
High-Performance Thin-Layer Chromatography-Densitometry-Tandem ESI-MS to Evaluate Phospholipid Content in Exosomes of Cancer Cells

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
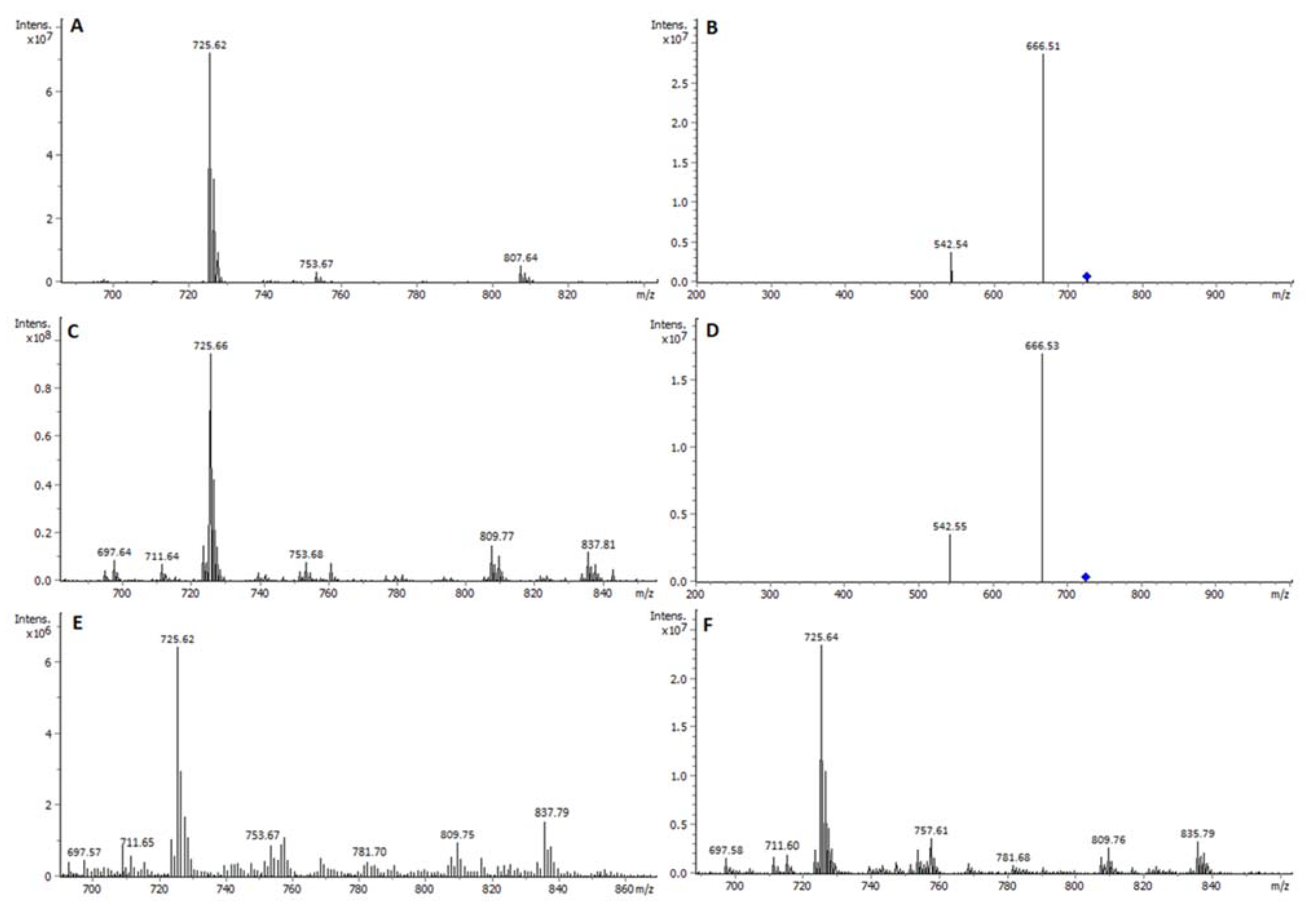
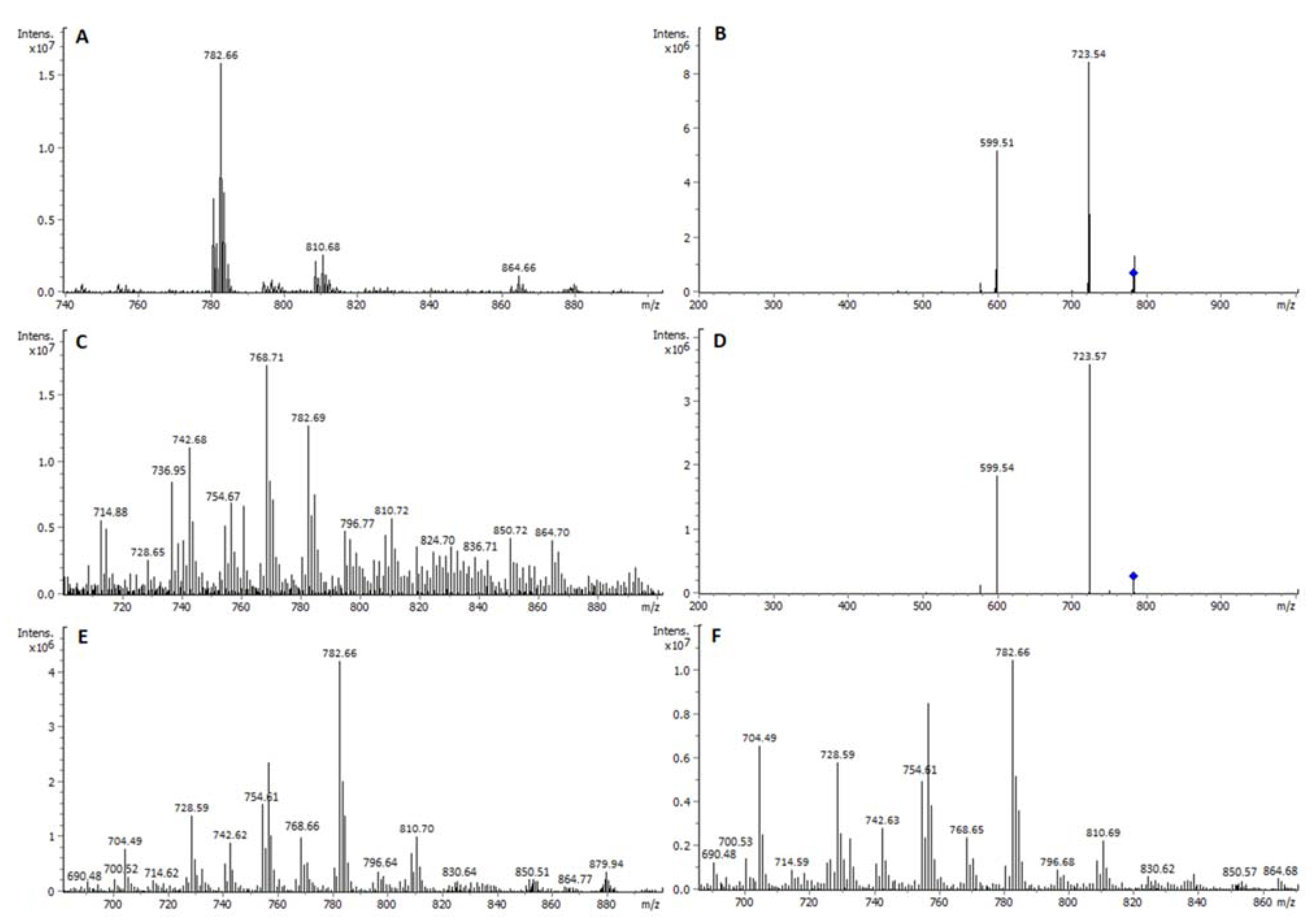
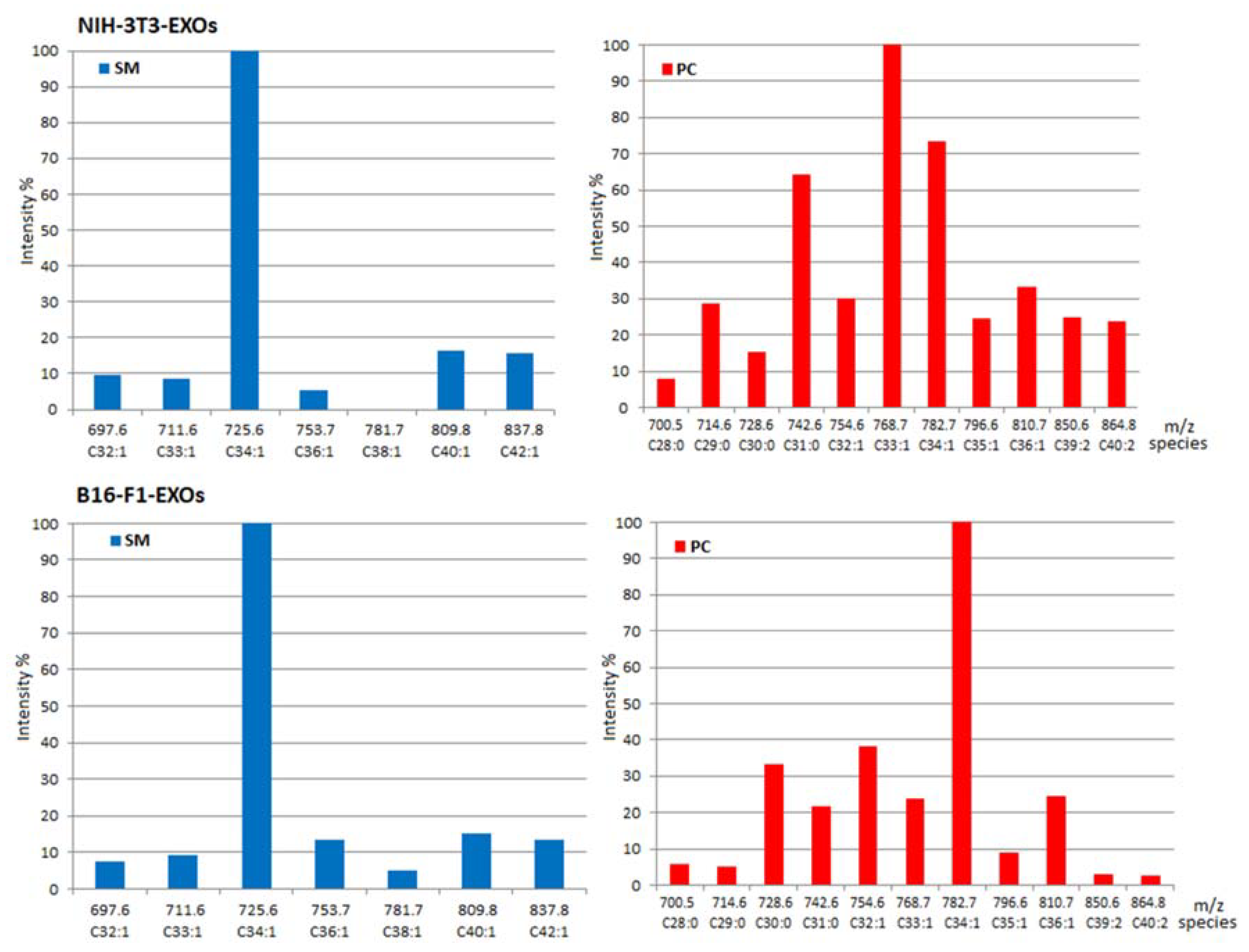
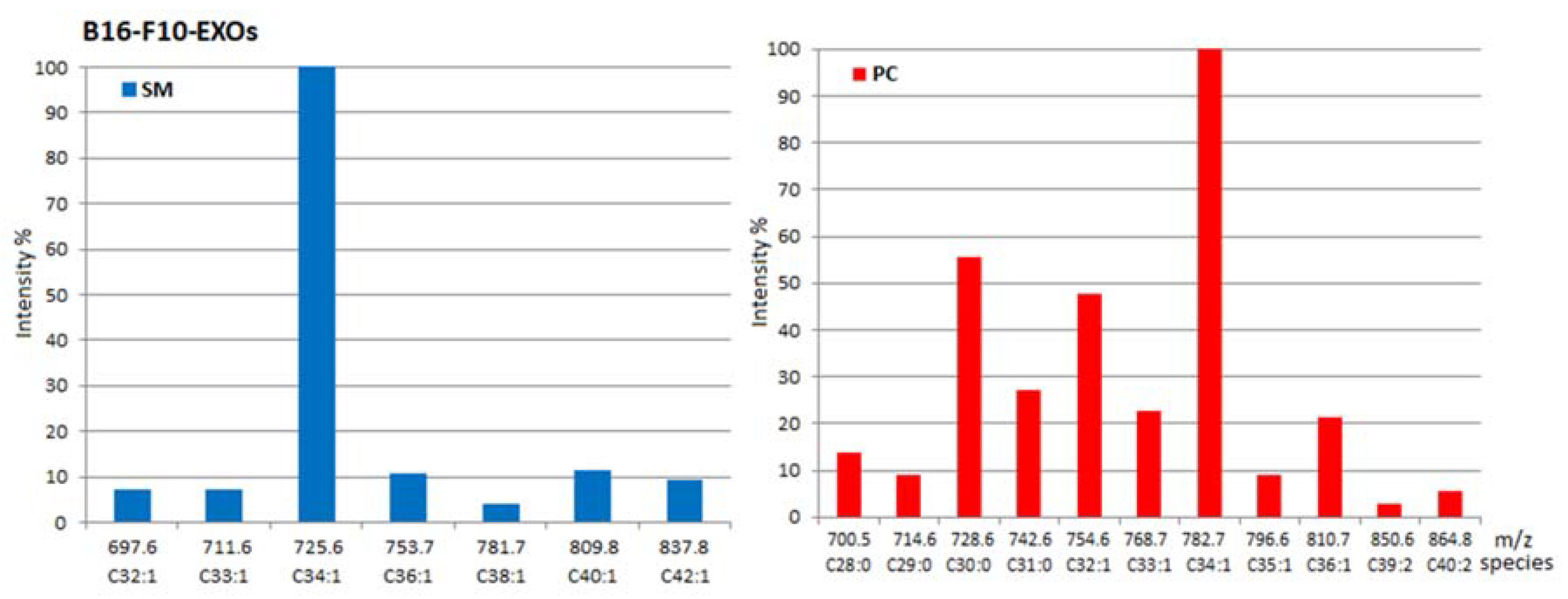
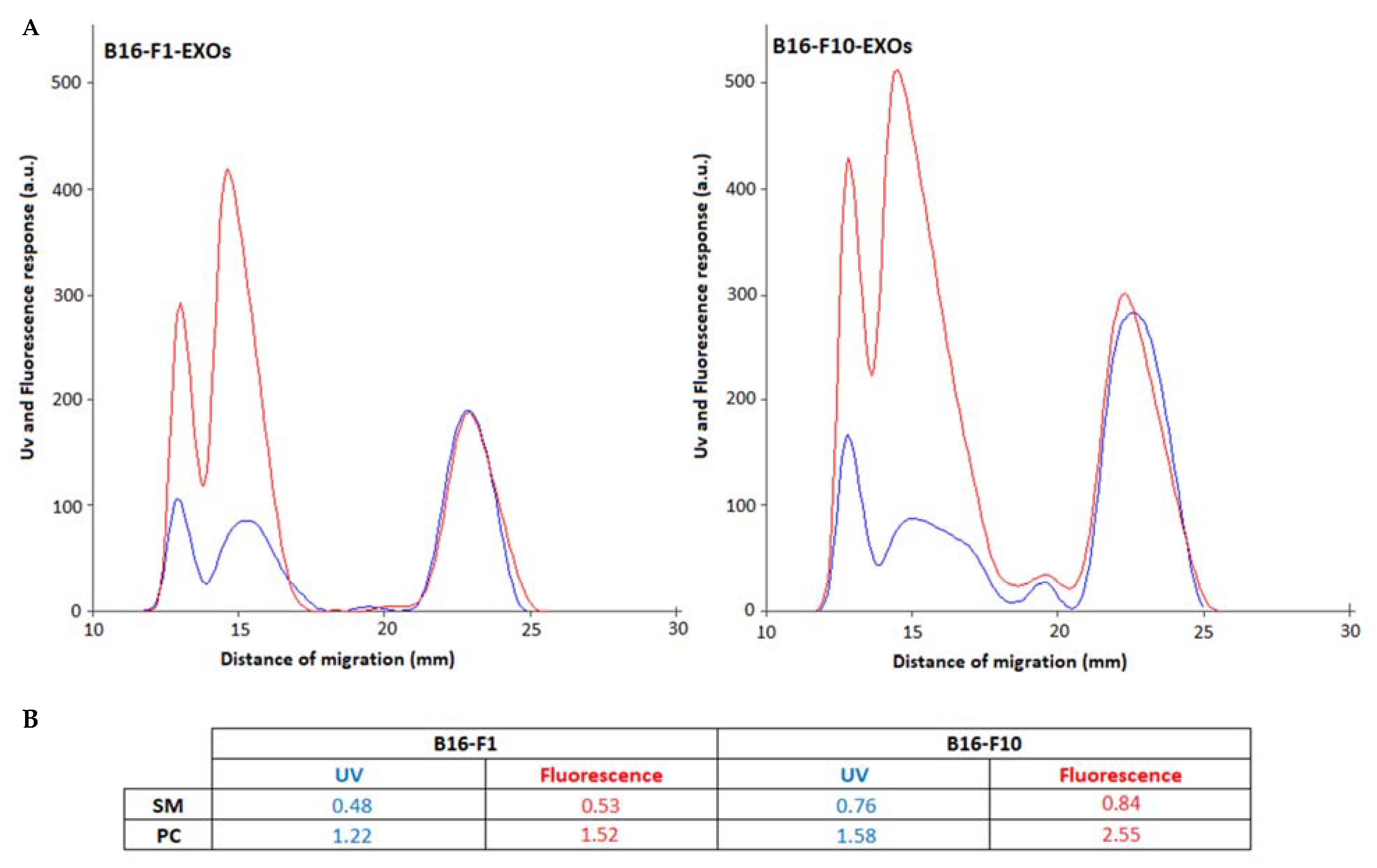
2.1. Identity of PL Classes and Their Molecular Species
2.2. Quantitative Densitometry for PL Classes
3. Materials and Methods
3.1. Cell Culture Conditions
3.2. Exosome Isolation
3.3. Total Lipid Isolation from Exosomes
3.4. Standards and Chemicals
3.5. HPTLC-Densitometry
3.6. HPTLC-MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Skotland, T.; Sagini, K.; Sandvig, K.; Llorente, A. An emerging focus on lipids in extracellular vesicles. Adv. Drug Deliv. Rev. 2020, 159, 308–321. [Google Scholar] [CrossRef]
- Skotland, T.; Sandvig, K.; Llorente, A. Lipids in Exosomes: Current Knowledge and the Way Forward. Prog. Lipid Res. 2017, 66, 30–41. [Google Scholar] [CrossRef]
