
Small Molecule Compounds of Natural Origin Target Cellular Receptors to Inhibit Cancer Development and Progression


Abstract
:1. Introduction
2. Natural Small Molecule Compound Targeting Receptor and Its Anti-Cancer Activity
2.1. Ion Channel Coupled Receptor
2.1.1. Alkaloid Compounds Inhibiting Ion Channel Receptor P2X7R
2.1.2. Flavonoids Inhibiting Ion Channel Receptor nAChRs
2.2. Enzyme-Linked Receptor-Tyrosine Kinases
2.2.1. Small Natural Compounds Inhibiting Epidermal Growth Factor Receptor
2.2.2. Small Natural Compounds Inhibiting Vascular Endothelial Growth Factor Receptors
2.2.3. Flavonoids and Polyphenols Inhibiting FLT3, c-Met, EphA2R
2.3. G Protein-Coupled Receptors
2.3.1. Small Natural Compounds Inhibiting Protease-Activated Receptor-2
2.3.2. Small Natural Compounds Inhibiting Prostaglandin E Receptors
2.3.3. Flavonoids Inhibiting Cannabinoid Receptors
2.3.4. Small Natural Compounds Inhibiting Frizzled Receptors
2.4. Nuclear Receptor Superfamily
Polyphenols Inhibiting Estrogen Receptor and Androgen Receptor
2.5. Other Receptors
2.5.1. Small Natural Compounds Combined with TRAIL to Inhibit the Death Receptor DR5
2.5.2. Small Natural Compounds Inhibiting Peroxisome Proliferator-Activated Receptors and Toll-Like Receptor 4
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer 2021, 127, 3029–3030. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Soerjomataram, I.; Bray, F. Planning for tomorrow: Global cancer incidence and the role of prevention 2020–2070. Nat. Rev. Clin. Oncol. 2021, 18, 663–672. [Google Scholar] [CrossRef]
- Tomas, A.; Futter, C.E.; Eden, E.R. EGF receptor trafficking: Consequences for signaling and cancer. Trends Cell Biol 2014, 24, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Thomas, C.; Gustafsson, J. The different roles of ER subtypes in cancer biology and therapy. Nat. Rev. Cancer 2011, 11, 597–608. [Google Scholar] [CrossRef]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat. Rev. Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef]
- Herz, J.M.; Thomsen, W.J.; Yarbrough, G.G. Molecular approaches to receptors as targets for drug discovery. J. Recept. Signal. Transduct. Res. 1997, 17, 671–776. [Google Scholar] [CrossRef]
- Serrano-Marín, J.; Reyes-Resina, I.; Martínez-Pinilla, E.; Navarro, G.; Franco, R. Natural Compounds as Guides for the Discovery of Drugs Targeting G-Protein-Coupled Receptors. Molecules 2020, 25, 5060. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Lappano, R.; Maggiolini, M. G protein-coupled receptors: Novel targets for drug discovery in cancer. Nat. Rev. Drug Discov. 2011, 10, 47–60. [Google Scholar] [CrossRef]
- Nieto Gutierrez, A.; McDonald, P.H. GPCRs: Emerging anti-cancer drug targets. Cell Signal. 2018, 41, 65–74. [Google Scholar] [CrossRef]
- Liu, Y.; An, S.; Ward, R.; Yang, Y.; Guo, X.X.; Li, W.; Xu, T.R. G protein-coupled receptors as promising cancer targets. Cancer Lett. 2016, 376, 226–239. [Google Scholar] [CrossRef]
- Allen, L.F.; Lefkowitz, R.J.; Caron, M.G.; Cotecchia, S. G-protein-coupled receptor genes as protooncogenes: Constitutively activating mutation of the alpha 1B-adrenergic receptor enhances mitogenesis and tumorigenicity. Proc. Natl. Acad. Sci. USA 1991, 88, 11354–11358. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z. ErbB Receptors and Cancer. Methods Mol. Biol. 2017, 1652, 3–35. [Google Scholar]
- Bublil, E.M.; Yarden, Y. The EGF receptor family: Spearheading a merger of signaling and therapeutics. Curr. Opin. Cell Biol. 2007, 19, 124–134. [Google Scholar] [CrossRef]
- Cho, H.S.; Leahy, D.J. Structure of the extracellular region of HER3 reveals an interdomain tether. Science 2002, 297, 1330–1333. [Google Scholar] [CrossRef]
- Kumagai, S.; Koyama, S.; Nishikawa, H. Antitumour immunity regulated by aberrant ERBB family signalling. Nat. Rev. Cancer 2021, 21, 181–197. [Google Scholar] [CrossRef]
- Arteaga, C.L.; Engelman, J.A. ERBB receptors: From oncogene discovery to basic science to mechanism-based cancer therapeutics. Cancer Cell 2014, 25, 282–303. [Google Scholar] [CrossRef] [Green Version]
- Baselga, J.; Arteaga, C.L. Critical update and emerging trends in epidermal growth factor receptor targeting in cancer. J. Clin. Oncol. 2005, 23, 2445–2459. [Google Scholar] [CrossRef]
- Harari, P.M.; Allen, G.W.; Bonner, J.A. Biology of interactions: Antiepidermal growth factor receptor agents. J. Clin. Oncol. 2007, 25, 4057–4065. [Google Scholar] [CrossRef]
- Ono, M.; Kuwano, M. Molecular mechanisms of epidermal growth factor receptor (EGFR) activation and response to gefitinib and other EGFR-targeting drugs. Clin. Cancer Res. 2006, 12, 7242–7251. [Google Scholar] [CrossRef] [Green Version]
- Nyati, M.K.; Morgan, M.A.; Feng, F.Y.; Lawrence, T.S. Integration of EGFR inhibitors with radiochemotherapy. Nat. Rev. Cancer 2006, 6, 876–885. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Muratspahić, E.; Freissmuth, M.; Gruber, C.W. Nature-Derived Peptides: A Growing Niche for GPCR Ligand Discovery. Trends Pharmacol. Sci. 2019, 40, 309–326. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Song, X.; Guo, Y.; Song, P.; Duan, D.; Chen, Z.S. Natural Products: A Promising Therapeutics for Targeting Tumor Angiogenesis. Front. Oncol. 2021, 11, 772915. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, T.; Wang, Y.; Jiang, Y.; Wang, Y. Mechanisms and Advances in Anti-Ovarian Cancer with Natural Plants Component. Molecules 2021, 26, 5949. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Yang, D.; Li, Z.; Zhang, X.; Pu, L. Co-delivery of paclitaxel and tanespimycin in lipid nanoparticles enhanced anti-gastric-tumor effect in vitro and in vivo. Artif. Cells Nanomed. Biotechnol. 2018, 46, 904–911. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.M.; Stow, J.L. Nuclear translocation of cell-surface receptors: Lessons from fibroblast growth factor. Traffic 2005, 6, 947–954. [Google Scholar] [CrossRef]
