
K143R Amino Acid Substitution in 14-α-Demethylase (Erg11p) Changes Plasma Membrane and Cell Wall Structure of Candida albicans


Abstract
:1. Introduction
2. Results
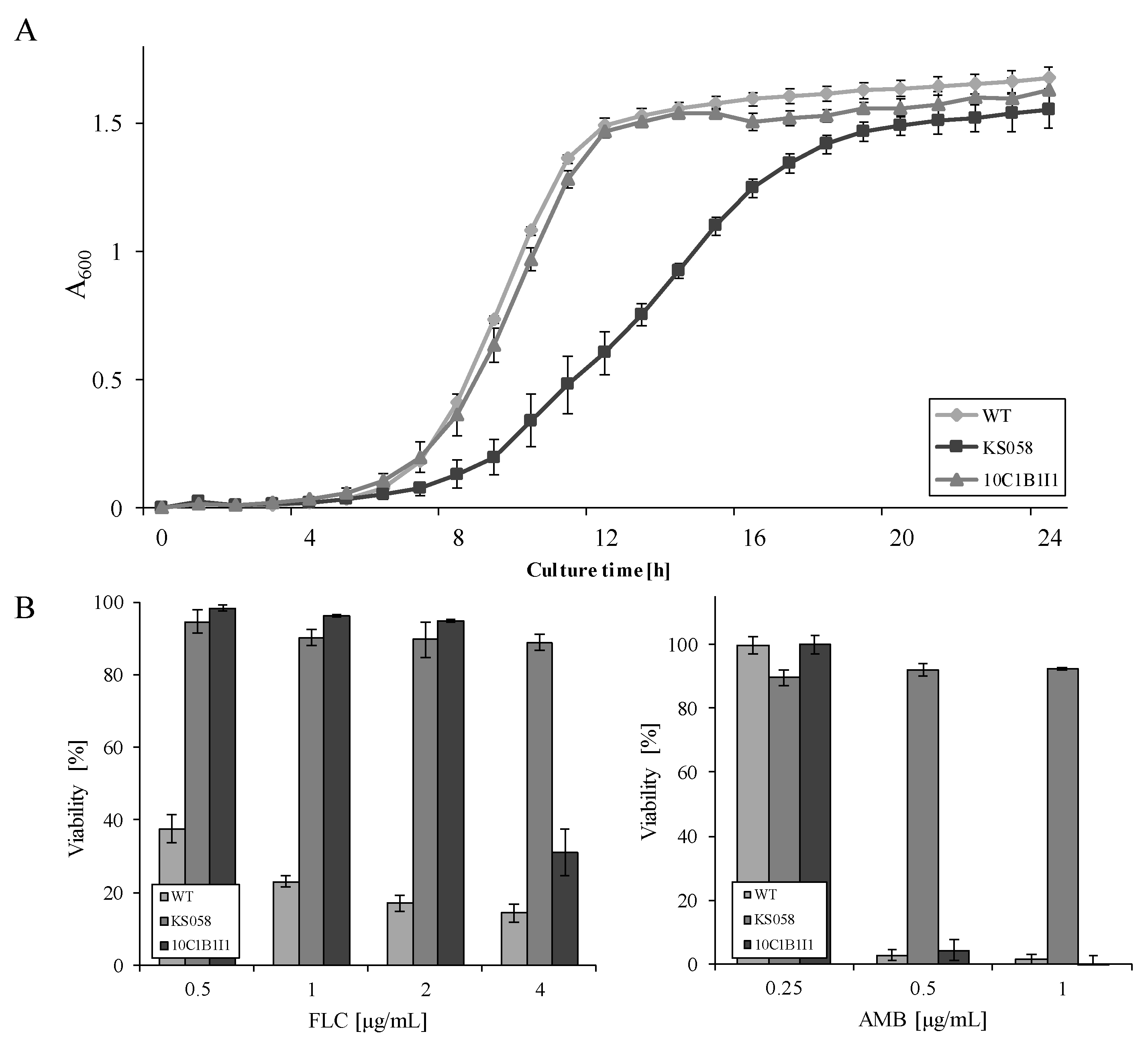
2.1. Point Mutation in ERG11 Does Not Affect the Growth of the C. albicans ERG11K143R/K143R Strain and Contributes to Increased Fluconazole Resistance
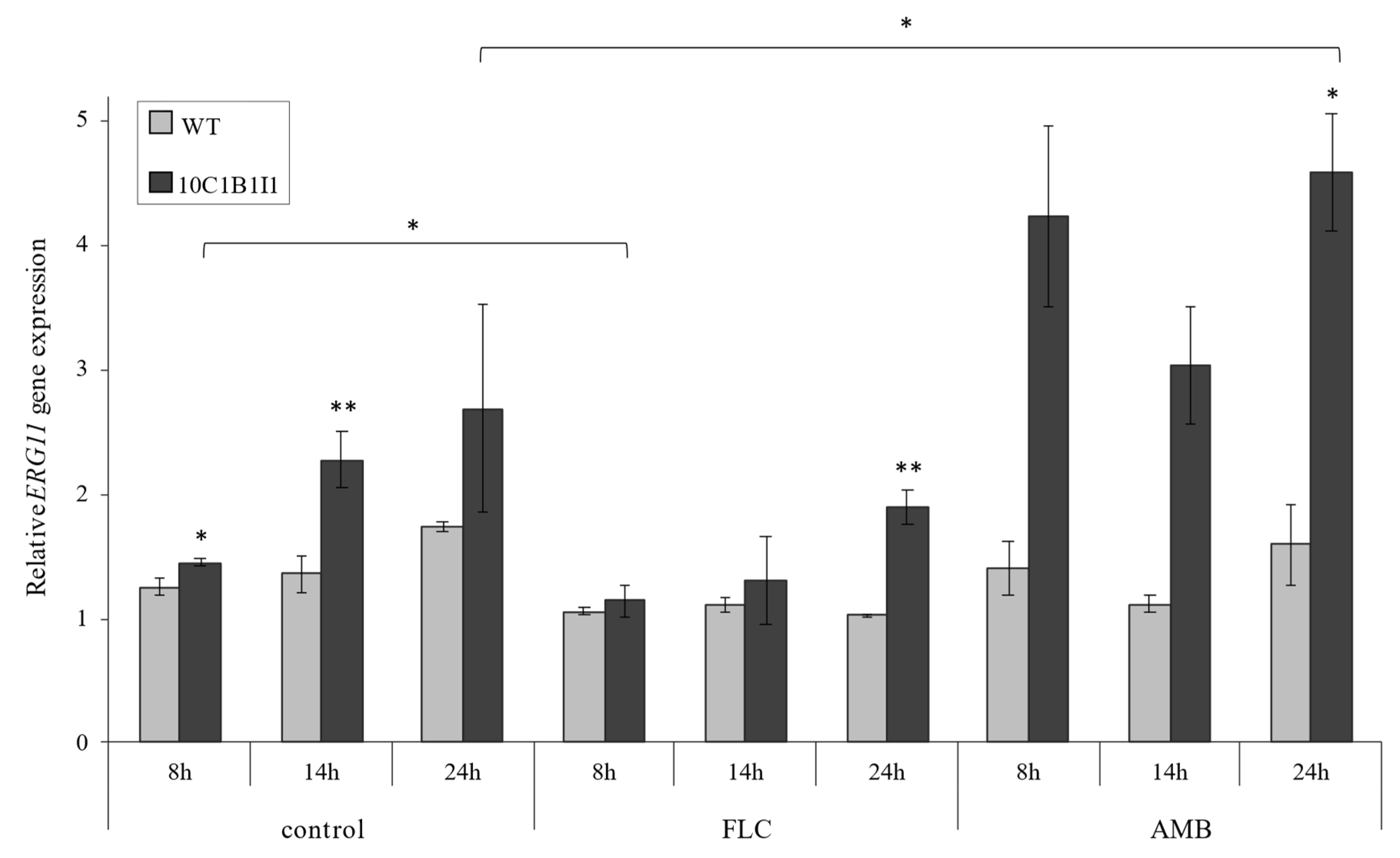
2.2. The Point Mutation ERG11K143R/K143R Results in an Increased ERG11 Gene Expression in the C. albicans 10C1B1I1 Strain
2.3. The Overexpression of the ERG11 Gene Corresponds to an Elevated Ergosterol Content in the C. albicans ERG11K143R/K143R Strain
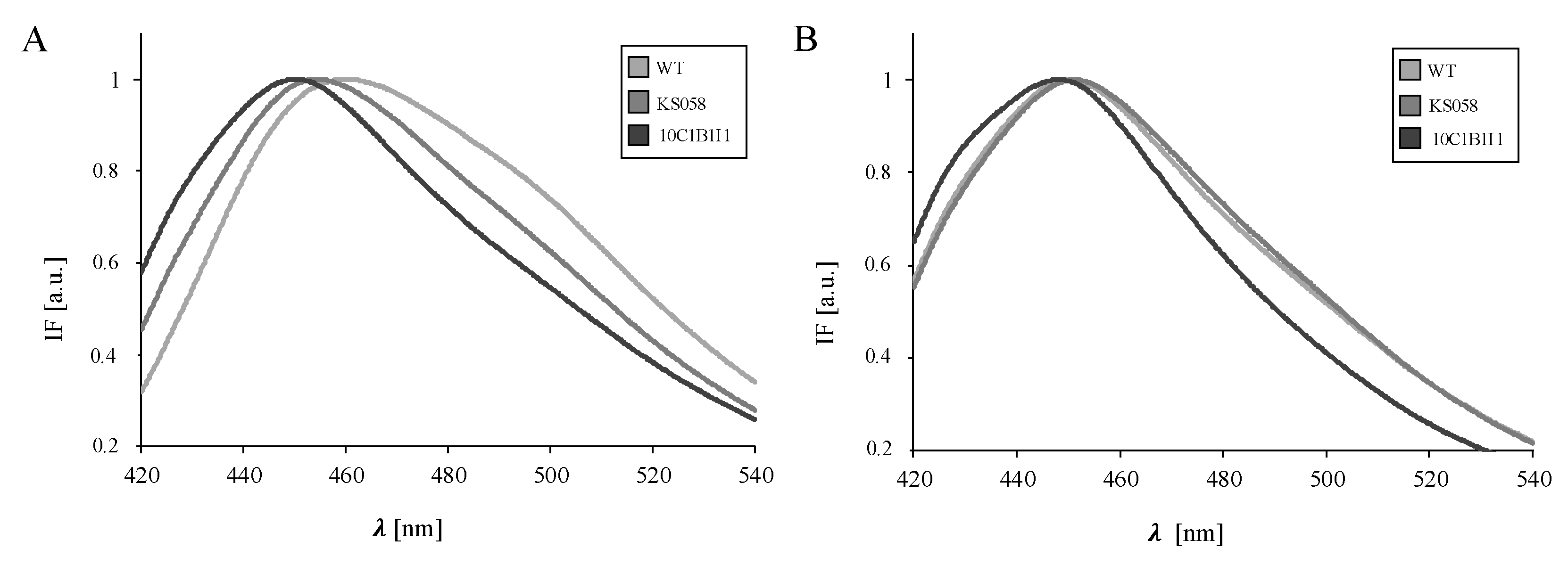
2.4. An Increased Ergosterol Content in C. albicans ERG11K143R/K143R Results in Decreased Plasma Membrane Fluidity
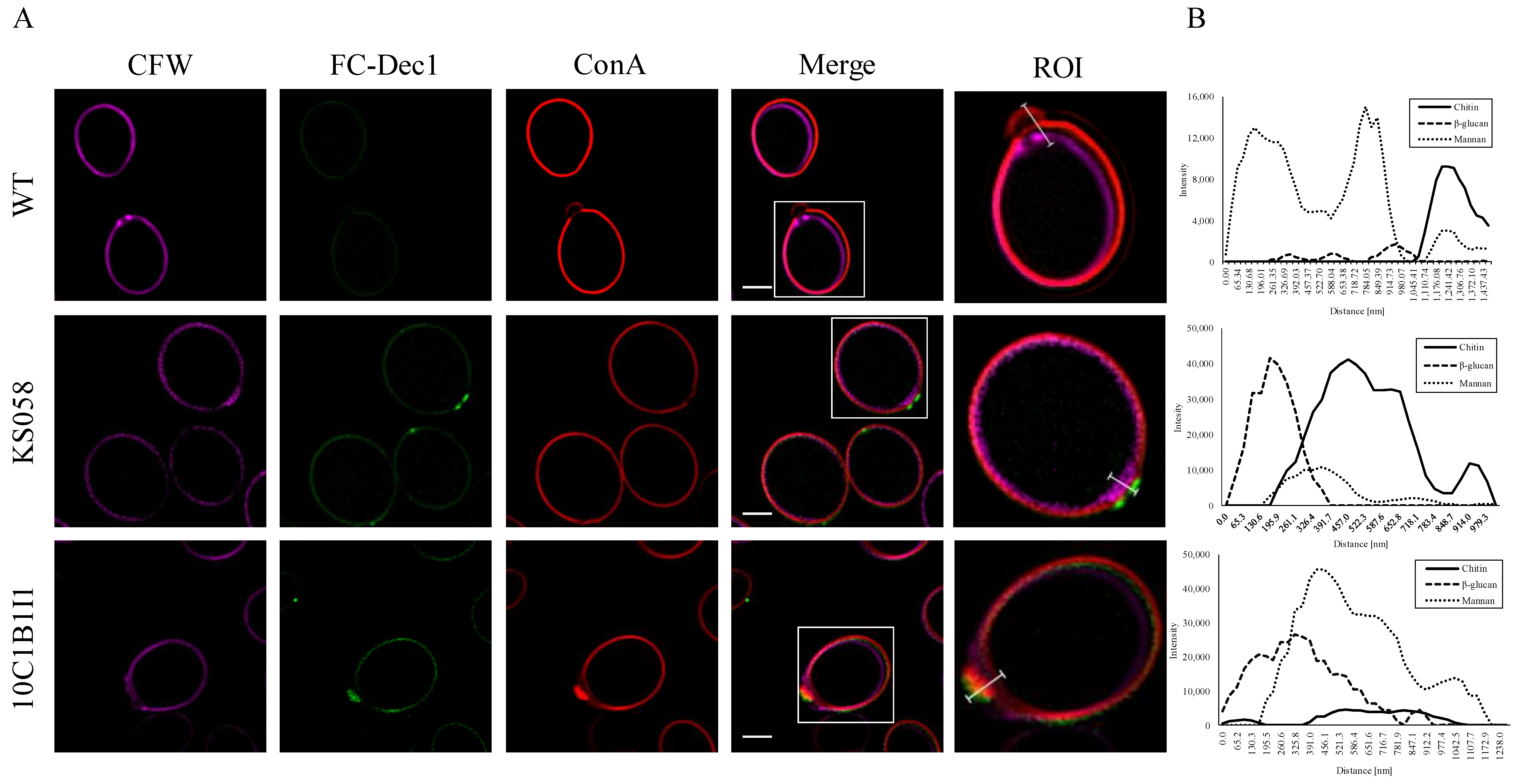
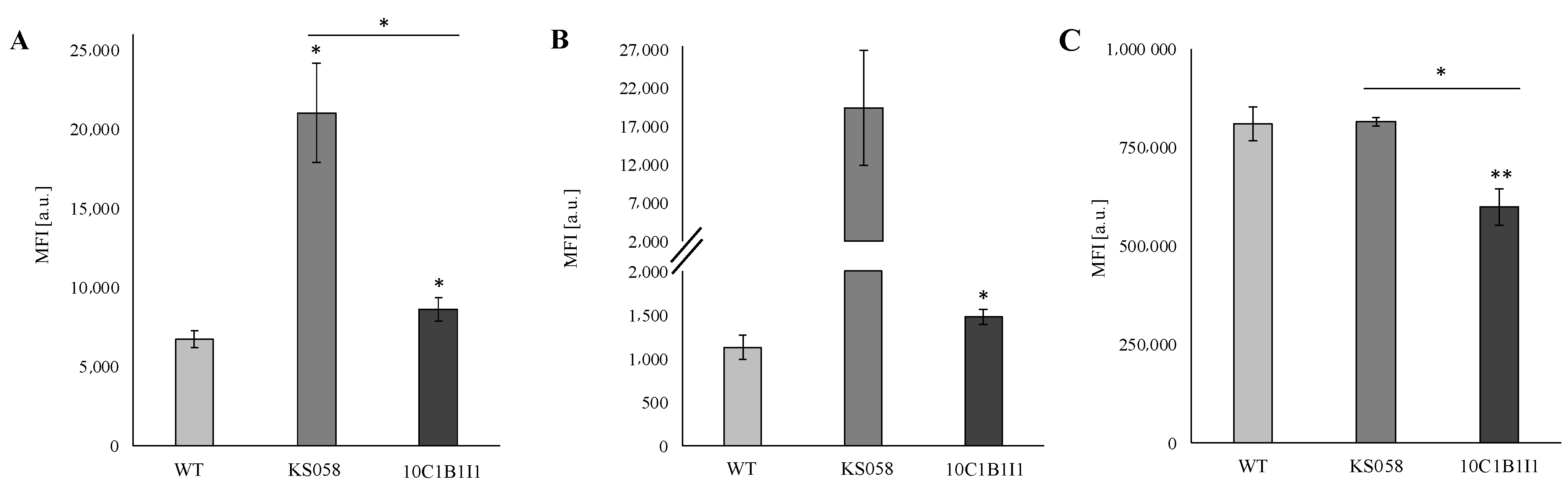
2.5. The Deletion and Increased Expression of the C. albicans ERG11 Gene Correlates with CW Remodeling
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Strains and Culture Conditions
4.3. The Determination of Growth Phases (Growth Curve)
4.4. Minimal Inhibitory Concentration (MIC)
4.5. Real-Time Quantitative PCR (RT-qPCR) Reaction
4.6. Plasma Membrane Isolation and Sterol Analyses
4.7. GC-MS Sterol Analysis in the Plasma Membrane
4.8. The Plasma Membrane Fluidity Measurement (Laurdan Fluorescent Probe)
4.9. Staining of Cell Wall and Structured Illumination Microscopy (SIM)
4.10. Fluorescence-Activated Cell Sorting (FACS) Analyses
4.11. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gao, Y.; Liang, G.; Wang, Q.; She, X.; Shi, D.; Shen, Y.; Su, X.; Wang, X.; Wang, W.; Li, D.; et al. Different Host Immunological Response to C. albicans by Human Oral and Vaginal Epithelial Cells. Mycopathologia 