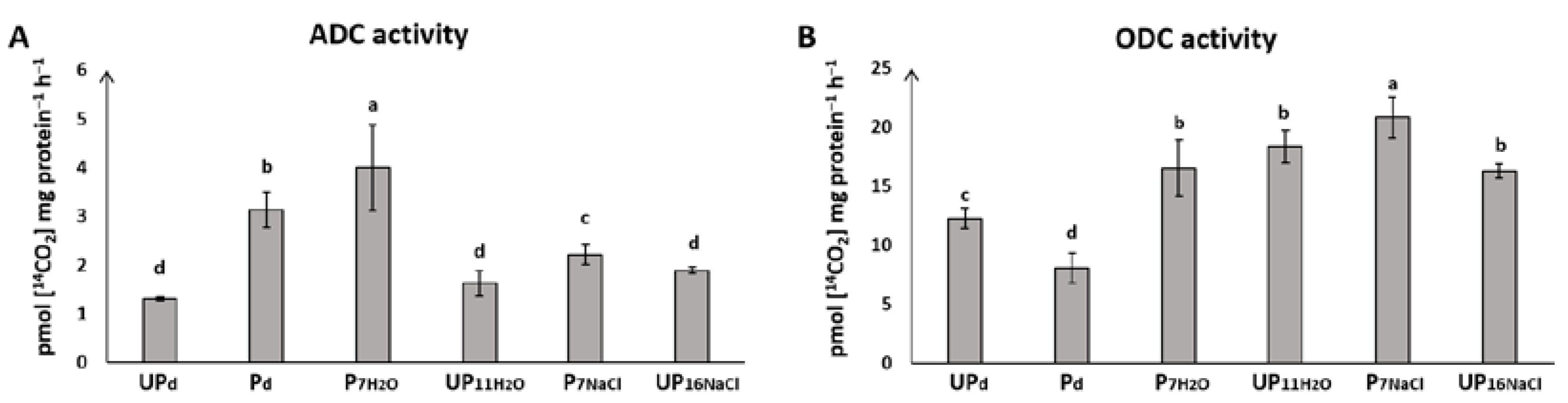
2.4. Seed Priming Modulates ODC and ADC Activity during Seed Germination under Salt Stress
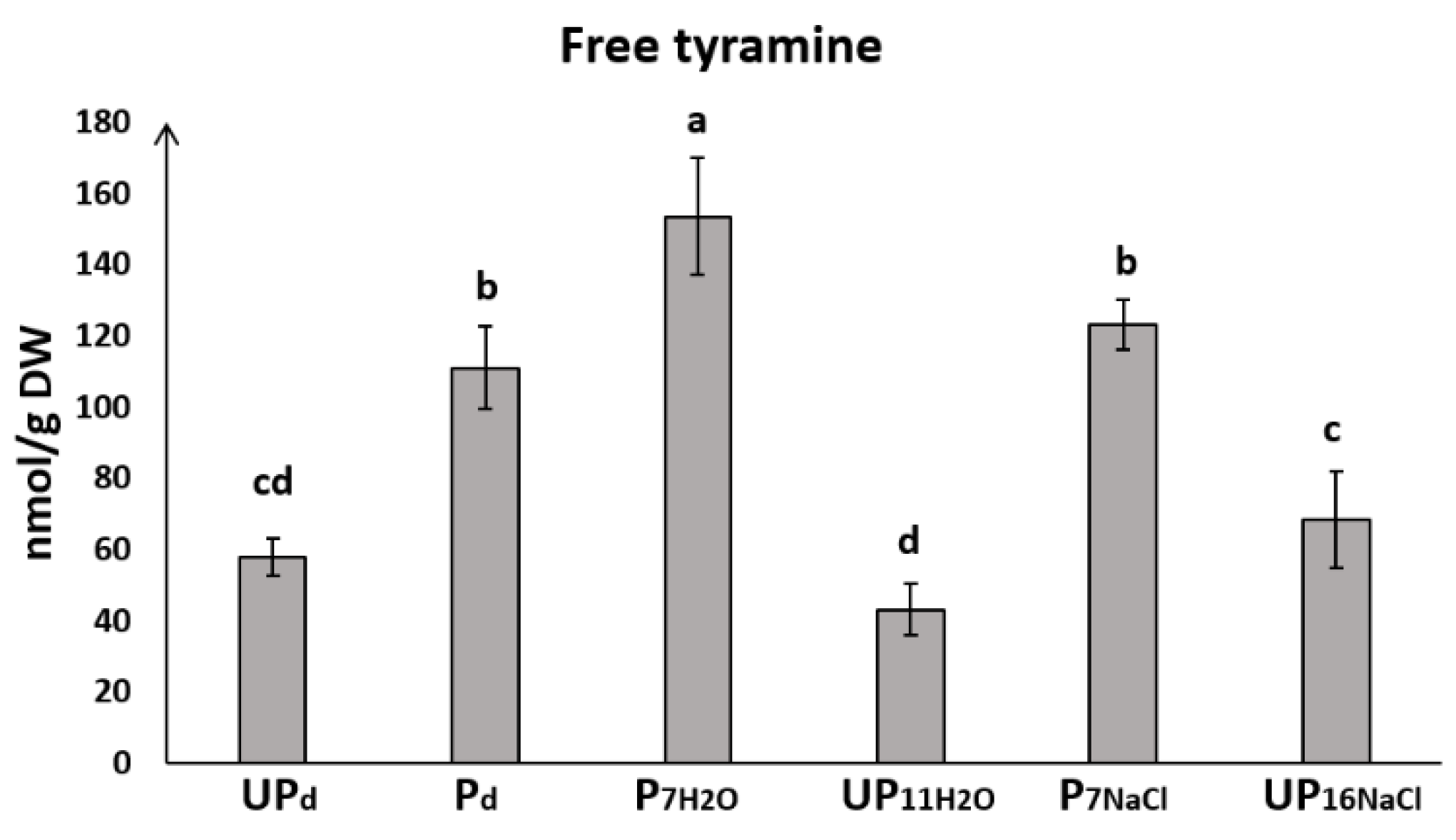
2.5. Seed Priming Modulates the Level of Free Tyramine
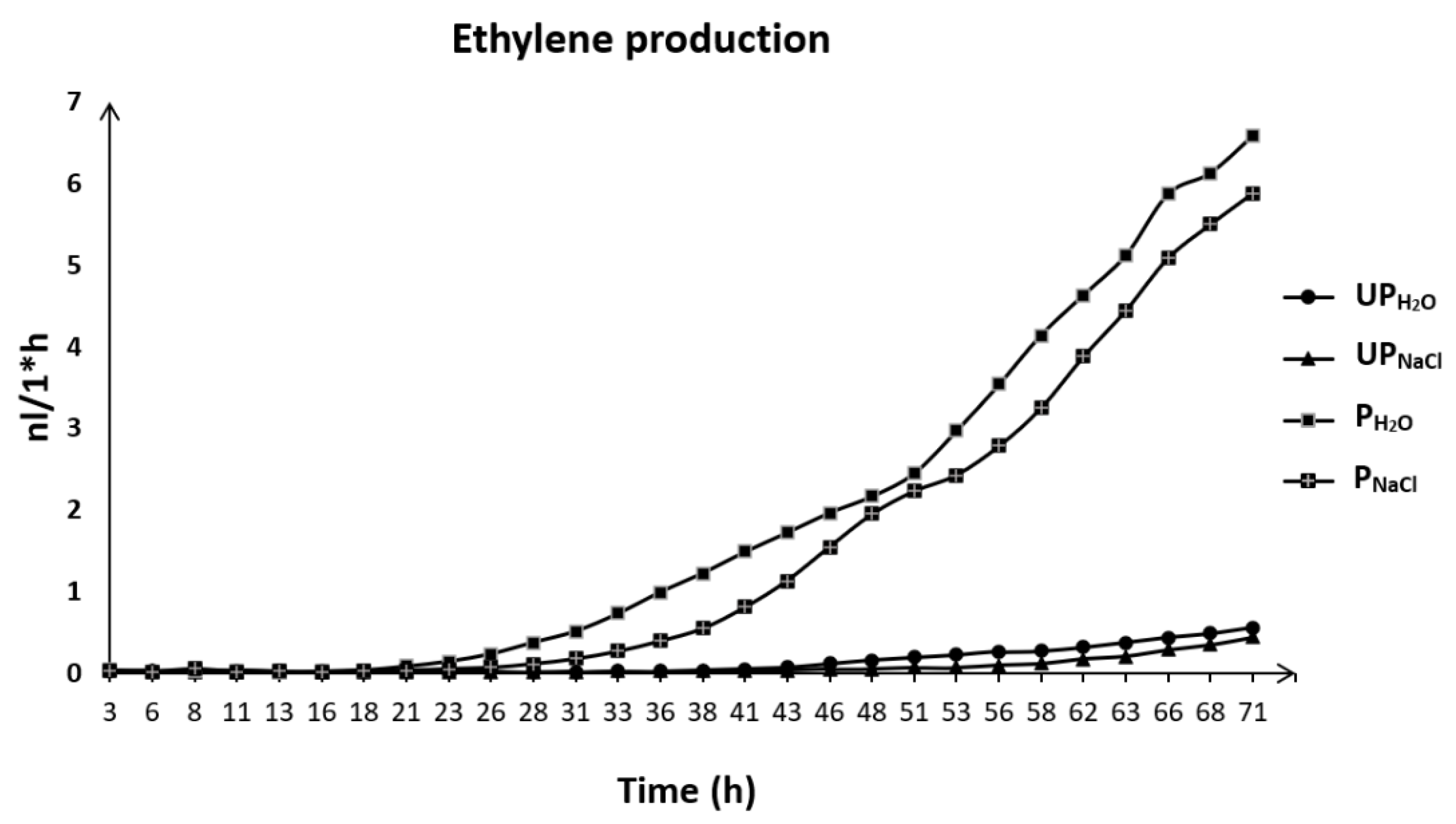
2.6. Priming-Induced Changes in Ethylene Production during Early Seedling Growth under Salt Stress
3. Discussion
4. Materials and Methods
4.1. Seed Osmopriming Procedure
4.2. Germination Conditions, Gemination Test and Experimental Design
4.3. Extraction and Quantification of PAs and Tyramine
