
Fluorine-Containing Drug Administration in Rats Results in Fluorination of Selected Proteins in Liver and Brain Tissue

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Study Limitations
5. Materials and Methods
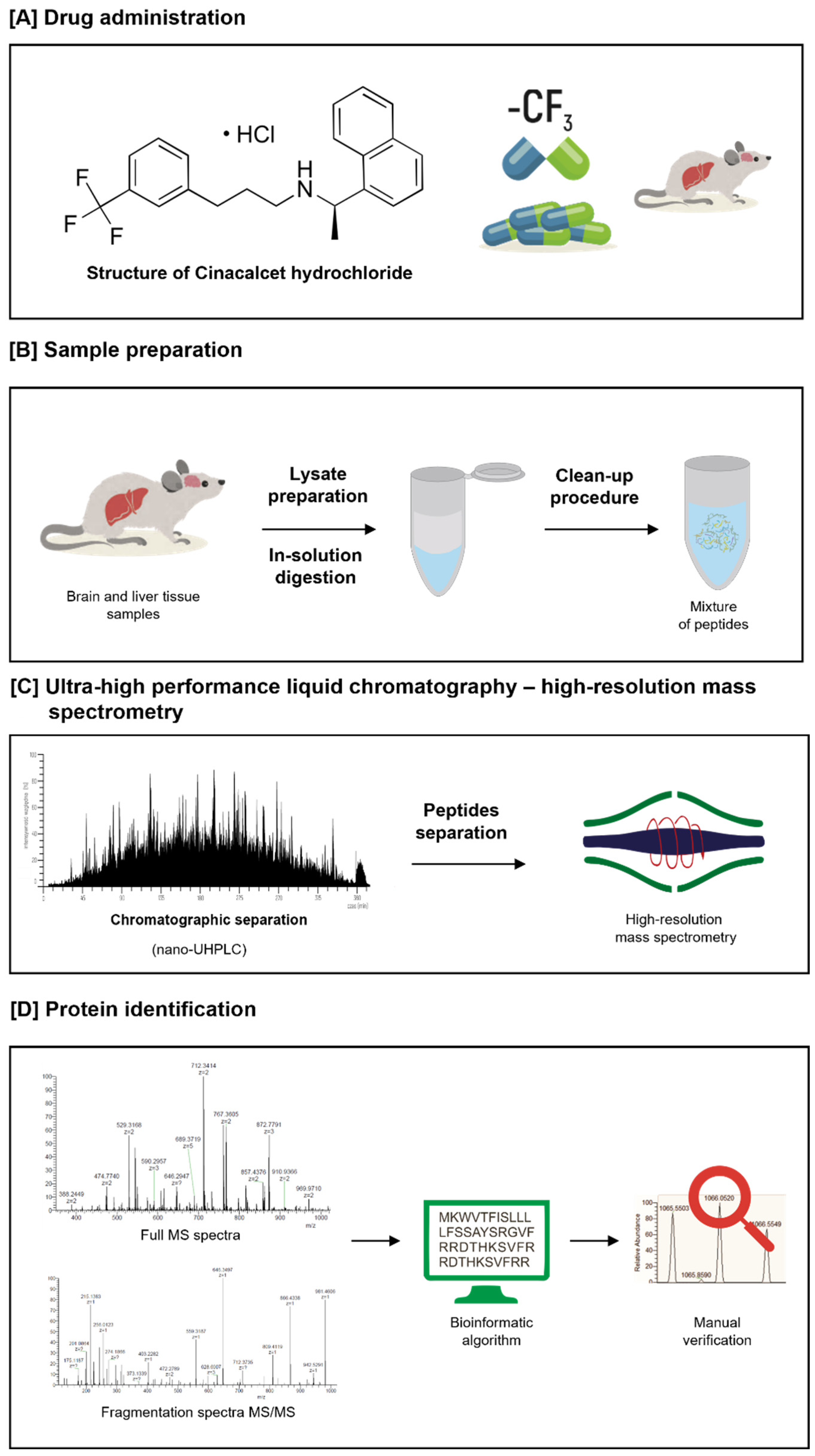
5.1. Chemicals, Reagents, and Instrumentation
5.2. Sample Preparation and Liquid Chromatography–MS/MS Analysis
5.3. Data Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, P.; Westwell, A.D. The role of fluorine in medicinal chemistry. J. Enzym. Inhib. Med. Chem. 2007, 22, 527–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calero, P.; Volke, D.C.; Lowe, P.T.; Gotfredsen, C.H.; O’Hagan, D.; Nike, P.I. A fluoride-responsive genetic circuit enables in vivo biofluorination in engineered Pseudomonas putida. Nat. Commun. 2020, 11, 5045. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, C.; Chaudhary, M.; De Oliveira, R.N.; Borbas, A.; Kempaiah, P.; Singh, P.; Rathi, B. Fluorinated scaffolds for antimalarial drug discovery. Expert Opin. Drug Discov. 2020, 15, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Huhmann, S.; Nyakatura, E.K.; Rohrhofer, A.; Moschner, J.; Schmidt, B.; Eichler, J.; Roth, C.; Koksch, B. Systematic Evaluation of Fluorination as Modification for Peptide-Based Fusion Inhibitors against HIV-1 Infection. ChemBioChem 2021, 22, 3443–3451. [Google Scholar] [CrossRef]
- Park, B.K.; Kitteringham, N.R.; O’Neill, P.M. Metabolism of fluorine-containing drugs. Annu. Rev. Pharm. Toxicol. 2001, 41, 443–470. [Google Scholar] [CrossRef]
- Inoue, M.; Sumii, Y.; Shibata, N. Contribution of Organofluorine Compounds to Pharmaceuticals. ACS Omega 2020, 5, 10633–10640. [Google Scholar] [CrossRef]
- Johnson, B.M.; Shu, Y.Z.; Zhuo, X.; Meanwell, N.A. Metabolic and Pharmaceutical Aspects of Fluorinated Compounds. J. Med. Chem. 2020, 63, 6315–6386. [Google Scholar] [CrossRef]
- Summary of Product Characteristics. Available online: https://www.ema.europa.eu/en/documents/product-information/mimpara-epar-product-information_en.pdf (accessed on 6 December 2021).
- Bover, J.; Ureña, P.; Ruiz-García, C.; DaSilva, I.; Lescano, P.; del Carpio, J.; Ballarín, J.; Cozzolino, M. Clinical and Practical Use of Calcimimetics in Dialysis Patients with Secondary Hyperparathyroidism. Clin. J. Am. Soc. Nephrol. 2016, 11, 161–174. [Google Scholar] [CrossRef] [Green Version]
- Thompson, D.C.; Perera, K.; London, R. Spontaneous Hydrolysis of 4-Trifluoromethylphenol to a Quinone Methide and Subsequent Protein Alkylation. Chem. Biol. Interact. 2000, 126, 1–14. [Google Scholar] [CrossRef]