2019, 184, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yeşilbağ, Z.; Şeker, Y.T.; Avci, K.D.; Şenoğlu, S.; Çukurova, Z.; Hergünsel, G.O. Changing epidemiology and risk factors for candidemia in critically ill patients. J. Surg. Med. 2021, 5, 97–102. [Google Scholar] [CrossRef]
- Tsay, S.V.; Mu, Y.; Williams, S.; Epson, E.; Nadle, J.; Bamberg, W.M.; Barter, D.M.; Johnston, H.L.; Farley, M.M.; Harb, S.; et al. Burden of Candidemia in the United States, 2017. Clin. Infect. Dis. 2020, 71, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Patterson, L.; McMullan, R.; Harrison, D.A. Individual risk factors and critical care unit effects on Invasive Candida Infec-tion occurring in critical care units in the UK: A multilevel model. Mycoses 2019, 62, 790–795. [Google Scholar] [PubMed]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Maertens, J. History of the development of azole derivatives. Clin. Microbiol. Infect. 2004, 10, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida albicans and Emerging Non-albicans Candida Species. Front. Microbiol. 2017, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Fumiyoshi, A.; Toshiki, H. Mechanistic role of ergosterol in membrane rigidity and cycloheximide resistance in Saccharomyces cerevisiae. Biochim. Biophys. Acta (BBA)-Biomembr. 2009, 1788, 743–752. [Google Scholar]
- Francois, E.J.A.I.; Cammue, B.P.A.; Borgers, M.; Ausma, J.; Dispersyn, G.D.; Thevissen, K. Azoles: Mode of Antifungal Action and Resistance Development. Effect of Miconazole on Endogenous Reactive Oxygen Species Production in Candida albicans. Anti-Infect. Agents Med. Chem. 2006, 5, 3–13. [Google Scholar] [CrossRef]
- Pristov, K.E.; Ghannoum, M.A. Resistance of Candida to azoles and echinocandins worldwide. Clin. Microbiol. Infect. 2019, 25, 792–798. [Google Scholar] [CrossRef]
