
Delayed Minocycline Treatment Ameliorates Hydrocephalus Development and Choroid Plexus Inflammation in Spontaneously Hypertensive Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
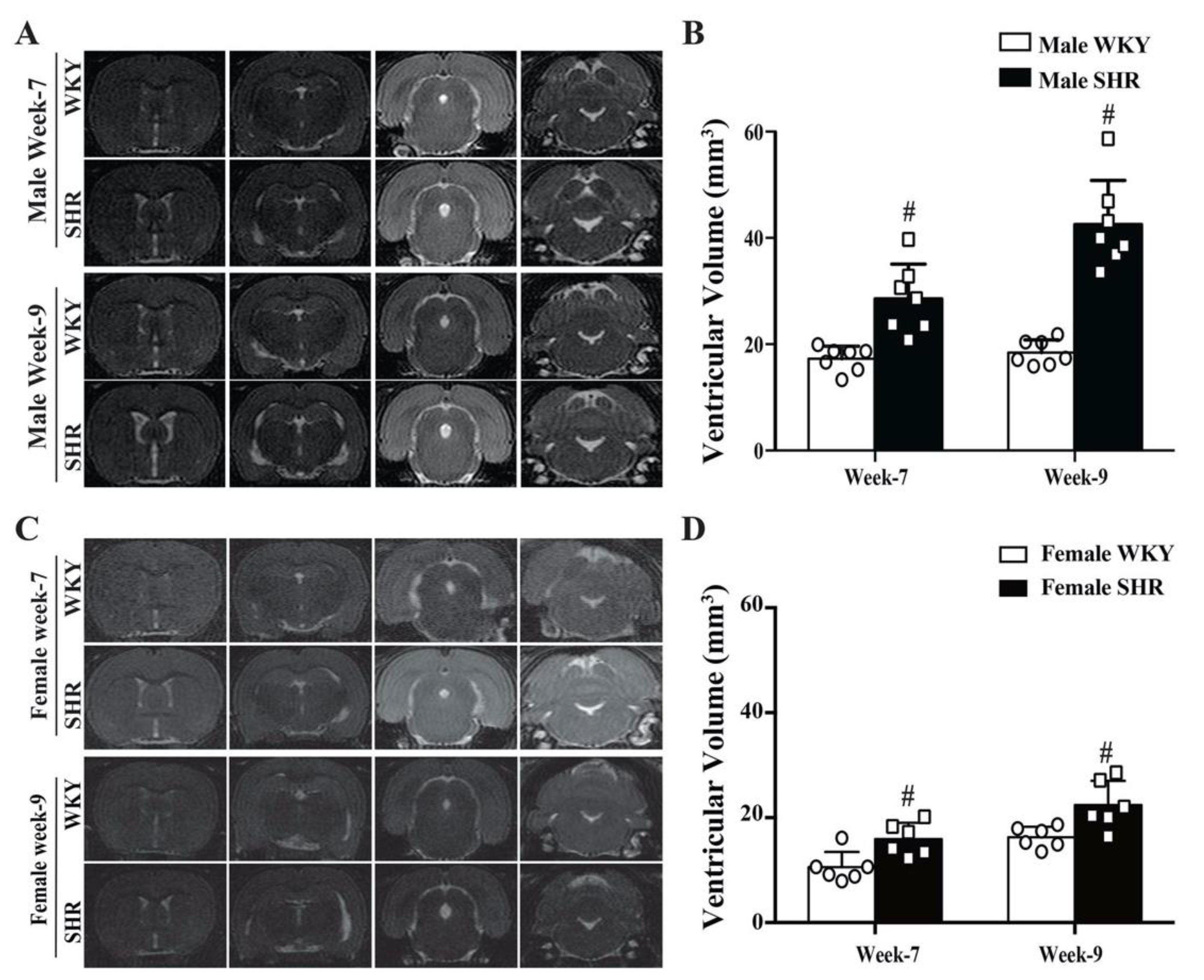
2.1. Spontaneous Hydrocephalus Developed in SHRs
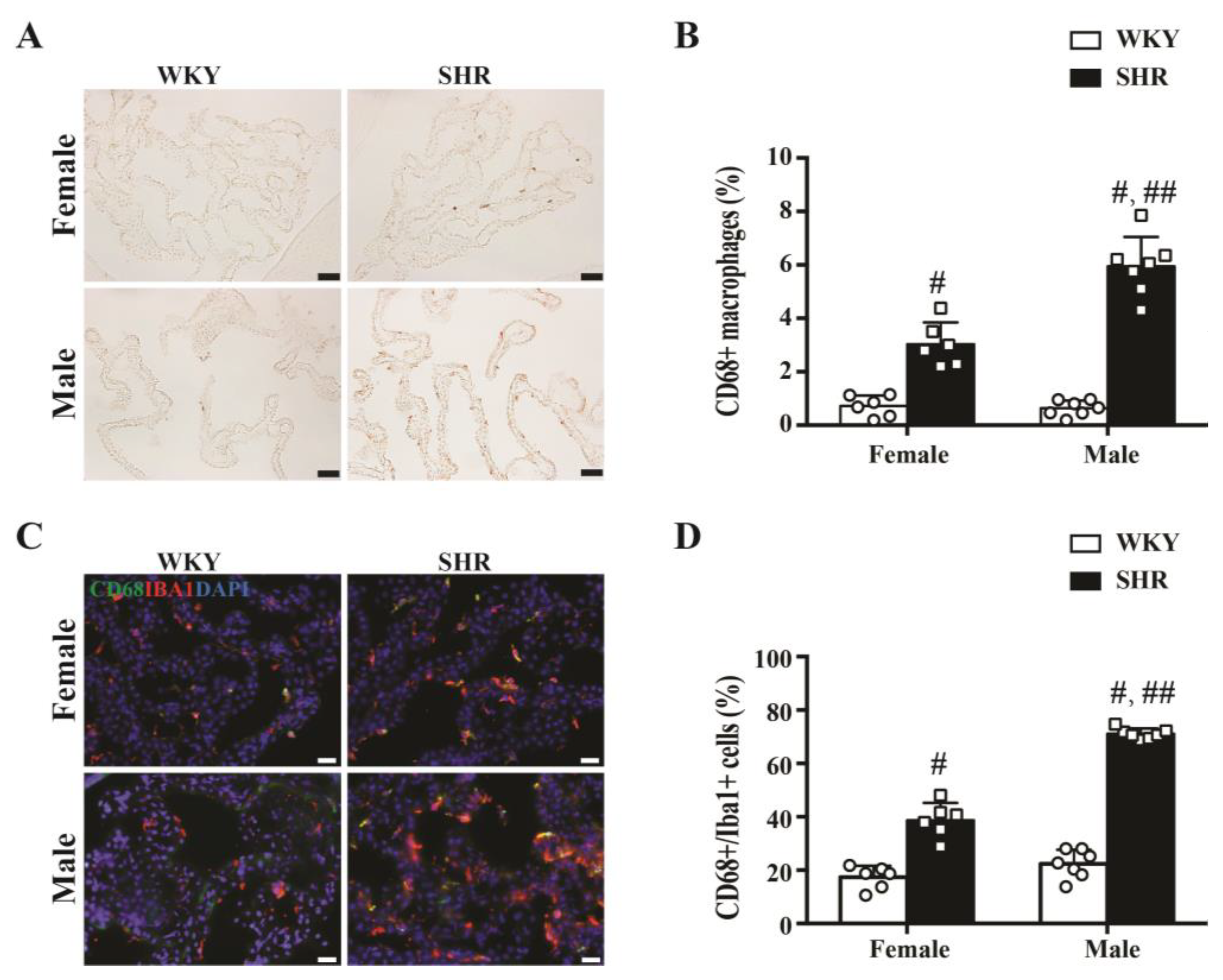
2.2. Activation of Epiplexus Macrophages in SHRs
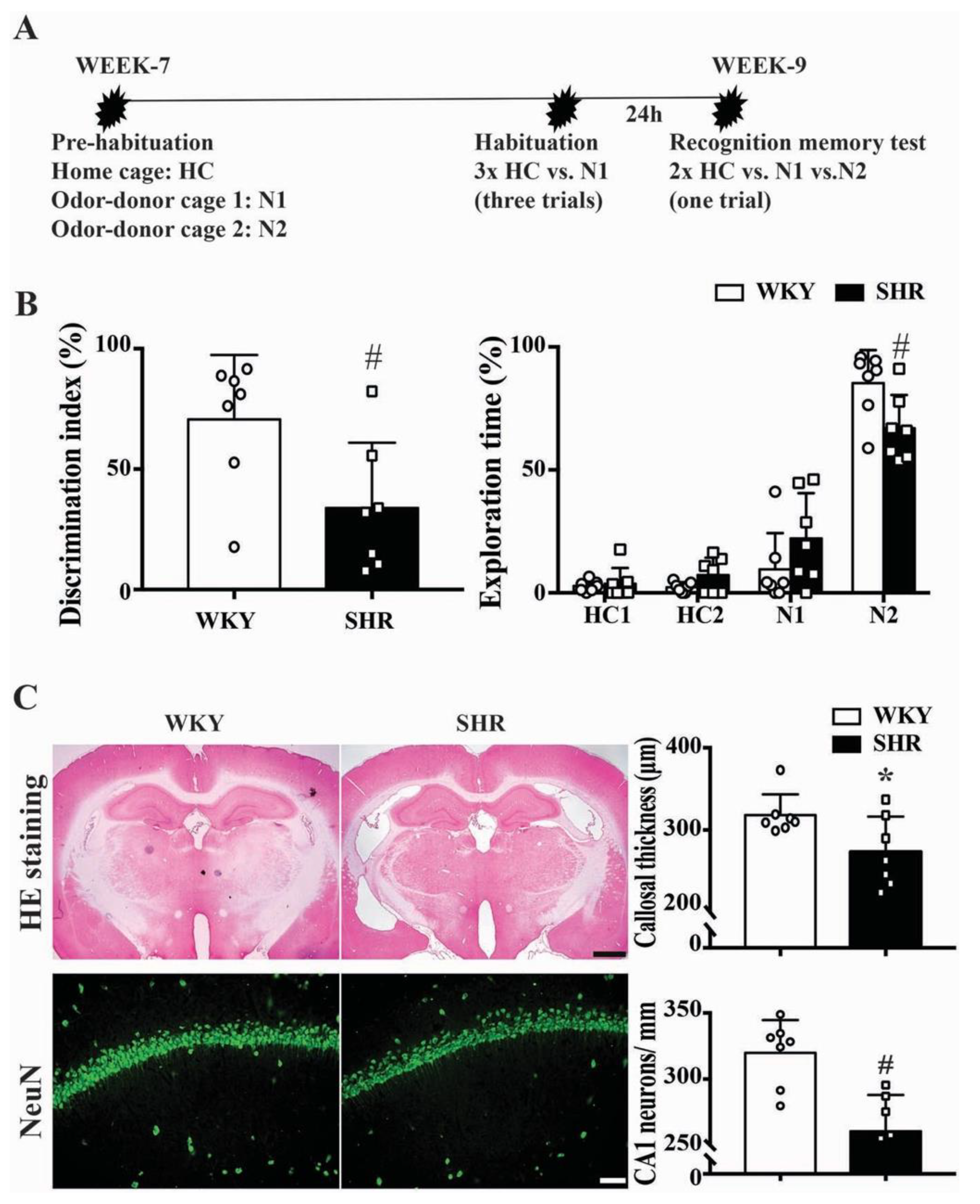
2.3. Cognitive Function and Neuronal Loss in Male SHRs
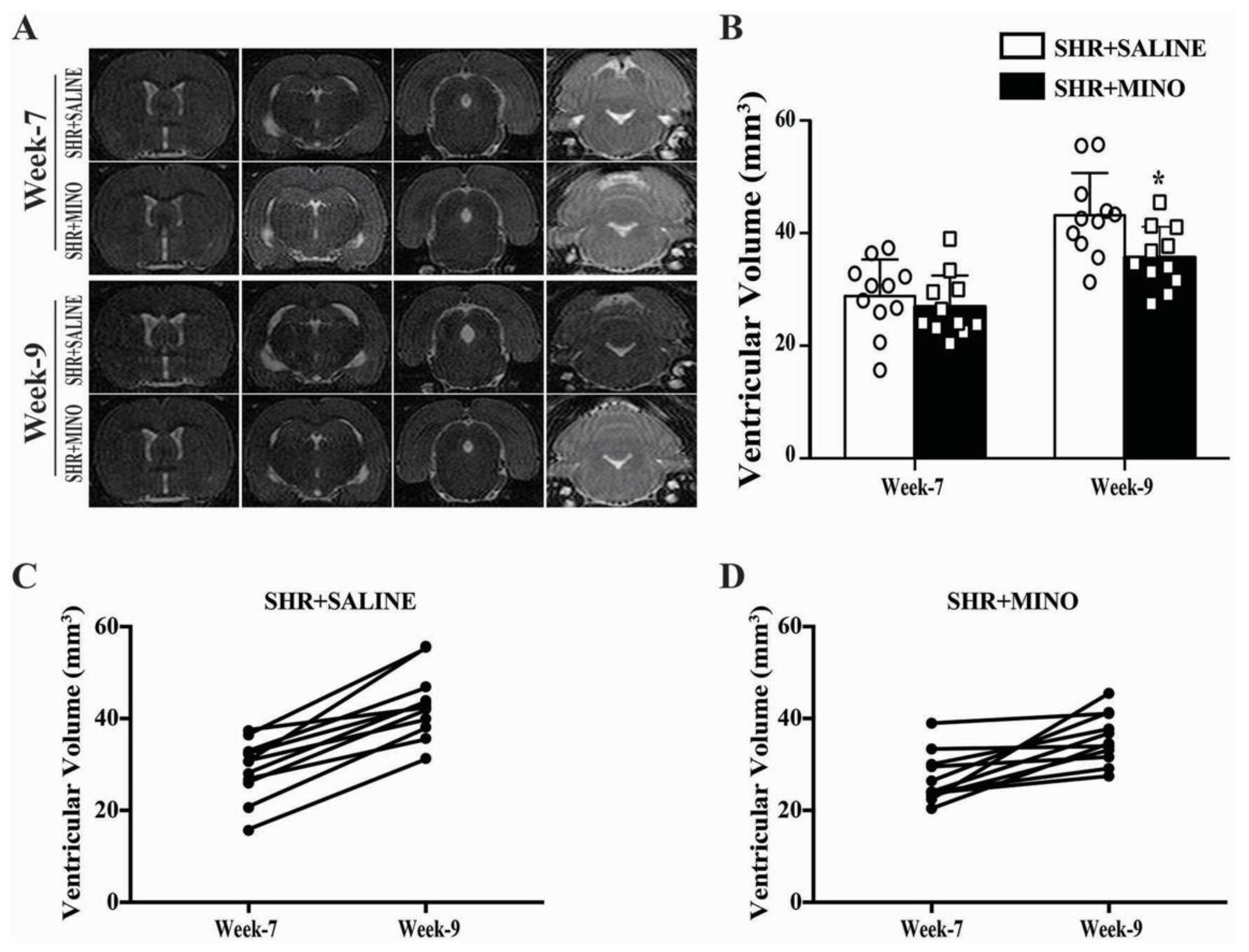
2.4. Hydrocephalus and Blood Pressure after Minocycline Treatment in Male SHRs
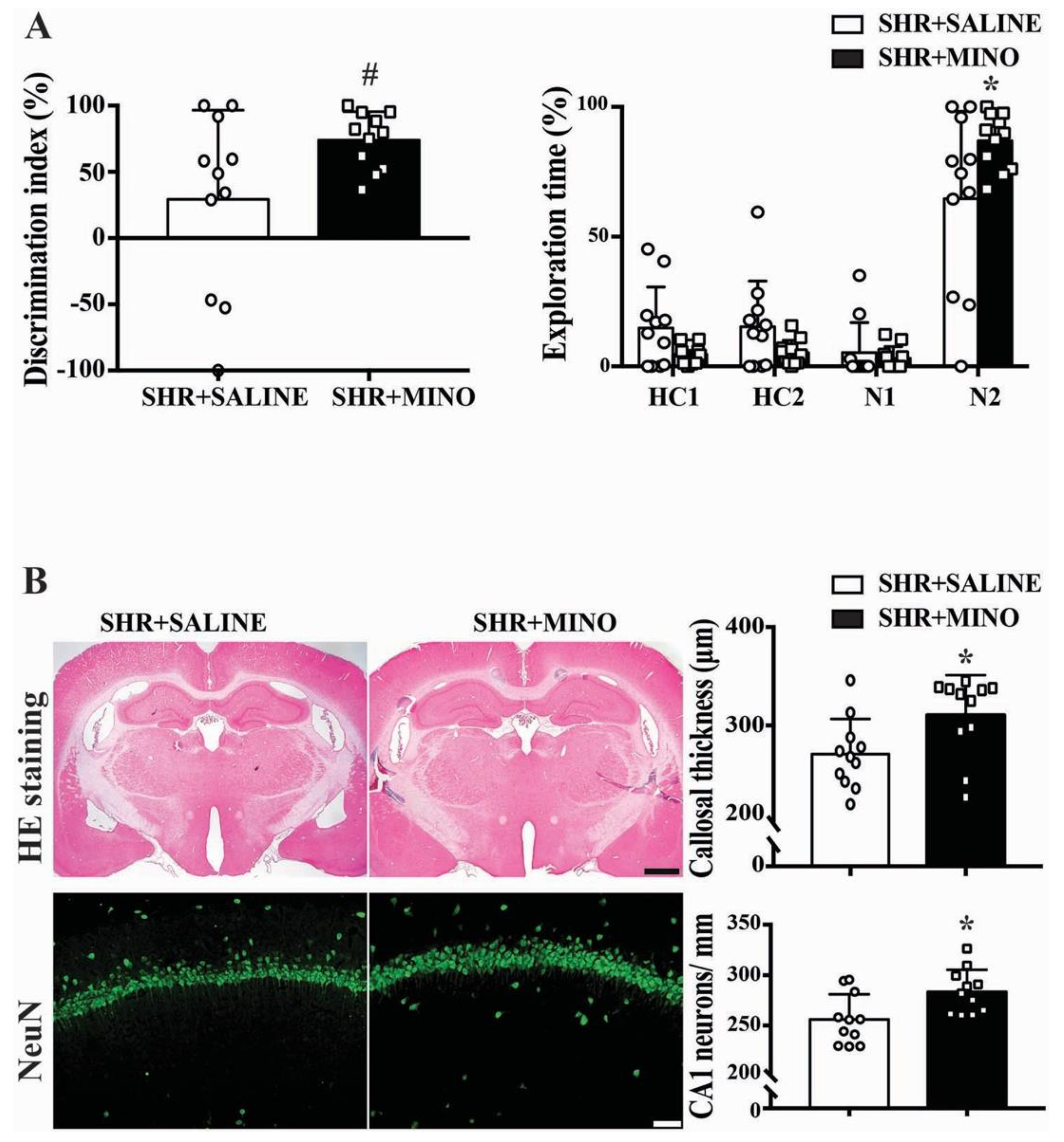
2.5. Cognitive Function and Neuron Loss after Minocycline Treatment
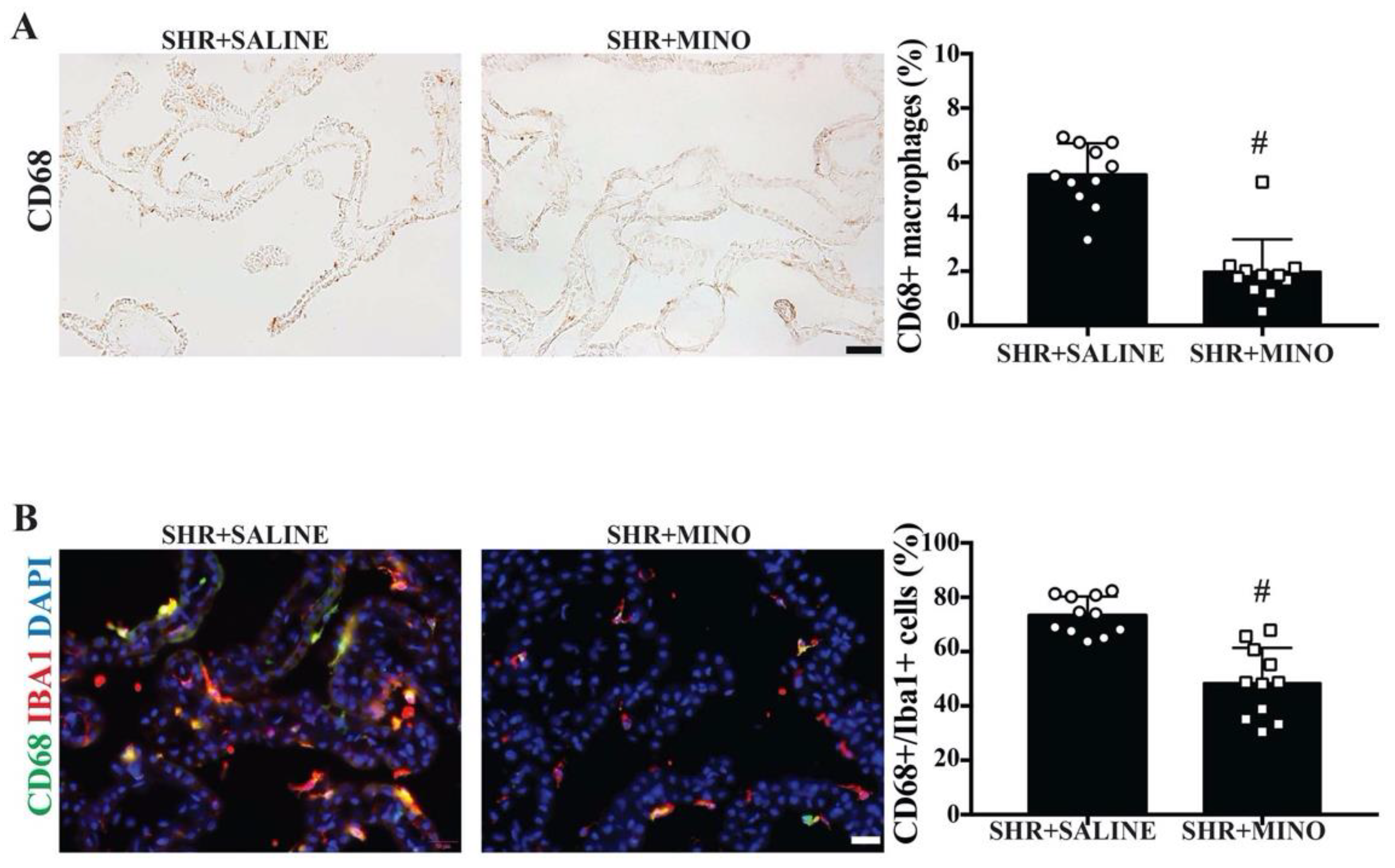
2.6. Activation of Epiplexus Macrophages after Minocycline Treatment
3. Discussion
4. Materials and Methods
4.1. The Animal Preparation
4.2. Experimental Groups
4.3. Minocycline Administration
4.4. Magnetic Resonance Imaging (MRI) and Ventricle Volume Measurement
4.5. Immunohistochemistry and Hematoxylin and Eosin (HE) Staining
4.6. Novel Object Recognition Test
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hemphill, J.C., 3rd; Greenberg, S.M.; Anderson, C.S.; Becker, K.; Bendok, B.R.; Cushman, M.; Fung, G.L.; Goldstein, J.N.; Macdonald, R.L.; Mitchell, P.H.; et al. Guidelines for the Management of Spontaneous Intracerebral Hemorrhage: A Guideline for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2015, 46, 2032–2060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, Z.; Larrew, T.W.; Eskandari, R. Pediatric Hydrocephalus: Current State of Diagnosis and Treatment. Pediatr. Rev. 2016, 37, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Del Bigio, M.R.; Di Curzio, D.L. Nonsurgical therapy for hydrocephalus: A comprehensive and critical review. Fluids Barriers CNS 2016, 13, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, S.; Dinh, T.T. Progressive postnatal dilation of brain ventricles in spontaneously hypertensive rats. Brain Res. 1986, 370, 327–332. [Google Scholar] [CrossRef]
