
Native and Engineered Probiotics: Promising Agents against Related Systemic and Intestinal Diseases

Guangdong Provincial Key Laboratory of Protein Function and Regulation in Agricultural Organisms, College of Life Sciences, South China Agricultural University, Guangzhou 510642, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(2), 594; https://doi.org/10.3390/ijms23020594
Submission received: 9 December 2021
/
Revised: 29 December 2021
/
Accepted: 3 January 2022
/
Published: 6 January 2022
(This article belongs to the Special Issue The Effect of Probiotics on Gut Homeostasis Regulation 2.0)
Abstract
:Intestinal homeostasis is a dynamic balance involving the interaction between the host intestinal mucosa, immune barrier, intestinal microecology, nutrients, and metabolites. Once homeostasis is out of balance, it will increase the risk of intestinal diseases and is also closely associated with some systemic diseases. Probiotics (Escherichia coli Nissle 1917, Akkermansia muciniphila, Clostridium butyricum, lactic acid bacteria and Bifidobacterium spp.), maintaining the gut homeostasis through direct interaction with the intestine, can also exist as a specific agent to prevent, alleviate, or cure intestinal-related diseases. With genetic engineering technology advancing, probiotics can also show targeted therapeutic properties. The aims of this review are to summarize the roles of potential native and engineered probiotics in oncology, inflammatory bowel disease, and obesity, discussing the therapeutic applications of these probiotics.
1. Introduction
The benefits of probiotics are well known, and both traditional and recently discovered probiotics have received a lot of attention. They can also be quite successful therapeutic agents. There is a large number of reviews that focus on describing single probiotics [1,2,3], their derivatives [4,5], or probiotics in terms of health/disease [6,7]. The variety of probiotic bacteria and the complex relationships between them and various diseases cannot be summarized in a single review. We aim to enumerate and discuss the beneficial effects of probiotics (Escherichia coli Nissle 1917, Akkermansia muciniphila, Clostridium butyricum, lactic acid bacteria and Bifidobacterium spp.) and, once intestinal homeostasis is compromised, their impacts on cancer, inflammatory bowel disease (IBD), obesity, and other systemic disorders. Moreover, we will elaborate on the benefits of their modified bacteria (including their derivatives). In this review, comprehensive elucidation of these probiotics, both native and modified, is expected to have a significant impact on a deeper understanding of probiotics.
2. Intestinal Probiotics
Escherichia coli Nissle 1917 (EcN) was initially isolated from the feces of a German soldier who was clear of intestinal disease in a Shigella spp. heavily contaminated area [8]. Since its discovery, it has been commonly used to regulate intestinal microbiota [9,10], relieve inflammation [11,12], etc. Furthermore, EcN has good colonization [13,14], is nonimmunogenic, and has a clear genetic background [15], making it easily genetically engineered for therapeutic use.
Akkermansia muciniphila (AM), a strictly anaerobic, non-motile, non-spore-forming, intestinal mucin-degrading, Gram-negative bacteria, was isolated in 2004 by Derrien et al. [16]. Recently, the EFSA Panel on NDA concluded that the novel food (NF), pasteurized AM, is safe for the target population at 3.4 × 1010 cells/day, provided that the number of viable AM is below 10 cells/g NF [17]. AM is a resident probiotic with a high abundance in the mucus layer of the intestine [18,19]. It is thought to modulate host immune homeostasis, improve metabolism, protect the intestinal barrier [20], show anti-aging and anti-cancer effects [21], and play a role in the microbiota–brain–gut axis [22]. AM is currently being extensively researched, and expected to be the next generation of probiotics.
Clostridium butyricum (CB) is a strictly anaerobic, butyric acid-producing, Gram-positive bacteria, colonizing predominantly the distal small intestine and colon [23]. CB can be used as a probiotic preparation to promote host health, including regulation of intestinal microbiota [24], production of beneficial metabolites [25,26], prevention of intestinal inflammation [27,28], promotion of growth [29,30], etc. Another major advantage of the CB application is that it is a bacillus that forms heat-resistant endospores and survives in adverse conditions [31].
Over the past few decades, lactic acid bacteria and Bifidobacterium spp. have been used in dairy fermentation around the world [32,33]. Lactic acid bacteria is an aerotolerant anaerobic Gram-positive bacteria, which usually colonies the mucosal surface of the intestine [34] in the function of maintaining the balance of intestinal microbiota [35,36] and improving immune function [37,38]. Similarly, Bifidobacterium spp. is a Gram-positive bacterium, which has a variety of probiotic functions. It is able to adhere to the epithelial cells of the intestine [39] to secrete antibacterial substances [40] and synthesize vitamins [41]. In addition, they both have a hydrolytic effect on bile, which can cause oxidative stress and cellular damage [42]. Their tremendous worth in the fields of food and medicine has been demonstrated through many years of research.
The intestinal microbiota maintains the balance of the intestinal micro-ecology and links to almost all digestive diseases. Digestive disorders are the underlying basis for many systemic diseases, including endocrine diseases. Increased intestinal permeability and abnormal nutrient absorption contribute to these diseases, and further affect a range of metabolic changes. Probiotics, when used correctly, are a crucial part of maintaining human health.
3. Native Probiotics and Diseases
3.1. Native Probiotics and Cancer
Cancer is a major public health problem worldwide. Traditional cancer treatments include surgery, radiation therapy, and chemotherapy, which often lead to serious side effects as well as the destruction of adjacent normal cells [43]. Intestinal carcinogenesis is closely associated with alterations in intestinal native microbiota [44]. This may suggest that some probiotic bacteria can be utilized to assist in cancer prevention or therapy (Figure 1).
3.1.1. Native Escherichia coli Nissle 1917 (EcN) and Cancer
EcN can selectively colonize and replicate within solid tumors [45]. Except for tumor colonization, EcN can also trigger apoptosis in colon cancer HT-29 cells through a dose- and time-dependent mechanism [46]. In subcutaneous tumor-bearing mice, combined treatment of EcN and TGF-β blockers has been shown to be superior to monotherapy, and this synergy is probably mediated by the interaction of tumor-specific effector T cells, antigen-specific IFN-γ+ CD8+ T cells, and the gut microbiota [47]. However, 1917 can also produce colibactin in vitro and in vivo, inducing DNA damage, which may contribute to the development of colorectal cancer [48].
3.1.2. Native Akkermansia muciniphila (AM) and Cancer
AM is closely associated with cancer. It is found to be more abundant in non-small cell lung cancer patients than normal healthy people [49], and the abundance of AM gradually decreases during the progression of cirrhosis only to hepatocellular carcinoma-cirrhosis [50]. AM in combination with current anti-tumor therapies, including cisplatin treatment and IL-2-based immunotherapy, is also a promising treatment strategy [51,52]. In addition, the drug treatments of castration-resistant prostate cancer [53], pancreatic ductal adenocarcinoma [54], lung cancer [55], and colorectal cancer [56,57] may also be driven by AM.
AM has some specific proteins that may impact the progression of tumors. Among these, the Amuc_1434* degrades mucin 2 in colon cancer cells and promotes apoptosis of human colon cancer LS174T cells [58,59], and its specific outer membrane protein Amuc_1100 can inhibit colitis-associated colorectal cancer by regulating CD8+ cytotoxic T lymphocytes [60].
AM also plays an essential role in immunization. AM has been reported to suppress colorectal tumorigenesis and prostate cancer by inducing tumor-associated macrophages [61,62]. Notably, AM may contribute to immune checkpoint blockade (ICB). AM is a responder during anti-PD-1 immunotherapy and has an impact on effectiveness [63,64,65], perhaps due to the alterations in glycerophospholipid metabolism [66] and AM-derived cdAMP [67].
3.1.3. Native Clostridium butyricum (CB) and Cancer
CB reduces colitis-related colon cancer [68] and intestinal tumor [69] in mice, which may be related to the regulation of the NF-κB, Wnt signaling pathway. In addition, CB and its related metabolites also play a role in regulating the gut microbiota and promoting cancer therapy. CB achieved 18% eradication rates of Helicobacter pylori infection (associated with gastric cancer) among outpatients in a pilot study [70]. The cell-free supernatants derived from CB possess an antibiofilm effect and inhibit the growth of Enterotoxigenic Bacteroides fragilis [71], which has a risk of causing colorectal cancer. Since CB belongs to bacillus, the orally spores encapsulated by prebiotics have also been reported to specifically enrich and suppress colon cancer, regulating the intestinal microbiota [72]. It is notable that CB also influences microRNAs; it promotes the upregulation of tumor suppressor mir-200c, leading to the regulation of colitis-induced oncogenesis [73].
CB can also be used as an adjunct to current therapies. The research of Tian et al. has shown that CB reduces adverse events, especially diarrhea, caused by chemotherapy for lung cancer [74]. Furthermore, CB exhibits a synergistic effect when combined with lapatinib treatment for colon tumors in mice [75]. In ICB therapy, CB therapy can enhance the efficacy even in antibiotic-treated patients and is associated with longer progression-free survival and overall survival [76].
3.1.4. Native Bifidobacterium spp., Lactic Acid Bacteria and Cancer
Bifidobacterium spp. and lactic acid bacteria have been widely used in food, healthcare and medical applications. As an important component of intestinal probiotics, they also have an important contribution to anti-cancer. Chou et al. showed that dietary Limosilactobacillus fermentum appears to modulate the intestinal microbiota and reduce inflammation, which may well be helpful in mitigating the development of colon cancer [77], and the exopolysaccharides of lactic acid bacteria have the capability to induce apoptosis in cancer cells [78]. In addition, solid fermented grains of Limosilactobacillus reuteri and Lactiplantibacillus plantarum subsp. plantarum are also anti-proliferative against cancer cells in vitro, which may be related to released bioactive peptides and degraded polysaccharides [79].
A decrease in lactic acid bacteria and Bifidobacterium spp. was observed in patients with colorectal cancer [80]. In addition, enteral administration of Bifidobacterium breve can usually maintain the total fecal organic acids of patients with malignancies on chemotherapy above 100 μmol/g, with a pH below 7.0, and reduce the frequency of fever and antibiotic use (compared to the placebo group) [81]. This all suggests that we can intervene in the development of cancer through probiotics. Currently, lactic acid bacteria have been used as a vector to successfully transport therapeutic agents into MCF-7 cancer cells in vitro [82]. However, the safety and efficacy of this drug vector in vivo have yet to be explored. On the other hand, encouragingly, the significant independent antitumor effects and adjuvant effect on anti-PD-L1 of live Bifidobacterium spp. have been demonstrated in mice melanomas, which may be related to an anti-tumor T-cell response [83]. This bodes well for the use of Bifidobacterium spp. in human immunotherapies.
3.2. Native Probiotics and Inflammatory Bowel Disease (IBD)
IBD is a chronic recurrent inflammation of the gastrointestinal tract, mainly including Crohn’s disease and ulcerative colitis (UC) [84]. At present, more and more novel and emerging therapies towards IBD are put into research and development [85]. A general consensus has been established that gut microbiota play pivotal roles in the development of IBD [86]. Probiotics and their produced metabolites are crucial for gut homeostasis, suggesting they may have a role in the treatment of IBD [87,88] (Figure 1).
3.2.1. Native Escherichia coli Nissle 1917 (EcN) and IBD
EcN has been assessed in clinical trials and has shown a remarkable therapeutic effect, especially in UC treatment [89]. As early as 2004, EcN was shown to have the same effects as regular mesalazine but required a considerably lower dosage to reduce UC symptoms. [90]. EcN was also reported to protect intestinal epithelial cells by inducing human beta-defensin-2, which showed a higher expression level in UC than in CD, through NF-κB and activator protein 1 [91]. Meanwhile, EcN is a gut-friendly microorganism. Specific pathogen-free, but not germ-free, IL-2−/− mice generally develop colitis. Strikingly, GF IL-2−/− mice with EcN colonized did not induce colitis [92]. This is consistent with EcN being more effective in treating UC in the clinic. Remarkably, EcN was also shown to affect the regulation of micro-RNAs involved in the inflammatory response (miR-143, miR-150, miR-155, miR-223, and miR-375). For example, EcN could reverse the downregulation of miR-150, which reduced the expression of c-Myb and thereby compromised the integrity of the intestinal barrier [93,94]. It could also suppress the up-regulation of miR-155 and miR-233, which is consistent with the activities of IL-1β expression and the Th1/Th17 response, respectively [95].
EcN can also assist in the performance of other drugs. Common drugs for the treatment of IBD, such as 5-ASA and mesalamine, may affect the degradation rates of the polysaccharide-based drug delivery system. Fortunately, the utilization rate of oral drugs has been shown to be improved under the assistance of probiotics including EcN [96]. Additionally, EcN outer membrane vesicles (OMVs) have also been proved to have intestinal barrier protective and anti-inflammatory effects towards IBD, and it could be a promising treatment because of its non-replication and safety [91,97].
3.2.2. Native Akkermansia muciniphila (AM) and IBD
As an anaerobic bacteria specialized in the utilization of mucin as a carbon and nitrogen source, AM became a key force to protect the integrity of the epithelial barrier and update the mucus layer, which plays a key role in the onset of IBD [98,99,100]. The abundance of AM can be markedly enriched, along with restoring the epithelium barriers in acute colitis, by hyaluronic acid-bilirubin nanomedicine [101]. Additionally, AM could firmly bind to the laminin and cultured colonic epithelial cell lines Caco-2 and HT-29, thus gaining competitive exclusion of pathogens [102]. Plenty of investigations on IBD patients have identified that the abundance of AM is critically influenced. Through obtaining colonic biopsies and mucus brushings from UC patients, Earley et al. found that lower abundances of AM had a strong association with a lower percentage of sulphated mucin and higher inflammation [103]. Furthermore, 53.7% of IBD patients who underwent washed microbiota transplantation had a clinical response while also showing a higher colonization rate of AM [104]. Strikingly, different strains of AM could perform special functions on immune modulation. Both strain 139 and strain ATCC BAA-835 were sufficient to downregulate the expression of the pro-inflammatory cytokines (TNF-α and IFN-γ), while ATCC BAA-835 could modulate the differentiation of regulatory T (Treg) cells and increase the production of short-chain fatty acids, but strain 139 could not [105]. This suggests that more research needs to be done to better apply AM in IBD treatment.
3.2.3. Native Clostridium butyricum (CB) and IBD
Generally, microbial dysbiosis in IBD patients is associated with a decrease in short-chain fatty acid (SCFA)-producing bacteria [106,107]. CB can consume undigested dietary fiber and generate SCFAs, especially butyrate and acetate, and is listed as a potential probiotic for treating IBD [1]. Accordingly, the derivatives of CB have been shown to effectively prevent bloody diarrhea, therefore treating dextran sulfate sodium (DSS)-induced experimental colitis in rats [108]. It has also been reported that antibiotics used early in life play an important role in gut microbiota disorders, and can lead to IBD, while supplementation of antibiotics with butyrate can restore the intestinal macrophage response and prevent T cell dysfunction [109]. Compared with mesalamine and sodium butyrate, CB CGMCC0313 performed better on lowering the serum levels of both IL-23 and TNF-α as well as appearing to restore the intestinal microbiota more quickly [110]. CB CBM588 was shown to directly trigger the TLR2/MyD88 pathway of intestinal macrophages in the mucosa, resulting in the generation of IL-10, which can suppress colitis by regulating Treg cells [111,112]. In addition, CB can also be combined with other prebiotics or immunotherapy to assist in the treatment of IBD. For instance, CB capsules combined with specific immunotherapy can significantly inhibit UC and down-regulate the expression of proinflammatory cytokines in the clinics [113]. CB also employs germinated barley foodstuff, which exerts a therapeutic effect on IBD as a fermentation substrate. Combining them, according to this, CB can effectively treat colitis [114].
3.2.4. Native Bifidobacterium spp., Lactic acid Bacteria and IBD
Bifidobacterium spp. and Lactic acid bacteria are probiotics that have been widely applied in the food and medicine industry [115]. Plenty of plant extracts in the treatment of IBD showed that the reduction of colitis symptoms in mice was accompanied by a significant increase in Bifidobacterium spp. and Lactic acid bacteria, such as purple sweet potato anthocyanin extract, glycerol monolaurate, polysaccharides from tea flowers, etc. [116,117,118]. The underlying mechanism of action is closely related to the activation of Toll-like receptors and their downstream signaling pathways [119]. On the one hand, it has been shown that the TLR4/MYD88/NF- κB signal pathway can be inhibited by Bifidobacterium animalis subsp. lactis XLTG11, causing downregulation of pro-inflammatory cytokines to alleviate DSS-induced colitis in mice [120]. On the other hand, the mice can also be protected by Bifidobacterium adolescentis IF1-03, which activated macrophages through the TLR2/ERK/p38 MAPK signal pathway and induced differential regulation of the inflammatory immune response in the Treg/Th17 axis [121]. Mediated by the same signal pathway, it has been found that strain Bifidobacterium bifidum 1 could also induce significant and continuous enhancement of the intestinal tight junction (TJ) barrier in Caco-2 monolayers [122]. In IBD mediated by pathogenic bacteria, they have been proven to regulate the tight junction of Caco-2 cells [123], counteract Adherent-Invasive Escherichia coli virulence in CD patients, and reduce the secretion of cytokines related to the IL-23/Th17 axis in UC patients [124]. Furthermore, secreted factors from Bifidobacterium dentium can also down-regulate the endoplasmic reticulum stress gene and regulate the unfolded protein response to promote MUC2 secretion [125].
3.3. Native Probiotics and Obesity with Associated Diseases
Obesity is prone to a number of complications and is particularly associated with the development of diabetes [126]. The main reason for the development of obesity is a high-fat diet (HFD). HFD can imbalance the gut microbiota, reducing some probiotic bacteria such as Bifidobacterium, Lactobacillus and Akkermansia [127]. Moreover, it damages the intestinal barrier and even leads to a leaky gut, resulting in a range of metabolic diseases [128]. On the other hand, consumption of probiotics can improve the integrity of the mucosal barrier [129]. Furthermore, the intestinal microbiota has a regulatory role in obesity, with probiotics having a potentially beneficial effect on various metabolic parameters [130,131] (Figure 1).
3.3.1. Native Escherichia coli Nissle 1917 (EcN) and Obesity with Associated Diseases
To our knowledge, there are no reports related to the direct effects of native EcN on obesity and diabetes. However, EcN increases the expression of tight junction proteins to stabilize the intestinal barrier, which may be related to its secretion of OMVs and soluble factors [132,133]. One of the mechanisms is through the inhibition of TNF-α and IFN-γ-induced activation of NF-κB p65, thereby decreasing the binding of NF-κB in the myosin light-chain kinase promoter to reduce the disruption of tight junctions [134]. In addition, during this process, some miRNAs (miR-203, miR-483-3p, miR-595) may also contribute to tight junctions [135]. Subsequently, EcN was shown to have a protective effect against enteropathogenic E. coli-induced intestinal epithelial barrier dysfunction [136] and 5-fluorouracil-induced epithelial cell damage [137]. Considering the protective effect of EcN against pathogen-induced and chemical-induced intestinal barrier damage, it may also mitigate the disruption of the intestinal barrier caused by HFD to hinder the development of other metabolic diseases.
3.3.2. Native Akkermansia muciniphila (AM) and Obesity with Associated Diseases
It is now generally accepted that the abundance of AM is negatively correlated with obesity [138,139,140]. Dietary habits, one of the main causes of obesity, can also change the abundance of AM [141]. Dietary intervention in mice on an HFD can increase the abundance of AM [142]. Interestingly, a large amount of research has shown that while using some substances (including grape polyphenols [143], epigallocatechin-3-gallate [144], rhubarb extract [145], polysaccharides obtained from Cordyceps militaris [146], protein-bound β-glucan [147], puerarin [148], Bofutsushosan [149,150], etc.) to combat obesity, it has also been found that they can regulate the abundance of AM to improve efficacy. On the other hand, low AM abundance is associated with type 2 diabetes [139,141] and leads to impaired insulin secretion and disturbed glucose homeostasis even in lean individuals [151]. Therefore, it is natural that research on AM is focusing on the fight against obesity and its related diseases, especially diabetes.