- Banerjee, A.; Pata, J.; Sharma, S.; Monk, B.C.; Falson, P.; Prasad, R. Directed Mutational Strategies Reveal Drug Binding and Transport by the MDR Transporters of Candida albicans. J. Fungi 2021, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, P.; Ferrari, S.; Coste, A.T. Antifungal Resistance and New Strategies to Control Fungal Infections. Int. J. Microbiol. 2012, 2012, 713687. [Google Scholar] [CrossRef] [PubMed]
- Bossche, H.V.; Marichal, P.; Odds, F.C. Molecular mechanisms of drug resistance in fungi. Trends Microbiol. 1994, 2, 393–400. [Google Scholar] [CrossRef]
- Letscher-Bru, V.; Herbrecht, R. Caspofungin: The first representative of a new antifungal class. J. Antimicrob. Chemother. 2003, 51, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Wester, M.J.; Graus, M.S.; Lidke, K.A.; Neumann, A.K. Nanoscopic cell-wall architecture of an immunogenic ligand in Candida albicans during antifungal drug treatment. Mol. Biol. Cell 2016, 27, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; De Oliveira, H.C.; Rivera, J.; Trevijano-Contador, N. The Fungal Cell Wall: Candida, Cryptococcus, and Aspergillus Species. Front. Microbiol. 2020, 10, 2993. [Google Scholar] [CrossRef] [PubMed]
- Gow, N.A.; Hube, B. Importance of the Candida albicans cell wall during commensalism and infection. Curr. Opin. Microbiol. 2012, 15, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Shibata, N.; Suzuki, A.; Kobayashi, H.; Okawa, Y. Chemical structure of the cell-wall mannan of Candida albicans serotype A and its difference in yeast and hyphal forms. Biochem. J. 2007, 404, 365–372. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, J.; Derkacz, D.; Muraszko, J.; Panek, J.J.; Jezierska, A.; Łukaszewicz, M.; Krasowska, A. Fluconazole and Lipopeptide Surfactin Interplay During Candida albicans Plasma Membrane and Cell Wall Remodeling Increases Fungal Immune System Exposure. Pharmaceutics 2020, 12, 314. [Google Scholar] [CrossRef] [Green Version]