4.3.1. Free Polyamines and Tyramine Extraction
4.3.2. Conjugated Polyamine Extraction
4.3.3. Bound Polyamine Extraction
4.3.4. Sample Dansylation and HPLC Analysis
4.4. Enzyme Assay
4.5. Ethylene Measurement
4.6. Statistical Analysis
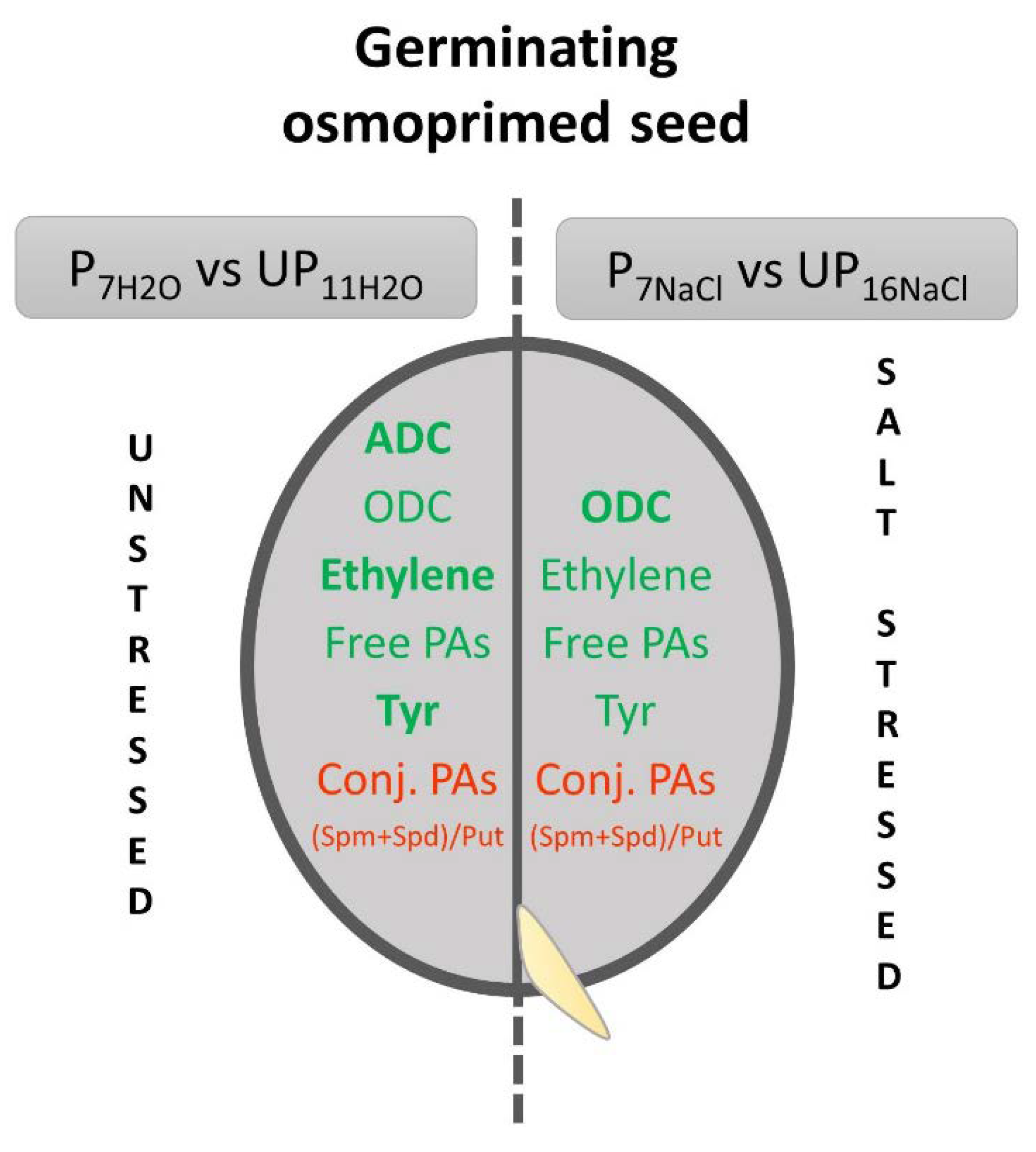
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADC | Arginine decarboxylase |
| dcSAM | Decarboxylated S-adenosylmethionine |
| ODC | Ornithine decarboxylase |
| P5CS | Pyrroline-5-carboxylate synthetase gene |
| P7H2O | Primed seeds germinating for 7 h in water |
| P7NaCl | Primed seeds germinating for 7 h in NaCl |
| PAs | Polyamines |
| Pd | Primed dried seeds |
| PDH | Proline dehydrogenase gene |
| PEG | Polyethylene glycol |
| Pro | Proline |
| Put | Putrescine |
| SAM | S-adenosylmethionine |
| Spd | Spermidine |
| Spm | Spermine |
| Tyr | Tyramine |
| UP11H2O | Unprimed seeds germinating 11 h in water |
| UP16NaCl | Unprimed seeds germinating 16 h in NaCl |
| UPd | Dry unprimed seeds |
References
- Global Map of Salt-Affected Soils|FAO SOILS PORTAL|Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 18 December 2021).