The anti-obesity properties of AM have been almost comprehensively evaluated. It improves vital obesity parameters and insulin sensitivity, and rescues glucose homeostasis in HFD mice [152]. Moreover, AM treatment can increase intestinal endogenous cannabinoid levels to further maintain intestinal homeostasis [139]. Clinical studies have also shown that the higher the AM abundance, the healthier the insulin sensitivity in individuals with calorie-restricted weight loss [153]. Meanwhile, AM-metabolites have been reported to affect various transcription factors and to be involved in peripheral lipid metabolism [154]. In addition to systemic obesity, AM has shown excellent performance in improving fatty liver and liver function through the enterohepatic axis, possibly in part by increasing L-aspartate levels from the intestine to the liver and regulating genes involved in lipid synthesis [152,155,156]. AM also makes a significant contribution to the intestinal barrier. There is evidence that AM helps to repair the intestinal barrier [152], and contrarily AM deficiency damages the intestinal barrier integrity [157]. This maintenance of the intestinal barrier may be related to the inflammation-relieving and anti-cancer properties of its membrane protein, Amuc_1100 [60]. Interacting with Toll-like receptor 2, Amuc_1100 also improves metabolism in obese and diabetic mice [158]. Combined with the reducing perilipin 2 (associated with lipid storage) in brown and white adipose tissues as well as increased energy expenditure and excretion by pasteurized AM [159], we can conclude that the beneficial effects of pasteurized AM on obesity and diabetes still exist.
3.3.3. Native Clostridium butyricum (CB) and Obesity with Associated Diseases
The tendency to accumulate body fat is negatively correlated with the abundance of CB and an HFD will lead to a reduction in the amount of CB [160]. In turn, CB administration reduced adipose tissue mass, blood glucose, and serum triglycerides [161,162,163], increased peroxisome abundance, and modulated related genes to improve lipid metabolism [161]. Additionally, CB can also mitigate leaky gut and reinforce the intestinal barrier by upregulating TJ proteins (claudin-1 and occludin) [162]. The cell wall components and secretory products of CB (butyrate) have also been observed to stimulate ANGPTL4 production, which affects the triglyceride deposition in adipocytes [164].
On the other hand, CB has a beneficial effect on the postoperative period of bariatric-related surgery. A comparison of the modified Gastrointestinal Quality of Life Index before and after the intervention shows that CB improves gastrointestinal symptoms after gastric bypass surgery, which may be caused by a combination of altered digestive environment and microbiota [165]. In addition, CB has an interventional effect on bone metabolism. Studies have shown that administration of CB can promote osteoblast autophagy to reduce bone loss after bariatric surgery, possibly also by altering the microbial composition of the gut [166].
Moreover, CB improves glucose homeostasis and insulin resistance in type 2 diabetic mice [167]. In autoimmune diabetes, CB also has a protective effect, which involves the regulation of intestinal immunity, particularly pancreatic regulatory T-cells and intestinal microbiota [168]. Through the brain–gut axis, CB was shown to attenuate cerebral I/R injury with diabetes and to reverse neuronal apoptosis [169]. In conclusion, CB has a promising application in improving obesity, diabetes, and other diseases, as well as helping patients to recover after surgery.
3.3.4. Native Bifidobacterium spp., Lactic Acid Bacteria and Obesity with Associated Diseases
Significant differences in microbial composition were found in the childhood obesity survey, with a significant decrease in the abundance of Bifidobacterium [170]. Unsurprisingly, a relatively high abundance of Lacticaseibacillus paracasei subsp. paracasei has an effect against obesity in children [171]. Adjusting the abundance of probiotics can help prevent obesity in the early stages. Bifidobacterium animalis subsp. lactis CECT 8145 treatment inhibited intake and significantly increased plasma adiponectin levels in obese mice, which may play a key role in the regulation of insulin sensitivity and glucose metabolism [172]. Bifidobacterium animalis subsp. lactis A6 reduces body weight and fat mass in obese mice, even showing better results than AM [173]. It increases the expression of endothelial nitric oxide synthase which alters mitochondrial metabolism and reduces the risk of obesity [173]. Intervention with Lactiplantibacillus plantarum subsp. plantarum reduces the rate of weight gain in mice through the peroxisome proliferators-activated receptor pathway, improves related parameters, attenuates obesity-related oxidative damage and inflammatory responses [174]. Similarly, Bifidobacterium longum [175], Bifidobacterium pseudolongum [176], Lactiplantibacillus plantarum subsp. plantarum [177], Lacticaseibacillus rhamnosus [178], Companilactobacillus crustorum [178], Latilactobacillus sakei subsp. sakei [179,180], and Lactobacillus johnsonii [181] all induce weight loss, shown by reduced fat accumulation in HFD-induced obese mice. Notably, via the gut–bone axis, Bifidobacterium pseudocatenulatum mitigates the adverse effects of obesity on bone metabolism; one of the reasons for this may be through activation of the Wnt/β-catenin pathway [182].
In addition, Bifidobacterium longum contributes to glucose tolerance, regulates lipid metabolism, protects the liver and pancreas, and improves glucagon-like peptide-1 (GLP-1) [183]. After inactivation, it still has the potential to improve obesity and diabetes-related parameters [184]. Lacticaseibacillus paracasei subsp. paracasei was also reported to have an ameliorative effect on rats with type 2 diabetes mellitus, even with parameters comparable to those of normal mice after the intervention [185]. In clinical studies, Limosilactobacillus reuteri has been reported to improve insulin sensitivity in some patients [186]. It has good therapeutic effects in patients with type 2 diabetes and can increase the abundance of Bifidobacterium [187]. Lactobacillus acidophilus improves the expression of genes related to glucose and lipid metabolism and regulates the gut microbiota to combat diabetes [188]. Meanwhile, it also enhances the intestinal barrier function [188]. Likewise, Bifidobacterium has a modulating function on intestinal permeability and has a promising application in intestinal leakage, but with attention to the antagonistic effects when combined with prebiotics [189].
The combination of Lacticaseibacillus rhamnosus and Bifidobacterium lactis showed a protective effect on islet B cell function in children with newly diagnosed type 1 diabetes [190]. The culture supernatant of Bifidobacterium longum and Lacticaseibacillus rhamnosus promotes the “browning” of adipose tissue, altering energy metabolism and activating thermogenesis through PKA/CREB to treat obesity [191]. The mechanism of the combination of Bifidobacterium spp. and Lactic acid bacteria in the treatment of obesity still remains to be explored and the appropriate form of microbiota supplementation needs to be screened.
3.4. Summary
Currently, the anti-tumor effects of probiotics have mainly been reported in digestive system cancer and lung cancer. Probiotic therapies used in oncology are mainly investigated in cells and mice. This cancer treatment is mostly used as an adjuvant, with mechanisms of probiotics that modulate the composition of the gut microbia and promote immunological responses. Although probiotic therapy is still in its infancy, it can be added to the daily diet for cancer prevention. In addition, probiotics can also be used in combination with drugs and ICB in clinical treatment.
The research on probiotic therapy in IBD treatment is mainly carried out on clinical patients or experimental mouse models, especially colitis. The main responsibilities of probiotics are intestinal microbiota regulation, intestinal epithelial barrier protection, and abnormal inflammatory response inhibition. Existing probiotic therapy can be combined with prebiotics or immunotherapy to achieve a significant therapeutic effect; some of them even catch up with the general clinical drugs.
Obesity is closely linked to gut microbes. One of the mechanisms of using probiotics to treat obesity lies in the regulation of the intestinal microbiota composition. The microbiota affects glucose and lipid metabolism, obesity-related biochemical parameters, inflammation, and oxidative stress levels, insulin sensitivity, and the intestinal barrier through various signaling pathways. When the impacts are negative, a series of metabolic disturbances and pathological changes can lead to the development of obesity. The utilization of probiotics as an appropriate supplement is expected to reverse this process, counteracting the development of obesity and its complications.
Native probiotics have a wide range of applications and are beneficial to health. They show a preventive and palliative effect on many diseases.
4. Modified Probiotics and Diseases
It is possible to modify the properties of probiotics in a targeted way, even transforming its auxiliary role in disease treatment into a mainstay. Purposefully engineered probiotics can be more multi-functional and efficient.
4.1. Modified Escherichia coli Nissle 1917, EcN
The complete genomic DNA sequence has given a strong impetus to the engineering methods of EcN [15]. As an engineered bacterium, EcN is expected to achieve better pharmacological properties. In recent years, the engineered EcN has been studied in a variety of fields (Figure 2) (Table 1).
4.1.1. Express Direct Therapeutic Factors
One of the key strategies for using the engineered EcN in disease therapy is to modify it to directly express therapeutic factors. The administration of engineered HlyE-expressing EcN in cytolytic therapy of tumors promotes regression of tumor tissues in mice [192]. In this research, EcN was equipped with a strictly regulated expression system. This process includes extinguishing the araFGH operon and araBAD operon, and knocking down ptsG to ensure persistent inducibility of L-arabinose, which induces PBAD, thereby expressing HlyE. In addition, engineered EcN expressing soluble Tum-5 (a fragment of tumstatin) [193] or azurin, a copper-containing redox protein that can initiate cancer cell apoptosis [194], effectively inhibits tumor growth in xenograft mice tumor cells. Similarly, the tumor-targeting characteristics allow the introduction of a cluster of biosynthetic genes for cytotoxic compounds into EcN for tumor therapy [45,195].
In terms of intestinal inflammation, EcN is used in the expression of schistosome immunoregulatory proteins to continuously protect the body against colitis [196]. In this study, the sequence encoding Sj16 peptide was linked to the HlyA C-terminal signaling sequence and contained in an expression vector alongside the HlyB and HlyD. Using HlyB, HlyD, and TolC proteins to form the pore recognizing HlyA protein signal, Sj16 was secreted directly into the extracellular environment, rescuing gut microbial compositions and effectively alleviating DSS-induced colitis in mice. The heterologous synthesis of (R)-3-hydroxybutyrate (3HB) by altering the EcN metabolic pathway has also been studied [197]. This process involved screening for acetyl-CoA acetyltransferase, 3HB-CoA dehydrogenase, and thioesterase isozymes, inserting the optimum 3HB synthesis system into the EcN genome, and knocking out adhE or ldhA to inhibit the branch pathways and increase 3HB production, thereby alleviating colitis. Optogenetics has also been combined with probiotics, using upconversion microgels to convert near-infrared light into local visible blue light [198]. Through this technology, the recombinant EcN is activated to secrete autotransporter 43 adhesins antigen, achieving precise spatiotemporal regulation, which effectively alleviates colitis in mice [198]. Recently, recombinant IL-10-secreting EcN has been used in the optotheranostic nanosystem for UC as a real-time intervention module for long-lasting relief of the intestinal inflammatory responses [199], making it possible to diagnose and treat intestinal inflammation at home.
In particular, to combat obesity and prevent related chronic diseases, EcN has been modified to secrete dipeptidyl peptidase 4-degradation-resistant GLP-1 [200] or N-acyl-phosphatidylethanolamines. Both possess anti-obesity properties [201], which have numerous benefits for obese mice. These studies offer an oral treatment option for obesity and even chronic diseases.
The engineered EcN also has great potential in terms of antimicrobial activity. EcN was selected to deliver anti-enterococcal peptides, which were expressed as a fusion of Microcin V secretion tag and a mature bacteriocin [202]. These probiotics were subsequently demonstrated to reduce the levels of Enterococcus faecium and Enterococcus faecalis in the feces of male Balb/cJ mice. During intestinal infection with Pseudomonas aeruginosa, the EcN containing the ‘Sense-Kill’ construct could express dispersin B, which destabilizes biofilms against P. aeruginosa infection [203].
The engineered EcN has also been applied in cardiometabolic diseases. Producing N-phosphatidylethanolamines, endogenous anorexigenic lipids, EcN was constructed, which has been shown to reduce body weight, liver inflammation, fibrosis, and atherosclerotic necrosis, thus alleviating cardiovascular metabolic disease in low-density lipoprotein receptor null mice [204].
In metabolic diseases, EcN can still play a role. The modified probiotic EcN, producing pyrroloquinoline quinone (PQQ) and other metabolizing enzymes, is reported to relieve heavy metal toxicity [205], iron deficiency [206], fructose-induced dyslipidemia, hyperglycemia, and hepatic steatosis [206,207]. Moreover, to alleviate hyperammonemia, Kurtz et al. deleted the negative regulator of L-arginine biosynthesis and inserted a feedback-resistant L-arginine biosynthetic enzyme that exhausts ammonia for L-arginine biosynthesis in EcN, which yielded satisfactory results in a Phase I clinical study [208]. Modification of EcN is also an alternative strategy to fight against phenylketonuria (PKU). The genes encoding phenylalanine ammonia-lyase and L-amino acid deaminase are inserted into the genome to facilitate EcN consumption of phenylalanine in patients with PKU [209]. These findings all demonstrated the potential of EcN to be used in the treatment of metabolic disorders.
4.1.2. Express Adjuvant Therapeutic Factors
Another alternative strategy is to express adjuvant therapeutic factors. Engineered EcN is also used in combination therapy with photodynamic therapy. It was tailored by metabolic engineering to promote 5-aminolevulinic acid (5-ALA) production, which can be converted to protoporphyrin IX (the photosensitizer for photodynamic therapy) [210]. In particular, Salmonella arizona hemAM and endogenous hemL were co-expressed in EcN to increase 5-ALA accumulation. Meanwhile, levulinic acid was used to inhibit downstream biosynthesis for 5-ALA production. Moreover, to improve the efficacy of radiotherapy, a continuous catalase (CAT)-secreting EcN has been constructed, using CAT to catalyze the production of O2 from H2O2 in tumor cells, alleviating the hypoxic environment and increasing the sensitivity of tumors to radiotherapy [211].
Engineered EcN also serves for ICB treatment. Considering the enhancement on T-cell anti-tumor activity of L-arginine in tumors [226], similarly to Kurtz et al. [208], EcN has been engineered to maintain a high local level of L-arginine in tumors, which was achieved by deleting the arginine repressor gene (ArgR) and integrating feedback-resistant mutant N-acetyl glutamate synthase ArgAfbr [212]. Therefore, PD-L1 blockade therapy can be coordinated with this probiotic, which utilizes ammonia, metabolic waste in tumors, to produce L-arginine.
4.1.3. EcN or EcN-derivatives as a Targeted Delivery System of Therapeutic Factors
The engineered EcN can also be employed as a delivery system of therapeutic factors. Xie et al. used acid-labile materials that are hydrolyzed responsively to the acidic tumor microenvironment to conjugate therapeutic promicelle polymers on EcN [215,216]. The polymers are released and self-assembled into hybrid micelles that are endocytosed into tumor cells in response to cytosolic GSH and release synergetic anti-tumor drugs [215], achieving a tumor-targeted release of drugs in vivo. Considering that this drug loading is friendly to bacterial motion profiles and activity and that the drug accumulation is higher than that of commonly used nanocarriers [216], this technology offers an effective strategy for targeted delivery.
Interestingly, Joshi et al. constructed an EcN that secretes a curli-fused healing bio-signature, where these curli fibers bind firmly to the inflammation site of the intestinal mucus layer like a band-aid [217], and they systematically designed the pMUT plasmid containing the curli secretion system to export curli proteins with specific therapeutic factors to the intestine in vivo [218]. This study could be a boon for patients with IBD and even intestinal trauma.
As a new adjuvant and antigen delivery platform, the bacterial ghosts (BGs) based on EcN to treat cancer have also been studied in recent years. Kraśko et al. showed that a course of three subcutaneous inoculations with Lewis lung carcinoma (LLC) lysate supplemented with EcN-BGs significantly increased the survival rate of mice after removal of LLC tumors [219]. Effective drug delivery is also a major challenge in drug therapy, and thus cell-targeted drug delivery vehicles are receiving a lot of attention in oncology drug therapy. EcN is reported to effectively load Epothilones and induce apoptosis in HeLa cells [220]. Recently, Xie et al. used the photothermal effect of EcN-BGs surface-modified nanorods to modulate the temporal and spatial release of contained drugs [221], which suggests that it is a promising candidate for drug carrier. Except for cancer, EcN-derived BGs expressing chlamydial antigens have been constructed, providing a new insight into the treatment of ocular surface diseases [222].
Based on the OMVs lacking infectivity while retaining pathogen-associated molecular pattern molecules and the specific regulation of T cells via TLR2 [223,227], the display of exogenous antigens on the surface of EcN OMVs using a ClyA fusion chimera to trigger cellular immune responses has also proven to be a great strategy for the development of recombinant subunit vaccines [223].
EcN may be used in the development of live vaccines. Porcine postweaning diarrhea is closely associated with F4 and F18 fimbriae enterotoxigenic Escherichia coli (ETEC), and studies have shown that EcN integrated with F4 or F18 fimbriae cluster genes expressed in the surface can activate adhesion inhibition of F4+ and F18+ ETEC by piglets [224]. This evidence suggests that EcN is a promising candidate for a live vaccine.
Interestingly, in addition to regular-sized forms, the nanosized forms of EcN have been studied in the production of tumor-targeting delivery. By deleting the minCD gene and enhancing the expression of the minE gene, EcN-derived minicells were produced on the scale [225]. Compared with traditional nanoparticles, these minicells have higher biocompatibility, lower immunogenicity and toxicity, greater drug loading and therapeutic index [228], providing an innovative means of drug delivery. The EcN-derived minicells harboring a low-pH insertion peptide (to increase molecular accumulation in the acidic tumor microenvironment) and doxorubicin (antibiotic antineoplastics) show a significant tumor regression ability in a mouse model [225].
4.2. Modified Akkermansia muciniphila
AM was isolated in 2004 [16], and in 2011 its genome was sequenced and annotated by Smidt et al. [229], laying the foundation for genetic modification of AM. However, to our knowledge, there are no reports of AM genetic modification. Apart from the probiotic effect of AM, studies have focused on its OMVs (Table 2). AM-OMVs can influence obesity-related genes, as well as metabolism, to treat obesity, improve the tight junction to control leaky gut [230,231,232], and decrease the level of pro-inflammatory cytokines to ameliorate colitis [233], maintaining intestinal homeostasis. Subsequently, AM-OMVs may alleviate pathological damage to the liver and prevent liver fibrosis via the hepatic portal vein [234,235]. Not limited to the intestine axis, though the gut–brain axis, AM-derived OMVs can also affect the expression of key genes in the serotonin system and promote serotonin concentrations [236,237]. In addition, AM-OMVs exhibit significant osteogenic effects and inhibit osteoclast differentiation, conveying the potential to regulate bone homeostasis [238]. Moreover, AM-OMVs elevate the proportion of GZMB+ and IFN-γ+ lymphocytes in CD8+ T cells and recruit macrophages, which represent their antitumor potential [62].
Based on their powerful and broad therapeutic properties, AM-OMVs may be designed for drug delivery in the future. For example, through engineering, AM-OMVs can act on the surface as immunotherapeutic agents. In fact, it is not only OMVs; given the probiotic effect of AM (Section 3.1.2, Section 3.2.2 and Section 3.3.2), its ability to colonize, and the clear genetic background, AM could be genetically modified using λ-Red recombination system/CRISPR-Cas system/nisin-controlled gene expression system to display or secrete the therapeutic factors for application in more fields to treat diseases.
4.3. Modified Clostridium Butyricum
Currently, CB is hardly ever engineered for health maintenance and disease prevention (Table 2), while it is mainly applied in biosynthesis [262,263,264]. Most of these studies involve modifying metabolic pathways to optimize production through gene overexpression or inactivation. From a health perspective, our laboratory constructed a porcine epidermal growth factor overexpressing CB to enhance its gut protective function [240]. However, it should be mentioned that the major hindrance to the application of engineered CB in health care is the paucity of viable genetic tools for CB, or in other words, genetic manipulation of CB is prone to failure.
In general, for gene editing in CB, a plasmid system for shuttling and replication in E. coli and CB is first required. A number of plasmids have been successfully applied to Clostridium spp. including plasmids such as pMTL007 [263,265] and pMTL82151 [240,266]. In terms of gene editing methods, Heap et al. developed the ClosTron system in 2007 [267]. The principle of this technique is to use type II intron RNA to self-splice into lariat RNAs that complementarily pair with the target DNA sequence, thus achieving inactivation of the target gene [267]. Subsequently, this team developed another effective Clostridium spp. gene-editing technique, the allele-coupled exchange, which achieves gene knockout by bi-directional screening based on the pyrE gene [268]. Recently, efficient gene editing tools developed in CB using the heterologous Type II CRISPR-Cas9 system and endogenous Type I-B CRISPR-Cas system have also been reported [269]. This easily applied technology reduces the occurrence of the polarity effect in genetic manipulation for CB and lays the foundation for CB-based therapies.
In addition, CB-OMVs are highly productive and can stimulate the innate immune system [239], which suggests that CB-OMVs also have the potential to be used as vectors to deliver antigens or carry therapeutic factors by genetic manipulation.
4.4. Modified Lactic Acid Bacteria and Bifidobacterium spp.
4.4.1. Treatment of Diseases
As traditional probiotics engineered Lactic acid bacteria and Bifidobacterium spp. have also been used in recent years (Table 2). Bifidobacterium longum was reported to be modified to express a single-chain variable fragment that bound to the receptor on the HER2-positive cancer cell surface [270] or Tum [271], thus inhibiting the growth and proliferation of cancer cells.