- Jiang, L.H.; Adinolfi, E.; Roger, S. Editorial: Ion Channel Signalling in Cancer: From Molecular Mechanisms to Therapeutics. Front. Pharmacol. 2021, 12, 711593. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Fan, X.; Yuan, B.; Takagi, N.; Liu, S.; Han, X.; Ren, J.; Liu, J. Berberine inhibits NLRP3 Inflammasome pathway in human triple-negative breast cancer MDA-MB-231 cell. BMC Complement. Altern. Med. 2019, 19, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelassi, B.; Anchelin, M.; Chamouton, J.; Cayuela, M.L.; Clarysse, L.; Li, J.; Goré, J.; Jiang, L.H.; Roger, S. Anthraquinone emodin inhibits human cancer cell invasiveness by antagonizing P2X7 receptors. Carcinogenesis 2013, 34, 1487–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, K.F.; Gutierres, J.M.; Pillat, M.M.; Rissi, V.B.; Santos Araújo, M.D.; Bertol, G.; Gonçalves, P.B.; Schetinger, M.R.; Morsch, V.M. Uncaria tomentosa extract alters the catabolism of adenine nucleotides and expression of ecto-5’-nucleotidase/CD73 and P2X7 and A1 receptors in the MDA-MB-231 cell line. J. Ethnopharmacol. 2016, 194, 108–116. [Google Scholar] [CrossRef]
- Shih, Y.L.; Liu, H.C.; Chen, C.S.; Hsu, C.H.; Pan, M.H.; Chang, H.W.; Chang, C.H.; Chen, F.C.; Ho, C.T.; Yang, Y.Y.; et al. Combination treatment with luteolin and quercetin enhances antiproliferative effects in nicotine-treated MDA-MB-231 cells by down-regulating nicotinic acetylcholine receptors. J. Agric. Food Chem. 2010, 58, 235–241. [Google Scholar] [CrossRef]
- Tu, S.H.; Ku, C.Y.; Ho, C.T.; Chen, C.S.; Huang, C.S.; Lee, C.H.; Chen, L.C.; Pan, M.H.; Chang, H.W.; Chang, C.H.; et al. Tea polyphenol (-)-epigallocatechin-3-gallate inhibits nicotine- and estrogen-induced α9-nicotinic acetylcholine receptor upregulation in human breast cancer cells. Mol. Nutr. Food Res. 2011, 55, 455–466. [Google Scholar] [CrossRef]
- Chen, C.S.; Lee, C.H.; Hsieh, C.D.; Ho, C.T.; Pan, M.H.; Huang, C.S.; Tu, S.H.; Wang, Y.J.; Chen, L.C.; Chang, Y.J.; et al. Nicotine-induced human breast cancer cell proliferation attenuated by garcinol through down-regulation of the nicotinic receptor and cyclin D3 proteins. Breast Cancer Res. Treat. 2011, 125, 73–87. [Google Scholar] [CrossRef]
- Hou, X.; Du, H.; Quan, X.; Shi, L.; Zhang, Q.; Wu, Y.; Liu, Y.; Xiao, J.; Li, Y.; Lu, L.; et al. Silibinin Inhibits NSCLC Metastasis by Targeting the EGFR/LOX Pathway. Front. Pharmacol. 2018, 9, 21. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, L.; Zhang, J.; Dai, B.; Wang, N.; Chen, Y.; He, L. Antitumor activity of taspine by modulating the EGFR signaling pathway of Erk1/2 and Akt in vitro and in vivo. Planta Med. 2011, 77, 1774–1781. [Google Scholar] [CrossRef]
- Wang, Y.; Romigh, T.; He, X.; Orloff, M.S.; Silverman, R.H.; Heston, W.D.; Eng, C. Resveratrol regulates the PTEN/AKT pathway through androgen receptor-dependent and -independent mechanisms in prostate cancer cell lines. Hum. Mol. Genet. 2010, 19, 4319–4329. [Google Scholar] [CrossRef]
- Leeman-Neill, R.J.; Cai, Q.; Joyce, S.C.; Thomas, S.M.; Bhola, N.E.; Neill, D.B.; Arbiser, J.L.; Grandis, J.R. Honokiol inhibits epidermal growth factor receptor signaling and enhances the antitumor effects of epidermal growth factor receptor inhibitors. Clin. Cancer Res. 2010, 16, 2571–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.J.; Kim, M.J.; Kim, Y.K.; Kim, S.M.; Park, J.Y.; Myoung, H. Combined cetuximab and genistein treatment shows additive anti-cancer effect on oral squamous cell carcinoma. Cancer Lett. 2010, 292, 54–63. [Google Scholar] [CrossRef]
- Rho, J.K.; Choi, Y.J.; Jeon, B.S.; Choi, S.J.; Cheon, G.J.; Woo, S.K.; Kim, H.R.; Kim, C.H.; Choi, C.M.; Lee, J.C. Combined treatment with silibinin and epidermal growth factor receptor tyrosine kinase inhibitors overcomes drug resistance caused by T790M mutation. Mol. Cancer Ther. 2010, 9, 3233–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar] [PubMed]
- Chuang, T.C.; Wu, K.; Lin, Y.Y.; Kuo, H.P.; Kao, M.C.; Wang, V.; Hsu, S.C.; Lee, S.L. Dual down-regulation of EGFR and ErbB2 by berberine contributes to suppression of migration and invasion of human ovarian cancer cells. Environ. Toxicol. 2021, 36, 737–747. [Google Scholar] [CrossRef]
- Kim, Y.; Sugihara, Y.; Kim, T.Y.; Cho, S.M.; Kim, J.Y.; Lee, J.Y.; Yoo, J.S.; Song, D.; Han, G.; Rezeli, M.; et al. Identification and Validation of VEGFR2 Kinase as a Target of Voacangine by a Systematic Combination of DARTS and MSI. Biomolecules 2020, 10, 508. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Wang, Z.Y.; Mo, S.L.; Loo, T.Y.; Wang, D.M.; Luo, H.B.; Yang, D.P.; Chen, Y.L.; Shen, J.G.; Chen, J.P. Ellagic acid, a phenolic compound, exerts anti-angiogenesis effects via VEGFR-2 signaling pathway in breast cancer. Breast Cancer Res. Treat. 2012, 134, 943–955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceci, C.; Tentori, L.; Atzori, M.G.; Lacal, P.M.; Bonanno, E.; Scimeca, M.; Cicconi, R.; Mattei, M.; de Martino, M.G.; Vespasiani, G.; et al. Ellagic Acid Inhibits Bladder Cancer Invasiveness and In Vivo Tumor Growth. Nutrients 2016, 8, 744. [Google Scholar] [CrossRef]
- Liao, X.Z.; Tao, L.T.; Liu, J.H.; Gu, Y.Y.; Xie, J.; Chen, Y.; Lin, M.G.; Liu, T.L.; Wang, D.M.; Guo, H.Y.; et al. Matrine combined with cisplatin synergistically inhibited urothelial bladder cancer cells via down-regulating VEGF/PI3K/Akt signaling pathway. Cancer Cell Int. 2017, 17, 124. [Google Scholar] [CrossRef]
- Cao, Z.X.; Wen, Y.; He, J.L.; Huang, S.Z.; Gao, F.; Guo, C.J.; Liu, Q.Q.; Zheng, S.W.; Gong, D.Y.; Li, Y.Z.; et al. Isoliquiritigenin, an Orally Available Natural FLT3 Inhibitor from Licorice, Exhibits Selective Anti-Acute Myeloid Leukemia Efficacy In Vitro and In Vivo. Mol. Pharmacol. 2019, 96, 589–599. [Google Scholar] [CrossRef]
- Giorgio, C.; Mena, P.; Del Rio, D.; Brighenti, F.; Barocelli, E.; Hassan-Mohamed, I.; Callegari, D.; Lodola, A.; Tognolini, M. The ellagitannin colonic metabolite urolithin D selectively inhibits EphA2 phosphorylation in prostate cancer cells. Mol. Nutr. Food Res. 2015, 59, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Cheng, C.Y.; Su, T.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.; Kwan, H.Y.; Yu, H.; Yu, Z.L. Quercetin inhibits HGF/c-Met signaling and HGF-stimulated melanoma cell migration and invasion. Mol. Cancer 2015, 14, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Zhou, H.; Wang, T.; Mu, Y.; Wu, B.; Guo, D.L.; Zhang, X.M.; Wu, Y. Epigallocatechin-3-gallate inhibits proliferation and migration of human colon cancer SW620 cells in vitro. Acta Pharmacol. Sin. 2012, 33, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, S.M.; Min, K.J.; Chae, I.G.; Chun, K.S.; Kwon, T.K. Silymarin suppresses the PGE2 -induced cell migration through inhibition of EP2 activation; G protein-dependent PKA-CREB and G protein-independent Src-STAT3 signal pathways. Mol. Carcinog. 2015, 54, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Chang, Y.; Wei, W.; He, Y.F.; Hu, S.S.; Wang, D.; Wu, Y.J. Prostanoid EP1 receptor as the target of (-)-epigallocatechin-3-gallate in suppressing hepatocellular carcinoma cells in vitro. Acta Pharmacol. Sin. 2012, 33, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Kundu, N.; Collin, P.D.; Goloubeva, O.; Fulton, A.M. Frondoside A inhibits breast cancer metastasis and antagonizes prostaglandin E receptors EP4 and EP2. Breast Cancer Res. Treat. 2012, 132, 1001–1008. [Google Scholar] [CrossRef] [Green Version]