- Zaman, M.; Shahid, S.A.; Pharis, R.P. Salinity a Serious Threat to Food Security–Where Do We Stand? Soils Newsl. 2016, 39, 9–10. [Google Scholar]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Zaman, M., Shahid, S.A., Heng, L., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 43–53. ISBN 978-3-319-96190-3. [Google Scholar]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the Physiological and Biochemical Impacts of Salt Stress on Plant Growth and Development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, e701596. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Genes and Salt Tolerance: Bringing Them Together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Hanin, M.; Ebel, C.; Ngom, M.; Laplaze, L.; Masmoudi, K. New Insights on Plant Salt Tolerance Mechanisms and Their Potential Use for Breeding. Front. Plant Sci. 2016, 7, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant Salinity Stress: Many Unanswered Questions Remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Rejeb, I.B.; Pastor, V.; Mauch-Mani, B. Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants (Basel) 2014, 3, 458–475. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt Stress in Maize: Effects, Resistance Mechanisms, and Management. A Review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Guo, J.; Wang, C.; Li, K.; Zhang, X.; Yang, Z.; Li, M.; Wang, B. An Effective Screening Method and a Reliable Screening Trait for Salt Tolerance of Brassica Napus at the Germination Stage. Front. Plant Sci. 2019, 10, 530. [Google Scholar] [CrossRef]
- Wahid, A.; Farooq, M.; Basra, S.M.A.; Rasul, E.; Siddique, K. Germination of Seeds and Propagules under Salt Stress. In Handbook of Plant and Crop Stress, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 321–337. [Google Scholar]
- Wojtyla, L.; Lechowska, K.; Kubala, S.; Garnczarska, M. Molecular Processes Induced in Primed Seeds-Increasing the Potential to Stabilize Crop Yields under Drought Conditions. J. Plant Physiol. 2016, 203, 116–126. [Google Scholar] [CrossRef]
- Lutts, S.; Benincasa, P.; Wojtyla, L.; Kubala, S.; Pace, R.; Lechowska, K.; Quinet, M.; Garnczarska, M. Seed Priming: New Comprehensive Approaches for an Old Empirical Technique. In New Challenges in Seed Biology–Basic and Translational Research Driving Seed Technology; Araújo, S., Balestrazzi, A., Eds.; InTechOpen: London, UK, 2016. [Google Scholar]
- Marthandan, V.; Geetha, R.; Kumutha, K.; Renganathan, V.G.; Karthikeyan, A.; Ramalingam, J. Seed Priming: A Feasible Strategy to Enhance Drought Tolerance in Crop Plants. Int. J. Mol. Sci. 2020, 21, 8258. [Google Scholar] [CrossRef]
- Kubala, S.; Garnczarska, M.; Wojtyla, Ł.; Clippe, A.; Kosmala, A.; Żmieńko, A.; Lutts, S.; Quinet, M. Deciphering Priming-Induced Improvement of Rapeseed (Brassica napus L.) Germination through an Integrated Transcriptomic and Proteomic Approach. Plant Sci. 2015, 231, 94–113. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.P.; Hamza, J.H.; Rihan, H.Z.; Al-Issawi, M. Germination of Primed Seed under NaCl Stress in Wheat. ISRN Bot. 2012, 2012, e167804. [Google Scholar] [CrossRef] [Green Version]
- Feghhenabi, F.; Hadi, H.; Khodaverdiloo, H.; van Genuchten, M.T. Seed Priming Alleviated Salinity Stress during Germination and Emergence of Wheat (Triticum aestivum L.). Agric. Water Manag. 2020, 231, 106022. [Google Scholar] [CrossRef]
- Theerakulpisut, P.; Kanawapee, N.; Panwong, B. Seed Priming Alleviated Salt Stress Effects on Rice Seedlings by Improving Na+/K+ and Maintaining Membrane Integrity. Int. J. Plant Biol. 2016, 7, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Rauf, H. Inducing Salt Tolerance in Maize (Zea mays L.) through Seed Priming with Chloride Salts: Growth and Ion Transport at Early Growth Stages. Acta Physiol. Plant 2001, 23, 407–414. [Google Scholar] [CrossRef]