- Khojasteh, S.C.; Argikar, U.A.; Driscoll, J.P.; Heck, C.J.S.; King, L.; Jackson, K.D.; Jian, W.; Kalgutkar, A.S.; Miller, G.P.; Kramlinger, V.; et al. Novel advances in biotransformation and bioactivation research—2020 year in review. Drug Metab. Rev. 2021, 53, 384–433. [Google Scholar] [CrossRef]
- Ekstrand., J.; Alván, G.; Boréus, L.O.; Norlin, A. Pharmacokinetics of fluoride in man after single and multiple oral doses. Eur. J. Clin. Pharm. 1977, 12, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, J.; Liang, C.; Yang, W.; Zhu, Q.; Luo, H.; Liu, X.; Wang, J.; Zhang, J. Potential Protective Effect of Riboflavin Against Pathological Changes in the Main Organs of Male Mice Induced by Fluoride Exposure. Biol. Trace Elem. Res. 2021, 200, 1262–1273. [Google Scholar] [CrossRef] [PubMed]
- Ning, H.; Li, C.; Yin, Z.; Hu, D.; Ge, Y.; Chen, L. Fluoride exposure decreased neurite formation on cerebral cortical neurons of SD rats in vitro. Environ. Sci. Pollut. Res. Int. 2021, 28, 50975–50982. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, L.; Govindarajan, V.; Ameeramja, J.; Nair, H.R.; Perumal, E. Single oral acute fluoride exposure causes changes in cardiac expression of oxidant and antioxidant enzymes, apoptotic and necrotic markers in male rats. Biochimie 2015, 119, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Gawor, A.; Konopka, A.; Ruszczyńska, A.; Pączek, L.; Gajewski, Z.; Bulska, E. Molecular Absorption and Mass Spectrometry for Complementary Analytical Study of Fluorinated Drugs in Animal Organisms. J. Anal. Spectrom. 2020, 35, 1840–1847. [Google Scholar] [CrossRef]
- Eriksson, J.; Chait, B.T.; Fenyö, D. A statistical basis for testing the significance of mass spectrometric protein identification results. Anal. Chem. 2000, 72, 999–1005. [Google Scholar] [CrossRef]
- Resing, K.A.; Meyer-Arendt, K.; Mendoza, A.M.; Aveline-Wolf, L.D.; Jonscher, K.R.; Pierce, K.G.; Old, W.M.; Cheung, H.T.; Russell, S.; Wattawa, J.L.; et al. Improving reproducibility and sensitivity in identifying human proteins by shotgun proteomics. Anal. Chem. 2004, 76, 3556–3568. [Google Scholar] [CrossRef]
- Padhi, D.; Harris, R. Clinical pharmacokinetic and pharmacodynamic profile of cinacalcet hydrochloride. Clin. Pharmacokinet. 2009, 48, 303–311. [Google Scholar] [CrossRef]
- Buer, B.C.; Meagher, J.L.; Stuckey, J.A.; Marsh, E.N. Structural basis for the enhanced stability of highly fluorinated proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 4810–4815. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.H.; Sproul, C.; Poppe, L.; Turner, S.; Gohdes, M.; Ghoborah, H.; Padhi, D.; Roskos, L. Metabolism and disposition of calcimimetic agent cinacalcet HCl in humans and animal models. Drug Metab. Dispos. 2004, 32, 1491–1500. [Google Scholar] [CrossRef] [Green Version]