Besides cancer, some studies also use Bifidobacterium longum to express antioxidant enzymes [241], anti-inflammatory cytokines [243], and hormones [242] against inflammatory diseases. Engineered Lacticaseibacillus paracasei subsp. paracasei F19 can provide the enzyme N-acyl-phosphatidylethanolamine-preferring phospholipase D, which indirectly produces a natural lipid, palmitoylethanolamide, playing a role in regulating inflammation [250]. Moreover, through modulating the intestinal microbiota and reducing bacterial translocation to the liver, IL-22-expressing Limosilactobacillus reuteri has been reported to alleviate ethanol-induced steatohepatitis [247] and reduce nonalcoholic fatty liver diseases [246]. These constructed probiotics exert a powerful anti-inflammatory capacity, especially shown in the hepatic and intestinal axis.
Additionally, metabolic disorders are closely associated with the development of inflammation [272,273]. To alleviate metabolic disorders, Bifidobacterium longum has also been genetically engineered to express hormones originally released from the intestine (for example, oxyntomodulin and GLP-1) or its derivatives to facilitate the administration of drugs [244,245]. These engineered probiotics improve the absorption of nutrients, providing a new way to treat diabetes and obesity.
In fact, considering the brain–intestine axis, this may also be applied to the treatment of neurological disorders. Chen et al. constructed GLP-1-producing Lactococcus lactis MG1363 and demonstrated its neuroprotective effects in Alzheimer’s disease mice and Parkinson’s disease mice, possibly through the TLR4/NF-κB and AKT/GSK3β signaling pathways [248]. Orally, angiotensin-expressing Lacticaseibacillus paracasei subsp. paracasei has also been found to promote beneficial circulating neurotransmitters and reduce neuroinflammatory cytokine expression in the prefrontal cortex [249]. Interestingly, the engineered Lactic acid bacteria in these studies both increased the abundance of AM, indicating that they may have a synergistic effect and can be considered in combination in treatment regimens.
4.4.2. Probiotic Vaccines
Another crucial application for them is shown in probiotic vaccines (Table 2). Many studies have used probiotics to express the antigens of some pathogens and stimulate the body to secrete antibodies in large quantities. In pig farming, Lacticaseibacillus casei was modified to constitutively express E2 protein, the envelope protein of bovine viral diarrhea virus (BVDV) that possesses important epitopes, fused with cholera toxin B as an adjuvant to help antigen presentation [251]. Similarly, this strategy has also been applied to prevent the infection of porcine transmissible gastroenteritis virus (TGEV) and porcine epidemic diarrhea virus (PEDV) [252,253,254,259]. In fish culture, the recombinant Lacticaseibacillus casei expressing antigen VP2 of infectious pancreatic necrosis virus (IPNV) can induce a higher systemic immune response against infection to protect rainbow trout [255,256]. In addition to viruses, the presence of pathogenic bacteria can also cause economic damage to the pig industry. Lacticaseibacillus casei was chosen to express F4 fimbrial adhesin main subunit and conjunction with adjuvants to combat F4+ enterotoxigenic Escherichia coli infections [258]. The anti-bacteria effect of the engineered Lacticaseibacillus casei carrying antigens of the corresponding pathogenic bacteria was also demonstrated in mouse models of Staphylococcus aureus [260] and Clostridium perfringens [257] infection. In general, Lactic acid bacteria have the potential to become oral vaccines against infection.
Another strategy is to use probiotics to express the receptor for the virus, thereby trapping the virus and reducing its ability to spread. Lactobacillus acidophilus was constructed by chromosomal integration to surface display human CD4 [261]. This engineered bacterium can capture HIV-1 and reduce the efficiency of HIV-1 transmission. However, this strategy may be difficult to intercept non-enterohepatic axis-invading pathogens.
4.5. Summary
Currently, the main strategy for the modification of probiotics is the expression of therapeutic factors for direct or adjuvant therapy. The others are to use them or their derivatives (BGs, OMVs, minicells, etc.) as carriers, utilizing their targeting or immunoactivities to transport therapeutic factors or to display antigens on the surface for probiotic vaccines. Probiotic treatment is mainly manifested in the intestinal tract and dispersed through the intestine–liver axis, the brain–intestinal axis, and so on. It has a greater potential for the treatment of digestive and metabolic diseases and some neurological disorders. Probiotic engineering has also excelled in the fight against infection via competition, quorum sensing, or masquerading itself as a host cell to hinder pathogens, but this often seems to be limited to the gut.
5. Challenges and Outlooks
As our friends, probiotics can bring positive health benefits and offer new treatments for diseases. However, the application of probiotic therapy still faces a range of risks and challenges.
The security of probiotics is an essential challenge for their applications. Notably, there is a pathogenicity island on the genome of EcN that produces colibactin, a genotoxin that induces DNA damage [48]. While much research has focused on its positive effects, we can never lose sight of this negative one. To develop EcN without colibactin risk, polyphosphate kinase inhibitors (e.g., mesalamine) can be used to inhibit colibactin production [274]. Another strategy is to delete or mutate virulence genes/virulence expression-dependent genes. However, we have to ascertain whether such manipulation will affect the normal metabolic function of the corresponding bacteria or not.
Although AM currently appears to be a safe probiotic. There are still some contrary observations implying that AM is not a probiotic in all cases. For example, a high abundance of AM was detected in samples from patients with type 2 diabetes [275], and associated with the development of inflammation in some cases [276,277]. In addition, as a mucosal degrading bacterium, AM also disrupts mucus barrier function [278], which may lead to mucosal immune damage. These controversies may be due to various periods/stages of disease development or to different host states/microenvironments. For the maximum value of AM, it is now imperative to clarify its exact mechanism and how it interacts with the host and commensal bacteria, so we can prevent damage caused by its misuse.
CB has also been reported to acquire the type E botulinum neurotoxin gene [279,280]. Therefore, we should carefully select the engineered strains and manage them to avoid the acquisition of exogenous genes and to prevent the flow of their genes to other organisms. However, during the period of clinical treatment, isolated adverse cases have been reported where the administration of CB for biliary cancer has led to bacteremia [281]. This may be related to reduced immunity and a broken epithelial barrier, enlightening us to take into account the condition of patients to determine if probiotic therapy is appropriate.
In contrast, Lactic acid bacteria and Bifidobacterium spp. appear to be relatively safe. They are commonly added as ingredients in food products and no serious adverse effects have been observed.
Additionally, there are risks associated with the use of modified bacteria. Exogenous genes can be transferred to other organisms through genetic drift, resulting in contamination of the gene pool in the ecosystem. Moreover, specific antibiotic resistance genes are often used as marker genes in the genetic manipulation of bacteria. This resistance gene has a certain probability of being transferred to other organisms, making them develop resistance. In modified probiotics application, attention also needs to be paid to the possibility of these bacteria triggering a violent immune response (due to the exogenous proteins) that could jeopardize the safety of the organism. It has to be admitted that every treatment involves a certain amount of risk. Some bacteria may have side-effects reported, but we cannot cover the whole picture with bias. Considering the positive impact of probiotics in the treatment of various diseases, we should not abandon the use of promising probiotics for isolated cases. Therefore, it is necessary to determine the population for which probiotics are suitable, including assessments of the age, gender, health level, and so on.
Common methods of administration include oral, injectable, and nebulized inhalation methods, while probiotic administration can be most conveniently done orally. Therefore, the ability of probiotics to adhere and colonize can often be critical to the efficacy of the treatment. Yang et al. found that EcN with defective adsorption to epithelial cells did not exhibit the original efficacy against tumors [47]. Meanwhile, adherence is the basis for colonization, and the capacity to adhere can also affect the dose of probiotics. Therefore, we should seek to improve the adherence of probiotics when applying them, for example, heterologous expressing surface proteins or cellular components that affect adhesion [282]. Following this, we still need to reassess the safety (paying particular attention to whether immunogenicity is altered) and efficacy of the engineered probiotics.
Currently, the mechanism of how probiotics achieve a protective effect on a particular organ is under-researched. Therefore, the strategy of using modifications to strengthen the original bacteria can be hampered. Furthermore, the interactions between multi-organs are still ambiguous, which limits the scope of probiotic application. Combined with the complexity of interactions among strains and the complex dietary factors [283], these difficulties make it hard for us to judge the role played by a specific probiotic in a particular microenvironment. In general, for the development of probiotic therapy, we need to strengthen the basic research related to probiotics, including the mechanism of the interactions between probiotics and different organs, the relationship between probiotics and commensal microorganisms, etc.
6. Conclusions
Native probiotics can confer health on the host when applied in moderation. They can be effective in the treatment of cancer, IBD, obesity, and other related diseases. The mechanisms behind them mainly include the regulation of the composition of the microbiota in the microenvironment, the beneficial effects of secreted substances and their bacteriological components, and activation of the immune system on the host.
Further modification of probiotics can give these native properties more targeted therapeutic factors that will deepen their efficacy and expand their application areas. However, the modification of probiotics is mainly based on the understanding of how probiotics play a probiotic role and how they interact with the organism/microenvironment. The existing inadequate theories of the underlying mechanisms may lead to unjustified modifications. Therefore, the safety of modified probiotics needs to be rigorously verified at the time of application.
Author Contributions
Writing—original draft preparation, H.S., Z.Z. (Zitong Zhao); writing—review and editing, Y.C., L.Z.; visualization, Z.Z. (Zengjue Zhao); supervision, L.Z.; project administration, L.Z.; funding acquisition, L.Z. All authors have read and agreed to the published version of the manuscript.
Funding
The study was funded by the Guangzhou Science and Technology Project (202002020056, 201903010078), Lingnan Modern Agriculture Key Project (mmkj2020026).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O.; Eid, J. Butyrate-Producing Human Gut Symbiont, Clostridium Butyricum, and Its Role in Health and Disease. Gut Microbes 2021, 13, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Lin, C.; Yu, J.; Qi, Q.; Wang, Q. Bioengineered Escherichia coli Nissle 1917 for Tumour-Targeting Therapy. Microb. Biotechnol. 2020, 13, 629–636. [Google Scholar] [CrossRef] [Green Version]
- Zou, Y.; Chen, T. Engineered Akkermansia muciniphila: A Promising Agent against Diseases (Review). Exp. Ther. Med. 2020, 20, 285. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface Components and Metabolites of Probiotics for Regulation of Intestinal Epithelial Barrier. Microb. Cell Fact. 2020, 19, 23. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Milagro, F.I.; Riezu-Boj, J.I.; Lorente-Cebrián, S. Effects of Gut Microbiota-Derived Extracellular Vesicles on Obesity and Diabetes and Their Potential Modulation through Diet. J. Physiol. Biochem. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Routy, B.; Gopalakrishnan, V.; Daillère, R.; Zitvogel, L.; Wargo, J.A.; Kroemer, G. The Gut Microbiota Influences Anticancer Immunosurveillance and General Health. Nat. Rev. Clin. Oncol. 2018, 15, 382–396. [Google Scholar] [CrossRef]
- Liu, Q.; Gai, Y.; Chen, Y.; Lan, X.; Jiang, D. Escherichia coli Nissle 1917 as a Novel Microrobot for Tumor-Targeted Imaging and Therapy. Pharmaceutics 2021, 13, 1226. [Google Scholar] [CrossRef] [PubMed]
- Sonnenborn, U. Escherichia coli Strain Nissle 1917—From Bench to Bedside and Back: History of a Special Escherichia coli Strain with Probiotic Properties. FEMS Microbiol. Lett. 2016, 363, fnw212. [Google Scholar] [CrossRef] [Green Version]
- Hu, R.; Lin, H.; Li, J.; Zhao, Y.; Wang, M.; Sun, X.; Min, Y.; Gao, Y.; Yang, M. Probiotic Escherichia coli Nissle 1917—Derived Outer Membrane Vesicles Enhance Immunomodulation and Antimicrobial Activity in RAW264.7 Macrophages. BMC Microbiol. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, L.; Chen, S.; Guo, S.; Yue, T.; Hou, Q.; Feng, M.; Xu, H.; Liu, Y.; Wang, P.; et al. The Administration of Escherichia coli Nissle 1917 Ameliorates Irinotecan-Induced Intestinal Barrier Dysfunction and Gut Microbial Dysbiosis in Mice. Life Sci. 2019, 231, 116529. [Google Scholar] [CrossRef]
- Secher, T.; Kassem, S.; Benamar, M.; Bernard, I.; Boury, M.; Barreau, F.; Oswald, E.; Saoudi, A. Oral Administration of the Probiotic Strain Escherichia coli Nissle 1917 Reduces Susceptibility to Neuroinflammation and Repairs Experimental Autoimmune Encephalomyelitis-Induced Intestinal Barrier Dysfunction. Front. Immunol. 2017, 8, 1096. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.L.S.; Campos, C.L.V.; Reis, D.C.; Cassali, G.D.; Generoso, S.V.; Cardoso, V.N.; Azevedo, V.; Medeiros, J.D.; Fernandes, G.R.; Nicoli, J.R.; et al. Beneficial Effects Resulting from Oral Administration of Escherichia coli Nissle 1917 on a Chronic Colitis Model. Benef. Microbes 2020, 11, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Maltby, R.; Leatham-Strains Jensen, M.P.; Gibson, T.; Cohen, P.S.; Conway, T. Nutritional Basis for Colonization Resistance by Human Commensal Escherichia coli HS and Nissle 1917 against E. Coli O157:H7 in the Mouse Intestine. PLoS ONE 2013, 8, e53957. [Google Scholar] [CrossRef] [Green Version]
- Schinner, S.A.C.; Mokszycki, M.E.; Adediran, J.; Leatham-Jensen, M.; Conway, T.; Cohen, P.S. Escherichia coli EDL933 Requires Gluconeogenic Nutrients to Successfully Colonize the Intestines of Streptomycin-Treated Mice Precolonized with E. Coli Nissle 1917. Infect. Immun. 2015, 83, 1983–1991. [Google Scholar] [CrossRef] [Green Version]
- Reister, M.; Hoffmeier, K.; Krezdorn, N.; Rotter, B.; Liang, C.; Rund, S.; Dandekar, T.; Sonnenborn, U.; Oelschlaeger, T.A. Complete Genome Sequence of the Gram-Negative Probiotic Escherichia coli Strain Nissle 1917. J. Biotechnol. 2014, 187, 106–107. [Google Scholar] [CrossRef] [PubMed]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia muciniphila Gen. Nov., Sp. Nov., a Human Intestinal Mucin-Degrading Bacterium. Int. J. Syst Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Bohn, T.; Castenmiller, J.; De Henauw, S.; Hirsch-Ernst, K.I.; Maciuk, A.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Safety of Pasteurised Akkermansia muciniphila as a Novel Food Pursuant to Regulation (EU) 2015/2283. EFSA J. 2021, 19, e06780. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Q.; Feng, S.; Arjan, N.; Chen, W. A next Generation Probiotic, Akkermansia muciniphila. Crit. Rev. Food Sci. Nutr. 2019, 59, 3227–3236. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.; Li, X.; Sun, Y.; Zhao, J.; Miao, S.; Xiong, Q.; Zhang, Y.; Zhang, G. Comparative Genomic and Functional Analysis of Akkermansia muciniphila and Closely Related Species. Genes Genom. 2019, 41, 1253–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottman, N.; Reunanen, J.; Meijerink, M.; Pietilä, T.E.; Kainulainen, V.; Klievink, J.; Huuskonen, L.; Aalvink, S.; Skurnik, M.; Boeren, S.; et al. Pili-like Proteins of Akkermansia muciniphila Modulate Host Immune Responses and Gut Barrier Function. PLoS ONE 2017, 12, e0173004. [Google Scholar] [CrossRef]
- Grajeda-Iglesias, C.; Durand, S.; Daillère, R.; Iribarren, K.; Lemaitre, F.; Derosa, L.; Aprahamian, F.; Bossut, N.; Nirmalathasan, N.; Madeo, F.; et al. Oral Administration of Akkermansia muciniphila Elevates Systemic Antiaging and Anticancer Metabolites. Aging 2021, 13, 6375–6405. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Corleto, J.; Ruegger, P.M.; Logan, G.D.; Peacock, B.B.; Mendonca, S.; Yamaki, S.; Adamson, T.; Ermel, R.; McKemy, D.; et al. Dominant Role of the Gut Microbiota in Chemotherapy Induced Neuropathic Pain. Sci. Rep. 2019, 9, 20324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Wang, J.; Zhang, H.; Wu, S.; Qi, G. Supplemental Clostridium Butyricum Modulates Lipid Metabolism Through Shaping Gut Microbiota and Bile Acid Profile of Aged Laying Hens. Front. Microbiol. 2020, 11, 600. [Google Scholar] [CrossRef]
- Xia, X.; Chen, J.; Xia, J.; Wang, B.; Liu, H.; Yang, L.; Wang, Y.; Ling, Z. Role of Probiotics in the Treatment of Minimal Hepatic Encephalopathy in Patients with HBV-Induced Liver Cirrhosis. J. Int. Med. Res. 2018, 46, 3596–3604. [Google Scholar] [CrossRef]