- Imran, M.; Boelt, B.; Mühling, K.-H. Zinc Seed Priming Improves Salt Resistance in Maize. J. Agron. Crop Sci. 2018, 204, 390–399. [Google Scholar] [CrossRef]
- Soliman, M.; Elkelish, A.; Souad, T.; Alhaithloul, H.; Farooq, M. Brassinosteroid Seed Priming with Nitrogen Supplementation Improves Salt Tolerance in Soybean. Physiol. Mol. Biol. Plants 2020, 26, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, C.L.; Araújo, H.T.N.; de Brito, S.F.; da Maia, M.S.; da Viana, J.S.; Filho, S.M. Seed Priming and Tolerance to Salt and Water Stress in Divergent Grain Sorghum Genotypes. Am. J. Plant Sci. 2018, 9, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Lechowska, K.; Kubala, S.; Wojtyla, Ł.; Nowaczyk, G.; Quinet, M.; Lutts, S.; Garnczarska, M. New Insight on Water Status in Germinating Brassica Napus Seeds in Relation to Priming-Improved Germination. Int. J. Mol. Sci. 2019, 20, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubala, S.; Wojtyla, Ł.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced Expression of the Proline Synthesis Gene P5CSA in Relation to Seed Osmopriming Improvement of Brassica Napus Germination under Salinity Stress. J. Plant Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. Food Outlook–Biannual Report on Global Food Markets: June 2021; Food Outlook; FAO: Rome, Italy, 2021; ISBN 978-92-5-134334-0. [Google Scholar]
- Lutts, S.; Hausman, J.; Quinet, M.; Lefèvre, I. Polyamines and Their Roles in the Alleviation of Ion Toxicities in Plants. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M., Prasad, M., Eds.; Springer: New York, NY, USA, 2013; pp. 315–353. [Google Scholar]
- Kusano, T.; Berberich, T.; Tateda, C.; Takahashi, Y. Polyamines: Essential Factors for Growth and Survival. Planta 2008, 228, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Verma, R.; Singh, K.; Nisha, N.; Keisham, M.; Bhati, K.K.; Kim, S.T.; Gupta, R. Ethylene: A Master Regulator of Salinity Stress Tolerance in Plants. Biomolecules 2020, 10, 959. [Google Scholar] [CrossRef]
- Ghabriche, R.; Ghnaya, T.; Mnasri, M.; Zaier, H.; Baioui, R.; Vromman, D.; Abdelly, C.; Lutts, S. Polyamine and Tyramine Involvement in NaCl-Induced Improvement of Cd Resistance in the Halophyte Inula chrithmoides L. J. Plant Physiol. 2017, 216, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Krasuska, U.; Ciacka, K.; Bogatek, R.; Gniazdowska, A. Polyamines and Nitric Oxide Link in Regulation of Dormancy Removal and Germination of Apple (Malus domestica Borkh.) Embryos. J. Plant Growth Regul. 2014, 33, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Basra, A.S.; Singh, B.; Malik, C.P. Amelioration of the Effects of Ageing in Onion Seeds by Osmotic Priming and Associated Changes in Oxidative Metabolism. Biol. Plant. 1994, 36, 365–371. [Google Scholar] [CrossRef]
- Bueno, M.; Cordovilla, M.-P. Polyamines in Halophytes. Front. Plant Sci. 2019, 10, 439. [Google Scholar] [CrossRef] [Green Version]
- Puga-Hermida, M.I.; Gallardo, M.; Rodriguez-Gacio, M.C.; Matilla, A.J. Polyamine Contents, Ethylene Synthesis, and BrACO2 Expression during Turnip Germination. Biol. Plant 2006, 50, 574–580. [Google Scholar] [CrossRef]
- Sińska, I.; Lewandowska, U. Polyamines and Ethylene in the Removal of Embryonal Dormancy in Apple Seeds. Physiol. Plant. 1991, 81, 59–64. [Google Scholar] [CrossRef]
- Aragão, V.; Navarro, B.; Zanchetta Passamani, L.; Macedo, A.; Floh, E.; Silveira, V.; Santa Catarina, C. Free Amino Acids, Polyamines, Soluble Sugars and Proteins during Seed Germination and Early Seedling Growth of Cedrela Fissilis Vellozo (Meliaceae), an Endangered Hardwood Species from the Atlantic Forest in Brazil. Theor. Exp. Plant Physiol. 2015, 27, 157–169. [Google Scholar] [CrossRef]