- Marcu, J.P.; Christian, R.T.; Lau, D.; Zielinski, A.J.; Horowitz, M.P.; Lee, J.; Pakdel, A.; Allison, J.; Limbad, C.; Moore, D.H.; et al. Cannabidiol enhances the inhibitory effects of delta9-tetrahydrocannabinol on human glioblastoma cell proliferation and survival. Mol. Cancer Ther. 2010, 9, 180–189. [Google Scholar] [CrossRef] [Green Version]
- Refolo, M.G.; D’Alessandro, R.; Malerba, N.; Laezza, C.; Bifulco, M.; Messa, C.; Caruso, M.G.; Notarnicola, M.; Tutino, V. Anti Proliferative and Pro Apoptotic Effects of Flavonoid Quercetin Are Mediated by CB1 Receptor in Human Colon Cancer Cell Lines. J. Cell Physiol. 2015, 230, 2973–2980. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, C.; Ma, T.; Jiang, L.; Tang, L.; Shi, T.; Zhang, S.; Zhang, L.; Zhu, P.; Li, J.; et al. Reversal effect of quercetin on multidrug resistance via FZD7/β-catenin pathway in hepatocellular carcinoma cells. Phytomedicine 2018, 43, 37–45. [Google Scholar] [CrossRef]
- Shi, X.; Zhu, M.; Kang, Y.; Yang, T.; Chen, X.; Zhang, Y. Wnt/β-catenin signaling pathway is involved in regulating the migration by an effective natural compound brucine in LoVo cells. Phytomedicine 2018, 46, 85–92. [Google Scholar] [CrossRef]
- Papoutsi, Z.; Kassi, E.; Tsiapara, A.; Fokialakis, N.; Chrousos, G.P.; Moutsatsou, P. Evaluation of estrogenic/antiestrogenic activity of ellagic acid via the estrogen receptor subtypes ERalpha and ERbeta. J. Agric. Food Chem. 2005, 53, 7715–7720. [Google Scholar] [CrossRef] [PubMed]
- Chiu, F.L.; Lin, J.K. Downregulation of androgen receptor expression by luteolin causes inhibition of cell proliferation and induction of apoptosis in human prostate cancer cells and xenografts. Prostate 2008, 68, 61–71. [Google Scholar] [CrossRef]
- Peng, Z.; Guan, Q.; Luo, J.; Deng, W.; Liu, J.; Yan, R.; Wang, W. Sophoridine exerts tumor-suppressive activities via promoting ESRRG-mediated β-catenin degradation in gastric cancer. BMC Cancer 2020, 20, 582. [Google Scholar] [CrossRef] [PubMed]
- Jung, E.M.; Lim, J.H.; Lee, T.J.; Park, J.W.; Choi, K.S.; Kwon, T.K. Curcumin sensitizes tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis through reactive oxygen species-mediated upregulation of death receptor 5 (DR5). Carcinogenesis 2005, 26, 1905–1913. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Meng, Y.; Sun, Q.; Zhang, Z.; Guo, X.; Sheng, X.; Tai, G.; Cheng, H.; Zhou, Y. Ginsenoside compound K sensitizes human colon cancer cells to TRAIL-induced apoptosis via autophagy-dependent and -independent DR5 upregulation. Cell Death Dis. 2016, 7, e2334. [Google Scholar] [CrossRef]
- Siraj, F.M.; SathishKumar, N.; Kim, Y.J.; Kim, S.Y.; Yang, D.C. Ginsenoside F2 possesses anti-obesity activity via binding with PPARγ and inhibiting adipocyte differentiation in the 3T3-L1 cell line. J. Enzyme Inhib. Med. Chem. 2015, 30, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, C.C.; Loo, S.Y.; Gee, V.; Yap, C.W.; Sethi, G.; Kumar, A.P.; Tan, K.H. Anticancer activity of thymoquinone in breast cancer cells: Possible involvement of PPAR-γ pathway. Biochem. Pharmacol. 2011, 82, 464–475. [Google Scholar] [CrossRef]
- Tan, B.S.; Kang, O.; Mai, C.W.; Tiong, K.H.; Khoo, A.S.; Pichika, M.R.; Bradshaw, T.D.; Leong, C.O. 6-Shogaol inhibits breast and colon cancer cell proliferation through activation of peroxisomal proliferator activated receptor γ (PPARγ). Cancer Lett. 2013, 336, 127–139. [Google Scholar] [CrossRef]
- Youn, H.S.; Saitoh, S.I.; Miyake, K.; Hwang, D.H. Inhibition of homodimerization of Toll-like receptor 4 by curcumin. Biochem. Pharmacol. 2006, 72, 62–69. [Google Scholar] [CrossRef]
- Pasqualetto, G.; Brancale, A.; Young, M.T. The Molecular Determinants of Small-Molecule Ligand Binding at P2X Receptors. Front. Pharmacol. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Virgilio, F.; Dal Ben, D.; Sarti, A.C.; Giuliani, A.L.; Falzoni, S. The P2X7 Receptor in Infection and Inflammation. Immunity 2017, 47, 15–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deli, T.; Varga, N.; Adám, A.; Kenessey, I.; Rásó, E.; Puskás, L.G.; Tóvári, J.; Fodor, J.; Fehér, M.; Szigeti, G.P.; et al. Functional genomics of calcium channels in human melanoma cells. Int. J. Cancer 2007, 121, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Solini, A.; Cuccato, S.; Ferrari, D.; Santini, E.; Gulinelli, S.; Callegari, M.G.; Dardano, A.; Faviana, P.; Madec, S.; Di Virgilio, F.; et al. Increased P2X7 receptor expression and function in thyroid papillary cancer: A new potential marker of the disease? Endocrinology 2008, 149, 389–396. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Wang, L.; Feng, Y.H.; Li, X.; Zeng, R.; Gorodeski, G.I. P2X7 receptor-mediated apoptosis of human cervical epithelial cells. Am. J. Physiol. Cell Physiol. 2004, 287, C1349–C1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zitvogel, L.; Kepp, O.; Galluzzi, L.; Kroemer, G. Inflammasomes in carcinogenesis and anticancer immune responses. Nat. Immunol. 2012, 13, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Di Virgilio, F. Liaisons dangereuses: P2X(7) and the inflammasome. Trends Pharmacol. Sci. 2007, 28, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zhang, J. Inflammasomes in Inflammation-Induced Cancer. Front. Immunol. 2017, 8, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, Y.; Du, X.; Ma, H.; Yao, J. The Anti-Cancer Mechanisms of Berberine: A Review. Cancer Manag. Res. 2020, 12, 695–702. [Google Scholar] [CrossRef] [Green Version]
- Mohammadinejad, R.; Ahmadi, Z.; Tavakol, S.; Ashrafizadeh, M. Berberine as a potential autophagy modulator. J. Cell Physiol. 2019, 234, 14914–14926. [Google Scholar] [CrossRef]
- Eo, S.H.; Kim, J.H.; Kim, S.J. Induction of G₂/M Arrest by Berberine via Activation of PI3K/Akt and p38 in Human Chondrosarcoma Cell Line. Oncol. Res. 2014, 22, 147–157. [Google Scholar] [CrossRef]