- Han, X.L. Lipidomics. Comprehensive Mass Spectrometry of Lipids; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Skotland, T.; Hessvik, N.P.; Sandvig, K.; Llorente, A. Exosomal Lipid Composition and the Role of Ether Lipids and Phosphoinositides in Exosome Biology. J. Lipid Res. 2019, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.W.; Megna, J.S.; Nguyen, J. Composition, Physicochemical and Biological Properties of Exosomes Secreted From Cancer Cells. In Diagnostic and Therapeutic Applications of Exosomes in Cancer; Amiji, M., Ramesh, R., Eds.; Academic Press: London, UK, 2015; Volume 3, pp. 45–47. [Google Scholar]
- Skotland, T.; Ekroos, K.; Kauhanen, D.; Simolin, E.; Seierstad, T.; Berge, V.; Sandvig, K.; Llorente, A. Molecular lipid species in urinary exosomes as potential prostate cancer biomarkers. Eur. J. Cancer 2017, 70, 122–132. [Google Scholar] [CrossRef]
- Llorente, A.; Skotland, T.; Sylvänne, T.; Kauhanen, D.; Róg, T.; Orłowski, A.; Ekroos, K.; Sandvig, K. Molecular lipidomics of exosomes released by PC-3 prostate cancer cells. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2013, 1831, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Donoso-Quezada, J.; Ayala-Mar, S.; González-Valdez, J. The role of lipids in exosome biology and intercelular communciation: Function, analytics and applications. Traffic 2021, 22, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Corcuff, J.B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and inmunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Haraszti, R.A.; Didiot, M.C.; Sapp, E.; Leszyk, J.; Shaffer, S.A.; Rockwell, H.E.; Gao, F.; Narain, N.R.; DiFiglia, M.; Kiebish, M.A.; et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J. Extracell. Vesicles 2016, 5, 32570. [Google Scholar] [CrossRef]
- Beloribi, S.; Ristorcelli, E.; Breuzard, G.; Silvy, F.; Bertrand-Michel, J.; Beraud, E.; Verine, A.; Lombardo, D. Exosomal lipids impact notch signaling and induce death of human pancreatic tumoral SOJ-6 cells. PLoS ONE 2012, 7, e47480. [Google Scholar] [CrossRef]
- Cebolla, V.L.; Jarne, C.; Vela, J.; Garriga, R.; Membrado, L.; Galbán, J. Scanning densitometry and mass spectrometry for HPTLC analysis of lipids: The last 10 years. J. Liq. Chromatogr. Relat. Technol. 2021, 44, 148–170. [Google Scholar] [CrossRef]
- Singhto, N.; Vinaiphat, A.; Thongboonkerd, V. Discrimination of Urinary Exosomes from Microvesicles by Lipidomics Using Thin Layer Liquid Chromatography (TLC) Coupled with MALDI-TOF Mass Spectrometry. Sci. Rep. 2019, 9, 