- Buer, B.C.; Marsh, E.N. Fluorine: A new element in protein design. Protein Sci. 2012, 21, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robalo, J.R.; Vila Verde, A. Unexpected trends in the hydrophobicity of fluorinated amino acids reflect competing changes in polarity and conformation. Phys. Chem. Chem. Phys. 2019, 21, 2029–2038. [Google Scholar] [CrossRef] [Green Version]
- Marsh, E.N. Fluorinated proteins: From design and synthesis to structure and stability. Acc. Chem. Res. 2014, 47, 2878–2886. [Google Scholar] [CrossRef] [PubMed]
- Yoo, T.H.; Link, A.J.; Tirrell, D.A. Evolution of a fluorinated green fluorescent protein. Proc. Natl. Acad. Sci. USA 2007, 104, 13887–13890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, B.; Liu, H.; Liu, Z.; Doenen, R.; Nash, M.A. Influence of Fluorination on Single-Molecule Unfolding and Rupture Pathways of a Mechanostable Protein Adhesion Complex. Nano Lett. 2020, 20, 8940–8950. [Google Scholar] [CrossRef]
- Naarmann, N.; Bilgiçer, B.; Meng, H.; Kumar, K.; Steinem, C. Fluorinated interfaces drive self-association of transmembrane alpha helices in lipid bilayers. Angew. Chem. Int. Ed. Engl. 2006, 45, 2588–2591. [Google Scholar] [CrossRef]
- Welte, H.; Zhou, T.; Mihajlenko, X.; Mayans, O.; Kovermann, M. What does fluorine do to a protein? Thermodynamic, and highly-resolved structural insights into fluorine-labelled variants of the cold shock protein. Sci. Rep. 2020, 10, 2640. [Google Scholar] [CrossRef]
- Sun, X.; Dyson, H.J.; Wright, P.E. Fluorotryptophan Incorporation Modulates the Structure and Stability of Transthyretin in a Site-Specific Manner. Biochemistry 2017, 56, 5570–5581. [Google Scholar] [CrossRef]
- Meng, H.; Krishnaji, S.T.; Beinborn, M.; Kumar, K. Influence of selective fluorination on the biological activity and proteolytic stability of glucagon-like peptide-1. J. Med. Chem. 2008, 51, 7303–7307. [Google Scholar] [CrossRef] [Green Version]
- Tang, H.C.; Lin, Y.J.; Horng, J.C. Modulating the folding stability and ligand binding affinity of Pin1 WW domain by proline ring puckering. Proteins 2014, 82, 67–76. [Google Scholar] [CrossRef]
- Dominguez, M.A., Jr.; Thornton, K.C.; Melendez, M.G.; Dupureur, C.M. Differential effects of isomeric incorporation of fluorophenylalanines into PvuII endonuclease. Proteins 2001, 45, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Uniprot. Available online: https://www.uniprot.org (accessed on 30 October 2021).