- Lu, L.; Huang, J.; Xue, X.; Wang, T.; Huang, Z.; Li, J. Berberine Regulated miR150-5p to Inhibit P2X7 Receptor, EMMPRIN and MMP-9 Expression in oxLDL Induced Macrophages. Front. Pharmacol. 2021, 12, 639558. [Google Scholar] [CrossRef] [PubMed]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A. Emodin-A natural anthraquinone derivative with diverse pharmacological activities. Phytochemistry 2021, 190, 112854. [Google Scholar] [CrossRef]
- Heitzman, M.E.; Neto, C.C.; Winiarz, E.; Vaisberg, A.J.; Hammond, G.B. Ethnobotany, phytochemistry and pharmacology of Uncaria (Rubiaceae). Phytochemistry 2005, 66, 5–29. [Google Scholar] [CrossRef]
- De Martino, L.; Martinot, J.L.; Franceschelli, S.; Leone, A.; Pizza, C.; De Feo, V. Proapoptotic effect of Uncaria tomentosa extracts. J. Ethnopharmacol. 2006, 107, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Pilarski, R.; Filip, B.; Wietrzyk, J.; Kuraś, M.; Gulewicz, K. Anticancer activity of the Uncaria tomentosa (Willd.) DC. preparations with different oxindole alkaloid composition. Phytomedicine 2010, 17, 1133–1139. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Huang, C.S.; Chen, C.S.; Tu, S.H.; Wang, Y.J.; Chang, Y.J.; Tam, K.W.; Wei, P.L.; Cheng, T.C.; Chu, J.S.; et al. Overexpression and activation of the alpha9-nicotinic receptor during tumorigenesis in human breast epithelial cells. J. Natl. Cancer Inst. 2010, 102, 1322–1335. [Google Scholar] [CrossRef] [PubMed]
- Schaal, C.; Padmanabhan, J.; Chellappan, S. The Role of nAChR and Calcium Signaling in Pancreatic Cancer Initiation and Progression. Cancers 2015, 7, 1447–1471. [Google Scholar] [CrossRef]
- Moura, M.A.; Bergmann, A.; Aguiar, S.S.; Thuler, L.C. The magnitude of the association between smoking and the risk of developing cancer in Brazil: A multicenter study. BMJ Open 2014, 4, e003736. [Google Scholar] [CrossRef]
- Wong, H.P.; Yu, L.; Lam, E.K.; Tai, E.K.; Wu, W.K.; Cho, C.H. Nicotine promotes cell proliferation via alpha7-nicotinic acetylcholine receptor and catecholamine-synthesizing enzymes-mediated pathway in human colon adenocarcinoma HT-29 cells. Toxicol. Appl. Pharmacol. 2007, 221, 261–267. [Google Scholar] [CrossRef]
- Chen, R.J.; Ho, Y.S.; Guo, H.R.; Wang, Y.J. Rapid activation of Stat3 and ERK1/2 by nicotine modulates cell proliferation in human bladder cancer cells. Toxicol. Sci. 2008, 104, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.H.; Lee, C.H.; Ho, Y.S. Nicotinic acetylcholine receptor-based blockade: Applications of molecular targets for cancer therapy. Clin. Cancer Res. 2011, 17, 3533–3541. [Google Scholar] [CrossRef] [Green Version]
- Reschke, M.; Mihic-Probst, D.; van der Horst, E.H.; Knyazev, P.; Wild, P.J.; Hutterer, M.; Meyer, S.; Dummer, R.; Moch, H.; Ullrich, A. HER3 is a determinant for poor prognosis in melanoma. Clin. Cancer Res. 2008, 14, 5188–5197. [Google Scholar] [CrossRef] [Green Version]
- Milanezi, F.; Carvalho, S.; Schmitt, F.C. EGFR/HER2 in breast cancer: A biological approach for molecular diagnosis and therapy. Expert Rev. Mol. Diagn. 2008, 8, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Koutras, A.K.; Fountzilas, G.; Kalogeras, K.T.; Starakis, I.; Iconomou, G.; Kalofonos, H.P. The upgraded role of HER3 and HER4 receptors in breast cancer. Crit. Rev. Oncol. Hematol. 2010, 74, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, A.; Schlessinger, J. Signal transduction by receptors with tyrosine kinase activity. Cell 1990, 61, 203–212. [Google Scholar] [CrossRef]
- Chu, E. Clinical colorectal cancer: The epidermal growth factor receptor signaling pathway as a chemotherapeutic target. Clin. Colorectal Cancer 2003, 2, 202–203. [Google Scholar] [CrossRef] [PubMed]
- Barton, S.; Starling, N.; Swanton, C. Predictive molecular markers of response to epidermal growth factor receptor (EGFR) family-targeted therapies. Curr. Cancer Drug Targets 2010, 10, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.; Peng, L.; Shou, J.; Wang, J.; Jin, Y.; Liang, F.; Zhao, J.; Wu, M.; Li, Q.; Zhang, B.; et al. EGFR-Mutated Squamous Cell Lung Cancer and Its Association with Outcomes. Front. Oncol. 2021, 11, 680804. [Google Scholar] [CrossRef]
- Reese, D.M.; Slamon, D.J. HER-2/neu signal transduction in human breast and ovarian cancer. Stem Cells 1997, 15, 1–8. [Google Scholar] [CrossRef]
- Davis-Searles, P.R.; Nakanishi, Y.; Kim, N.C.; Graf, T.N.; Oberlies, N.H.; Wani, M.C.; Wall, M.E.; Agarwal, R.; Kroll, D.J. Milk thistle and prostate cancer: Differential effects of pure flavonolignans from Silybum marianum on antiproliferative end points in human prostate carcinoma cells. Cancer Res. 2005, 65, 4448–4457. [Google Scholar] [CrossRef] [Green Version]
- Rollinger, J.M.; Schuster, D.; Baier, E.; Ellmerer, E.P.; Langer, T.; Stuppner, H. Taspine: Bioactivity-guided isolation and molecular ligand-target insight of a potent acetylcholinesterase inhibitor from Magnolia x soulangiana. J. Nat. Prod. 2006, 69, 1341–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaisberg, A.J.; Milla, M.; Planas, M.C.; Cordova, J.L.; de Agusti, E.R.; Ferreyra, R.; Mustiga, M.C.; Carlin, L.; Hammond, G.B. Taspine is the cicatrizant principle in Sangre de Grado extracted from Croton lechleri. Planta Med. 1989, 55, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Feng, W.; Ji, Y.; Jin, L. Resveratrol mediates cell cycle arrest and cell death in human esophageal squamous cell carcinoma by directly targeting the EGFR signaling pathway. Oncol. Lett. 2017, 13, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Ma, X.; Li, N.; Liu, H.; Dong, Q.; Zhang, J.; Yang, C.; Liu, Y.; Liang, Q.; Zhang, S.; et al. Resveratrol inhibits Hexokinases II mediated glycolysis in non-small cell lung cancer via targeting Akt signaling pathway. Exp. Cell Res. 2016, 349, 320–327. [Google Scholar] [CrossRef]
- Zhu, Y.; He, W.; Gao, X.; Li, B.; Mei, C.; Xu, R.; Chen, H. Resveratrol overcomes gefitinib resistance by increasing the intracellular gefitinib concentration and triggering apoptosis, autophagy and senescence in PC9/G NSCLC cells. Sci. Rep. 2015, 5, 17730. [Google Scholar] [CrossRef] [Green Version]
- Mikhailova, M.; Wang, Y.; Bedolla, R.; Lu, X.H.; Kreisberg, J.I.; Ghosh, P.M. AKT regulates androgen receptor-dependent growth and PSA expression in prostate cancer. Adv. Exp. Med. Biol. 2008, 617, 397–405. [Google Scholar]