13834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiller, J.; Fuchs, B.; Suss, R.; Popkova, Y.; Griesinger, H.; Matheis, K.; Oberle, M.; Schulz, M. TLC/MALDI MS for the Analysis of Lipids. In Planar Chromatography-Mass Spectrometry; Kowalska, T., Sajewicz, M., Sherma, J., Eds.; Chromatography Science Series; CRC Press/Taylor & Francis Group: Boca Raton, FL, USA, 2016; Volume 110, pp. 213–232. [Google Scholar]
- Jarne, C.; Membrado, L.; Savirón, M.; Vela, J.; Orduna, J.; Garriga, R.; Galbán, J.; Cebolla, V.L. Globotriaosylceramide-related biomarkers of fabry disease identified in plasma by high-performance thin-layer chromatography-densitometry-mass spectrometry. J. Chromatogr. A 2021, 1638, 461895. [Google Scholar] [CrossRef] [PubMed]
- Engel, K.M.; Schiller, J. The value of coupling thin-layer chromatography to mass spectrometry in lipid research—A review. J. Chromatogr. B 2021, 1185, 123001. [Google Scholar] [CrossRef] [PubMed]
- Morlock, G.; Schwack, W. Coupling of planar chromatography to mass spectrometry. TrAC 2010, 29, 1157–1171. [Google Scholar] [CrossRef]
- Jamin, E.L.; Jacques, C.; Jourdes, L.; Tabet, J.-C.; Borotra, N.; Bessou-Touya, S.; Debrauwer, L.; Duplan, H. Identification of Lipids of the Stratum Corneum by High Performance Thin Layer Chromatography and Mass Spectrometry. Eur. J. Mass Spectrom. 2019, 25, 278–290. [Google Scholar] [CrossRef]