- Vieira, A.T.; Fukumori, C.; Ferreira, C.M. New Insights into Therapeutic Strategies for Gut Microbiota Modulation in Inflammatory Diseases. Clin. Transl. Immunol. 2016, 5, e87. [Google Scholar] [CrossRef] [Green Version]
- Patil, P.; Bhandary, S.K.; Haridas, V.; Sarathkumar, E.; Shetty, P. Is Butyrate a Natural Alternative to Dexamethasone in the Management of COVID-19? F1000 Res. 2021, 10, 273. [Google Scholar] [CrossRef]
- Zhao, Q.; Yang, W.-R.; Wang, X.-H.; Li, G.-Q.; Xu, L.-Q.; Cui, X.; Liu, Y.; Zuo, X.-L. Clostridium Butyricum Alleviates Intestinal Low-Grade Inflammation in TNBS-Induced Irritable Bowel Syndrome in Mice by Regulating Functional Status of Lamina Propria Dendritic Cells. World J. Gastroenterol. 2019, 25, 5469–5482. [Google Scholar] [CrossRef]
- Matsuoka, K.; Kanai, T. The Gut Microbiota and Inflammatory Bowel Disease. Semin. Immunopathol. 2015, 37, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Zhang, L.; Zhan, X.; Zeng, X.; Zhou, L.; Cao, G.; Chen, A.; Yang, C. Effects of Dietary Supplementation of Probiotic, Clostridium Butyricum, on Growth Performance, Immune Response, Intestinal Barrier Function, and Digestive Enzyme Activity in Broiler Chickens Challenged with Escherichia coli K88. J. Anim. Sci. Biotechnol. 2016, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.M.; Cao, G.T.; Ferket, P.R.; Liu, T.T.; Zhou, L.; Zhang, L.; Xiao, Y.P.; Chen, A.G. Effects of Probiotic, Clostridium Butyricum, on Growth Performance, Immune Function, and Cecal Microflora in Broiler Chickens. Poult. Sci. 2012, 91, 2121–2129. [Google Scholar] [CrossRef] [PubMed]
- Brunt, J.; Cross, K.L.; Peck, M.W. Apertures in the Clostridium Sporogenes Spore Coat and Exosporium Align to Facilitate Emergence of the Vegetative Cell. Food Microbiol. 2015, 51, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishizaki, A.; Bi, X.; Nguyen, L.V.; Matsuda, K.; Pham, H.V.; Phan, C.T.T.; Khu, D.T.K.; Ichimura, H. Effects of Short-Term Probiotic Ingestion on Immune Profiles and Microbial Translocation among HIV-1-Infected Vietnamese Children. Int. J. Mol. Sci. 2017, 18, 2185. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Tan, C.; Chen, D.; Liu, S. Potential of Three Probiotic Lactobacilli in Transforming Star Fruit Juice into Functional Beverages. Food Sci. Nutr. 2018, 6, 2141–2150. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Roos, S.; Eeckhaut, V.; MacKenzie, D.A.; Derde, M.; Verstraete, W.; Marzorati, M.; Possemiers, S.; Vanhoecke, B.; Van Immerseel, F.; et al. Incorporating a Mucosal Environment in a Dynamic Gut Model Results in a More Representative Colonization by Lactobacilli. Microb. Biotechnol. 2012, 5, 106–115. [Google Scholar] [CrossRef] [Green Version]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the Probiotic Potential of Lactic Acid Bacteria from African Traditional Fermented Foods and Beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Li, J.; Zhang, H.; Xie, Y.; Xiong, L.; Liu, H.; Wang, F. Effects of a Probiotic on the Growth Performance, Intestinal Flora, and Immune Function of Chicks Infected with Salmonella Pullorum. Poult. Sci. 2020, 99, 5316–5323. [Google Scholar] [CrossRef]
- Zou, Y.-J.; Xu, J.-J.; Wang, X.; Zhu, Y.-H.; Wu, Q.; Wang, J.-F. Lactobacillus Johnsonii L531 Ameliorates Escherichia coli-Induced Cell Damage via Inhibiting NLRP3 Inflammasome Activity and Promoting ATG5/ATG16L1-Mediated Autophagy in Porcine Mammary Epithelial Cells. Vet. Sci. 2020, 7, 112. [Google Scholar] [CrossRef]
- Kim, J.E.; Sharma, A.; Sharma, G.; Lee, S.Y.; Shin, H.S.; Rudra, D.; Im, S.-H. Lactobacillus Pentosus Modulates Immune Response by Inducing IL-10 Producing Tr1 Cells. Immune Netw. 2019, 19, e39. [Google Scholar] [CrossRef] [PubMed]
- Guglielmetti, S.; Tamagnini, I.; Mora, D.; Minuzzo, M.; Scarafoni, A.; Arioli, S.; Hellman, J.; Karp, M.; Parini, C. Implication of an Outer Surface Lipoprotein in Adhesion of Bifidobacterium Bifidum to Caco-2 Cells. Appl. Environ. Microbiol. 2008, 74, 4695–4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheikhyoussef, A.; Pogori, N.; Chen, H.; Tian, F.; Chen, W.; Tang, J.; Zhang, H. Antimicrobial Activity and Partial Characterization of Bacteriocin-like Inhibitory Substances (BLIS) Produced by Bifidobacterium Infantis BCRC 14602. Food Control 2009, 20, 553–559. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as Vitamin Suppliers to Their Host: A Gut Microbiota Perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile Resistance Mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.K.; Ranjan, A.; Srivastava, A.K.; Singh, M.; Shukla, A.K.; Atri, N.; Mishra, A.; Singh, A.K.; Singh, S.K. Cytotoxic and Apoptotic Inducing Activity of Amoora Rohituka Leaf Extracts in Human Breast Cancer Cells. J. Ayurveda Integr. Med. 2020, 11, 383–390. [Google Scholar] [CrossRef]
- Thomas, A.M.; Manghi, P.; Asnicar, F.; Pasolli, E.; Armanini, F.; Zolfo, M.; Beghini, F.; Manara, S.; Karcher, N.; Pozzi, C.; et al. Metagenomic Analysis of Colorectal Cancer Datasets Identifies Cross-Cohort Microbial Diagnostic Signatures and a Link with Choline Degradation. Nat. Med. 2019, 25, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Helbig, L.; Fu, J.; Bian, X.; Herrmann, J.; Baumann, M.; Stewart, A.F.; Müller, R.; Li, A.; Zips, D.; et al. Expressing Cytotoxic Compounds in Escherichia coli Nissle 1917 for Tumor-Targeting Therapy. Res. Microbiol. 2019, 170, 74–79. [Google Scholar] [CrossRef]
- Alizadeh, S.; Esmaeili, A.; Omidi, Y. Anti-Cancer Properties of Escherichia coli Nissle 1917 against HT-29 Colon Cancer Cells through Regulation of Bax/Bcl-XL and AKT/PTEN Signaling Pathways. Iran. J. Basic Med. Sci. 2020, 23, 886–893. [Google Scholar] [CrossRef]
- Shi, L.; Sheng, J.; Wang, M.; Luo, H.; Zhu, J.; Zhang, B.; Liu, Z.; Yang, X. Combination Therapy of TGF-β Blockade and Commensal-Derived Probiotics Provides Enhanced Antitumor Immune Response and Tumor Suppression. Theranostics 2019, 9, 4115–4129. [Google Scholar] [CrossRef] [PubMed]
- Nougayrède, J.-P.; Chagneau, C.V.; Motta, J.-P.; Bossuet-Greif, N.; Belloy, M.; Taieb, F.; Gratadoux, J.-J.; Thomas, M.; Langella, P.; Oswald, E. A Toxic Friend: Genotoxic and Mutagenic Activity of the Probiotic Strain Escherichia coli Nissle 1917. mSphere 2021, 6, e0062421. [Google Scholar] [CrossRef]
- Vernocchi, P.; Gili, T.; Conte, F.; Del Chierico, F.; Conta, G.; Miccheli, A.; Botticelli, A.; Paci, P.; Caldarelli, G.; Nuti, M.; et al. Network Analysis of Gut Microbiome and Metabolome to Discover Microbiota-Linked Biomarkers in Patients Affected by Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2020, 21, 8730. [Google Scholar] [CrossRef]
- Lapidot, Y.; Amir, A.; Nosenko, R.; Uzan-Yulzari, A.; Veitsman, E.; Cohen-Ezra, O.; Davidov, Y.; Weiss, P.; Bradichevski, T.; Segev, S.; et al. Alterations in the Gut Microbiome in the Progression of Cirrhosis to Hepatocellular Carcinoma. mSystems 2020, 5, e00153-20. [Google Scholar] [CrossRef]
- Chen, Z.; Qian, X.; Chen, S.; Fu, X.; Ma, G.; Zhang, A. Akkermansia muciniphila Enhances the Antitumor Effect of Cisplatin in Lewis Lung Cancer Mice. J. Immunol. Res. 2020, 2020, 2969287. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Sheng, J.; Chen, G.; Zhu, P.; Shi, C.; Li, B.; Park, C.; Wang, J.; Zhang, B.; Liu, Z.; et al. Combining IL-2-Based Immunotherapy with Commensal Probiotics Produces Enhanced Antitumor Immune Response and Tumor Clearance. J. Immunother. Cancer 2020, 8, e000973. [Google Scholar] [CrossRef]
- Daisley, B.A.; Chanyi, R.M.; Abdur-Rashid, K.; Al, K.F.; Gibbons, S.; Chmiel, J.A.; Wilcox, H.; Reid, G.; Anderson, A.; Dewar, M.; et al. Abiraterone Acetate Preferentially Enriches for the Gut Commensal Akkermansia muciniphila in Castrate-Resistant Prostate Cancer Patients. Nat. Commun. 2020, 11, 4822. [Google Scholar] [CrossRef]
- Panebianco, C.; Adamberg, K.; Jaagura, M.; Copetti, M.; Fontana, A.; Adamberg, S.; Kolk, K.; Vilu, R.; Andriulli, A.; Pazienza, V. Influence of Gemcitabine Chemotherapy on the Microbiota of Pancreatic Cancer Xenografted Mice. Cancer Chemother. Pharmacol. 2018, 81, 773–782. [Google Scholar] [CrossRef]
- Teng, L.; Wang, K.; Chen, W.; Wang, Y.; Bi, L. HYR-2 Plays an Anti-Lung Cancer Role by Regulating PD-L1 and Akkermansia muciniphila. Pharmacol. Res. 2020, 160, 105086. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, C.; Zhong, Y.N.; Zhao, F.; Hao, Z.; Xu, Y.; Lai, R.; Shen, G.; Yin, X. Effect and Mechanism of Vitamin D on the Development of Colorectal Cancer Based on Intestinal Flora Disorder. J. Gastroenterol. Hepatol. 2020, 35, 1023–1031. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Li, J.; Zhang, Y.; Guo, Y.; Chang, Q.; Chen, L.; Wang, Y.; Wang, S.; Song, Y.; et al. Sini Decoction Ameliorates Colorectal Cancer and Modulates the Composition of Gut Microbiota in Mice. Front. Pharmacol. 2021, 12, 609992. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Wang, W.; Lan, T.; Yang, W.; Yu, D.; Fang, X.; Wu, H. A Purified Aspartic Protease from Akkermansia muciniphila Plays an Important Role in Degrading Muc2. Int. J. Mol. Sci. 2019, 21, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Zhang, J.; Wu, H.; Yu, D.; Fang, X. Akkermansia muciniphila Aspartic Protease Amuc_1434* Inhibits Human Colorectal Cancer LS174T Cell Viability via TRAIL-Mediated Apoptosis Pathway. Int. J. Mol. Sci. 2020, 21, 3385. [Google Scholar] [CrossRef]
- Wang, L.; Tang, L.; Feng, Y.; Zhao, S.; Han, M.; Zhang, C.; Yuan, G.; Zhu, J.; Cao, S.; Wu, Q.; et al. A Purified Membrane Protein from Akkermansia muciniphila or the Pasteurised Bacterium Blunts Colitis Associated Tumourigenesis by Modulation of CD8+ T Cells in Mice. Gut 2020, 69, 1988–1997. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Xu, C.; Ge, Q.; Lin, Y.; Wong, C.C.; Qi, Y.; Ye, B.; Lian, Q.; Zhuo, W.; Si, J.; et al. A. Muciniphila Suppresses Colorectal Tumorigenesis by Inducing TLR2/NLRP3-Mediated M1-Like TAMs. Cancer Immunol. Res. 2021, 9, 1111–1124. [Google Scholar] [CrossRef]
- Luo, Z.-W.; Xia, K.; Liu, Y.-W.; Liu, J.-H.; Rao, S.-S.; Hu, X.-K.; Chen, C.-Y.; Xu, R.; Wang, Z.-X.; Xie, H. Extracellular Vesicles from Akkermansia muciniphila Elicit Antitumor Immunity Against Prostate Cancer via Modulation of CD8+ T Cells and Macrophages. Int. J. Nanomed. 2021, 16, 2949–2963. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, T.; Tu, X.; Huang, Y.; Zhang, H.; Tan, D.; Jiang, W.; Cai, S.; Zhao, P.; Song, R.; et al. Gut Microbiome Affects the Response to Anti-PD-1 Immunotherapy in Patients with Hepatocellular Carcinoma. J. Immunother. Cancer 2019, 7, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut Microbiome Influences Efficacy of PD-1-Based Immunotherapy against Epithelial Tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salgia, N.J.; Bergerot, P.G.; Maia, M.C.; Dizman, N.; Hsu, J.; Gillece, J.D.; Folkerts, M.; Reining, L.; Trent, J.; Highlander, S.K.; et al. Stool Microbiome Profiling of Patients with Metastatic Renal Cell Carcinoma Receiving Anti-PD-1 Immune Checkpoint Inhibitors. Eur. Urol. 2020, 78, 498–502. [Google Scholar] [CrossRef]
- Xu, X.; Lv, J.; Guo, F.; Li, J.; Jia, Y.; Jiang, D.; Wang, N.; Zhang, C.; Kong, L.; Liu, Y.; et al. Gut Microbiome Influences the Efficacy of PD-1 Antibody Immunotherapy on MSS-Type Colorectal Cancer via Metabolic Pathway. Front. Microbiol. 2020, 11, 814. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.C.; Araya, R.E.; Huang, A.; Chen, Q.; Di Modica, M.; Rodrigues, R.R.; Lopès, A.; Johnson, S.B.; Schwarz, B.; Bohrnsen, E.; et al. Microbiota Triggers STING-Type I IFN-Dependent Monocyte Reprogramming of the Tumor Microenvironment. Cell 2021, 184, 5338–5356.e21. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xie, W.; Wan, X.; Deng, T. Clostridium Butyricum Modulates Gut Microbiota and Reduces Colitis Associated Colon Cancer in Mice. Int. Immunopharmacol. 2020, 88, 106862. [Google Scholar] [CrossRef]
- Chen, D.; Jin, D.; Huang, S.; Wu, J.; Xu, M.; Liu, T.; Dong, W.; Liu, X.; Wang, S.; Zhong, W.; et al. Clostridium Butyricum, a Butyrate-Producing Probiotic, Inhibits Intestinal Tumor Development through Modulating Wnt Signaling and Gut Microbiota. Cancer Lett. 2020, 469, 456–467. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, J.; Li, D.; Chen, M.; Liu, J.; Feng, C.; He, Q.; Zhao, J.; Zhang, L.; Chen, J.; et al. The Efficacy and Safety of Clostridium Butyricum and Bacillus Coagulans in Helicobacter Pylori Eradication Treatment: An Open-Label, Single-Arm Pilot Study. Medicine 2020, 99, e22976. [Google Scholar] [CrossRef]
- Shin, D.-S.; Rhee, K.-J.; Eom, Y.-B. Effect of Probiotic Clostridium Butyricum NCTC 7423 Supernatant on Biofilm Formation and Gene Expression of Bacteroides Fragilis. J. Microbiol. Biotechnol. 2020, 30, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.-W.; Li, R.-Q.; An, J.-X.; Xie, T.-Q.; Han, Z.-Y.; Xu, R.; Fang, Y.; Zhang, X.-Z. Prebiotics-Encapsulated Probiotic Spores Regulate Gut Microbiota and Suppress Colon Cancer. Adv. Mater. 2020, 32, e2004529. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Dai, X.; Li, K.; Gui, G.; Liu, J.; Yang, H. Clostridium Butyricum Partially Regulates the Development of Colitis-Associated Cancer through MiR-200c. Cell Mol. Biol. 2017, 63, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.; Li, M.; Song, W.; Jiang, R.; Li, Y.Q. Effects of Probiotics on Chemotherapy in Patients with Lung Cancer. Oncol. Lett. 2019, 17, 2836–2848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, M.; Xie, Q.; Ma, L.; An, G.; Xiao, J.; Li, J.; Liu, X.; Gao, P.; Liang, J.; Li, Y. Synergistic Anti-Tumour Effects of Clostridium Butyricum in Combination with Apatinib in CT26 Colorectal Tumour-Bearing Mice. Anticancer Drugs 2019, 30, 991–997. [Google Scholar] [CrossRef]
- Tomita, Y.; Ikeda, T.; Sakata, S.; Saruwatari, K.; Sato, R.; Iyama, S.; Jodai, T.; Akaike, K.; Ishizuka, S.; Saeki, S.; et al. Association of Probiotic Clostridium Butyricum Therapy with Survival and Response to Immune Checkpoint Blockade in Patients with Lung Cancer. Cancer Immunol. Res. 2020, 8, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.-C.; Ho, P.-Y.; Chen, W.-J.; Wu, S.-H.; Pan, M.-H. Lactobacillus Fermentum V3 Ameliorates Colitis-Associated Tumorigenesis by Modulating the Gut Microbiome. Am. J. Cancer Res. 2020, 10, 1170–1181. [Google Scholar]
- Tukenmez, U.; Aktas, B.; Aslim, B.; Yavuz, S. The Relationship between the Structural Characteristics of Lactobacilli-EPS and Its Ability to Induce Apoptosis in Colon Cancer Cells in Vitro. Sci. Rep. 2019, 9, 8268. [Google Scholar] [CrossRef] [Green Version]
- Ayyash, M.; Johnson, S.K.; Liu, S.-Q.; Mesmari, N.; Dahmani, S.; Al Dhaheri, A.S.; Kizhakkayil, J. In Vitro Investigation of Bioactivities of Solid-State Fermented Lupin, Quinoa and Wheat Using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef]
- Rezasoltani, S.; Asadzadeh Aghdaei, H.; Dabiri, H.; Akhavan Sepahi, A.; Modarressi, M.H.; Nazemalhosseini Mojarad, E. The Association between Fecal Microbiota and Different Types of Colorectal Polyp as Precursors of Colorectal Cancer. Microb. Pathog. 2018, 124, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Wada, M.; Nagata, S.; Saito, M.; Shimizu, T.; Yamashiro, Y.; Matsuki, T.; Asahara, T.; Nomoto, K. Effects of the Enteral Administration of Bifidobacterium Breve on Patients Undergoing Chemotherapy for Pediatric Malignancies. Support. Care Cancer 2010, 18, 751–759. [Google Scholar] [CrossRef]
- Raj, R.; Das, S. Development and Application of Anticancer Fluorescent CdS Nanoparticles Enriched Lactobacillus Bacteria as Therapeutic Microbots for Human Breast Carcinoma. Appl. Microbiol. Biotechnol. 2017, 101, 5439–5451. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium Promotes Antitumor Immunity and Facilitates Anti-PD-L1 Efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of Gut Microbiota in Inflammatory Bowel Disease (IBD): Cause or Consequence? IBD Treatment Targeting the Gut Microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Bawardy, B.; Shivashankar, R.; Proctor, D.D. Novel and Emerging Therapies for Inflammatory Bowel Disease. Front. Pharmacol. 2021, 12, 651415. [Google Scholar] [CrossRef]
- Jukic, A.; Bakiri, L.; Wagner, E.F.; Tilg, H.; Adolph, T.E. Calprotectin: From Biomarker to Biological Function. Gut 2021, 70, 1978–1988. [Google Scholar] [CrossRef] [PubMed]
- Damaskos, D.; Kolios, G. Probiotics and Prebiotics in Inflammatory Bowel Disease: Microflora “on the Scope”. Br. J. Clin. Pharmacol. 2008, 65, 453–467. [Google Scholar] [CrossRef] [Green Version]
- Yoshimatsu, Y.; Mikami, Y.; Kanai, T. Bacteriotherapy for Inflammatory Bowel Disease. Inflamm. Regen. 2021, 41, 3. [Google Scholar] [CrossRef] [PubMed]
- Schultz, M. Clinical Use of E. Coli Nissle 1917 in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2008, 14, 1012–1018. [Google Scholar] [CrossRef]
- Kruis, W.; Fric, P.; Pokrotnieks, J.; Lukás, M.; Fixa, B.; Kascák, M.; Kamm, M.A.; Weismueller, J.; Beglinger, C.; Stolte, M.; et al. Maintaining Remission of Ulcerative Colitis with the Probiotic Escherichia coli Nissle 1917 Is as Effective as with Standard Mesalazine. Gut 2004, 53, 1617–1623. [Google Scholar] [CrossRef]
- Behrouzi, A.; Mazaheri, H.; Falsafi, S.; Tavassol, Z.H.; Moshiri, A.; Siadat, S.D. Intestinal Effect of the Probiotic Escherichia coli Strain Nissle 1917 and Its OMV. J. Diabetes Metab. Disord. 2020, 19, 597–604. [Google Scholar] [CrossRef]
- Waidmann, M.; Bechtold, O.; Frick, J.-S.; Lehr, H.-A.; Schubert, S.; Dobrindt, U.; Loeffler, J.; Bohn, E.; Autenrieth, I.B. Bacteroides Vulgatus Protects against Escherichia coli-Induced Colitis in Gnotobiotic Interleukin-2-Deficient Mice. Gastroenterology 2003, 125, 162–177. [Google Scholar] [CrossRef]
- Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Rodríguez-Sojo, M.J.; Rodríguez-Cabezas, M.E.; Olivares, M.; García, F.; Gálvez, J.; Morón, R.; Rodríguez-Nogales, A. Intestinal Anti-Inflammatory Effects of Probiotics in DNBS-Colitis via Modulation of Gut Microbiota and MicroRNAs. Eur. J. Nutr. 2021, 60, 2537–2551. [Google Scholar] [CrossRef]