- Cao, A.H.; Yu, L.; Wang, Y.W.; Wang, J.M.; Yang, L.J.; Lei, G.F. Effects of methylphenidate on attentional set-shifting in a genetic model of attention-deficit/hyperactivity disorder. Behav. Brain Funct. 2012, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Meneses, A.; Castillo, C.; Ibarra, M.; Hong, E. Effects of aging and hypertension on learning, memory, and activity in rats. Physiol. Behav. 1996, 60, 341–345. [Google Scholar] [CrossRef]
- Kantak, K.M.; Singh, T.; Kerstetter, K.A.; Dembro, K.A.; Mutebi, M.M.; Harvey, R.C.; Deschepper, C.F.; Dwoskin, L.P. Advancing the spontaneous hypertensive rat model of attention deficit/hyperactivity disorder. Behav. Neurosci. 2008, 122, 340–357. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, D.; Weise, G.; Moller, K.; Scheibe, J.; Posel, C.; Baasch, S.; Gawlitza, M.; Lobsien, D.; Diederich, K.; Minnerup, J.; et al. Spontaneous white matter damage, cognitive decline and neuroinflammation in middle-aged hypertensive rats: An animal model of early-stage cerebral small vessel disease. Acta Neuropathol. Commun. 2014, 2, 169. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Hao, X.; Li, J.; Hua, Y.; Keep, R.F.; Xi, G. Effects of minocycline on epiplexus macrophage activation, choroid plexus injury and hydrocephalus development in spontaneous hypertensive rats. J. Cereb. Blood Flow Metab. 2019, 39, 1936–1948. [Google Scholar] [CrossRef]
- Wan, Y.; Hua, Y.; Garton, H.J.L.; Novakovic, N.; Keep, R.F.; Xi, G. Activation of epiplexus macrophages in hydrocephalus caused by subarachnoid hemorrhage and thrombin. CNS Neurosci. Ther. 2019, 25, 1134–1141. [Google Scholar] [CrossRef]
- Chen, T.; Tan, X.; Xia, F.; Hua, Y.; Keep, R.F.; Xi, G. Hydrocephalus Induced by Intraventricular Peroxiredoxin-2: The Role of Macrophages in the Choroid Plexus. Biomolecules 2021, 11, 654. [Google Scholar] [CrossRef] [PubMed]