- Manastır, L.; Ergon, M.C.; Yucesoy, M. Investigation of mutations in Erg11 gene of fluconazole resistant Candida albicans isolates from Turkish hospitals. Mycoses 2011, 54, 99–104. [Google Scholar] [CrossRef]
- Healey, K.R.; Kordalewska, M.; Jiménez Ortigosa, C.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 Mu-tations Identified in Isolates of Candida auris Directly Contribute to Reduced Azole Susceptibility. Antimicrob. Agents Chemother. 2018, 62, e01427-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Muraszko, J.; Bernat, P.; Krasowska, A. A Crucial Role for Ergosterol in Plasma Membrane Composition, Localisation, and Activity of Cdr1p and H+-ATPase in Candida albicans. Microorganisms 2019, 7, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Primers 2018, 4, 18026. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.; Avelar, G.M.; Bain, J.M.; Childers, D.; Pelletier, C.; Larcombe, D.E.; Shekhova, E.; Netea, M.G.; Brown, G.D.; Erwig, L.; et al. Non-canonical signalling mediates changes in fungal cell wall PAMPs that drive im-mune evasion. FEMS Yeast Res. 2019, 10, 5315. [Google Scholar]
- Xiang, M.J.; Liu, J.Y.; Ni, P.H.; Wang, S.; Shi, C.; Wei, B.; Ni, Y.X.; Ge, H.L. Erg11 mutations associated with azole re-sistance in clinical isolates of Candida albicans. FEMS Yeast Res. 2013, 13, 386–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargrove, T.Y.; Friggeri, L.; Wawrzak, Z.; Qi, A.; Hoekstra, W.J.; Schotzinger, R.J.; York, J.D.; Guengerich, F.P.; Lepesheva, G.I. Structural analyses of Candida albicans sterol 14α-demethylase complexed with azole drugs address the molecular basis of azole-mediated inhibition of fungal sterol biosynthesis. J. Biol. Chem. 2017, 292, 6728–6743. [Google Scholar] [CrossRef] [Green Version]