- Bian, Z.; Li, L.; Cui, J.; Zhang, H.; Liu, Y.; Zhang, C.-Y.; Zen, K. Role of MiR-150-Targeting c-Myb in Colonic Epithelial Disruption during Dextran Sulphate Sodium-Induced Murine Experimental Colitis and Human Ulcerative Colitis. J. Pathol. 2011, 225, 544–553. [Google Scholar] [CrossRef]
- Rodríguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Vezza, T.; Utrilla, M.P.; Chueca, N.; Fernández-Caballero, J.A.; García, F.; Rodríguez-Cabezas, M.E.; Gálvez, J. The Administration of Escherichia coli Nissle 1917 Ameliorates Development of DSS-Induced Colitis in Mice. Front. Pharmacol. 2018, 9, 468. [Google Scholar] [CrossRef]
- Zhang, X.; Han, Y.; Huang, W.; Jin, M.; Gao, Z. The Influence of the Gut Microbiota on the Bioavailability of Oral Drugs. Acta Pharm. Sin. B 2021, 11, 1789–1812. [Google Scholar] [CrossRef] [PubMed]
- Fábrega, M.-J.; Rodríguez-Nogales, A.; Garrido-Mesa, J.; Algieri, F.; Badía, J.; Giménez, R.; Gálvez, J.; Baldomà, L. Intestinal Anti-Inflammatory Effects of Outer Membrane Vesicles from Escherichia coli Nissle 1917 in DSS-Experimental Colitis in Mice. Front. Microbiol. 2017, 8, 1274. [Google Scholar] [CrossRef]
- Geerlings, S.Y.; Kostopoulos, I.; de Vos, W.M.; Belzer, C. Akkermansia muciniphila in the Human Gastrointestinal Tract: When, Where, and How? Microorganisms 2018, 6, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belzer, C.; de Vos, W.M. Microbes Inside—From Diversity to Function: The Case of Akkermansia. ISME J. 2012, 6, 1449–1458. [Google Scholar] [CrossRef]
- Paone, P.; Cani, P.D. Mucus Barrier, Mucins and Gut Microbiota: The Expected Slimy Partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Sugihara, K.; Gillilland, M.G.; Jon, S.; Kamada, N.; Moon, J.J. Hyaluronic Acid-Bilirubin Nanomedicine for Targeted Modulation of Dysregulated Intestinal Barrier, Microbiome and Immune Responses in Colitis. Nat. Mater. 2020, 19, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Reunanen, J.; Kainulainen, V.; Huuskonen, L.; Ottman, N.; Belzer, C.; Huhtinen, H.; de Vos, W.M.; Satokari, R. Akkermansia muciniphila Adheres to Enterocytes and Strengthens the Integrity of the Epithelial Cell Layer. Appl. Environ. Microbiol. 2015, 81, 3655–3662. [Google Scholar] [CrossRef] [Green Version]
- Earley, H.; Lennon, G.; Balfe, Á.; Coffey, J.C.; Winter, D.C.; O’Connell, P.R. The Abundance of Akkermansia muciniphila and Its Relationship with Sulphated Colonic Mucins in Health and Ulcerative Colitis. Sci. Rep. 2019, 9, 15683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Li, P.; Wu, X.; Lu, G.; Marcella, C.; Ji, X.; Ji, G.; Zhang, F. Alterations of Akkermansia muciniphila in the Inflammatory Bowel Disease Patients with Washed Microbiota Transplantation. Appl. Microbiol. Biotechnol. 2020, 104, 10203–10215. [Google Scholar] [CrossRef]
- Zhai, R.; Xue, X.; Zhang, L.; Yang, X.; Zhao, L.; Zhang, C. Strain-Specific Anti-Inflammatory Properties of Two Akkermansia muciniphila Strains on Chronic Colitis in Mice. Front. Cell Infect. Microbiol. 2019, 9, 239. [Google Scholar] [CrossRef] [PubMed]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short Chain Fatty Acids and Its Producing Organisms: An Overlooked Therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Kaczmarczyk, O.; Dąbek-Drobny, A.; Woźniakiewicz, M.; Paśko, P.; Dobrowolska-Iwanek, J.; Woźniakiewicz, A.; Piątek-Guziewicz, A.; Zagrodzki, P.; Mach, T.; Zwolińska-Wcisło, M. Fecal Levels of Lactic, Succinic and Short-Chain Fatty Acids in Patients with Ulcerative Colitis and Crohn Disease: A Pilot Study. J. Clin. Med. 2021, 10, 4701. [Google Scholar] [CrossRef]
- Araki, Y.; Andoh, A.; Takizawa, J.; Takizawa, W.; Fujiyama, Y. Clostridium Butyricum, a Probiotic Derivative, Suppresses Dextran Sulfate Sodium-Induced Experimental Colitis in Rats. Int. J. Mol. Med. 2004, 13, 577–580. [Google Scholar] [CrossRef]
- Scott, N.A.; Andrusaite, A.; Andersen, P.; Lawson, M.; Alcon-Giner, C.; Leclaire, C.; Caim, S.; Le Gall, G.; Shaw, T.; Connolly, J.P.R.; et al. Antibiotics Induce Sustained Dysregulation of Intestinal T Cell Immunity by Perturbing Macrophage Homeostasis. Sci. Transl. Med. 2018, 10, eaao4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.-Q.; Ding, T.T.; Zhao, J.-S.; Yang, X.; Zhang, H.-X.; Zhang, J.-J.; Cui, Y.-L. Therapeutic Effects of Clostridium Butyricum on Experimental Colitis Induced by Oxazolone in Rats. World J. Gastroenterol. 2009, 15, 1821–1828. [Google Scholar] [CrossRef]
- Hayashi, A.; Sato, T.; Kamada, N.; Mikami, Y.; Matsuoka, K.; Hisamatsu, T.; Hibi, T.; Roers, A.; Yagita, H.; Ohteki, T.; et al. A Single Strain of Clostridium Butyricum Induces Intestinal IL-10-Producing Macrophages to Suppress Acute Experimental Colitis in Mice. Cell Host Microbe 2013, 13, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Kanai, T.; Mikami, Y.; Hayashi, A. A Breakthrough in Probiotics: Clostridium Butyricum Regulates Gut Homeostasis and Anti-Inflammatory Response in Inflammatory Bowel Disease. J. Gastroenterol. 2015, 50, 928–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, M.; Zeng, L.; Li, L.-J.; Mo, L.-H.; Xie, R.-D.; Feng, B.-S.; Zheng, P.-Y.; Liu, Z.-G.; Liu, Z.-J.; Yang, P.-C. Specific Immunotherapy Ameliorates Ulcerative Colitis. Allergy Asthma Clin. Immunol. 2016, 12, 37. [Google Scholar] [CrossRef] [Green Version]
- Araki, Y.; Fujiyama, Y.; Andoh, A.; Koyama, S.; Kanauchi, O.; Bamba, T. The Dietary Combination of Germinated Barley Foodstuff plus Clostridium Butyricum Suppresses the Dextran Sulfate Sodium-Induced Experimental Colitis in Rats. Scand. J. Gastroenterol. 2000, 35, 1060–1067. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and Prebiotics in Intestinal Health and Disease: From Biology to the Clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chen, G.; Chen, C.; Zeng, X.; Ye, H. Prebiotics Effects in Vitro of Polysaccharides from Tea Flowers on Gut Microbiota of Healthy Persons and Patients with Inflammatory Bowel Disease. Int. J. Biol. Macromol. 2020, 158, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Mo, Q.; Liu, T.; Fu, A.; Ruan, S.; Zhong, H.; Tang, J.; Zhao, M.; Li, Y.; Zhu, S.; Cai, H.; et al. Novel Gut Microbiota Patterns Involved in the Attenuation of Dextran Sodium Sulfate-Induced Mouse Colitis Mediated by Glycerol Monolaurate via Inducing Anti-Inflammatory Responses. mBio 2021, 12, e0214821. [Google Scholar] [CrossRef]
- Mu, J.; Xu, J.; Wang, L.; Chen, C.; Chen, P. Anti-Inflammatory Effects of Purple Sweet Potato Anthocyanin Extract in DSS-Induced Colitis: Modulation of Commensal Bacteria and Attenuated Bacterial Intestinal Infection. Food Funct. 2021, 12, 11503–11514. [Google Scholar] [CrossRef]
- Ghavami, S.B.; Yadegar, A.; Aghdaei, H.A.; Sorrentino, D.; Farmani, M.; Mir, A.S.; Azimirad, M.; Balaii, H.; Shahrokh, S.; Zali, M.R. Immunomodulation and Generation of Tolerogenic Dendritic Cells by Probiotic Bacteria in Patients with Inflammatory Bowel Disease. Int. J. Mol. Sci. 2020, 21, 6266. [Google Scholar] [CrossRef]
- Wang, N.; Wang, S.; Xu, B.; Liu, F.; Huo, G.; Li, B. Alleviation Effects of Bifidobacterium Animalis Subsp. Lactis XLTG11 on Dextran Sulfate Sodium-Induced Colitis in Mice. Microorganisms 2021, 9, 2093. [Google Scholar] [CrossRef]
- Yu, R.; Zuo, F.; Ma, H.; Chen, S. Exopolysaccharide-Producing Bifidobacterium Adolescentis Strains with Similar Adhesion Property Induce Differential Regulation of Inflammatory Immune Response in Treg/Th17 Axis of DSS-Colitis Mice. Nutrients 2019, 11, 782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Dharmaprakash, V.; Nighot, P.; Guo, S.; Nighot, M.; Do, T.; Ma, T.Y. Bifidobacterium Bifidum Enhances the Intestinal Epithelial Tight Junction Barrier and Protects against Intestinal Inflammation by Targeting the Toll-like Receptor-2 Pathway in an NF-ΚB-Independent Manner. Int. J. Mol. Sci. 2021, 22, 8070. [Google Scholar] [CrossRef]
- Zhao, L.; Xie, Q.; Etareri Evivie, S.; Liu, D.; Dong, J.; Ping, L.; Liu, F.; Li, B.; Huo, G. Bifidobacterium Dentium N8 with Potential Probiotic Characteristics Prevents LPS-Induced Intestinal Barrier Injury by Alleviating the Inflammatory Response and Regulating the Tight Junction in Caco-2 Cell Monolayers. Food Funct. 2021, 12, 7171–7184. [Google Scholar] [CrossRef] [PubMed]
- Leccese, G.; Bibi, A.; Mazza, S.; Facciotti, F.; Caprioli, F.; Landini, P.; Paroni, M. Probiotic Lactobacillus and Bifidobacterium Strains Counteract Adherent-Invasive Escherichia coli (AIEC) Virulence and Hamper IL-23/Th17 Axis in Ulcerative Colitis, but Not in Crohn’s Disease. Cells 2020, 9, 1824. [Google Scholar] [CrossRef]
- Engevik, M.A.; Herrmann, B.; Ruan, W.; Engevik, A.C.; Engevik, K.A.; Ihekweazu, F.; Shi, Z.; Luck, B.; Chang-Graham, A.L.; Esparza, M.; et al. Bifidobacterium Dentium-Derived y-Glutamylcysteine Suppresses ER-Mediated Goblet Cell Stress and Reduces TNBS-Driven Colonic Inflammation. Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef]
- Boles, A.; Kandimalla, R.; Reddy, P.H. Dynamics of Diabetes and Obesity: Epidemiological Perspective. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1026–1036. [Google Scholar] [CrossRef]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial Dysbiosis-Induced Obesity: Role of Gut Microbiota in Homoeostasis of Energy Metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef]
- Serino, M.; Luche, E.; Gres, S.; Baylac, A.; Bergé, M.; Cenac, C.; Waget, A.; Klopp, P.; Iacovoni, J.; Klopp, C.; et al. Metabolic Adaptation to a High-Fat Diet Is Associated with a Change in the Gut Microbiota. Gut 2012, 61, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Ashaolu, T.J.; Fernández-Tomé, S. Gut Mucosal and Adipose Tissues as Health Targets of the Immunomodulatory Mechanisms of Probiotics. Trends Food Sci. Technol. 2021, 112, 764–779. [Google Scholar] [CrossRef]
- Cerdó, T.; García-Santos, J.A.; Bermúdez, M.G.; Campoy, C. The Role of Probiotics and Prebiotics in the Prevention and Treatment of Obesity. Nutrients 2019, 11, 635. [Google Scholar] [CrossRef] [Green Version]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [Green Version]
- Ukena, S.N.; Singh, A.; Dringenberg, U.; Engelhardt, R.; Seidler, U.; Hansen, W.; Bleich, A.; Bruder, D.; Franzke, A.; Rogler, G.; et al. Probiotic Escherichia coli Nissle 1917 Inhibits Leaky Gut by Enhancing Mucosal Integrity. PLoS ONE 2007, 2, e1308. [Google Scholar] [CrossRef]
- Alvarez, C.-S.; Badia, J.; Bosch, M.; Giménez, R.; Baldomà, L. Outer Membrane Vesicles and Soluble Factors Released by Probiotic Escherichia coli Nissle 1917 and Commensal ECOR63 Enhance Barrier Function by Regulating Expression of Tight Junction Proteins in Intestinal Epithelial Cells. Front. Microbiol. 2016, 7, 1981. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.; Chen, S.; Ma, J.; Ma, Y.; Zhu, J.; Ma, Y.; Liu, Y.; Wang, P.; Pan, Y. Escherichia coli Nissle 1917 Protects Intestinal Barrier Function by Inhibiting NF-ΚB-Mediated Activation of the MLCK-P-MLC Signaling Pathway. Mediat. Inflamm. 2019, 2019, 5796491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veltman, K.; Hummel, S.; Cichon, C.; Sonnenborn, U.; Schmidt, M.A. Identification of Specific MiRNAs Targeting Proteins of the Apical Junctional Complex That Simulate the Probiotic Effect of E. Coli Nissle 1917 on T84 Epithelial Cells. Int. J. Biochem. Cell Biol. 2012, 44, 341–349. [Google Scholar] [CrossRef]
- Alvarez, C.-S.; Giménez, R.; Cañas, M.-A.; Vera, R.; Díaz-Garrido, N.; Badia, J.; Baldomà, L. Extracellular Vesicles and Soluble Factors Secreted by Escherichia coli Nissle 1917 and ECOR63 Protect against Enteropathogenic E. Coli-Induced Intestinal Epithelial Barrier Dysfunction. BMC Microbiol. 2019, 19, 166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prisciandaro, L.D.; Geier, M.S.; Chua, A.E.; Butler, R.N.; Cummins, A.G.; Sander, G.R.; Howarth, G.S. Probiotic Factors Partially Prevent Changes to Caspases 3 and 7 Activation and Transepithelial Electrical Resistance in a Model of 5-Fluorouracil-Induced Epithelial Cell Damage. Support. Care Cancer 2012, 20, 3205–3210. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Belda, E.; Prifti, E.; Everard, A.; Kayser, B.D.; Bouillot, J.-L.; Chevallier, J.-M.; Pons, N.; Le Chatelier, E.; Ehrlich, S.D.; et al. Akkermansia muciniphila Abundance Is Lower in Severe Obesity, but Its Increased Level after Bariatric Surgery Is Not Associated with Metabolic Health Improvement. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E446–E459. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.A.; Collier, F.; Mohebbi, M.; Stuart, A.L.; Loughman, A.; Pasco, J.A.; Jacka, F.N. Obesity, Akkermansia muciniphila, and Proton Pump Inhibitors: Is There a Link? Obes. Res. Clin. Pract. 2020, 14, 524–530. [Google Scholar] [CrossRef]
- Verhoog, S.; Taneri, P.E.; Roa Díaz, Z.M.; Marques-Vidal, P.; Troup, J.P.; Bally, L.; Franco, O.H.; Glisic, M.; Muka, T. Dietary Factors and Modulation of Bacteria Strains of Akkermansia muciniphila and Faecalibacterium Prausnitzii: A Systematic Review. Nutrients 2019, 11, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Zhou, K.; Zhang, Y.; Han, X.; Zhao, A.; Liu, J.; Qu, C.; Ge, K.; Huang, F.; Hernandez, B.; et al. Food Withdrawal Alters the Gut Microbiota and Metabolome in Mice. FASEB J. 2018, 32, 4878–4888. [Google Scholar] [CrossRef] [Green Version]
- Roopchand, D.E.; Carmody, R.N.; Kuhn, P.; Moskal, K.; Rojas-Silva, P.; Turnbaugh, P.J.; Raskin, I. Dietary Polyphenols Promote Growth of the Gut Bacterium Akkermansia muciniphila and Attenuate High-Fat Diet-Induced Metabolic Syndrome. Diabetes 2015, 64, 2847–2858. [Google Scholar] [CrossRef] [Green Version]
- Sheng, L.; Jena, P.K.; Liu, H.-X.; Hu, Y.; Nagar, N.; Bronner, D.N.; Settles, M.L.; Bäumler, A.J.; Wan, Y.-J.Y. Obesity Treatment by Epigallocatechin-3-Gallate-Regulated Bile Acid Signaling and Its Enriched Akkermansia muciniphila. FASEB J. 2018, 32, fj201800370R. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Régnier, M.; Rastelli, M.; Morissette, A.; Suriano, F.; Le Roy, T.; Pilon, G.; Delzenne, N.M.; Marette, A.; Van Hul, M.; Cani, P.D. Rhubarb Supplementation Prevents Diet-Induced Obesity and Diabetes in Association with Increased Akkermansia muciniphila in Mice. Nutrients 2020, 12, 2932. [Google Scholar] [CrossRef]
- Lee, B.-H.; Chen, C.-H.; Hsu, Y.-Y.; Chuang, P.-T.; Shih, M.-K.; Hsu, W.-H. Polysaccharides Obtained from Cordyceps Militaris Alleviate Hyperglycemia by Regulating Gut Microbiota in Mice Fed a High-Fat/Sucrose Diet. Foods 2021, 10, 1870. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, P.; Zhang, P.; Chang, Y.; Cui, M.; Duan, J. Protein-Bound β-Glucan from Coriolus Versicolor Has Potential for Use Against Obesity. Mol. Nutr. Food Res. 2019, 63, e1801231. [Google Scholar] [CrossRef]
- Wang, L.; Wu, Y.; Zhuang, L.; Chen, X.; Min, H.; Song, S.; Liang, Q.; Li, A.-D.; Gao, Q. Puerarin Prevents High-Fat Diet-Induced Obesity by Enriching Akkermansia muciniphila in the Gut Microbiota of Mice. PLoS ONE 2019, 14, e0218490. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, M.; Ohtake, N.; Kaneko, A.; Tsuchiya, N.; Imamura, S.; Iizuka, S.; Ishizawa, S.; Nishi, A.; Yamamoto, M.; Taketomi, A.; et al. Increase of Akkermansia muciniphila by a Diet Containing Japanese Traditional Medicine Bofutsushosan in a Mouse Model of Non-Alcoholic Fatty Liver Disease. Nutrients 2020, 12, 839. [Google Scholar] [CrossRef] [Green Version]
- Fujisaka, S.; Usui, I.; Nawaz, A.; Igarashi, Y.; Okabe, K.; Furusawa, Y.; Watanabe, S.; Yamamoto, S.; Sasahara, M.; Watanabe, Y.; et al. Bofutsushosan Improves Gut Barrier Function with a Bloom of Akkermansia muciniphila and Improves Glucose Metabolism in Mice with Diet-Induced Obesity. Sci. Rep. 2020, 10, 5544. [Google Scholar] [CrossRef]
- Zhang, J.; Ni, Y.; Qian, L.; Fang, Q.; Zheng, T.; Zhang, M.; Gao, Q.; Zhang, Y.; Ni, J.; Hou, X.; et al. Decreased Abundance of Akkermansia muciniphila Leads to the Impairment of Insulin Secretion and Glucose Homeostasis in Lean Type 2 Diabetes. Adv. Sci. 2021, 8, e2100536. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Bose, S.; Lim, S.; Seo, J.; Shin, J.; Lee, D.; Chung, W.-H.; Song, E.-J.; Nam, Y.-D.; Kim, H. Beneficial Effects of Newly Isolated Akkermansia muciniphila Strains from the Human Gut on Obesity and Metabolic Dysregulation. Microorganisms 2020, 8, 1413. [Google Scholar] [CrossRef] [PubMed]
- Dao, M.C.; Everard, A.; Aron-Wisnewsky, J.; Sokolovska, N.; Prifti, E.; Verger, E.O.; Kayser, B.D.; Levenez, F.; Chilloux, J.; Hoyles, L.; et al. Akkermansia muciniphila and Improved Metabolic Health during a Dietary Intervention in Obesity: Relationship with Gut Microbiome Richness and Ecology. Gut 2016, 65, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Lukovac, S.; Belzer, C.; Pellis, L.; Keijser, B.J.; de Vos, W.M.; Montijn, R.C.; Roeselers, G. Differential Modulation by Akkermansia muciniphila and Faecalibacterium Prausnitzii of Host Peripheral Lipid Metabolism and Histone Acetylation in Mouse Gut Organoids. mBio 2014, 5, e01438-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, Y.; Kuang, Z.; Li, C.; Guo, S.; Xu, Y.; Zhao, D.; Hu, Y.; Song, B.; Jiang, Z.; Ge, Z.; et al. Gut Akkermansia muciniphila Ameliorates Metabolic Dysfunction-Associated Fatty Liver Disease by Regulating the Metabolism of L-Aspartate via Gut-Liver Axis. Gut Microbes 2021, 13, 1–19. [Google Scholar] [CrossRef]
- Kim, S.; Lee, Y.; Kim, Y.; Seo, Y.; Lee, H.; Ha, J.; Lee, J.; Choi, Y.; Oh, H.; Yoon, Y. Akkermansia muciniphila Prevents Fatty Liver Disease, Decreases Serum Triglycerides, and Maintains Gut Homeostasis. Appl. Environ. Microbiol. 2020, 86, e03004–e03019. [Google Scholar] [CrossRef]