- Salwiczek, M.; Nyakatura, E.K.; Gerling, U.I.; Ye, S.; Koksch, B. Fluorinated amino acids: Compatibility with native protein structures and effects on protein-protein interactions. Chem. Soc. Rev. 2012, 41, 2135–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Examined Tissue | Peptide Sequence | Variable Modification | Experimental Mass | Theoretical Mass | Δ Mass [ppm] | Peptide Score | Protein Name | Gene Name |
|---|---|---|---|---|---|---|---|---|
| Liver | GASIPQFTNSPTMVIMVGLPAR | fluorination (A) | 2304.1958 | 2304.1770 | 8.15 | 120 | 6-phosphofructo-2-kinase/ fructose-2,6-bisphosphatase 1 (Fragment) | Pfkfb1 |
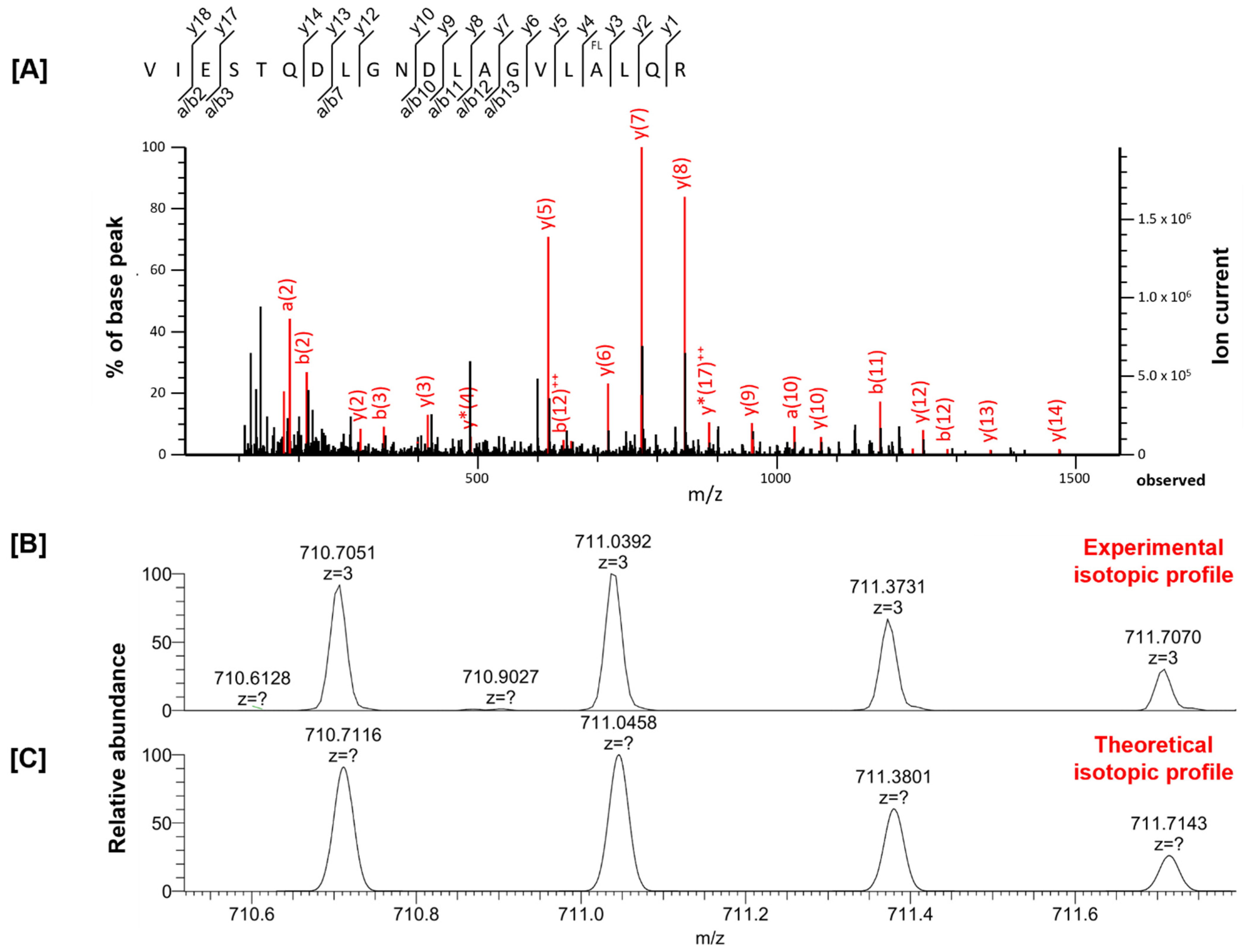
| VIESTQDLGNDLAGVLALQR | fluorination (A) | 2129.0930 | 2129.1128 | −9.29 | 112 | Spectrin beta chain | Sptbn2 | |
| LFATEATSDWLNANNVPATPV AWPSQEGQNPSLSSIR | fluorination (A) | 3985.9498 | 3985.9246 | 6.31 | 79 | Carbamoyl-phosphate synthase [ammonia], mitochondrial | Cps1 | |
| Brain | GVVVFGEPITASLGTDGSHYWSK | fluorination (F) | 2424.1777 | 2424.1762 | 0.61 | 73 | Dihydropyrimidinase-related protein 4 (Fragment) | Dpysl4 |
| DLDLLNPAAR | fluorination (A) | 1114.5835 | 1114.5782 | 4.49 | 47 | Prominin-2 | Prom2 | |