- Flowers, S.A.; Barker, K.S.; Berkow, E.L.; Toner, G.; Chadwick, S.G.; Gygax, S.E.; Morschhäuser, J.; Rogers, P.D. Gain-of-function mutations in UPC2 are a frequent cause of ERG11 upregulation in azole-resistant clinical isolates of Candida albicans. Eukaryot. Cell 2012, 11, 1289–1299. [Google Scholar] [CrossRef] [Green Version]
- Oliver, B.G.; Song, J.L.; Choiniere, J.H.; White, T.C. Cis-Acting elements within the Candida albicans ERG11 promoter me-diate the azole response through transcription factor Upc2p. Eukaryot. Cell 2007, 6, 2231–2239. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Ma, W.; Fan, J.; Liu, X.; Wang, Y.; Zhou, X. The A756T Mutation of the ERG11 Gene Associated With Resistance to Itraconazole in Candida Krusei Isolated From Mycotic Mastitis of Cows. Front. Vet. Sci. 2021, 8, 634286. [Google Scholar] [CrossRef]
- Vu, B.G.; Moye-Rowley, W.S. Multiple mechanisms impact fluconazole resistance of mutant Erg11 proteins in Candida glabrata. bioRxiv 2021, 449691. [Google Scholar] [CrossRef]
- Gale, E.F. The Release of Potassium Ions from Candida albicans in the Presence of Polyene Antibiotics. Microbiology 1974, 80, 451–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Liao, B.; Ye, X.; Zhou, Y.; Chen, X.; Liao, M.; Cheng, L.; Zhou, X.; Ren, B. Artemisinin elevates ergosterol levels of Candida albicans to synergise with amphotericin B against oral candidiasis. Int. J. Antimicrob. Agents 2021, 58, 106394. [Google Scholar] [CrossRef]
- Sanglard, D.; Ischer, F.; Parkinson, T.; Falconer, D.; Bille, J. Candida albicans mutations in the ergosterol biosynthetic path-way and resistance to several antifungal agents. Antimicrob. Agents Chemother. 2003, 47, 2404–2412. [Google Scholar] [CrossRef] [Green Version]