- Huang, Y.; Lin, C.; He, F.; Li, Z.; Guan, Y.; Hu, Q.; Hu, J. Exogenous Spermidine Improves Seed Germination of Sweet Corn via Involvement in Phytohormone Interactions, H2O2 and Relevant Gene Expression. BMC Plant Biol. 2017, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- Mirza, J.I.; Bagni, N. Effects of Exogenous Polyamines and Difluoromethylornithine on Seed Germination and Root Growth of Arabidopsis Thaliana. Plant Growth Regul. 1991, 10, 163–168. [Google Scholar] [CrossRef]
- Sidhu, G.K.; Tuan, P.A.; Renault, S.; Daayf, F.; Ayele, B.T. Polyamine-Mediated Transcriptional Regulation of Enzymatic Antioxidative Response to Excess Soil Moisture during Early Seedling Growth in Soybean. Biology 2020, 9, 185. [Google Scholar] [CrossRef] [PubMed]
- Kubala, S.; Wojtyla, L.; Garnczarska, M. Seed Priming Improves Salt Stress Tolerance during Germination by Modulation of Antioxidative Capacity. BioTechnologia 2013, 94, 223. [Google Scholar]
- Wojtyla, L.; Kubala, S.; Lechowska, K.; Garnczarska, M. Modulation of Antioxidative Metabolism in Response to Osmopriming in Brassica Napus Seeds Improves Germination under Salt Stress. BioTechnologia 2013, 94, 394. [Google Scholar]
- Lechowska, K.; Wojtyla, Ł.; Garnczarska, M. The Impact of Seed Hydropriming on Antioxidative Defense System of Brassica Napus during Germination and Growth under Saline Conditions. In Communication in Plants: From Cell to Environment; Cieresszko, I., Bajguz, A., Eds.; Polish Society of Experimental Plant Biology: Białystok, Poland, 2017; p. 136. [Google Scholar]
- Dhansu, P.; Goyal, V.; Devi, S.; Munjal, R. Role of Polyamines in Protecting Plants from Oxidative Stress. In Reactive Oxygen, Nitrogen and Sulfur Species in Plants: Production, Metabolism, Signaling and Defense Mechanisms; Wiley: Hoboken, NJ, USA, 2019; pp. 143–157. ISBN 978-1-119-46867-7. [Google Scholar]
- Pieruzzi, F.P.; Dias, L.L.C.; Balbuena, T.S.; Santa-Catarina, C.; dos Santos, A.L.W.; Floh, E.I.S. Polyamines, IAA and ABA during Germination in Two Recalcitrant Seeds: Araucaria Angustifolia (Gymnosperm) and Ocotea Odorifera (Angiosperm). Ann. Bot. 2011, 108, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Paul, S.; Roychoudhury, A. Seed Priming with Spermine and Spermidine Regulates the Expression of Diverse Groups of Abiotic Stress-Responsive Genes during Salinity Stress in the Seedlings of Indica Rice Varieties. Plant Gene 2017, 11, 124–132. [Google Scholar] [CrossRef]
- Minocha, R.; Shortle, W.C.; Coughin, D.J.J.; Minocha, S.C. Effects of Aluminum on Growth, Polyamine Metabolism, and Inorganic Ions in Suspension Cultures of Red Spruce (Picea rubens). Can. J. For. Res. 1996, 26, 550–559. [Google Scholar] [CrossRef]
- Kakkar, R.K.; Nagar, P.K.; Ahuja, P. Polyamines and Plant Morphogenesis. Biol. Plant. 2000, 43, 1–11. [Google Scholar] [CrossRef]
- Chen, K.; Arora, R. Priming Memory Invokes Seed Stress-Tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Li, S.; Cui, L.; Zhang, Y.; Wang, Y.; Mao, P. The Variation Tendency of Polyamines Forms and Components of Polyamine Metabolism in Zoysiagrass (Zoysia japonica Steud.) to Salt Stress with Exogenous Spermidine Application. Front. Physiol. 2017, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Tiburcio, A.F.; Altabella, T.; Borrell, A.; Masgrau, C. Polyamine Metabolism and Its Regulation. Physiol. Plant. 1997, 100, 664–674. [Google Scholar] [CrossRef]
- Urano, K.; Hobo, T.; Shinozaki, K. Arabidopsis ADC Genes Involved in Polyamine Biosynthesis Are Essential for Seed Development. FEBS Lett. 2005, 579, 1557–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castiglione, M.R. Polyamine Metabolism during Early Germination of Triticum durum Desf. Cv. Cappelli. G. Bot. Ital. 1996, 130, 795–800. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M. Anhydrobiosis. Annu. Rev. Physiol. 1992, 54, 579–599. [Google Scholar] [CrossRef]