- Koduri, S.; Daou, B.; Hua, Y.; Keep, R.; Xi, G.; Pandey, A.S. Mechanisms of Post-Hemorrhagic Stroke Hydrocephalus Development: The Role of Kolmer Epiplexus Cells. World Neurosurg. 2020, 144, 256–257. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Tan, G.; Zhang, S.; Zhu, H.; Liu, F.; Huang, C.; Zhang, F.; Wang, Z. Minocycline reduces reactive gliosis in the rat model of hydrocephalus. BMC Neurosci. 2012, 13, 148. [Google Scholar] [CrossRef] [Green Version]
- McAllister, J.P., 2nd; Miller, J.M. Minocycline inhibits glial proliferation in the H-Tx rat model of congenital hydrocephalus. Cerebrospinal Fluid Res. 2010, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Chen, Q.; Tang, J.; Zhang, J.; Tao, Y.; Li, L.; Zhu, G.; Feng, H.; Chen, Z. Minocycline-induced attenuation of iron overload and brain injury after experimental germinal matrix hemorrhage. Brain Res. 2015, 1594, 115–124. [Google Scholar] [CrossRef]
- Bushnell, C.D.; Chaturvedi, S.; Gage, K.R.; Herson, P.S.; Hurn, P.D.; Jimenez, M.C.; Kittner, S.J.; Madsen, T.E.; McCullough, L.D.; McDermott, M.; et al. Sex differences in stroke: Challenges and opportunities. J. Cereb. Blood Flow Metab. 2018, 38, 2179–2191. [Google Scholar] [CrossRef]
- Hurn, P.D. 2014 Thomas Willis Award Lecture: Sex, stroke, and innovation. Stroke 2014, 45, 3725–3729. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.M.; Mulcahey, T.A.; Filipek, N.C.; Wise, P.M. Production of proinflammatory cytokines and chemokines during neuroinflammation: Novel roles for estrogen receptors alpha and beta. Endocrinology 2010, 151, 4916–4925. [Google Scholar] [CrossRef] [Green Version]
- Vegeto, E.; Benedusi, V.; Maggi, A. Estrogen anti-inflammatory activity in brain: A therapeutic opportunity for menopause and neurodegenerative diseases. Front. Neuroendocrinol. 2008, 29, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Tu, T.W.; Turtzo, L.C.; Williams, R.A.; Lescher, J.D.; Dean, D.D.; Frank, J.A. Imaging of spontaneous ventriculomegaly and vascular malformations in Wistar rats: Implications for preclinical research. J. Neuropathol. Exp. Neurol. 2014, 73, 1152–1165. [Google Scholar] [CrossRef] [Green Version]