- Mukund, V.; Mukund, D.; Sharma, V.; Mannarapu, M.; Alam, A. Genistein: Its role in metabolic diseases and cancer. Crit. Rev. Oncol. Hematol. 2017, 119, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Valley, C.C.; Arndt-Jovin, D.J.; Karedla, N.; Steinkamp, M.P.; Chizhik, A.I.; Hlavacek, W.S.; Wilson, B.S.; Lidke, K.A.; Lidke, D.S. Enhanced dimerization drives ligand-independent activity of mutant epidermal growth factor receptor in lung cancer. Mol. Biol. Cell 2015, 26, 4087–4099. [Google Scholar] [CrossRef]
- Wang, L.; Cao, H.; Lu, N.; Liu, L.; Wang, B.; Hu, T.; Israel, D.A.; Peek, R.M., Jr.; Polk, D.B.; Yan, F. Berberine inhibits proliferation and down-regulates epidermal growth factor receptor through activation of Cbl in colon tumor cells. PLoS ONE 2013, 8, e56666. [Google Scholar] [CrossRef] [Green Version]
- Goel, H.L.; Mercurio, A.M. VEGF targets the tumour cell. Nat. Rev. Cancer 2013, 13, 871–882. [Google Scholar] [CrossRef]
- Lian, L.; Li, X.L.; Xu, M.D.; Li, X.M.; Wu, M.Y.; Zhang, Y.; Tao, M.; Li, W.; Shen, X.M.; Zhou, C.; et al. VEGFR2 promotes tumorigenesis and metastasis in a pro-angiogenic-independent way in gastric cancer. BMC Cancer 2019, 19, 183. [Google Scholar] [CrossRef] [PubMed]
- Kamba, T.; McDonald, D.M. Mechanisms of adverse effects of anti-VEGF therapy for cancer. Br. J. Cancer 2007, 96, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Kazi, J.U.; Rönnstrand, L. FMS-like Tyrosine Kinase 3/FLT3: From Basic Science to Clinical Implications. Physiol. Rev. 2019, 99, 1433–1466. [Google Scholar] [CrossRef] [PubMed]
- Ambinder, A.J.; Levis, M. Potential targeting of FLT3 acute myeloid leukemia. Haematologica 2021, 106, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Shah, N.P.; Talpaz, M.; Deininger, M.W.; Mauro, M.J.; Flinn, I.W.; Bixby, D.; Lustgarten, S.; Gozgit, J.M.; Clackson, T.; Turner, C.D.; et al. Ponatinib in patients with refractory acute myeloid leukaemia: Findings from a phase 1 study. Br. J. Haematol. 2013, 162, 548–552. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.M.; Mandrekar, S.J.; Sanford, B.L.; Laumann, K.; Geyer, S.; Bloomfield, C.D.; Thiede, C.; Prior, T.W.; Döhner, K.; Marcucci, G.; et al. Midostaurin plus Chemotherapy for Acute Myeloid Leukemia with a FLT3 Mutation. N. Engl. J. Med. 2017, 377, 454–464. [Google Scholar] [CrossRef]
- Cortes, J.; Perl, A.E.; Döhner, H.; Kantarjian, H.; Martinelli, G.; Kovacsovics, T.; Rousselot, P.; Steffen, B.; Dombret, H.; Estey, E.; et al. Quizartinib, an FLT3 inhibitor, as monotherapy in patients with relapsed or refractory acute myeloid leukaemia: An open-label, multicentre, single-arm, phase 2 trial. Lancet Oncol. 2018, 19, 889–903. [Google Scholar] [CrossRef]
- Albers, C.; Leischner, H.; Verbeek, M.; Yu, C.; Illert, A.L.; Peschel, C.; von Bubnoff, N.; Duyster, J. The secondary FLT3-ITD F691L mutation induces resistance to AC220 in FLT3-ITD+ AML but retains in vitro sensitivity to PKC412 and Sunitinib. Leukemia 2013, 27, 1416–1418. [Google Scholar] [CrossRef]
- Xiao, T.; Xiao, Y.; Wang, W.; Tang, Y.Y.; Xiao, Z.; Su, M. Targeting EphA2 in cancer. J. Hematol. Oncol. 2020, 13, 114. [Google Scholar] [CrossRef]
- Moosavi, F.; Giovannetti, E.; Saso, L.; Firuzi, O. HGF/MET pathway aberrations as diagnostic, prognostic, and predictive biomarkers in human cancers. Crit. Rev. Clin. Lab. Sci. 2019, 56, 533–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigelow, R.L.; Cardelli, J.A. The green tea catechins, (-)-Epigallocatechin-3-gallate (EGCG) and (-)-Epicatechin-3-gallate (ECG), inhibit HGF/Met signaling in immortalized and tumorigenic breast epithelial cells. Oncogene 2006, 25, 1922–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.J.; Wu, L.F.; Chen, W.K.; Wang, C.J.; Tseng, T.H. Inhibitory effect of luteolin on hepatocyte growth factor/scatter factor-induced HepG2 cell invasion involving both MAPK/ERKs and PI3K-Akt pathways. Chem. Biol. Interact. 2006, 160, 123–133. [Google Scholar] [CrossRef]
- Ungefroren, H.; Gieseler, F.; Kaufmann, R.; Settmacher, U.; Lehnert, H.; Rauch, B.H. Signaling Crosstalk of TGF-β/ALK5 and PAR2/PAR1: A Complex Regulatory Network Controlling Fibrosis and Cancer. Int. J. Mol. Sci. 2018, 19, 1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, N.; Jin, K.; He, K.; Cao, J.; Teng, L. Protease-activated receptors in cancer: A systematic review. Oncol. Lett. 2011, 2, 599–608. [Google Scholar] [CrossRef]
- Shi, X.; Gangadharan, B.; Brass, L.F.; Ruf, W.; Mueller, B.M. Protease-activated receptors (PAR1 and PAR2) contribute to tumor cell motility and metastasis. Mol. Cancer Res. 2004, 2, 395–402. [Google Scholar]
- Kakarala, K.K.; Jamil, K. Protease activated receptor-2 (PAR2): Possible target of phytochemicals. J. Biomol. Struct. Dyn. 2015, 33, 2003–2022. [Google Scholar] [CrossRef]
- Dohadwala, M.; Batra, R.K.; Luo, J.; Lin, Y.; Krysan, K.; Pold, M.; Sharma, S.; Dubinett, S.M. Autocrine/paracrine prostaglandin E2 production by non-small cell lung cancer cells regulates matrix metalloproteinase-2 and CD44 in cyclooxygenase-2-dependent invasion. J. Biol. Chem. 2002, 277, 50828–50833. [Google Scholar] [CrossRef] [Green Version]
- Masato, M.; Miyata, Y.; Kurata, H.; Ito, H.; Mitsunari, K.; Asai, A.; Nakamura, Y.; Araki, K.; Mukae, Y.; Matsuda, T.; et al. Oral administration of E-type prostanoid (EP) 1 receptor antagonist suppresses carcinogenesis and development of prostate cancer via upregulation of apoptosis in an animal model. Sci. Rep. 2021, 11, 20279. [Google Scholar] [CrossRef]
- Sun, X.; Li, Q. Prostaglandin EP2 receptor: Novel therapeutic target for human cancers (Review). Int. J. Mol. Med. 2018, 42, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Dubois, R.N. Prostaglandins and cancer. Gut 2006, 55, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hull, M.A.; Ko, S.C.; Hawcroft, G. Prostaglandin EP receptors: Targets for treatment and prevention of colorectal cancer? Mol. Cancer Ther. 2004, 3, 1031–1039. [Google Scholar] [PubMed]
- Piazza, P.V.; Cota, D.; Marsicano, G. The CB1 Receptor as the Cornerstone of Exostasis. Neuron 2017, 93, 1252–1274. [Google Scholar] [CrossRef] [PubMed]
- Hillard, C.J. Stress regulates endocannabinoid-CB1 receptor signaling. Semin. Immunol. 2014, 26, 380–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, J.L. Medical marijuana for cancer. CA Cancer J. Clin. 2015, 65, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Velasco, G.; Carracedo, A.; Blázquez, C.; Lorente, M.; Aguado, T.; Haro, A.; Sánchez, C.; Galve-Roperh, I.; Guzmán, M. Cannabinoids and gliomas. Mol. Neurobiol. 2007, 36, 60–67. [Google Scholar] [CrossRef]