- Lapieza, M.P.; Jungas, C.; Savirón, M.; Jarne, C.; Membrado, L.; Vela, J.; Orduna, J.; Garriga, R.; Galbán, J.; Cebolla, V.L. HPTLC Coupled to ESI-Tandem MS for Identifying Phospholipids Associated to Membrane Proteins in Photosynthetic Purple Bacteria. J. Liq. Chromatogr. Relat. Technol. 2019, 42, 1–8. [Google Scholar] [CrossRef]
- Cebolla, V.L.; Mateos, E.; Garriga, R.; Jarne, C.; Membrado, L.; Cossío, F.P.; Gálvez, E.M.; Matt, M.; Delgado-Camón, A. Changes in Fluorescent Emission Due to Non-Covalent Interactions as a General Detection Procedure for Thin-Layer Chromatography. ChemPhysChem 2012, 13, 291–299. [Google Scholar] [CrossRef]
- Domínguez, A.; Jarne, C.; Cebolla, V.L.; Galbán, J.; Savirón, M.; Orduna, J.; Membrado, L.; Lapieza, M.P.; Romero, E.; Sanz-Vicente, I.; et al. A Hyphenated Technique Based on High-Performance Thin Layer Chromatography for Determining Neutral Sphingolipids: A Proof of Concept. Chromatography 2015, 2, 167–187. [Google Scholar] [CrossRef] [Green Version]