- Guo, D.; Wilkinson, D.A.; Thompson, B.G.; Pandey, A.S.; Keep, R.F.; Xi, G.; Hua, Y. MRI Characterization in the Acute Phase of Experimental Subarachnoid Hemorrhage. Transl. Stroke Res. 2017, 8, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.J.; Klebe, D.; Tang, J.; Zhang, J.H. A comprehensive review of therapeutic targets that induce microglia/macrophage-mediated hematoma resolution after germinal matrix hemorrhage. J. Neurosci. Res. 2020, 98, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritter, S.; Dinh, T.T.; Stone, S.; Ross, N. Cerebroventricular dilation in spontaneously hypertensive rats (SHRs) is not attenuated by reduction of blood pressure. Brain Res. 1988, 450, 354–359. [Google Scholar] [CrossRef]
- Gonzalez-Marrero, I.; Hernandez-Abad, L.G.; Castaneyra-Ruiz, L.; Carmona-Calero, E.M.; Castaneyra-Perdomo, A. Changes in the choroid plexuses and brain barriers associated with high blood pressure and ageing. Neurologia (Engl. Ed.) 2018. [Google Scholar] [CrossRef]
- González-Marrero, I.; Castañeyra-Ruiz, L.; González-Toledo, J.M.; Castañeyra-Ruiz, A.; De Paz-Carmona, H.; Ruiz-Mayor, L.; Castañeyra-Perdomo, A.; Carmona Calero, E. High Blood Pressure Effects on the Brain Barriers and Choroid Plexus Secretion. Neurosci. Med. 2012, 3, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Palta, P.; Sharrett, A.R.; Wei, J.; Meyer, M.L.; Kucharska-Newton, A.; Power, M.C.; Deal, J.A.; Jack, C.R.; Knopman, D.; Wright, J.; et al. Central Arterial Stiffness Is Associated With Structural Brain Damage and Poorer Cognitive Performance: The ARIC Study. J. Am. Heart Assoc. 2019, 8, e011045. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini, M.; Catalani, A.; Consoli, C.; Marletta, N.; Tomassoni, D.; Avola, R. The hippocampus in spontaneously hypertensive rats: An animal model of vascular dementia? Mech. Ageing Dev. 2002, 123, 547–559. [Google Scholar] [CrossRef]
- Yang, Y.; Kimura-Ohba, S.; Thompson, J.; Rosenberg, G.A. Rodent Models of Vascular Cognitive Impairment. Transl. Stroke Res. 2016, 7, 407–414. [Google Scholar] [CrossRef]