- Guzmán, M.; Duarte, M.J.; Blázquez, C.; Ravina, J.; Rosa, M.C.; Galve-Roperh, I.; Sánchez, C.; Velasco, G.; González-Feria, L. A pilot clinical study of Delta9-tetrahydrocannabinol in patients with recurrent glioblastoma multiforme. Br. J. Cancer 2006, 95, 197–203. [Google Scholar] [CrossRef]
- Breijyeh, Z.; Jubeh, B.; Bufo, S.A.; Karaman, R.; Scrano, L. Cannabis: A Toxin-Producing Plant with Potential Therapeutic Uses. Toxins 2021, 13, 117. [Google Scholar] [CrossRef]
- Blázquez, C.; Carracedo, A.; Barrado, L.; Real, P.J.; Fernández-Luna, J.L.; Velasco, G.; Malumbres, M.; Guzmán, M. Cannabinoid receptors as novel targets for the treatment of melanoma. FASEB J. 2006, 20, 2633–2635. [Google Scholar] [CrossRef] [Green Version]
- Ligresti, A.; Moriello, A.S.; Starowicz, K.; Matias, I.; Pisanti, S.; De Petrocellis, L.; Laezza, C.; Portella, G.; Bifulco, M.; Di Marzo, V. Antitumor activity of plant cannabinoids with emphasis on the effect of cannabidiol on human breast carcinoma. J. Pharmacol. Exp. Ther. 2006, 318, 1375–1387. [Google Scholar] [CrossRef] [Green Version]
- McAllister, S.D.; Soroceanu, L.; Desprez, P.Y. The Antitumor Activity of Plant-Derived Non-Psychoactive Cannabinoids. J. Neuroimmune Pharmacol. 2015, 10, 255–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, G.; Sánchez, C.; Guzmán, M. Towards the use of cannabinoids as antitumour agents. Nat. Rev. Cancer 2012, 12, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Emerging strategies for exploiting cannabinoid receptor agonists as medicines. Br. J. Pharmacol. 2009, 156, 397–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, S.; Lorente, M.; Rodríguez-Fornés, F.; Hernández-Tiedra, S.; Salazar, M.; García-Taboada, E.; Barcia, J.; Guzmán, M.; Velasco, G. A combined preclinical therapy of cannabinoids and temozolomide against glioma. Mol. Cancer Ther. 2011, 10, 90–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, T.D.; Zhang, W.; Suto, M.J.; Li, Y. Frizzled7 as an emerging target for cancer therapy. Cell Signal. 2012, 24, 846–851. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Xu, M.; Lin, M.; Zhang, Y.; Zheng, X.; Sun, Y.; Deng, Y.; Pan, J.; Xu, Z.; Lu, X.; et al. Overexpression of FZD7 is associated with poor survival in patients with colon cancer. Pathol. Res. Pract. 2019, 215, 152478. [Google Scholar] [CrossRef]
- Yin, S.; Xu, L.; Bonfil, R.D.; Banerjee, S.; Sarkar, F.H.; Sethi, S.; Reddy, K.B. Tumor-initiating cells and FZD8 play a major role in drug resistance in triple-negative breast cancer. Mol. Cancer Ther. 2013, 12, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Xu, M.R.; Wei, P.F.; Suo, M.Z.; Hu, Y.; Ding, W.; Su, L.; Zhu, Y.D.; Song, W.J.; Tang, G.H.; Zhang, M.; et al. Brucine Suppresses Vasculogenic Mimicry in Human Triple-Negative Breast Cancer Cell Line MDA-MB-231. Biomed. Res. Int. 2019, 2019, 6543230. [Google Scholar] [CrossRef]
- Fuentes, N.; Silveyra, P. Estrogen receptor signaling mechanisms. Adv. Protein Chem. Struct. Biol. 2019, 116, 135–170. [Google Scholar]
- Solomon, Z.J.; Mirabal, J.R.; Mazur, D.J.; Kohn, T.P.; Lipshultz, L.I.; Pastuszak, A.W. Selective Androgen Receptor Modulators: Current Knowledge and Clinical Applications. Sex. Med. Rev. 2019, 7, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Goktas, S.; Ziada, A.; Crawford, E.D. Combined androgen blockade for advanced prostatic carcinoma. Prostate Cancer Prostatic Dis. 1999, 2, 172–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linja, M.J.; Savinainen, K.J.; Saramäki, O.R.; Tammela, T.L.; Vessella, R.L.; Visakorpi, T. Amplification and overexpression of androgen receptor gene in hormone-refractory prostate cancer. Cancer Res. 2001, 61, 3550–3555. [Google Scholar] [PubMed]
- Birt, D.F.; Hendrich, S.; Wang, W. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther. 2001, 90, 157–177. [Google Scholar] [CrossRef]
- Kang, M.H.; Choi, H.; Oshima, M.; Cheong, J.H.; Kim, S.; Lee, J.H.; Park, Y.S.; Choi, H.S.; Kweon, M.N.; Pack, C.G.; et al. Estrogen-related receptor gamma functions as a tumor suppressor in gastric cancer. Nat. Commun. 2018, 9, 1920. [Google Scholar] [CrossRef]
- Madhavan, S.; Gusev, Y.; Singh, S.; Riggins, R.B. ERRγ target genes are poor prognostic factors in Tamoxifen-treated breast cancer. J. Exp. Clin. Cancer Res. 2015, 34, 45. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Li, Y.; Li, K.W.; Zhou, C.Z. Sophoridine: A review of its pharmacology, pharmacokinetics and toxicity. Phytomedicine 2022, 95, 153756. [Google Scholar] [CrossRef]
- Wu, G.S. TRAIL as a target in anti-cancer therapy. Cancer Lett. 2009, 285, 1–5. [Google Scholar] [CrossRef]
- Yuan, X.; Gajan, A.; Chu, Q.; Xiong, H.; Wu, K.; Wu, G.S. Developing TRAIL/TRAIL death receptor-based cancer therapies. Cancer Metastasis Rev. 2018, 37, 733–748. [Google Scholar] [CrossRef]
- Mehta, K.; Pantazis, P.; McQueen, T.; Aggarwal, B.B. Antiproliferative effect of curcumin (diferuloylmethane) against human breast tumor cell lines. Anticancer Drugs 1997, 8, 470–481. [Google Scholar] [CrossRef]
- Christofides, A.; Konstantinidou, E.; Jani, C.; Boussiotis, V.A. The role of peroxisome proliferator-activated receptors (PPAR) in immune responses. Metabolism 2021, 114, 154338. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.; Wagner, K.D. The Role of PPARs in Disease. Cells 2020, 9, 2367. [Google Scholar] [CrossRef] [PubMed]
- Burits, M.; Bucar, F. Antioxidant activity of Nigella sativa essential oil. Phytother. Res. 2000, 14, 323–328. [Google Scholar] [CrossRef]
- Jehan, S.; Zhong, C.; Li, G.; Zulqarnain Bakhtiar, S.; Li, D.; Sui, G. Thymoquinone Selectively Induces Hepatocellular Carcinoma Cell Apoptosis in Synergism with Clinical Therapeutics and Dependence of p53 Status. Front. Pharmacol. 2020, 11, 555283. [Google Scholar] [CrossRef] [PubMed]
- Sang, S.; Hong, J.; Wu, H.; Liu, J.; Yang, C.S.; Pan, M.H.; Badmaev, V.; Ho, C.T. Increased growth inhibitory effects on human cancer cells and anti-inflammatory potency of shogaols from Zingiber officinale relative to gingerols. J. Agric. Food Chem. 2009, 57, 10645–10650. [Google Scholar] [CrossRef] [Green Version]
- Isa, Y.; Miyakawa, Y.; Yanagisawa, M.; Goto, T.; Kang, M.S.; Kawada, T.; Morimitsu, Y.; Kubota, K.; Tsuda, T. 6-Shogaol and 6-gingerol, the pungent of ginger, inhibit TNF-alpha mediated downregulation of adiponectin expression via different mechanisms in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2008, 373, 429–434. [Google Scholar] [CrossRef]