- Bodogai, M.; O’Connell, J.; Kim, K.; Kim, Y.; Moritoh, K.; Chen, C.; Gusev, F.; Vaughan, K.; Shulzhenko, N.; Mattison, J.A.; et al. Commensal Bacteria Contribute to Insulin Resistance in Aging by Activating Innate B1a Cells. Sci. Transl. Med. 2018, 10, eaat4271. [Google Scholar] [CrossRef]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A Purified Membrane Protein from Akkermansia muciniphila or the Pasteurized Bacterium Improves Metabolism in Obese and Diabetic Mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Depommier, C.; Van Hul, M.; Everard, A.; Delzenne, N.M.; De Vos, W.M.; Cani, P.D. Pasteurized Akkermansia muciniphila Increases Whole-Body Energy Expenditure and Fecal Energy Excretion in Diet-Induced Obese Mice. Gut Microbes 2020, 11, 1231–1245. [Google Scholar] [CrossRef] [Green Version]
- Obanda, D.N.; Husseneder, C.; Raggio, A.M.; Page, R.; Marx, B.; Stout, R.W.; Guice, J.; Coulon, D.; Keenan, M.J. Abundance of the Species Clostridium Butyricum in the Gut Microbiota Contributes to Differences in Obesity Phenotype in Outbred Sprague-Dawley CD Rats. Nutrition 2020, 78, 110893. [Google Scholar] [CrossRef]
- Weng, H.; Endo, K.; Li, J.; Kito, N.; Iwai, N. Induction of Peroxisomes by Butyrate-Producing Probiotics. PLoS ONE 2015, 10, e0117851. [Google Scholar] [CrossRef] [Green Version]
- Shang, H.; Sun, J.; Chen, Y.Q. Clostridium Butyricum CGMCC0313.1 Modulates Lipid Profile, Insulin Resistance and Colon Homeostasis in Obese Mice. PLoS ONE 2016, 11, e0154373. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Yang, H.; Heng, C.; Wang, H.; Chen, S.; Hu, Y.; Jiang, Z.; Yu, Q.; Wang, Z.; Qian, S.; et al. Amelioration of Non-Alcoholic Fatty Liver Disease by Sodium Butyrate Is Linked to the Modulation of Intestinal Tight Junctions in Db/Db Mice. Food Funct. 2020, 11, 10675–10689. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Guo, Y.; Liu, H.; Gao, J.; Nie, W. Clostridium Butyricum Reduce Lipogenesis through Bacterial Wall Components and Butyrate. Appl. Microbiol. Biotechnol. 2014, 98, 7549–7557. [Google Scholar] [CrossRef]
- Chen, J.-C.; Lee, W.-J.; Tsou, J.-J.; Liu, T.-P.; Tsai, P.-L. Effect of Probiotics on Postoperative Quality of Gastric Bypass Surgeries: A Prospective Randomized Trial. Surg. Obes. Relat. Dis. 2016, 12, 57–61. [Google Scholar] [CrossRef]
- Shang, X.; Zhang, X.; Du, C.; Ma, Z.; Jin, S.; Ao, N.; Yang, J.; Du, J. Clostridium Butyricum Alleviates Gut Microbiota Alteration-Induced Bone Loss after Bariatric Surgery by Promoting Bone Autophagy. J. Pharmacol. Exp. Ther. 2021, 377, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Li, D.; Feng, N.; Shamoon, M.; Sun, Z.; Ding, L.; Zhang, H.; Chen, W.; Sun, J.; Chen, Y.Q. Anti-Diabetic Effects of Clostridium Butyricum CGMCC0313.1 through Promoting the Growth of Gut Butyrate-Producing Bacteria in Type 2 Diabetic Mice. Sci. Rep. 2017, 7, 7046. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Shan, K.; Pan, L.-L.; Feng, N.; Lv, Z.; Sun, Y.; Li, J.; Wu, C.; Zhang, H.; Chen, W.; et al. Clostridium Butyricum CGMCC0313.1 Protects against Autoimmune Diabetes by Modulating Intestinal Immune Homeostasis and Inducing Pancreatic Regulatory T Cells. Front. Immunol. 2017, 8, 1345. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Wang, F.; Ling, Z.; Yu, X.; Chen, W.; Li, H.; Jin, J.; Pang, M.; Zhang, H.; Yu, J.; et al. Clostridium Butyricum Attenuates Cerebral Ischemia/Reperfusion Injury in Diabetic Mice via Modulation of Gut Microbiota. Brain Res. 2016, 1642, 180–188. [Google Scholar] [CrossRef]
- Da Silva, C.C.; Monteil, M.A.; Davis, E.M. Overweight and Obesity in Children Are Associated with an Abundance of Firmicutes and Reduction of Bifidobacterium in Their Gastrointestinal Microbiota. Child. Obes. 2020, 16, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Castañeda-Márquez, A.C.; Díaz-Benítez, C.E.; Bahena-Roman, M.; Campuzano-Benítez, G.E.; Galván-Portillo, M.; Campuzano-Rincón, J.C.; Lagunas-Martínez, A.; Bermudez-Morales, V.H.; Orbe-Orihuela, Y.C.; Peralta-Romero, J.; et al. Lactobacillus paracasei as a Protective Factor of Obesity Induced by an Unhealthy Diet in Children. Obes. Res. Clin. Pract. 2020, 14, 271–278. [Google Scholar] [CrossRef]
- Carreras, N.L.; Martorell, P.; Chenoll, E.; Genovés, S.; Ramón, D.; Aleixandre, A. Anti-Obesity Properties of the Strain Bifidobacterium Animalis Subsp. Lactis CECT 8145 in Zücker Fatty Rats. Benef. Microbes 2018, 9, 629–641. [Google Scholar] [CrossRef]
- Huo, Y.; Lu, X.; Wang, X.; Wang, X.; Chen, L.; Guo, H.; Zhang, M.; Li, Y. Bifidobacterium Animalis Subsp. Lactis A6 Alleviates Obesity Associated with Promoting Mitochondrial Biogenesis and Function of Adipose Tissue in Mice. Molecules 2020, 25, 1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, X.; Zeng, X.; Tan, F.; Yi, R.; Pan, Y.; Zhou, X.; Mu, J.; Zhao, X. Lactobacillus Plantarum KFY04 Prevents Obesity in Mice through the PPAR Pathway and Alleviates Oxidative Damage and Inflammation. Food Funct. 2020, 11, 5460–5472. [Google Scholar] [CrossRef]
- Schellekens, H.; Torres-Fuentes, C.; van de Wouw, M.; Long-Smith, C.M.; Mitchell, A.; Strain, C.; Berding, K.; Bastiaanssen, T.F.S.; Rea, K.; Golubeva, A.V.; et al. Bifidobacterium Longum Counters the Effects of Obesity: Partial Successful Translation from Rodent to Human. EBioMedicine 2021, 63, 103176. [Google Scholar] [CrossRef] [PubMed]
- Bo, T.-B.; Wen, J.; Zhao, Y.-C.; Tian, S.-J.; Zhang, X.-Y.; Wang, D.-H. Bifidobacterium Pseudolongum Reduces Triglycerides by Modulating Gut Microbiota in Mice Fed High-Fat Food. J. Steroid Biochem. Mol. Biol. 2020, 198, 105602. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Tang, M.-W.; Tan, F.; Zhou, X.-R.; Fan, L.; Xie, Y.-X.; Zhao, X. Anti-Obesity Effect of Lactobacillus Plantarum CQPC01 by Modulating Lipid Metabolism in High-Fat Diet-Induced C57BL/6 Mice. J. Food Biochem. 2020, 44, e13491. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yan, H.; Lu, Y.; Li, X.; Wang, X.; Shan, Y.; Yi, Y.; Liu, B.; Zhou, Y.; Lü, X. Anti-Obesity Effect of Lactobacillus rhamnosus LS-8 and Lactobacillus crustorum MN047 on High-Fat and High-Fructose Diet Mice Base on Inflammatory Response Alleviation and Gut Microbiota Regulation. Eur. J. Nutr. 2020, 59, 2709–2728. [Google Scholar] [CrossRef]
- Won, S.-M.; Chen, S.; Lee, S.Y.; Lee, K.E.; Park, K.W.; Yoon, J.-H. Lactobacillus sakei ADM14 Induces Anti-Obesity Effects and Changes in Gut Microbiome in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 3703. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.-M.; Han, S.-K.; Kim, J.-K.; Oh, S.-J.; Jang, H.-B.; Kim, D.-H. Lactobacillus sakei Alleviates High-Fat-Diet-Induced Obesity and Anxiety in Mice by Inducing AMPK Activation and SIRT1 Expression and Inhibiting Gut Microbiota-Mediated NF-ΚB Activation. Mol. Nutr. Food Res. 2019, 63, e1800978. [Google Scholar] [CrossRef]
- Yang, G.; Hong, E.; Oh, S.; Kim, E. Non-Viable Lactobacillus Johnsonii JNU3402 Protects against Diet-Induced Obesity. Foods 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Murga, M.L.; Olivares, M.; Sanz, Y. Bifidobacterium Pseudocatenulatum CECT 7765 Reverses the Adverse Effects of Diet-Induced Obesity through the Gut-Bone Axis. Bone 2020, 141, 115580. [Google Scholar] [CrossRef]
- Zhao, D.; Zhu, H.; Gao, F.; Qian, Z.; Mao, W.; Yin, Y.; Tan, J.; Chen, D. Antidiabetic Effects of Selenium-Enriched Bifidobacterium Longum DD98 in Type 2 Diabetes Model of Mice. Food Funct. 2020, 11, 6528–6541. [Google Scholar] [CrossRef] [PubMed]
- Ben Othman, M.; Sakamoto, K. Effect of Inactivated Bifidobacterium Longum Intake on Obese Diabetes Model Mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Yuan, Q.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative Effects of Probiotic Lactobacillus paracasei NL41 on Insulin Sensitivity, Oxidative Stress, and Beta-Cell Function in a Type 2 Diabetes Mellitus Rat Model. Mol. Nutr. Food Res. 2019, 63, e1900457. [Google Scholar] [CrossRef] [PubMed]
- Mobini, R.; Tremaroli, V.; Ståhlman, M.; Karlsson, F.; Levin, M.; Ljungberg, M.; Sohlin, M.; Bertéus Forslund, H.; Perkins, R.; Bäckhed, F.; et al. Metabolic Effects of Lactobacillus reuteri DSM 17938 in People with Type 2 Diabetes: A Randomized Controlled Trial. Diabetes Obes. Metab. 2017, 19, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, M.-C.; Tsai, W.-H.; Jheng, Y.-P.; Su, S.-L.; Wang, S.-Y.; Lin, C.-C.; Chen, Y.-H.; Chang, W.-W. The Beneficial Effects of Lactobacillus reuteri ADR-1 or ADR-3 Consumption on Type 2 Diabetes Mellitus: A Randomized, Double-Blinded, Placebo-Controlled Trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef]
- Yan, F.; Li, N.; Shi, J.; Li, H.; Yue, Y.; Jiao, W.; Wang, N.; Song, Y.; Huo, G.; Li, B. Lactobacillus acidophilus Alleviates Type 2 Diabetes by Regulating Hepatic Glucose, Lipid Metabolism and Gut Microbiota in Mice. Food Funct. 2019, 10, 5804–5815. [Google Scholar] [CrossRef]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Clarke, J.; Shawron, K.; Keshavarzian, A.; Walter, J. Probiotic Bifidobacterium Strains and Galactooligosaccharides Improve Intestinal Barrier Function in Obese Adults but Show No Synergism When Used Together as Synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef]
- Groele, L.; Szajewska, H.; Szypowska, A. Effects of Lactobacillus rhamnosus GG and Bifidobacterium Lactis Bb12 on Beta-Cell Function in Children with Newly Diagnosed Type 1 Diabetes: Protocol of a Randomised Controlled Trial. BMJ Open 2017, 7, e017178. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.; Park, D.-S.; Rahman, M.S.; Ki, S.-J.; Lee, Y.R.; Imran, K.M.; Yoon, D.; Heo, J.; Lee, T.-J.; Kim, Y.-S. Bifidobacterium Longum DS0956 and Lactobacillus rhamnosus DS0508 Culture-Supernatant Ameliorate Obesity by Inducing Thermogenesis in Obese-Mice. Benef. Microbes 2020, 11, 361–373. [Google Scholar] [CrossRef]
- Chiang, C.-J.; Huang, P.-H. Metabolic Engineering of Probiotic Escherichia coli for Cytolytic Therapy of Tumors. Sci. Rep. 2021, 11, 5853. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Yang, H.; Liu, F.; Chen, Y.; Tang, S.; Ji, W.; Tang, J.; Liu, Z.; Sun, Y.; Hu, S.; et al. Escherichia coli Nissle 1917 Engineered to Express Tum-5 Can Restrain Murine Melanoma Growth. Oncotarget 2017, 8, 85772–85782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, Y.; Xia, L.; Zhang, X.; Ding, X.; Yan, F.; Wu, F. Escherichia coli Nissle 1917 Targets and Restrains Mouse B16 Melanoma and 4T1 Breast Tumors through Expression of Azurin Protein. Appl. Environ. Microbiol. 2012, 78, 7603–7610. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.-J.; Hong, Y.-H. In Situ Delivery of Biobutyrate by Probiotic Escherichia coli for Cancer Therapy. Sci. Rep. 2021, 11, 18172. [Google Scholar] [CrossRef]
- Wang, L.; Liao, Y.; Yang, R.; Zhu, Z.; Zhang, L.; Wu, Z.; Sun, X. An Engineered Probiotic Secreting Sj16 Ameliorates Colitis via Ruminococcaceae/Butyrate/Retinoic Acid Axis. Bioeng. Transl. Med. 2021, 6, e10219. [Google Scholar] [CrossRef]
- Yan, X.; Liu, X.-Y.; Zhang, D.; Zhang, Y.-D.; Li, Z.-H.; Liu, X.; Wu, F.; Chen, G.-Q. Construction of a Sustainable 3-Hydroxybutyrate-Producing Probiotic Escherichia coli for Treatment of Colitis. Cell Mol. Immunol. 2021, 18, 2344–2357. [Google Scholar] [CrossRef]
- Cui, M.; Sun, T.; Li, S.; Pan, H.; Liu, J.; Zhang, X.; Li, L.; Li, S.; Wei, C.; Yu, C.; et al. NIR Light-Responsive Bacteria with Live Bio-Glue Coatings for Precise Colonization in the Gut. Cell Rep. 2021, 36, 109690. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Pang, G.; Zhang, T.; Sun, T.; Zhang, L.; Kang, R.; Xue, X.; Pan, H.; Yang, C.; Zhang, X.; et al. Optotheranostic Nanosystem with Phone Visual Diagnosis and Optogenetic Microbial Therapy for Ulcerative Colitis At-Home Care. ACS Nano 2021, 15, 7040–7052. [Google Scholar] [CrossRef]
- Ma, J.; Li, C.; Wang, J.; Gu, J. Genetically Engineered Escherichia coli Nissle 1917 Secreting GLP-1 Analog Exhibits Potential Antiobesity Effect in High-Fat Diet-Induced Obesity Mice. Obesity 2020, 28, 315–322. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Chen, Z.; Guo, Y.; McMillan, C.; Flynn, C.R.; Davies, S.S. Two-Week Administration of Engineered Escherichia coli Establishes Persistent Resistance to Diet-Induced Obesity Even without Antibiotic Pre-Treatment. Appl. Microbiol. Biotechnol. 2019, 103, 6711–6723. [Google Scholar] [CrossRef]
- Geldart, K.G.; Kommineni, S.; Forbes, M.; Hayward, M.; Dunny, G.M.; Salzman, N.H.; Kaznessis, Y.N. Engineered E. Coli Nissle 1917 for the Reduction of Vancomycin-Resistant Enterococcus in the Intestinal Tract. Bioeng. Transl. Med. 2018, 3, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Hwang, I.Y.; Koh, E.; Wong, A.; March, J.C.; Bentley, W.E.; Lee, Y.S.; Chang, M.W. Engineered Probiotic Escherichia coli Can Eliminate and Prevent Pseudomonas Aeruginosa Gut Infection in Animal Models. Nat. Commun. 2017, 8, 15028. [Google Scholar] [CrossRef] [Green Version]
- May-Zhang, L.S.; Chen, Z.; Dosoky, N.S.; Yancey, P.G.; Boyd, K.L.; Hasty, A.H.; Linton, M.F.; Davies, S.S. Administration of N-Acyl-Phosphatidylethanolamine Expressing Bacteria to Low Density Lipoprotein Receptor-/- Mice Improves Indices of Cardiometabolic Disease. Sci. Rep. 2019, 9, 420. [Google Scholar] [CrossRef] [Green Version]
- Raghuvanshi, R.; Chaudhari, A.; Kumar, G.N. Amelioration of Cadmium- and Mercury-Induced Liver and Kidney Damage in Rats by Genetically Engineered Probiotic Escherichia coli Nissle 1917 Producing Pyrroloquinoline Quinone with Oral Supplementation of Citric Acid. Nutrition 2016, 32, 1285–1294. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.S.; Raghuvanshi, R.; Kumar, G.N. Genetically Engineered Escherichia coli Nissle 1917 Synbiotic Counters Fructose-Induced Metabolic Syndrome and Iron Deficiency. Appl. Microbiol. Biotechnol. 2017, 101, 4713–4723. [Google Scholar] [CrossRef]
- Somabhai, C.A.; Raghuvanshi, R.; Nareshkumar, G. Genetically Engineered Escherichia coli Nissle 1917 Synbiotics Reduce Metabolic Effects Induced by Chronic Consumption of Dietary Fructose. PLoS ONE 2016, 11, e0164860. [Google Scholar] [CrossRef]
- Kurtz, C.B.; Millet, Y.A.; Puurunen, M.K.; Perreault, M.; Charbonneau, M.R.; Isabella, V.M.; Kotula, J.W.; Antipov, E.; Dagon, Y.; Denney, W.S.; et al. An Engineered E. Coli Nissle Improves Hyperammonemia and Survival in Mice and Shows Dose-Dependent Exposure in Healthy Humans. Sci. Transl. Med. 2019, 11, eaau7975. [Google Scholar] [CrossRef] [PubMed]
- Puurunen, M.K.; Vockley, J.; Searle, S.L.; Sacharow, S.J.; Phillips, J.A.; Denney, W.S.; Goodlett, B.D.; Wagner, D.A.; Blankstein, L.; Castillo, M.J.; et al. Safety and Pharmacodynamics of an Engineered E. Coli Nissle for the Treatment of Phenylketonuria: A First-in-Human Phase 1/2a Study. Nat. Metab. 2021, 3, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, X.; Liu, Y.; Su, T.; Lin, C.; Shao, L.; Li, L.; Li, W.; Niu, G.; Yu, J.; et al. Engineering a Probiotic Strain of Escherichia coli to Induce the Regression of Colorectal Cancer through Production of 5-Aminolevulinic Acid. Microb. Biotechnol. 2021, 14, 2130–2139. [Google Scholar] [CrossRef]
- Huang, C.; Wang, F.-B.; Liu, L.; Jiang, W.; Liu, W.; Ma, W.; Zhao, H. Hypoxic Tumor Radiosensitization Using Engineered Probiotics. Adv. Healthc. Mater. 2021, 10, e2002207. [Google Scholar] [CrossRef] [PubMed]
- Canale, F.P.; Basso, C.; Antonini, G.; Perotti, M.; Li, N.; Sokolovska, A.; Neumann, J.; James, M.J.; Geiger, S.; Jin, W.; et al. Metabolic Modulation of Tumours with Engineered Bacteria for Immunotherapy. Nature 2021, 598, 662–666. [Google Scholar] [CrossRef]
- Duan, F.; March, J.C. Interrupting Vibrio Cholerae Infection of Human Epithelial Cells with Engineered Commensal Bacterial Signaling. Biotechnol. Bioeng. 2008, 101, 128–134. [Google Scholar] [CrossRef]
- Duan, F.; March, J.C. Engineered Bacterial Communication Prevents Vibrio Cholerae Virulence in an Infant Mouse Model. Proc. Natl. Acad. Sci. USA 2010, 107, 11260–11264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, S.; Chen, M.; Song, X.; Zhang, Z.; Zhang, Z.; Chen, Z.; Li, X. Bacterial Microbots for Acid-Labile Release of Hybrid Micelles to Promote the Synergistic Antitumor Efficacy. Acta Biomater. 2018, 78, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhao, L.; Song, X.; Tang, M.; Mo, C.; Li, X. Doxorubicin-Conjugated Escherichia coli Nissle 1917 Swimmers to Achieve Tumor Targeting and Responsive Drug Release. J. Control. Release 2017, 268, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Praveschotinunt, P.; Duraj-Thatte, A.M.; Gelfat, I.; Bahl, F.; Chou, D.B.; Joshi, N.S. Engineered E. Coli Nissle 1917 for the Delivery of Matrix-Tethered Therapeutic Domains to the Gut. Nat. Commun. 2019, 10, 5580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, A.; Gelfat, I.; Emani, S.; Praveschotinunt, P.; Joshi, N.S. Plasmid Vectors for in Vivo Selection-Free Use with the Probiotic E. Coli Nissle 1917. ACS Synth. Biol. 2021, 10, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Kraśko, J.A.; Žilionytė, K.; Darinskas, A.; Strioga, M.; Rjabceva, S.; Zalutsky, I.; Derevyanko, M.; Kulchitsky, V.; Lubitz, W.; Kudela, P.; et al. Bacterial Ghosts as Adjuvants in Syngeneic Tumour Cell Lysate-Based Anticancer Vaccination in a Murine Lung Carcinoma Model. Oncol. Rep. 2017, 37, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Hao, L.; Liu, X.; Borrás-Hidalgo, O.; Zhang, Y. Enhanced Anti-Proliferative Efficacy of Epothilone B Loaded with Escherichia coli Nissle 1917 Bacterial Ghosts on the HeLa Cells by Mitochondrial Pathway of Apoptosis. Drug Dev. Ind. Pharm. 2018, 44, 1328–1335. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhang, P.; Zhang, Z.; Liu, Y.; Chen, M.; Li, S.; Li, X. Bacterial Navigation for Tumor Targeting and Photothermally-Triggered Bacterial Ghost Transformation for Spatiotemporal Drug Release. Acta Biomater. 2021, 131, 172–184. [Google Scholar] [CrossRef]