- Sagvolden, T.; Russell, V.A.; Aase, H.; Johansen, E.B.; Farshbaf, M. Rodent models of attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1239–1247. [Google Scholar] [CrossRef]
- Sagvolden, T. Behavioral validation of the spontaneously hypertensive rat (SHR) as an animal model of attention-deficit/hyperactivity disorder (AD/HD). Neurosci. Biobehav. Rev. 2000, 24, 31–39. [Google Scholar] [CrossRef]
- Greven, C.U.; Bralten, J.; Mennes, M.; O’dwyer, L.; van Hulzen, K.J.; Rommelse, N.; Schweren, L.J.; Hoekstra, P.J.; Hartman, C.A.; Heslenfeld, D. Developmentally stable whole-brain volume reductions and developmentally sensitive caudate and putamen volume alterations in those with attention-deficit/hyperactivity disorder and their unaffected siblings. JAMA Psychiatry 2015, 72, 490–499. [Google Scholar] [CrossRef] [Green Version]
- Hoogman, M.; Bralten, J.; Hibar, D.P.; Mennes, M.; Zwiers, M.P.; Schweren, L.S.J.; van Hulzen, K.J.E.; Medland, S.E.; Shumskaya, E.; Jahanshad, N.; et al. Subcortical brain volume differences in participants with attention deficit hyperactivity disorder in children and adults: A cross-sectional mega-analysis. Lancet Psychiatry 2017, 4, 310–319. [Google Scholar] [CrossRef] [Green Version]
- van Leijsen, E.M.; Tay, J.; van Uden, I.W.; Kooijmans, E.C.; Bergkamp, M.I.; van der Holst, H.M.; Ghafoorian, M.; Platel, B.; Norris, D.G.; Kessels, R.P. Memory decline in elderly with cerebral small vessel disease explained by temporal interactions between white matter hyperintensities and hippocampal atrophy. Hippocampus 2018, 29, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Kril, J.J.; Patel, S.; Harding, A.J.; Halliday, G.M. Patients with vascular dementia due to microvascular pathology have significant hippocampal neuronal loss. J. Neurol. Neurosurg. Psychiatry 2002, 72, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimy, J.K.; Zhang, J.; Kurland, D.B.; Theriault, B.C.; Duran, D.; Stokum, J.A.; Furey, C.G.; Zhou, X.; Mansuri, M.S.; Montejo, J.; et al. Inflammation-dependent cerebrospinal fluid hypersecretion by the choroid plexus epithelium in posthemorrhagic hydrocephalus. Nat. Med. 2017, 23, 997–1003. [Google Scholar] [CrossRef]