- Quinet, M.; Ndayiragije, A.; Lefèvre, I.; Lambillotte, B.; Dupont-Gillain, C.C.; Lutts, S. Putrescine Differently Influences the Effect of Salt Stress on Polyamine Metabolism and Ethylene Synthesis in Rice Cultivars Differing in Salt Resistance. J. Exp. Bot. 2010, 61, 2719–2733. [Google Scholar] [CrossRef]
- Corbineau, F.; Xia, Q.; Bailly, C.; El-Maarouf-Bouteau, H. Ethylene, a Key Factor in the Regulation of Seed Dormancy. Front. Plant Sci. 2014, 5, 539. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J.; et al. Salt Stress Represses Soybean Seed Germination by Negatively Regulating GA Biosynthesis While Positively Mediating ABA Biosynthesis. Front. Plant Sci. 2017, 8, 1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Diao, P.; Kong, L.; Yu, R.; Zhang, M.; Zuo, T.; Fan, Y.; Niu, Y.; Yan, F.; Wuriyanghan, H. Ethylene Enhances Seed Germination and Seedling Growth Under Salinity by Reducing Oxidative Stress and Promoting Chlorophyll Content via ETR2 Pathway. Front. Plant Sci. 2020, 11, 1066. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.P.; Kirkham, M.; Long, Z.R.; Wassom, C. Genotypic Variation in Ethylene Production by Maize Grown under Nutrient Deficiency. J. Plant Physiol. 1991, 137, 483–487. [Google Scholar] [CrossRef]
- Pennazio, S.; Roggero, P. Effects of Exogenous Salicylate on Basal and Stress-Induced Ethylene Formation in Soybean. Biol. Plant. 1991, 33, 58–65. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.-M.; Bouharmont, J. Ethylene Production by Leaves of Rice (Oryza sativa L.) in Relation to Salinity Tolerance and Exogenous Putrescine Application. Plant Sci. 1996, 116, 15–25. [Google Scholar] [CrossRef]
- Datta, K.S.; Varma, S.K.; Angrish, R.; Kumar, B.; Kumari, P. Alleviation of Salt Stress by Plant Growth Regulators in Triticum aestivum L. Biol. Plant. 1997, 40, 269–275. [Google Scholar] [CrossRef]
- Zapata, P.; Serrano, M.; Pretel, M.T.; Amoros, A.; Botella, M.A. Changes in Ethylene Evolution and Polyamine Profiles of Seedlings of Nine Cultivars of Lactuca Sativa L. in Response to Salt Stress during Germination. Plant Sci. 2003, 164, 557–563. [Google Scholar] [CrossRef]
- Silva, N.C.Q.; de Souza, G.A.; Pimenta, T.M.; Brito, F.A.L.; Picoli, E.A.T.; Zsögön, A.; Ribeiro, D.M. Salt Stress Inhibits Germination of Stylosanthes Humilis Seeds through Abscisic Acid Accumulation and Associated Changes in Ethylene Production. Plant Physiol. Biochem. 2018, 130, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Qian, R.; Ma, X.; Zhang, X.; Hu, Q.; Liu, H.; Zheng, J. Effect of Exogenous Spermidine on Osmotic Adjustment, Antioxidant Enzymes Activity, and Gene Expression of Gladiolus Gandavensis Seedlings Under Salt Stress. J. Plant Growth Regul. 2021, 40, 1353–1367. [Google Scholar] [CrossRef]
- Sequera-Mutiozabal, M.; Antoniou, C.; Tiburcio, A.F.; Alcázar, R.; Fotopoulos, V. Polyamines: Emerging Hubs Promoting Drought and Salt Stress Tolerance in Plants. Curr. Mol. Biol. Rep. 2017, 3, 28–36. [Google Scholar] [CrossRef]
- Verma, S.; Mishra, S.N. Putrescine Alleviation of Growth in Salt Stressed Brassica Juncea by Inducing Antioxidative Defense System. J. Plant. Physiol. 2005, 162, 669–677. [Google Scholar] [CrossRef]
- Pál, M.; Janda, T. Role of Polyamine Metabolism in Plant Pathogen Interactions. J. Plant Sci. Phytopathol. 2017, 1, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Wojtyla, Ł.; Lechowska, K.; Kubala, S.; Garnczarska, M. Different Modes of Hydrogen Peroxide Action During Seed Germination. Front. Plant. Sci. 2016, 7, 66. [Google Scholar] [CrossRef] [Green Version]
- Bailly, C. The Signalling Role of ROS in the Regulation of Seed Germination and Dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef] [PubMed]
- Ebeed, H.T.; Hassan, N.M.; Aljarani, A.M. Exogenous Applications of Polyamines Modulate Drought Responses in Wheat through Osmolytes Accumulation, Increasing Free Polyamine Levels and Regulation of Polyamine Biosynthetic Genes. Plant Physiol. Biochem. 2017, 118, 438–448. [Google Scholar] [CrossRef]