- Abe, F.; Usui, K.; Hiraki, T. Fluconazole modulates membrane rigidity, heterogeneity, and water penetration into the plasma membrane in Saccharomyces cerevisiae. Biochemistry 2009, 48, 8494–8504. [Google Scholar] [CrossRef] [PubMed]
- Lenardon, M.D.; Munro, C.; Gow, N.A. Chitin synthesis and fungal pathogenesis. Curr. Opin. Microbiol. 2010, 13, 416–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballou, E.R.; Avelar, G.M.; Childers, D.S.; Mackie, J.; Bain, J.M.; Wagener, J.; Kastora, S.L.; Panea, M.D.; Hardison, S.E.; Walker, L.A.; et al. Lactate signalling regulates fungal β-glucan masking and immune evasion. Nat. Microbiol. 2016, 12, 16238. [Google Scholar] [CrossRef] [Green Version]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar] [CrossRef]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of Clinically Derived Mutations inERG11to Azole Resistance in Candida albicans. Antimicrob. Agents Chemother. 2014, 59, 450–460. [Google Scholar] [CrossRef] [Green Version]
- Szczepaniak, J.; Łukaszewicz, M.; Krasowska, A. Estimation of Candida albicans ABC Transporter Behavior in Real-Time via Fluorescence. Front. Microbiol. 2015, 6, 1382. [Google Scholar] [CrossRef] [Green Version]
- Suchodolski, J.; Muraszko, J.; Korba, A.; Bernat, P.; Krasowska, A. Lipid composition and cell surface hydrophobicity of Candida albicans influence the efficacy of fluconazole-gentamicin treatment. Yeast 2020, 37, 117–129. [Google Scholar] [CrossRef]