- Folch, J.; Less, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Brzozowski, J.S.; Jankowski, H.; Bond, D.R.; McCague, S.B.; Munro, B.R.; Predebon, M.J.; Scarlett, C.J.; Skelding, K.A.; Weidenhofer, J. Lipidomic profiling of extracellular vesicles derived from prostate and prostate cancer cell lines. Lipids Health Dis. 2018, 17, 211. [Google Scholar] [CrossRef] [Green Version]
- Keller, B.O.; Sui, J.; Young, A.B.; Whittal, R.M. Interferences and Contaminants Encountered in Modern Mass Spectrometry. Anal. Chim. Acta 2008, 627, 71–81. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inherent characteristics of HPTLC-densitometry-MS |
|
| Differential characteristics of HPTLC vs. shotgun MS by direct infusion |
|
| Differential characteristics vs. HPTLC on LC-MS |
| LC-MS is more efficient in separation than HPTLC. LC-MS quantification is complicated as there are variations in ionization of molecules of the same lipid class due to differences in the chemical environment ionization at different retention times. HPTLC quantification is usually carried out by densitometry. No studies have been published on HPTLC-MS quantification |
| PL | NIH-3T3-Exos | B16-F1-Exos | B16-F10-Exos |
|---|---|---|---|
| SM | 0.50 (RSD: 18.00%) | 0.48 (RSD: 17.40%) | 0.76 (RSD: 8.50%) |
| PC | 0.79 (RSD: 15.80%) | 1.22 (RSD: 6.30%) | 1.58 (RSD: 2.60%) |