- Montanaro, J.; Inic-Kanada, A.; Ladurner, A.; Stein, E.; Belij, S.; Bintner, N.; Schlacher, S.; Schuerer, N.; Mayr, U.B.; Lubitz, W.; et al. Escherichia coli Nissle 1917 Bacterial Ghosts Retain Crucial Surface Properties and Express Chlamydial Antigen: An Imaging Study of a Delivery System for the Ocular Surface. Drug Des. Dev. Ther. 2015, 9, 3741–3754. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, J.A.; Huang, C.-J.; Doody, A.M.; Leung, T.; Mineta, K.; Feng, D.D.; Wayne, E.C.; Nishimura, N.; Leifer, C.; DeLisa, M.P.; et al. Mechanistic Insight into the TH1-Biased Immune Response to Recombinant Subunit Vaccines Delivered by Probiotic Bacteria-Derived Outer Membrane Vesicles. PLoS ONE 2014, 9, e112802. [Google Scholar] [CrossRef] [PubMed]
- Ou, B.; Jiang, B.; Jin, D.; Yang, Y.; Zhang, M.; Zhang, D.; Zhao, H.; Xu, M.; Song, H.; Wu, W.; et al. Engineered Recombinant Escherichia coli Probiotic Strains Integrated with F4 and F18 Fimbriae Cluster Genes in the Chromosome and Their Assessment of Immunogenic Efficacy in Vivo. ACS Synth. Biol. 2020, 9, 412–426. [Google Scholar] [CrossRef]
- Zhang, Y.; Ji, W.; He, L.; Chen, Y.; Ding, X.; Sun, Y.; Hu, S.; Yang, H.; Huang, W.; Zhang, Y.; et al. E. Coli Nissle 1917-Derived Minicells for Targeted Delivery of Chemotherapeutic Drug to Hypoxic Regions for Cancer Therapy. Theranostics 2018, 8, 1690–1705. [Google Scholar] [CrossRef]
- Geiger, R.; Rieckmann, J.C.; Wolf, T.; Basso, C.; Feng, Y.; Fuhrer, T.; Kogadeeva, M.; Picotti, P.; Meissner, F.; Mann, M.; et al. L-Arginine Modulates T Cell Metabolism and Enhances Survival and Anti-Tumor Activity. Cell 2016, 167, 829–842.e13. [Google Scholar] [CrossRef] [Green Version]
- Sturm, A.; Rilling, K.; Baumgart, D.C.; Gargas, K.; Abou-Ghazalé, T.; Raupach, B.; Eckert, J.; Schumann, R.R.; Enders, C.; Sonnenborn, U.; et al. Escherichia coli Nissle 1917 Distinctively Modulates T-Cell Cycling and Expansion via Toll-like Receptor 2 Signaling. Infect. Immun. 2005, 73, 1452–1465. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.K.; Liu, Q.; Liang, K.; Li, P.; Kong, Q. Bacteria-Derived Minicells for Cancer Therapy. Cancer Lett. 2020, 491, 11–21. [Google Scholar] [CrossRef] [PubMed]
- van Passel, M.W.J.; Kant, R.; Zoetendal, E.G.; Plugge, C.M.; Derrien, M.; Malfatti, S.A.; Chain, P.S.G.; Woyke, T.; Palva, A.; de Vos, W.M.; et al. The Genome of Akkermansia muciniphila, a Dedicated Intestinal Mucin Degrader, and Its Use in Exploring Intestinal Metagenomes. PLoS ONE 2011, 6, e16876. [Google Scholar] [CrossRef] [Green Version]
- Ashrafian, F.; Shahriary, A.; Behrouzi, A.; Moradi, H.R.; Keshavarz Azizi Raftar, S.; Lari, A.; Hadifar, S.; Yaghoubfar, R.; Ahmadi Badi, S.; Khatami, S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles as a Mucosal Delivery Vector for Amelioration of Obesity in Mice. Front. Microbiol. 2019, 10, 2155. [Google Scholar] [CrossRef]
- Chelakkot, C.; Choi, Y.; Kim, D.-K.; Park, H.T.; Ghim, J.; Kwon, Y.; Jeon, J.; Kim, M.-S.; Jee, Y.-K.; Gho, Y.S.; et al. Akkermansia muciniphila-Derived Extracellular Vesicles Influence Gut Permeability through the Regulation of Tight Junctions. Exp. Mol. Med. 2018, 50, e450. [Google Scholar] [CrossRef] [PubMed]
- Ashrafian, F.; Behrouzi, A.; Shahriary, A.; Ahmadi Badi, S.; Davari, M.; Khatami, S.; Rahimi Jamnani, F.; Fateh, A.; Vaziri, F.; Siadat, S.D. Comparative Study of Effect of Akkermansia muciniphila and Its Extracellular Vesicles on Toll-like Receptors and Tight Junction. Gastroenterol. Hepatol. Bed Bench 2019, 12, 163–168. [Google Scholar] [PubMed]
- Kang, C.-S.; Ban, M.; Choi, E.-J.; Moon, H.-G.; Jeon, J.-S.; Kim, D.-K.; Park, S.-K.; Jeon, S.G.; Roh, T.-Y.; Myung, S.-J.; et al. Extracellular Vesicles Derived from Gut Microbiota, Especially Akkermansia muciniphila, Protect the Progression of Dextran Sulfate Sodium-Induced Colitis. PLoS ONE 2013, 8, e76520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keshavarz Azizi Raftar, S.; Ashrafian, F.; Yadegar, A.; Lari, A.; Moradi, H.R.; Shahriary, A.; Azimirad, M.; Alavifard, H.; Mohsenifar, Z.; Davari, M.; et al. The Protective Effects of Live and Pasteurized Akkermansia muciniphila and Its Extracellular Vesicles against HFD/CCl4-Induced Liver Injury. Microbiol. Spectr. 2021, 9, e0048421. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, D.; Zhuo, J.; Lin, Z.; Yang, M.; Xu, X. The Gut-Liver Axis in Immune Remodeling: New Insight into Liver Diseases. Int. J. Biol. Sci. 2020, 16, 2357–2366. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubfar, R.; Behrouzi, A.; Zare Banadkoki, E.; Ashrafian, F.; Lari, A.; Vaziri, F.; Nojoumi, S.A.; Fateh, A.; Khatami, S.; Siadat, S.D. Effect of Akkermansia muciniphila, Faecalibacterium Prausnitzii, and Their Extracellular Vesicles on the Serotonin System in Intestinal Epithelial Cells. Probiotics Antimicrob. Proteins 2021, 13, 1546–1556. [Google Scholar] [CrossRef] [PubMed]
- Yaghoubfar, R.; Behrouzi, A.; Ashrafian, F.; Shahryari, A.; Moradi, H.R.; Choopani, S.; Hadifar, S.; Vaziri, F.; Nojoumi, S.A.; Fateh, A.; et al. Modulation of Serotonin Signaling/Metabolism by Akkermansia muciniphila and Its Extracellular Vesicles through the Gut-Brain Axis in Mice. Sci. Rep. 2020, 10, 22119. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-H.; Chen, C.-Y.; Liu, Z.-Z.; Luo, Z.-W.; Rao, S.-S.; Jin, L.; Wan, T.-F.; Yue, T.; Tan, Y.-J.; Yin, H.; et al. Extracellular Vesicles from Child Gut Microbiota Enter into Bone to Preserve Bone Mass and Strength. Adv. Sci. 2021, 8, 2004831. [Google Scholar] [CrossRef]
- Morishita, M.; Horita, M.; Higuchi, A.; Marui, M.; Katsumi, H.; Yamamoto, A. Characterizing Different Probiotic-Derived Extracellular Vesicles as a Novel Adjuvant for Immunotherapy. Mol. Pharm. 2021, 18, 1080–1092. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.; Zhao, Z.; Liang, Q.; Shen, H.; Zhao, Z.; Chen, Z.; He, R.; Feng, S.; Cao, D.; Gan, G.; et al. Overexpression of PEGF Improved the Gut Protective Function of Clostridium Butyricum Partly through STAT3 Signal Pathway. Appl. Microbiol. Biotechnol. 2021, 105, 5973–5991. [Google Scholar] [CrossRef]
- Liu, M.; Li, S.; Zhang, Q.; Xu, Z.; Wang, J.; Sun, H. Oral Engineered Bifidobacterium Longum Expressing RhMnSOD to Suppress Experimental Colitis. Int. Immunopharmacol. 2018, 57, 25–32. [Google Scholar] [CrossRef]
- Wei, P.; Yang, Y.; Ding, Q.; Li, X.; Sun, H.; Liu, Z.; Huang, J.; Gong, Y. Oral Delivery of Bifidobacterium Longum Expressing α-Melanocyte-Stimulating Hormone to Combat Ulcerative Colitis. J. Med. Microbiol. 2016, 65, 160–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Huang, Z.; Sao, C.; Huang, Y.; Zhang, F.; Yang, J.; Lian, J.; Zeng, Z.; Luo, W.; Zeng, W.; et al. Bifidobacterium as an Oral Delivery Carrier of Interleukin-12 for the Treatment of Coxsackie Virus B3-Induced Myocarditis in the Balb/c Mice. Int. Immunopharmacol. 2012, 12, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Long, R.T.; Zeng, W.S.; Chen, L.Y.; Guo, J.; Lin, Y.Z.; Huang, Q.S.; Luo, S.Q. Bifidobacterium as an Oral Delivery Carrier of Oxyntomodulin for Obesity Therapy: Inhibitory Effects on Food Intake and Body Weight in Overweight Mice. Int. J. Obes. 2010, 34, 712–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, P.; Yang, Y.; Li, T.; Ding, Q.; Sun, H. A Engineered Bifidobacterium Longum Secreting a Bioative Penetratin-Glucagon-like Peptide 1 Fusion Protein Enhances Glucagon-like Peptide 1 Absorption in the Intestine. J. Microbiol. Biotechnol. 2015, in press. [Google Scholar]
- Oh, J.-H.; Schueler, K.L.; Stapleton, D.S.; Alexander, L.M.; Yen, C.-L.E.; Keller, M.P.; Attie, A.D.; van Pijkeren, J.-P. Secretion of Recombinant Interleukin-22 by Engineered Lactobacillus reuteri Reduces Fatty Liver Disease in a Mouse Model of Diet-Induced Obesity. mSphere 2020, 5, e00183-20. [Google Scholar] [CrossRef]
- Hendrikx, T.; Duan, Y.; Wang, Y.; Oh, J.-H.; Alexander, L.M.; Huang, W.; Stärkel, P.; Ho, S.B.; Gao, B.; Fiehn, O.; et al. Bacteria Engineered to Produce IL-22 in Intestine Induce Expression of REG3G to Reduce Ethanol-Induced Liver Disease in Mice. Gut 2019, 68, 1504–1515. [Google Scholar] [CrossRef]
- Fang, X.; Zhou, X.; Miao, Y.; Han, Y.; Wei, J.; Chen, T. Therapeutic Effect of GLP-1 Engineered Strain on Mice Model of Alzheimer’s Disease and Parkinson’s Disease. AMB Express 2020, 10, 80. [Google Scholar] [CrossRef]
- Buford, T.W.; Sun, Y.; Roberts, L.M.; Banerjee, A.; Peramsetty, S.; Knighton, A.; Verma, A.; Morgan, D.; Torres, G.E.; Li, Q.; et al. Angiotensin (1-7) Delivered Orally via Probiotic, but Not Subcutaneously, Benefits the Gut-Brain Axis in Older Rats. Geroscience 2020, 42, 1307–1321. [Google Scholar] [CrossRef]
- Esposito, G.; Pesce, M.; Seguella, L.; Lu, J.; Corpetti, C.; Del Re, A.; De Palma, F.D.E.; Esposito, G.; Sanseverino, W.; Sarnelli, G. Engineered Lactobacillus paracasei Producing Palmitoylethanolamide (PEA) Prevents Colitis in Mice. Int. J. Mol. Sci. 2021, 22, 2945. [Google Scholar] [CrossRef]
- Jia, S.; Huang, X.; Li, H.; Zheng, D.; Wang, L.; Qiao, X.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Immunogenicity Evaluation of Recombinant Lactobacillus casei W56 Expressing Bovine Viral Diarrhea Virus E2 Protein in Conjunction with Cholera Toxin B Subunit as an Adjuvant. Microb. Cell Fact. 2020, 19, 186. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Wang, L.; Ma, S.; Wang, X.; Wang, Y.; Xiao, Y.; Jiang, Y.; Qiao, X.; Tang, L.; Xu, Y.; et al. Immunogenicity of EGFP-Marked Recombinant Lactobacillus casei against Transmissible Gastroenteritis Virus and Porcine Epidemic Diarrhea Virus. Viruses 2017, 9, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Wang, L.; Huang, X.; Wang, X.; Chen, S.; Shi, W.; Qiao, X.; Jiang, Y.; Tang, L.; Xu, Y.; et al. Oral Recombinant Lactobacillus vaccine Targeting the Intestinal Microfold Cells and Dendritic Cells for Delivering the Core Neutralizing Epitope of Porcine Epidemic Diarrhea Virus. Microb. Cell Fact. 2018, 17, 20. [Google Scholar] [CrossRef]
- Hou, X.; Jiang, X.; Jiang, Y.; Tang, L.; Xu, Y.; Qiao, X.; Min, L.; Wen, C.; Ma, G.; Li, Y. Oral Immunization against PEDV with Recombinant Lactobacillus casei Expressing Dendritic Cell-Targeting Peptide Fusing COE Protein of PEDV in Piglets. Viruses 2018, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Duan, K.; Hua, X.; Wang, Y.; Wang, Y.; Chen, Y.; Shi, W.; Tang, L.; Li, Y.; Liu, M. Oral Immunization with a Recombinant Lactobacillus Expressing CK6 Fused with VP2 Protein against IPNV in Rainbow Trout (Oncorhynchus Mykiss). Fish Shellfish Immunol. 2018, 83, 223–231. [Google Scholar] [CrossRef]
- Chen, Y.; Hua, X.; Ren, X.; Duan, K.; Gao, S.; Sun, J.; Feng, Y.; Zhou, Y.; Guan, X.; Li, D.; et al. Oral Immunization with Recombinant Lactobacillus casei Displayed AHA1-CK6 and VP2 Induces Protection against Infectious Pancreatic Necrosis in Rainbow Trout (Oncorhynchus Mykiss). Fish Shellfish Immunol. 2020, 100, 18–26. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Y.; Wang, Z.; Bai, J.; Jia, S.; Feng, B.; Jiang, Y.; Cui, W.; Tang, L.; Li, Y.; et al. Oral Immunization of Mice with a Probiotic Lactobacillus casei Constitutively Expressing the α-Toxoid Induces Protective Immunity against Clostridium Perfringens α-Toxin. Virulence 2019, 10, 166–179. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Qi, R.; Chen, C.; Yin, J.; Ma, S.; Shi, W.; Wu, Y.; Ge, J.; Jiang, Y.; Tang, L.; et al. Immunogenicity of Recombinant Lactobacillus casei-Expressing F4 (K88) Fimbrial Adhesin FaeG in Conjunction with a Heat-Labile Enterotoxin A (LTAK63) and Heat-Labile Enterotoxin B (LTB) of Enterotoxigenic Escherichia coli as an Oral Adjuvant in Mice. J. Appl. Microbiol. 2017, 122, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Yue, J.; Deng, Y.; Yang, S.; Zhu, L.; Xu, Z. Immunogenicity of engineered Lactobacillus plantarum expressing porcine epidemic diarrhea virus S1 gene. Chin. J. Biotechnol. 2021, 37, 2779–2785. [Google Scholar] [CrossRef]
- Pan, N.; Liu, B.; Bao, X.; Zhang, H.; Sheng, S.; Liang, Y.; Pan, H.; Wang, X. Oral Delivery of Novel Recombinant Lactobacillus Elicit High Protection against Staphylococcus Aureus Pulmonary and Skin Infections. Vaccines 2021, 9, 984. [Google Scholar] [CrossRef]
- Wei, W.; Wiggins, J.; Hu, D.; Vrbanac, V.; Bowder, D.; Mellon, M.; Tager, A.; Sodroski, J.; Xiang, S.-H. Blocking HIV-1 Infection by Chromosomal Integrative Expression of Human CD4 on the Surface of Lactobacillus acidophilus ATCC 4356. J. Virol. 2019, 93, e01830-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, G.; Jin, B.; Monis, P.; Saint, C. A Genetic and Metabolic Approach to Redirection of Biochemical Pathways of Clostridium Butyricum for Enhancing Hydrogen Production. Biotechnol. Bioeng. 2013, 110, 338–342. [Google Scholar] [CrossRef]
- Cai, G.; Jin, B.; Saint, C.; Monis, P. Genetic Manipulation of Butyrate Formation Pathways in Clostridium Butyricum. J. Biotechnol. 2011, 155, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yun, J.; Zhang, H.; Magocha, T.A.; Zabed, H.; Xue, Y.; Fokum, E.; Sun, W.; Qi, X. Genetically Engineered Strains: Application and Advances for 1,3-Propanediol Production from Glycerol. Food Technol. Biotechnol. 2018, 56, 3–15. [Google Scholar] [CrossRef]
- Kuehne, S.A.; Minton, N.P. ClosTron-Mediated Engineering of Clostridium. Bioengineered 2012, 3, 247–254. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Du, Y.; Jiang, W.; Chang, W.-L.; Yang, S.-T.; Tang, I.-C. Effects of Different Replicons in Conjugative Plasmids on Transformation Efficiency, Plasmid Stability, Gene Expression and n-Butanol Biosynthesis in Clostridium Tyrobutyricum. Appl. Microbiol. Biotechnol. 2012, 93, 881–889. [Google Scholar] [CrossRef]
- Heap, J.T.; Pennington, O.J.; Cartman, S.T.; Carter, G.P.; Minton, N.P. The ClosTron: A Universal Gene Knock-out System for the Genus Clostridium. J. Microbiol. Methods 2007, 70, 452–464. [Google Scholar] [CrossRef]
- Ng, Y.K.; Ehsaan, M.; Philip, S.; Collery, M.M.; Janoir, C.; Collignon, A.; Cartman, S.T.; Minton, N.P. Expanding the Repertoire of Gene Tools for Precise Manipulation of the Clostridium Difficile Genome: Allelic Exchange Using PyrE Alleles. PLoS ONE 2013, 8, e56051. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Luo, H.; Wang, Y.; Wang, Y.; Tu, T.; Qin, X.; Su, X.; Bai, Y.; Yao, B.; et al. Exploiting Heterologous and Endogenous CRISPR-Cas Systems for Genome Editing in the Probiotic Clostridium Butyricum. Biotechnol. Bioeng. 2021, 118, 2448–2459. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shimizu, H.; Akiyama, Y.; Taniguchi, S. In Situ Delivery and Production System of Trastuzumab ScFv with Bifidobacterium. Biochem. Biophys. Res. Commun. 2017, 493, 306–312. [Google Scholar] [CrossRef]
- Wei, C.; Xun, A.Y.; Wei, X.X.; Yao, J.; Wang, J.Y.; Shi, R.Y.; Yang, G.H.; Li, Y.X.; Xu, Z.L.; Lai, M.G.; et al. Bifidobacteria Expressing Tumstatin Protein for Antitumor Therapy in Tumor-Bearing Mice. Technol. Cancer Res. Treat. 2016, 15, 498–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leite, A.Z.; Rodrigues, N.d.C.; Gonzaga, M.I.; Paiolo, J.C.C.; de Souza, C.A.; Stefanutto, N.A.V.; Omori, W.P.; Pinheiro, D.G.; Brisotti, J.L.; Matheucci Junior, E.; et al. Detection of Increased Plasma Interleukin-6 Levels and Prevalence of Prevotella Copri and Bacteroides Vulgatus in the Feces of Type 2 Diabetes Patients. Front. Immunol. 2017, 8, 1107. [Google Scholar] [CrossRef]
- Ling, Z.; Liu, F.; Shao, L.; Cheng, Y.; Li, L. Dysbiosis of the Urinary Microbiota Associated with Urine Levels of Proinflammatory Chemokine Interleukin-8 in Female Type 2 Diabetic Patients. Front. Immunol. 2017, 8, 1032. [Google Scholar] [CrossRef] [Green Version]
- Tang-Fichaux, M.; Chagneau, C.V.; Bossuet-Greif, N.; Nougayrède, J.-P.; Oswald, É.; Branchu, P. The Polyphosphate Kinase of Escherichia coli Is Required for Full Production of the Genotoxin Colibactin. mSphere 2020, 5, e01195-20. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A Metagenome-Wide Association Study of Gut Microbiota in Type 2 Diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Seregin, S.S.; Golovchenko, N.; Schaf, B.; Chen, J.; Pudlo, N.A.; Mitchell, J.; Baxter, N.T.; Zhao, L.; Schloss, P.D.; Martens, E.C.; et al. NLRP6 Protects Il10-/- Mice from Colitis by Limiting Colonization of Akkermansia muciniphila. Cell Rep. 2017, 19, 733–745. [Google Scholar] [CrossRef]
- Ganesh, B.P.; Klopfleisch, R.; Loh, G.; Blaut, M. Commensal Akkermansia muciniphila Exacerbates Gut Inflammation in Salmonella Typhimurium-Infected Gnotobiotic Mice. PLoS ONE 2013, 8, e74963. [Google Scholar] [CrossRef]
- Ijssennagger, N.; Belzer, C.; Hooiveld, G.J.; Dekker, J.; van Mil, S.W.C.; Müller, M.; Kleerebezem, M.; van der Meer, R. Gut Microbiota Facilitates Dietary Heme-Induced Epithelial Hyperproliferation by Opening the Mucus Barrier in Colon. Proc. Natl. Acad. Sci. USA 2015, 112, 10038–10043. [Google Scholar] [CrossRef] [Green Version]
- Ghoddusi, H.B.; Sherburn, R. Preliminary Study on the Isolation of Clostridium Butyricum Strains from Natural Sources in the UK and Screening the Isolates for Presence of the Type E Botulinal Toxin Gene. Int. J. Food Microbiol. 2010, 142, 202–206. [Google Scholar] [CrossRef]
- Abe, Y.; Negasawa, T.; Monma, C.; Oka, A. Infantile Botulism Caused by Clostridium Butyricum Type E Toxin. Pediatr. Neurol. 2008, 38, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Shimura, M.; Mizuma, M.; Nakagawa, K.; Aoki, S.; Miura, T.; Takadate, T.; Ariake, K.; Maeda, S.; Kawaguchi, K.; Masuda, K.; et al. Probiotic-Related Bacteremia after Major Hepatectomy for Biliary Cancer: A Report of Two Cases. Surg. Case Rep. 2021, 7, 133. [Google Scholar] [CrossRef] [PubMed]
- Mays, Z.J.S.; Chappell, T.C.; Nair, N.U. Quantifying and Engineering Mucus Adhesion of Probiotics. ACS Synth. Biol. 2020, 9, 356–367. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Ortega Moreno, L.; Chaparro, M.; Gisbert, J.P. Gut Microbiota and Dietary Factors as Modulators of the Mucus Layer in Inflammatory Bowel Disease. Int. J. Mol. Sci. 2021, 22, 10224. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of native probiotics in diseases. Escherichia coli Nissle 1917, Akkermansia muciniphila, Clostridium butyricum, lactic acid bacteria, and Bifidobacterium spp. have been proven to be promising therapeutic agents in cancer, IBD, and obesity.