- Amaro, M.; Reina, F.; Hof, M.; Eggeling, C.; Sezgin, E. Laurdan and Di-4-ANEPPDHQ probe different properties of the membrane. J. Phys. D Appl. Phys. 2017, 50, 134004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagener, J.; MacCallum, D.M.; Brown, G.D.; Gow, N.A. Candida albicans chitin in-creases arginase-1 activity in human macrophages, with an impact on macrophage antimicrobial functions. MBio 2017, 8, e01820-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchodolski, J.; Feder-Kubis, J.; Krasowska, A. Antifungal activity of ionic liquids based on (−)-menthol: A mechanism study. Microbiol. Res. 2017, 197, 56–64. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ergosterol | |||||||||
| Control | FLC | AMB | |||||||
| 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | |
| WT | 6.95 ± 1.46 | 49.49 ± 13.12 | 28.82 ± 2.06 | ND | ND | 0.73 ± 1.27 | 31.78 ± 4.48 | 17.14 ± 0.84 | 20.20 ± 0.96 |
| 10C1B1I1 | 7.84 ± 3.73 | 93.82 ± 8.63 * | 37.18 ± 1.53 ** | 5.83 ± 1.73 ** | 11.06 ± 0.71 *** | 5.58 ± 3.10 | 39.82 ± 2.15 * | 19.64 ± 2.73 | 16.19 ± 3.22 |
| Lanosterol | |||||||||
| Control | FLC | AMB | |||||||
| 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | |
| WT | 5.09 ± 2.11 | 2.52 ± 0.95 | 2.43 ± 0.36 | 8.73 ± 0.28 | 3.29 ± 0.77 | 5.87 ± 0.99 | 3.29 ± 0.08 | 4.92 ± 0.45 | 1.51 ± 0.10 |
| 10C1B1I1 | 5.11 ± 2.81 | 4.04 ± 0.93 | 2.31 ± 0.77 | 3.57 ± 0.99 *** | 8.79 ± 2.33 * | 5.72 ± 1.61 | 5.49 ± 1.60 | 1.77 ± 0.41 *** | 1.45 ± 0.33 |
| KS058 | 20.70 ± 0.43 ** | 11.45 ± 0.95 *** | 22.53 ± 0.46 *** | 26.06 ± 3.98 * | 11.33 ± 1.45 ** | 53.34 ± 6.89 ** | 21.41 ± 3.25 *** | 11.16 ± 2.44 * | 20.37 ± 1.00 *** |
| Eburicol | |||||||||
| Control | FLC | AMB | |||||||
| 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | |
| WT | ND | ND | ND | 2.88 ± 2.86 | 4.41 ± 3.91 | 3.41 ± 3.25 | ND | ND | ND |
| 10C1B1I1 | ND | ND | ND | 2.13 ± 2.43 | 1.97 ± 2.28 | 4.39 ± 4.43 | 0.17 ± 0.29 | 3.72 ± 6.32 | ND |
| KS058 | 9.88 ± 0.95 ** | 8.12 ± 1.53 * | 26.64 ± 2.14 ** | 9.41 ± 2.12 * | 9.47 ± 1.11 | 40.07 ± 2.60 *** | 10.32 ± 2.14 * | 7.76 ± 0.88 ** | 19.48 ± 1.22 ** |