| PS | 0.08 (RSD: 11.80%) | 0.02 (RSD: 55.00%) | 0.08 (RSD: 47.60%) |
| PE | 0.20 (RSD: 8.00%) | 1.99 (RSD: 2.80%) | 3.70 (RSD: 1.60%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho-Albero, M.; Jarne, C.; Savirón, M.; Martín-Duque, P.; Membrado, L.; Cebolla, V.L.; Santamaría, J. High-Performance Thin-Layer Chromatography-Densitometry-Tandem ESI-MS to Evaluate Phospholipid Content in Exosomes of Cancer Cells. Int. J. Mol. Sci. 2022, 23, 1150. https://doi.org/10.3390/ijms23031150
Sancho-Albero M, Jarne C, Savirón M, Martín-Duque P, Membrado L, Cebolla VL, Santamaría J. High-Performance Thin-Layer Chromatography-Densitometry-Tandem ESI-MS to Evaluate Phospholipid Content in Exosomes of Cancer Cells. International Journal of Molecular Sciences. 2022; 23(3):1150. https://doi.org/10.3390/ijms23031150
Chicago/Turabian StyleSancho-Albero, María, Carmen Jarne, María Savirón, Pilar Martín-Duque, Luis Membrado, Vicente L. Cebolla, and Jesús Santamaría. 2022. "High-Performance Thin-Layer Chromatography-Densitometry-Tandem ESI-MS to Evaluate Phospholipid Content in Exosomes of Cancer Cells" International Journal of Molecular Sciences 23, no. 3: 1150. https://doi.org/10.3390/ijms23031150