Figure 1.
Effects of native probiotics in diseases. Escherichia coli Nissle 1917, Akkermansia muciniphila, Clostridium butyricum, lactic acid bacteria, and Bifidobacterium spp. have been proven to be promising therapeutic agents in cancer, IBD, and obesity.

Figure 2.
Strategies of modified EcN in diseases. There are three main strategies for modified EcN: 1. Expressing direct therapeutic factors. 2. Using the expressed therapeutic factors to complement other therapies. 3. Using EcN proper or its derivatives as carriers, relying on targeting or immune activity to transport the therapeutic factors.
Figure 2.
Strategies of modified EcN in diseases. There are three main strategies for modified EcN: 1. Expressing direct therapeutic factors. 2. Using the expressed therapeutic factors to complement other therapies. 3. Using EcN proper or its derivatives as carriers, relying on targeting or immune activity to transport the therapeutic factors.


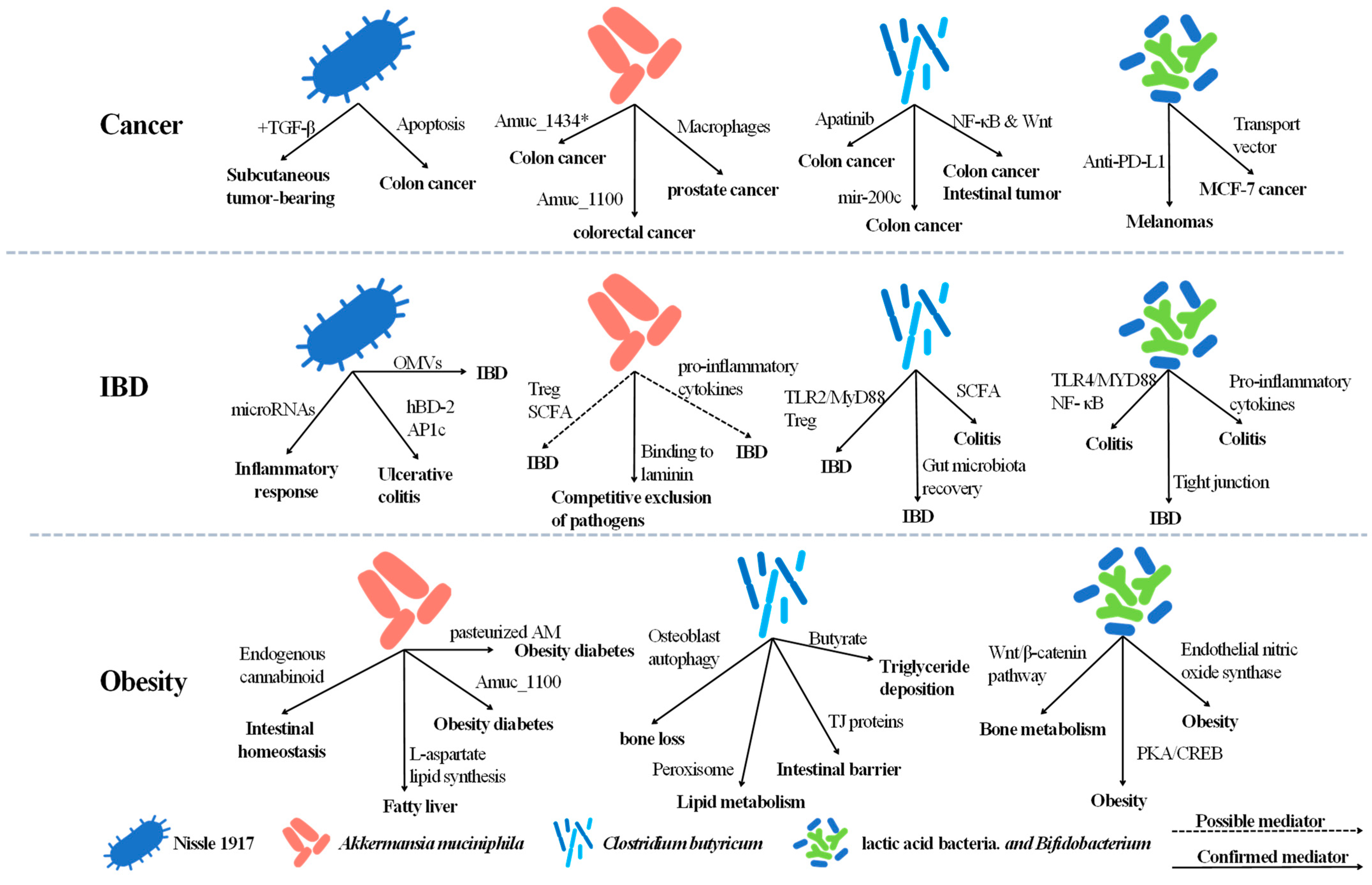{kind=link}
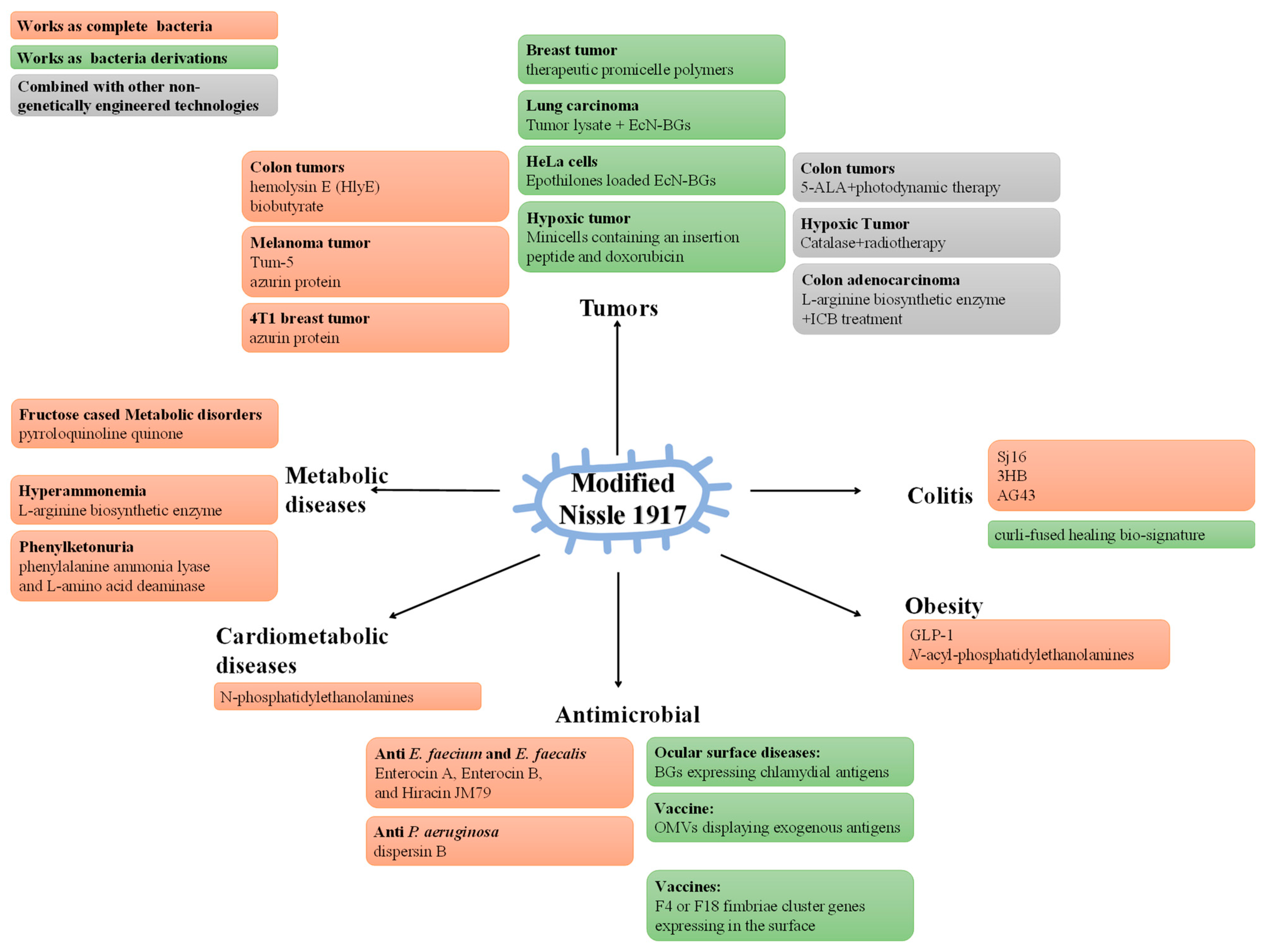{kind=link}
Table 1.
The strategy of modified EcN in the treatment of diseases.
| Strategy | Mechanisms | Functions/Benefits | Reference | |
|---|---|---|---|---|
| Express direct therapeutic factors | express HlyE | the cytotoxicity of released HlyE | against tumors | [192] |
| express Tum-5 | the anti-angiogenesis effects of released Tum-5 | against tumors | [193] | |
| express azurin | azurin selectively kills cancer cells | against tumors | [194] | |
| express colibactin | the cytotoxicity of released colibactin | against tumors | [45] | |
| express glidobactin | the cytotoxicity of released glidobactin | against tumors | [45] | |
| express luminmide | the cytotoxicity of released luminmide | against tumors | [45] | |
| express butyrate | the cytotoxicity of released butyrate | against tumors | [195] | |
| express Sj16 | via Ruminococcaceae/butyrate/retinoic acid axis | against colitis | [196] | |
| express 3HB | regulate gut microbiota, relieve architectural changes, inflammatory cell infiltrations, and epithelial injuries | against colitis | [197] | |
| express autotransporter 43 adhesins antigen | use optogenetics to activate secretion | against colitis/precise spatiotemporal colonization | [198] | |
| express IL-10 | phone visual diagnosis and optogenetics based secretion | against colitis/mobile health service | [199] | |
| express GLP-1 analog | GLP-1 analog diminishes food intake | against obesity | [200] | |
| express N-acyl-phosphatidylethanolamines | N-acyl-phosphatidylethanolamines induce satiety | against obesity | [201] | |
| express antimicrobial peptides | antimicrobial peptides target and kill Enterococcus | against vancomycin-resistant Enterococcus | [202] | |
| develop a synthetic genetic system | sense and kill Pseudomonas aeruginosa | against Pseudomonas aeruginosa | [203] | |
| express N-acyl-phosphatidylethanolamines | reduce body weight, liver inflammation, fibrosis and atherosclerotic necrosis | against cardiovascular metabolic disease | [204] | |
| express PQQ and other metabolizing enzymes | relieve oxidative stress | against heavy metal toxicity/iron deficiency/fructose-induced dyslipidemia/hyperglycemia/hepatic steatosis | [205,206,207] | |
| express l-arg biosynthetic enzyme | exhausts ammonia for L-arginine biosynthesis | against hyperammonemia | [208] | |
| express phenylalanine ammonia lyase and L-amino acid deaminase | consume phenylalanine within the gastrointestinal tract | against PKU | [209] | |
| Express adjuvant therapeutic factors | express 5-ALA | 5-ALA contributes to photodynamic therapy | against cancer | [210] |
| express CAT | increase the production of O2 contributing to radiotherapy | against cancer | [211] | |
| produce high local concentrations of arginine | the enhancement on T-cell anti-tumor activity of L-arginine in tumors | against cancer | [212] | |
| express cholera autoinducer 1 | use the qurom sensing of bacteria | against Vibrio cholerae | [213,214] | |
| Targeted delivery system | connect therapeutic promicelle polymers on bacteria | connection material responsively to acidic tumor microenvironment to release drug | against cancer | [215,216] |
| secrete curli-fused healing bio-signature | curli fibers bind firmly to inflammation site and transport therapeutic factors | against colitis | [217,218] | |
| use EcN-GBs as an adjuvant | induce cellular immune responses | against LLC | [219] | |
| EcN-GBs load Epothilones | the cytotoxicity of released Epothilones | against cancer | [220] | |
| modified EcN-GBs temporal and spatial release of contained drugs | using photothermal effect of nanorods to modulate release | release modulation | [221] | |
| express chlamydial antigens | induce cellular immune responses | against ocular surface diseases | [222] | |
| display exogenous antigens (ClyA fusion chimera) on the surface of EcN-OMVs | induce cellular immune responses | recombinant subunit vaccines deliver | [223] | |
| express F4 or F18 fimbriae in the surface of EcN | induce cellular immune responses | live vaccine application | [224] | |
| EcN-derived minicells loaded with a low-pH insertion peptide and doxorubicin | greater drug loading and therapeutic index contribute to doxorubicin tumor regression | against cancer | [225] | |
Table 2.
The strategy of modified probiotics or probiotic derivatives in the treatment of diseases.
| Probiotics | Strategy | Mechanisms | Functions/Benefits | Reference | |
|---|---|---|---|---|---|
| Akkermansia muciniphila | OMVS delivery/treatment | regulate inflammation, energy homeostasis, intestinal barrier | against obesity | [230] | |
| OMVS delivery/treatment | activate the AMPK pathway and increase TJ gene expressions | against leaky gut | [231,232] | ||
| OMVS delivery/treatment | regulate inflammation, epithelial stability | against DSS-induced colitis | [233] | ||
| OMVS delivery/treatment | reduce inflammation, reverse the activation of hepatic stellate cells and normalize serum glucose, lipid profiles, liver enzymes | against HFD/CCl4-induced liver fibrosis | [234] | ||
| OMVS delivery/treatment | promote osteogenesis and inhibit osteoclastogenesis | against osteoporosis | [238] | ||
| OMVS delivery/treatment | regulate CD8+ T cells and macrophages | against prostate cancer | [62] | ||
| OMVS delivery/treatment | induce serotonin biosynthesis | promote serotonin | [236] | ||
| Clostridium butyricum | OMVS delivery/treatment | enhance proinflammatory cytokine production | stimulate the immune system | [239] | |
| overexpress epidermal growth factor | activate STAT3 signal pathway and inhibit inflammation | gut protection | [240] | ||
| Bifidobacterium longum | express RhMnSOD | regulate inflammatory cytokines | against DSS-induced colitis | [241] | |
| express α-melanocyte-stimulating hormones | inhibit NF-κB p65 expression | against DSS-induced colitis | [242] | ||
| express interleukin-12 | upregulate the expression of Th1 cytokines (IFN-γ and TNF-α) | against coxsackie virus B3-induced myocarditis | [243] | ||
| express oxyntomodulin | reduces food intake, body weight and blood lipid levels | against obesity | [244] | ||
| express GLP-1 | improve the efficiency of glucose control | against type 2 diabetes | [245] | ||
| Lactic acid bacteria | Limosilactobacillus reuteri | express IL-22 | regulate REG3 via STAT3 | against nonalcoholic fatty liver | [246] |
| Limosilactobacillus reuteri | express IL-22 | induce expression of REG3G | against ethanol-induced steatohepatitis | [247] | |
| Lactococcus lactis | express GLP-1 | downregulate TLR4/NF-κB, upregulated the AKT/GSK3β signaling pathway and reverse disturbed microbiota | against Alzheimer/Parkinson | [248] | |
| Lacticaseibacillus paracasei subsp. paracasei | express angiotensin | increase beneficial circulating neurotransmitters and reduce neuro-inflammatory gene expression | benefits the gut-brain axis | [249] | |
| Lacticaseibacillus paracasei subsp. paracasei | express palmitoylethanolamide | block mucosal immune cell infiltration and the release of pro-inflammatory cytokines | against colitis | [250] | |
| Lacticaseibacillus casei | express bovine viral diarrhea virus E2 protein | induce cellular immune responses | against BVDV | [251] | |
| Lacticaseibacillus casei | express antigenic site of TGEV S protein and major antigen site of PEDV S protein | induce cellular immune responses | against TGEV and PEDV | [252,253,254] | |
| Lacticaseibacillus casei | express IPNV protein antigen VP2 | induce cellular immune responses | against IPNV | [255,256] | |
| Lacticaseibacillus casei | express toxoid of Clostridium perfringens α-toxin | induce cellular immune responses | against Clostridium perfringens | [257] | |
| Lacticaseibacillus casei | express the F4 fimbrial adhesin main subunit | induce cellular immune responses | against F4+ enterotoxigenic Escherichia coli | [258] | |
| Lactiplantibacillus plantarum subsp. plantarum | express PEDV S1 protein | induce cellular immune responses | against PEDV | [259] | |
| Lactiplantibacillus plantarum subsp. plantarum | express Staphylococcus aureus nontoxic mutated α-hemolysins | induce cellular immune responses | against Staphylococcus aureus | [260] | |
| Lactobacillus acidophilus | express Human CD4 on the surface | capture and neutralize HIV-1 | against HIV-1 | [261] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, H.; Zhao, Z.; Zhao, Z.; Chen, Y.; Zhang, L. Native and Engineered Probiotics: Promising Agents against Related Systemic and Intestinal Diseases. Int. J. Mol. Sci. 2022, 23, 594. https://doi.org/10.3390/ijms23020594
AMA Style
Shen H, Zhao Z, Zhao Z, Chen Y, Zhang L. Native and Engineered Probiotics: Promising Agents against Related Systemic and Intestinal Diseases. International Journal of Molecular Sciences. 2022; 23(2):594. https://doi.org/10.3390/ijms23020594
Chicago/Turabian StyleShen, Haokun, Zitong Zhao, Zengjue Zhao, Yuyi Chen, and Linghua Zhang. 2022. "Native and Engineered Probiotics: Promising Agents against Related Systemic and Intestinal Diseases" International Journal of Molecular Sciences 23, no. 2: 594. https://doi.org/10.3390/ijms23020594
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.