| VIESTQDLGNDLAGVLALQR | fluorination (A) | 2129.1112 | 2129.1128 | −0.78 | 43 | Spectrin beta chain | Sptbn2 | |
| ALSYALR | fluorination (A) | 810.4402 | 810.4399 | 0.37 | 36 | tRNA phosphotransferase 1 | Trpt1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gawor, A.; Gajewski, Z.; Paczek, L.; Czarkowska-Paczek, B.; Konopka, A.; Wryk, G.; Bulska, E. Fluorine-Containing Drug Administration in Rats Results in Fluorination of Selected Proteins in Liver and Brain Tissue. Int. J. Mol. Sci. 2022, 23, 4202. https://doi.org/10.3390/ijms23084202
Gawor A, Gajewski Z, Paczek L, Czarkowska-Paczek B, Konopka A, Wryk G, Bulska E. Fluorine-Containing Drug Administration in Rats Results in Fluorination of Selected Proteins in Liver and Brain Tissue. International Journal of Molecular Sciences. 2022; 23(8):4202. https://doi.org/10.3390/ijms23084202
Chicago/Turabian StyleGawor, Andrzej, Zdzislaw Gajewski, Leszek Paczek, Bozena Czarkowska-Paczek, Anna Konopka, Grzegorz Wryk, and Ewa Bulska. 2022. "Fluorine-Containing Drug Administration in Rats Results in Fluorination of Selected Proteins in Liver and Brain Tissue" International Journal of Molecular Sciences 23, no. 8: 4202. https://doi.org/10.3390/ijms23084202