- Hennessy, E.J.; Parker, A.E.; O’Neill, L.A. Targeting Toll-like receptors: Emerging therapeutics? Nat. Rev. Drug Discov. 2010, 9, 293–307. [Google Scholar] [CrossRef]
- Chuang, H.C.; Lin, H.Y.; Liao, P.L.; Huang, C.C.; Lin, L.L.; Hsu, W.M.; Chuang, J.H. Immunomodulator polyinosinic-polycytidylic acid enhances the inhibitory effect of 13-cis-retinoic acid on neuroblastoma through a TLR3-related immunogenic-apoptotic response. Lab. Invest. 2020, 100, 606–618. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, H.; Zhu, Z.; Gao, Y.; Xiang, B.; Wei, Q. The immunostimulatory effects and pro-apoptotic activity of rhCNB against Lewis lung cancer is mediated by Toll-like receptor 4. Cancer Med. 2019, 8, 4441–4453. [Google Scholar] [CrossRef]
- Boozari, M.; Butler, A.E.; Sahebkar, A. Impact of curcumin on toll-like receptors. J. Cell Physiol. 2019, 234, 12471–12482. [Google Scholar] [CrossRef]
- Li, P.M.; Li, Y.L.; Liu, B.; Wang, W.J.; Wang, Y.Z.; Li, Z. Curcumin inhibits MHCC97H liver cancer cells by activating ROS/TLR-4/caspase signaling pathway. Asian Pac. J. Cancer Prev. 2014, 15, 2329–2334. [Google Scholar] [CrossRef]
- Lozon, Y.; Sultan, A.; Lansdell, S.J.; Prytkova, T.; Sadek, B.; Yang, K.H.; Howarth, F.C.; Millar, N.S.; Oz, M. Inhibition of human α7 nicotinic acetylcholine receptors by cyclic monoterpene carveol. Eur. J. Pharmacol. 2016, 776, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, H.; Shaw, I.C. Food flavonoid ligand structure/estrogen receptor-α affinity relationships-toxicity or food functionality? Food Chem. Toxicol. 2019, 129, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Guamán Ortiz, L.M.; Tillhon, M.; Parks, M.; Dutto, I.; Prosperi, E.; Savio, M.; Arcamone, A.G.; Buzzetti, F.; Lombardi, P.; Scovassi, A.I. Multiple effects of berberine derivatives on colon cancer cells. Biomed. Res. Int. 2014, 2014, 924585. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tong, Y.; Yang, X.; Li, F.; Zheng, L.; Liu, W.; Wu, J.; Ou, R.; Zhang, G.; Hu, M.; et al. Novel histone deacetylase inhibitors derived from Magnolia officinalis significantly enhance TRAIL-induced apoptosis in non-small cell lung cancer. Pharmacol. Res. 2016, 111, 113–125. [Google Scholar] [CrossRef] [PubMed]
- Tanjak, P.; Thiantanawat, A.; Watcharasit, P.; Satayavivad, J. Genistein reduces the activation of AKT and EGFR, and the production of IL6 in cholangiocarcinoma cells involving estrogen and estrogen receptors. Int. J. Oncol. 2018, 53, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filardo, E.J.; Quinn, J.A.; Bland, K.I.; Frackelton, A.R., Jr. Estrogen-induced activation of Erk-1 and Erk-2 requires the G protein-coupled receptor homolog, GPR30, and occurs via trans-activation of the epidermal growth factor receptor through release of HB-EGF. Mol. Endocrinol. 2000, 14, 1649–1660. [Google Scholar] [CrossRef]
- Daub, H.; Wallasch, C.; Lankenau, A.; Herrlich, A.; Ullrich, A. Signal characteristics of G protein-transactivated EGF receptor. EMBO J. 1997, 16, 7032–7044. [Google Scholar] [CrossRef]


{kind=link}
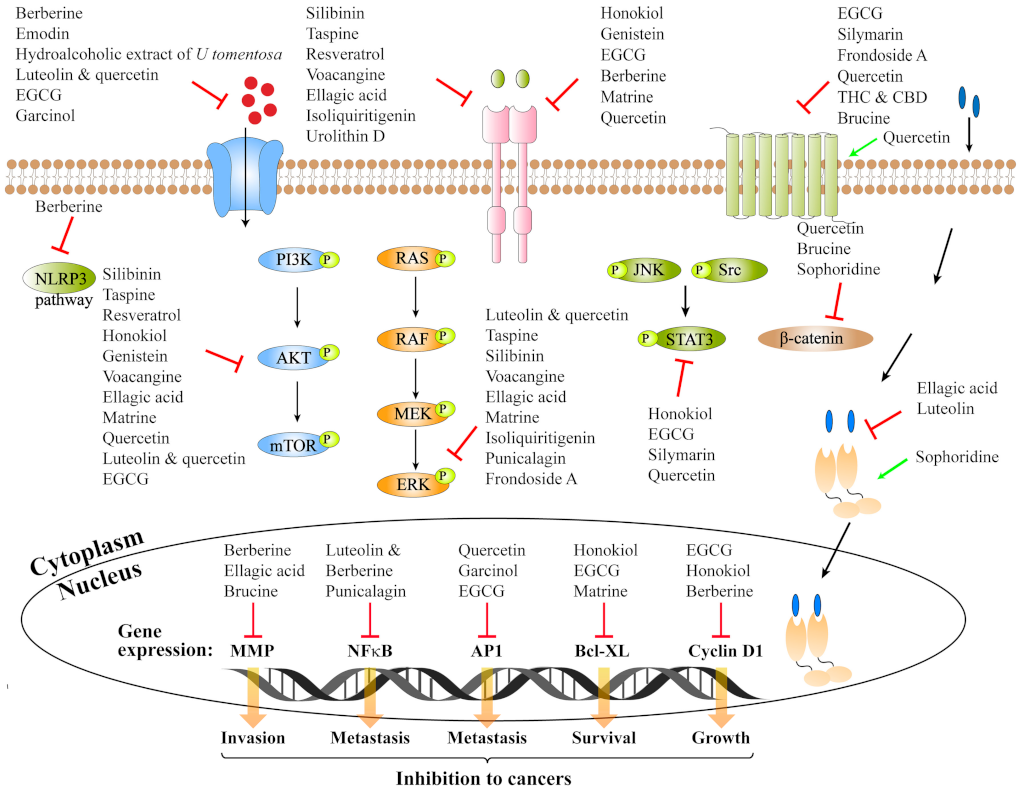
| Receptors | Targets | Compounds | Compound Categories | Main Sources | Biological Models | Pharmacological Activities | Solvents | Dosages | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Ion-channel-coupled | P2X7R | Berberine | Isoquinoline -alkaloid | Huanglian and Huangbai | MDA-MB-231 | Reduce P2X7R and NLRP3 expression; anti-metastasis | Water-soluble | 2.5–100 μg/mL | [32] |
| Emodin | Anthraquinone | Rheum- palmatum | MDA-MB-435 | Inhibit P2X7R-dependent Ca2+ signaling and ECM degradation; anti-metastasis | DMSO | 10 μM | [33] | ||
| Hydroalcoholic extract U tomentosa | Oxindole alkaloids | Uncaria tomentosa | MDA-MB-231 | Reduce P2X7R expression; anti-metastasis | 60% ethanol | 250 and 500 μg/mL | [34] | ||
| α9-nAChR | Luteolin and Quercetin | Flavonoid | Sophora and Japonicapeanut | MDA-MB-231 | Reduce α9-nAChR expression; anti-proliferation | DMSO | 0.5–1 μM | [35] | |
| Epigallocatechin-3-gallate | Flavonoid | Camelia sinensis | MCF-7 | Reduce α9-nAChR expression; anti-proliferation | DMSO | 1–10 μM | [36] | ||
| Garcinol (Gar) | Polyisoprenylated benzophenone | Garcinia indica fruit rind | MDA-MB-231 and MCF-7 | Reduce α9-nAChR and cyclin D3 expression; anti-proliferation | DMSO | 1–10 μM | [37] | ||
| Enzyme-linked receptor | EGFR | Silibinin | Polyphenolic | Silybium marianum | Head and neck squamous cell carcinoma | Bind to EGFR, inhibit EGFR phosphorylation; inhibit EGFR/LOX pathway; anti-migration; anti-metastasis | - | 5–20 μM | [38] |
| Taspine | Alkaloid | Magnolia spec. | A431 epidermoid cancer and HEK293/EGFR; A431 xenografted | Bind to EGFR, inhibit EGFR, AKT and ERK1/2 phosphorylation; in vivo significantly inhibit tumor growth; anti-proliferation | - | 0–6.4 μM | [39] | ||
| Resveratrol | Phytoalexin | Diverse plants | Prostate cancer DU145 (AR-) | Bind to EGFR, inhibit EGFR, AKT and ERK phosphorylation; anti-proliferation | - | 10–50 μM | [40] | ||
| Honokiol | Phenols | Magnolia officinalis | NSCLC; HNSCC xenograft | Inhibit EGFR, p-AKT and p-STAT3 expression; in vitro combination of erlotinib to inhibit invasion and metastasis; in vitro combination of honokiol and cetuximab to significantly enhance growth inhibition; (intraperitoneal injection) | 100% ethanol | 0.01–100 μM | [41] | ||
| Genistein | Isoflavonoid | Glycine max | HSC3 and KB oral squamous cell carcinoma; OSCC xenograft | Downregulate p-EGFR and p-AKT in HSC3, but not in KB cells; in vivo combination with cetuximab to significantly enhance growth inhibition (IP injection); anti-proliferation | DMSO | 0–80 μM | [42] | ||
| Silibinin | Polyphenolic | Silybium marianum | NSCLC and 293T; PC-9 xenografts | Inhibit EGFR activity by interfering with dimerization of EGFR; inhibit EGFR expression and EGFRAKT phosphorylation; in vitro combination with silibinin and erlotinib to induce apoptosis and growth inhibition (oral gavage); anti-proliferation | - | 0–200 μM | [43] | ||
| Epigallocatechin-3-gallate | Flavonoid | Camelia sinensis | YCU-N861 and YCU-H891 Head and neck squamous cell carcinoma | Inhibit p-EGFR, p-STAT3 and p-ERK expression; G1 cell cycle arrest, induce apoptosis; combination with 5-fluorouracil to significantly enhance growth inhibition | Water | 10–40 μg/mL | [44] | ||