| 14-α methylergosta-8-24(28)dienol | |||||||||
| Control | FLC | AMB | |||||||
| 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | 8 h | 14 h | 24 h | |
| WT | ND | ND | ND | ND | ND | 3.41 ± 3.24 | ND | ND | ND |
| 10C1B1I1 | ND | ND | ND | ND | ND | 4.39 ± 4.43 | ND | ND | ND |
| KS058 | 10.02 ± 0.32 *** | 9.00 ± 0.82 ** | 13.31 ± 0.69 *** | 17.06 ± 1.68 ** | 8.77 ± 1.64 * | 26.12 ± 5.07 ** | 12.4 ± 1.81 ** | 15.12 ± 2.35 ** | 12.18 ± 1.52 ** |
| Control | FLC | AMB | ||||
|---|---|---|---|---|---|---|
| 8 h | 24 h | 8 h | 24 h | 8 h | 24 h | |
| WT | −0.33 ± 0.046 | −0.07 ± 0.111 | −0.31 ± 0.049 | −0.35 ± 0.026 | −0.37 ± 0.038 | −0.38 ± 0.029 |
| KS058 | −0.21 ± 0.026 *** | −0.09 ± 0.122 | −0.29 ± 0.069 | −0.34 ± 0.017 | −0.37 ± 0.040 | −0.36 ± 0.049 |
| 10C1B1I1 | −0.10 ± 0.036 *** | 0.05 ± 0.023 * | −0.26 ± 0.085 | −0.32 ± 0.015 | −0.28 ± 0.021 ** | −0.35 ± 0.077 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derkacz, D.; Bernat, P.; Krasowska, A. K143R Amino Acid Substitution in 14-α-Demethylase (Erg11p) Changes Plasma Membrane and Cell Wall Structure of Candida albicans. Int. J. Mol. Sci. 2022, 23, 1631. https://doi.org/10.3390/ijms23031631
Derkacz D, Bernat P, Krasowska A. K143R Amino Acid Substitution in 14-α-Demethylase (Erg11p) Changes Plasma Membrane and Cell Wall Structure of Candida albicans. International Journal of Molecular Sciences. 2022; 23(3):1631. https://doi.org/10.3390/ijms23031631
Chicago/Turabian StyleDerkacz, Daria, Przemysław Bernat, and Anna Krasowska. 2022. "K143R Amino Acid Substitution in 14-α-Demethylase (Erg11p) Changes Plasma Membrane and Cell Wall Structure of Candida albicans" International Journal of Molecular Sciences 23, no. 3: 1631. https://doi.org/10.3390/ijms23031631