- Albanese, A.; Tuttolomondo, A.; Anile, C.; Sabatino, G.; Pompucci, A.; Pinto, A.; Licata, G.; Mangiola, A. Spontaneous chronic subdural hematomas in young adults with a deficiency in coagulation factor XIII. Report of three cases. J. Neurosurg. 2005, 102, 1130–1132. [Google Scholar] [CrossRef]
- Della Corte, V.; Tuttolomondo, A.; Pecoraro, R.; Di Raimondo, D.; Vassallo, V.; Pinto, A. Inflammation, Endothelial Dysfunction and Arterial Stiffness as Therapeutic Targets in Cardiovascular Medicine. Curr. Pharm. Des. 2016, 22, 4658–4668. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Casuccio, A.; Di Bona, D.; Aiello, A.; Accardi, G.; Arnao, V.; Clemente, G.; Corte, V.D.; et al. HLA and killer cell immunoglobulin-like receptor (KIRs) genotyping in patients with acute ischemic stroke. J. Neuroinflamm. 2019, 16, 88. [Google Scholar] [CrossRef] [Green Version]
- Botfield, H.; Gonzalez, A.M.; Abdullah, O.; Skjolding, A.D.; Berry, M.; McAllister, J.P., 2nd; Logan, A. Decorin prevents the development of juvenile communicating hydrocephalus. Brain 2013, 136, 2842–2858. [Google Scholar] [CrossRef] [Green Version]
- Ulfig, N.; Bohl, J.; Neudorfer, F.; Rezaie, P. Brain macrophages and microglia in human fetal hydrocephalus. Brain Dev. 2004, 26, 307–315. [Google Scholar] [CrossRef]
- Lattke, M.; Magnutzki, A.; Walther, P.; Wirth, T.; Baumann, B. Nuclear factor kappaB activation impairs ependymal ciliogenesis and links neuroinflammation to hydrocephalus formation. J. Neurosci. 2012, 32, 11511–11523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Hua, Y.; Keep, R.F.; Chaudhary, N.; Xi, G. Minocycline Effects on Intracerebral Hemorrhage-Induced Iron Overload in Aged Rats: Brain Iron Quantification With Magnetic Resonance Imaging. Stroke 2018, 49, 995–1002. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Hua, Y.; Keep, R.F.; Novakovic, N.; Fei, Z.; Xi, G. Minocycline attenuates brain injury and iron overload after intracerebral hemorrhage in aged female rats. Neurobiol. Dis. 2019, 126, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Hanley, D.F.; Thompson, R.E.; Muschelli, J.; Rosenblum, M.; McBee, N.; Lane, K.; Bistran-Hall, A.J.; Mayo, S.W.; Keyl, P.; Gandhi, D.; et al. Safety and efficacy of minimally invasive surgery plus alteplase in intracerebral haemorrhage evacuation (MISTIE): A randomised, controlled, open-label, phase 2 trial. Lancet Neurol. 2016, 15, 1228–1237. [Google Scholar] [CrossRef] [Green Version]
- Brundula, V.; Rewcastle, N.B.; Metz, L.M.; Bernard, C.C.; Yong, V.W. Targeting leukocyte MMPs and transmigration: Minocycline as a potential therapy for multiple sclerosis. Brain 2002, 125, 1297–1308. [Google Scholar] [CrossRef]
- Yong, V.W.; Wells, J.; Giuliani, F.; Casha, S.; Power, C.; Metz, L.M. The promise of minocycline in neurology. Lancet Neurol. 2004, 3, 744–751. [Google Scholar] [CrossRef]
- Kreisel, T.; Frank, M.G.; Licht, T.; Reshef, R.; Ben-Menachem-Zidon, O.; Baratta, M.V.; Maier, S.F.; Yirmiya, R. Dynamic microglial alterations underlie stress-induced depressive-like behavior and suppressed neurogenesis. Mol. Psychiatry 2014, 19, 699–709. [Google Scholar] [CrossRef] [Green Version]
- Papa, S.; Rossi, F.; Ferrari, R.; Mariani, A.; De Paola, M.; Caron, I.; Fiordaliso, F.; Bisighini, C.; Sammali, E.; Colombo, C.; et al. Selective nanovector mediated treatment of activated proinflammatory microglia/macrophages in spinal cord injury. ACS Nano 2013, 7, 9881–9895. [Google Scholar] [CrossRef]
- Zhao, F.; Hua, Y.; He, Y.; Keep, R.F.; Xi, G. Minocycline-induced attenuation of iron overload and brain injury after experimental intracerebral hemorrhage. Stroke 2011, 42, 3587–3593. [Google Scholar] [CrossRef] [Green Version]
- Ryu, J.K.; Franciosi, S.; Sattayaprasert, P.; Kim, S.U.; McLarnon, J.G. Minocycline inhibits neuronal death and glial activation induced by beta-amyloid peptide in rat hippocampus. Glia 2004, 48, 85–90. [Google Scholar] [CrossRef]