| EGFR/HER2 | Berberine | Isoquinoline -alkaloid | Huanglian and Huangbai | Human ovarian cancer cells; HT-29 cell xenograft | Downregulate EGFR and HER2 expression; inhibit EGFR-HER2/PI3K/Akt signaling pathway; inhibit migration and invasion | DMSO | 0–100 μM | [45] | |
| VEGFR2 | Voacangine | Alkaloid | Voacanga africana | Glioblastoma HUVEC; U87MG cells xenograft | Bind to VEGFR2; inhibit p-VEGFR2, p-ERK and P-AKT expression; in vitro combination with sunitinib to inhibit growth; (IP injection); anti-proliferation; | DMSO | 50 μM | [46] | |
| Ellagic acid | Polyphenol | Pomegranate | HUVECs and MDA-MB-231; Breast cancer xenograft | Bind to VEGFR2; inhibit p-VEGFR2, p-ERK, P-AKT and p-JNK expression; inhibit proliferation and invasion; in vivo inhibit growth (IP injection) | DMSO | 0–20 μM | [47] | ||
| Bladder cancer T24, UM-UC-3, 5637, HT-1376 and UM-UC-3 xenograft | Downregulate VEGFR2 expression; combination with mitomycin C to inhibit proliferation, migration and invasion; in vivo inhibit tumor growth; (IP injection) | DMSO | 5–60 μM | [48] | |||||
| Matrine | Alkaloid | Sophora flavescens | UBC cell lines | Inhibit VEGFR2 BCL-2, caspase-3, p-AKT and p-PI3K expression; increase Bax and cleaved caspase-3; combination with cisplatin to inhibit proliferation, migration and invasion | Physiological saline | 0–16 μM | [49] | ||
| FLT3 | Isoliquiritigenin | Flavonoid | Licorice | AML; AML xenograft model | Bind to FLT3; inhibit P-FLT3, p-STAT3, p-Erk1/2 and csapase3 expression; in vivo inhibit proliferation and induce apoptosis; (administered orally) | DMSO | 0–40 μM | [50] | |
| EphA2R | Ellagitannin -metabolite urolithin D | Polyphenol | Pomegranate | Prostate cancer cells | Bind to EphA2R; inhibit EphA2 phosphorylation; anti-proliferation | DMSO | 0–30 μM | [51] | |
| c-Met | Quercetin | Flavonoid | Sophora japonica | Melanoma | Reduce c-Met homo-dimerization; inhibit c-Met, Gab1, FAK and PAK phosphorylation; inhibit migration and invasion | DMSO | 0–80 μM | [52] | |
| G Protein-coupled receptors | PAR2 | Epigallocatechin-3-gallate | Flavonoid | Camelia sinensis | Human colon cancer SW620 | Inhibit that PAR2 activation by PAR2-AP or the TF/factor VIIa complex; inhibit ERK-1/2 and NF-κB activation; inhibit migration and proliferation; | - | 0–100 µg/mL | [53] |
| EP2 | Silymarin | Polyphenolic | Silybium marianum | Human renal carcinoma Caki | Inhibit migration; inhibit EP2, p-CREB, p-STAT3, P-Src expression | - | 20–50 µg/mL | [54] | |
| EP1 | Epigallocatechin-3-gallate | Flavonoid found | Camelia sinensis | HCC cell line | Inhibit migration and proliferation; inhibit EP1 expression; | - | 12.5–100 μg/mL | [55] | |
| EP2/EP4 | Frondoside A | Triterpene glycoside | Cucumaria frondosa | Balb/cByJ female mice | Selectively antagonize EP2/4; inhibit p-Erk expression; (IP injection); inhibit metastasis | PBS | 0.1–5 μM/L | [56] | |
| CB1/ CB2 | Tetrahydrocannabinol and cannabidiol | Phenols | Cannabis | Glioblastoma cell lines | Bind to CB1 and/or CB2 receptors; induce CB1 and/or CB2 receptor activation; inhibit invasiveness; induce apoptosis | - | 1.7 μM and 0.4 μM | [57] | |
| CB1 | Quercetin | Flavonoid | Sophora japonica | Caco2 and DLD-1 | Induce CB1-R expression; inhibit PI3K/AKT/mTOR and induce JNK/JUN pathways; inhibit the migration; induce apoptosis | DMSO or Absolute ethanol | 10–50 μM | [58] | |
| Frizzled-7 | BEL/5-FU and BEL-7402 | Suppress FZD7 and β-catenin expression; decrease ABCB1, ABCC1 and ABCC2 expression; reverse MDR | - | 0–160 μM | [59] | ||||
| Frizzled-8 | Brucine | Alkaloid | Strychnos nux-vomica L. | LoVo, SW480 and Caco-2; xenograft model | Downregulate Frizzled-8, MMP2, MMP3 and MMP9; inhibit Wnt/β-catenin signaling pathway; inhibit migration, and induce apoptosis; (orally administered) | - | 0–50 μM | [60] | |
| Nuclear receptor superfamily | ERβ | Ellagic acid | Polyphenol | Pomegranate | HeLa and MCF-7 | Estrogen antagonist | - | 10−6–10−8 M | [61] |
| AR | Luteolin | Flavonoid found | Peanut | Prostate cancer cell; LNCaP xenograft | Anti-proliferative; inhibit AR and PSA expression; in vivo inhibit tumor growth and downregulate AR and PSA (IP injection) | DMSO | 0–40 μM | [62] | |
| ESRRG | Sophoridine | Monomeric alkaloid | Sophora alopecuroides L. | Gastric cancer AGS and SGC7901 | Combination with cisplatin to inhibit proliferation, induce apoptosis; inhibit the migration and invasion; Enhance ESRRG expression and β-catenin degradation | DMSO | 3 μM | [63] | |
| Other receptors | DR5 | Curcumin | Diketones | Curcuma species | Human renal Caki cells | Enhance ROS and DR5 expression; induce apoptosis; | - | 0–30 μM | [64] |
| Ginsenoside C-K | Saponins | Ginseng | Colon cancer cells | induce apoptosis, induce Bax, Bid, cytochrome c expression; inhibit MCL-1, BCL-2, cFLIP and XIAP expression; enhance ROS, DR5 and p-JNK expression | DMSO | 0–100 μM | [65] | ||
| PPARγ | Ginsenoside F2 | Saponins | Ginseng | Mouse embryo fibroblast 3T3-L1 | Bind to PPARγ; inhibit PPARγ and perilipin expression | - | 10–100 μM | [66] | |
| Thymoquinone | - | Nigella sativa | Breast cancer cells | Bind to PPARγ; increase PPARγ activity; induce apoptosis | - | 0–80 μM | [67] | ||
| 6-Shogaol | - | Rhizomes of ginger | Breast and colon cancer cells | Binding to PPARγ; increase in PPARγ activity; induce apoptosis | DMSO | 0–100 μM | [68] | ||
| TLR4 | Curcumin | Diketones | Curcuma species | RAW264.7 | Inhibit dimerization of TLR4 | - | 10–50 μm | [69] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Li, D.; Zhao, B.; Kim, J.; Sui, G.; Shi, J. Small Molecule Compounds of Natural Origin Target Cellular Receptors to Inhibit Cancer Development and Progression. Int. J. Mol. Sci. 2022, 23, 2672. https://doi.org/10.3390/ijms23052672
Wang J, Li D, Zhao B, Kim J, Sui G, Shi J. Small Molecule Compounds of Natural Origin Target Cellular Receptors to Inhibit Cancer Development and Progression. International Journal of Molecular Sciences. 2022; 23(5):2672. https://doi.org/10.3390/ijms23052672
Chicago/Turabian StyleWang, Jinhua, Dangdang Li, Bo Zhao, Juhyok Kim, Guangchao Sui, and Jinming Shi. 2022. "Small Molecule Compounds of Natural Origin Target Cellular Receptors to Inhibit Cancer Development and Progression" International Journal of Molecular Sciences 23, no. 5: 2672. https://doi.org/10.3390/ijms23052672