- Paul, S.; Roychoudhury, A.; Banerjee, A.; Chaudhuri, N.; Ghosh, P. Seed Pre-Treatment with Spermidine Alleviates Oxidative Damages to Different Extent in the Salt (NaCl)-Stressed Seedlings of Three Indica Rice Cultivars with Contrasting Level of Salt Tolerance; Elsevier: Amsterdam, The Netherlands, 2017; Volume 11. [Google Scholar] [CrossRef]
- Aziz, A.; Martin-Tanguy, J.; Larher, F. Stress-Induced Changes in Polyamine and Tyramine Levels Can Regulate Proline Accumulation in Tomato Leaf Discs Treated with Sodium Chloride. Physiol. Plant. 1998, 104, 195–202. [Google Scholar] [CrossRef]
- Hohlfeld, H.; Scheel, D.; Strack, D. Purification of Hydroxycinnamoyl-CoA: Tyramine Hydroxycinnamoyltransferase from Cell-Suspension Cultures of Solanum tuberosum L. Cv. Datura. Planta 1996, 199, 166–168. [Google Scholar] [CrossRef]
- Borg-Olivier, O.; Monties, B. Lignin, Suberin, Phenolic Acids and Tyramine in the Suberized, Wound-Induced Potato Periderm. Phytochemistry 1993, 32, 601–606. [Google Scholar] [CrossRef]
- Back, K.; Jang, S.M.; Lee, B.C.; Schmidt, A.; Strack, D.; Kim, K.M. Cloning and Characterization of a Hydroxycinnamoyl-CoA: Tyramine N-(Hydroxycinnamoyl)Transferase Induced in Response to UV-C and Wounding from Capsicum Annuum. Plant Cell Physiol. 2001, 42, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Zhong, M.; Shu, S.; Du, N.; Sun, J.; Guo, S. Proteomic and Physiological Analyses Reveal Putrescine Responses in Roots of Cucumber Stressed by NaCl. Front. Plant Sci. 2016, 7, 1035. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Shu, S.; An, Y.; Zhou, H.; Guo, S.; Sun, J. Influence of Exogenous Spermidine on Carbon–Nitrogen Metabolism under Ca(NO3)2 Stress in Cucumber Root. Plant Growth Reg. 2017, 81, 103–115. [Google Scholar] [CrossRef]
- Lefèvre, I.; Gratia, E.; Lutts, S. Discrimination between the Ionic and Osmotic Components of Salt Stress in Relation to Free Polyamine Level in Rice (Oryza sativa). Plant Sci. 2001, 161, 943–952. [Google Scholar] [CrossRef]
- Quinet, M.; Bataille, G.; Dobrev, P.I.; Capel, C.; Gómez, P.; Capel, J.; Lutts, S.; Motyka, V.; Angosto, T.; Lozano, R. Transcriptional and Hormonal Regulation of Petal and Stamen Development by STAMENLESS, the Tomato (Solanum lycopersicum L.) Orthologue to the B-Class APETALA3 Gene. J. Exp. Bot. 2014, 65, 2243–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cristescu, S.M.; De Martinis, D.; te Lintel Hekkert, S.; Parker, D.H.; Harren, F.J.M. Ethylene Production by Botrytis Cinerea In Vitro and in Tomatoes. Appl. Environ. Microbiol. 2002, 68, 5342–5350. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PAs Ratio | UPd | Pd | P7H2O | UP11H2O | P7NaCl | UP16NaCl |
|---|---|---|---|---|---|---|
| (Spd + Spm)/Put | 12.266 (a) | 9.215 (c) | 8.912 (c) | 11.676 (b) | 9.102 (c) | 7.218 (d) |
| Put/Spd | 0.098 (c) | 0.133 (b) | 0.137 (b) | 0.103 (c) | 0.137 (b) | 0.167 (a) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechowska, K.; Wojtyla, Ł.; Quinet, M.; Kubala, S.; Lutts, S.; Garnczarska, M. Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress. Int. J. Mol. Sci. 2022, 23, 349. https://doi.org/10.3390/ijms23010349
Lechowska K, Wojtyla Ł, Quinet M, Kubala S, Lutts S, Garnczarska M. Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress. International Journal of Molecular Sciences. 2022; 23(1):349. https://doi.org/10.3390/ijms23010349
Chicago/Turabian StyleLechowska, Katarzyna, Łukasz Wojtyla, Muriel Quinet, Szymon Kubala, Stanley Lutts, and Małgorzata Garnczarska. 2022. "Endogenous Polyamines and Ethylene Biosynthesis in Relation to Germination of Osmoprimed Brassica napus Seeds under Salt Stress" International Journal of Molecular Sciences 23, no. 1: 349. https://doi.org/10.3390/ijms23010349