- Suzuki, H.; Sugimura, Y.; Iwama, S.; Suzuki, H.; Nobuaki, O.; Nagasaki, H.; Arima, H.; Sawada, M.; Oiso, Y. Minocycline prevents osmotic demyelination syndrome by inhibiting the activation of microglia. J. Am. Soc. Nephrol. 2010, 21, 2090–2098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, M.; Biala, G. The novel object recognition memory: Neurobiology, test procedure, and its modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monaghan, M.M.; Leddy, L.; Sung, M.L.; Albinson, K.; Kubek, K.; Pangalos, M.N.; Reinhart, P.H.; Zaleska, M.M.; Comery, T.A. Social odor recognition: A novel behavioral model for cognitive dysfunction in Parkinson’s disease. Neurodegener. Dis. 2010, 7, 153–159. [Google Scholar] [CrossRef]
- O’Dell, S.J.; Feinberg, L.M.; Marshall, J.F. A neurotoxic regimen of methamphetamine impairs novelty recognition as measured by a social odor-based task. Behav. Brain Res. 2011, 216, 396–401. [Google Scholar] [CrossRef]
- Liu, X.; Fan, B.; Szalad, A.; Wang, L.; Zhang, R.; Chopp, M.; Zhang, Z.G. Influence of sex on cognition and peripheral neurovascular function in diabetic mice. Front. Neurosci. 2018, 12, 795. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, X.; Ye, F.; Holste, K.G.; Hua, Y.; Garton, H.J.L.; Keep, R.F.; Xi, G. Delayed Minocycline Treatment Ameliorates Hydrocephalus Development and Choroid Plexus Inflammation in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2022, 23, 2306. https://doi.org/10.3390/ijms23042306
Hao X, Ye F, Holste KG, Hua Y, Garton HJL, Keep RF, Xi G. Delayed Minocycline Treatment Ameliorates Hydrocephalus Development and Choroid Plexus Inflammation in Spontaneously Hypertensive Rats. International Journal of Molecular Sciences. 2022; 23(4):2306. https://doi.org/10.3390/ijms23042306
Chicago/Turabian StyleHao, Xiaodi, Fenghui Ye, Katherine G. Holste, Ya Hua, Hugh J. L. Garton, Richard F. Keep, and Guohua Xi. 2022. "Delayed Minocycline Treatment Ameliorates Hydrocephalus Development and Choroid Plexus Inflammation in Spontaneously Hypertensive Rats" International Journal of Molecular Sciences 23, no. 4: 2306. https://doi.org/10.3390/ijms23042306