
Novel Biomarkers of Atherosclerotic Vascular Disease—Latest Insights in the Research Field

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
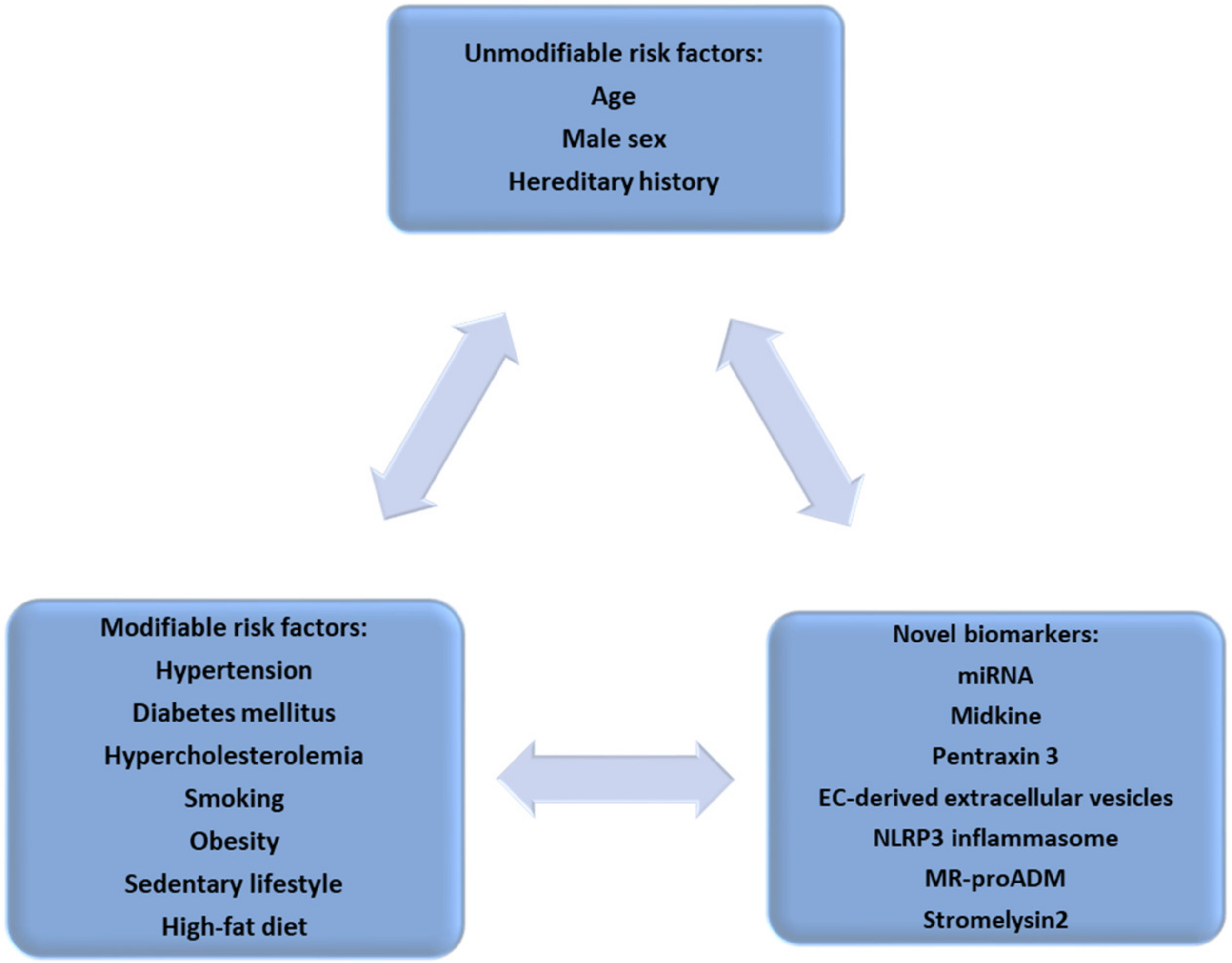
:1. Atherosclerotic Vascular Disease—A Cardiovascular Continuum
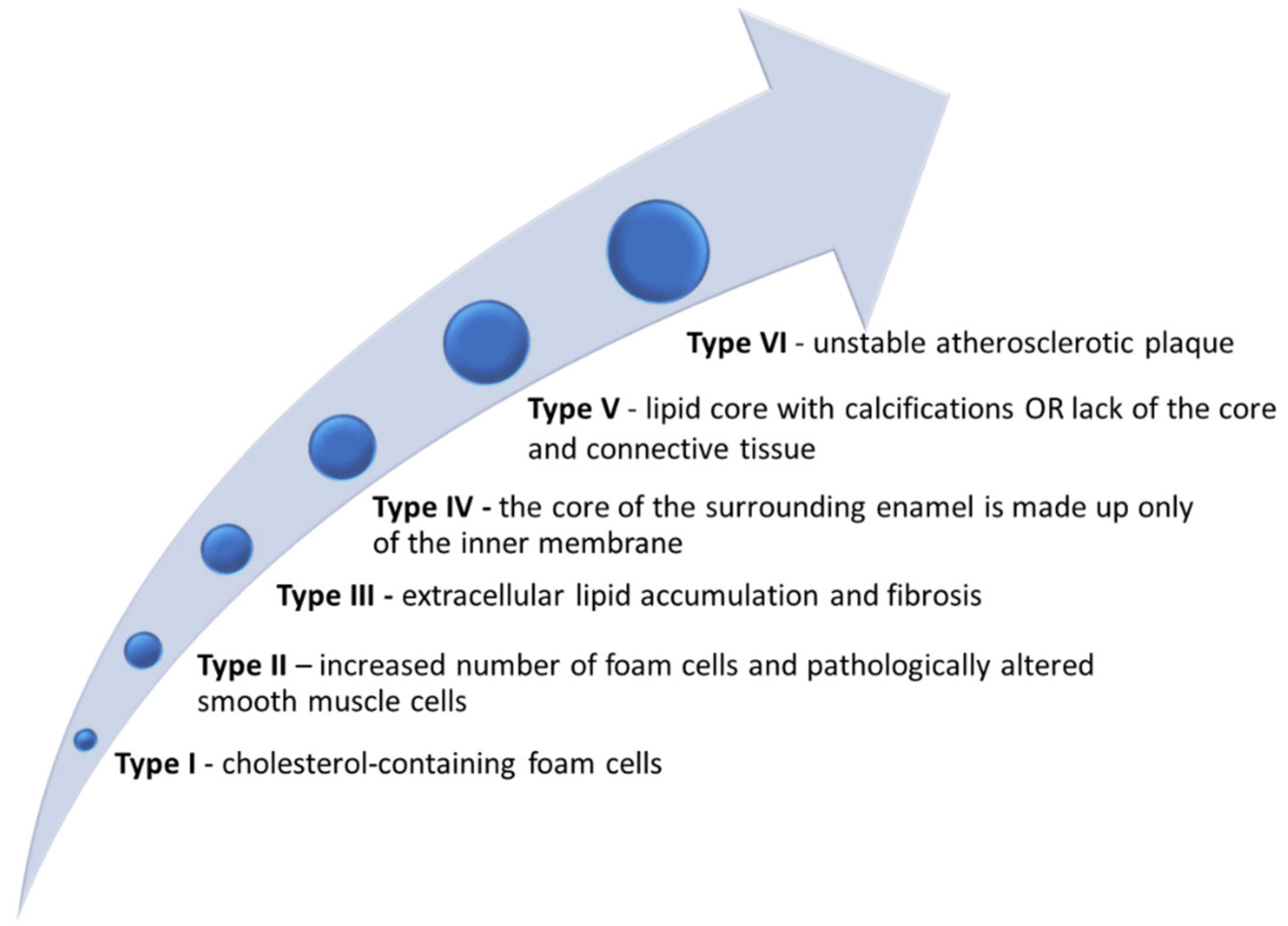
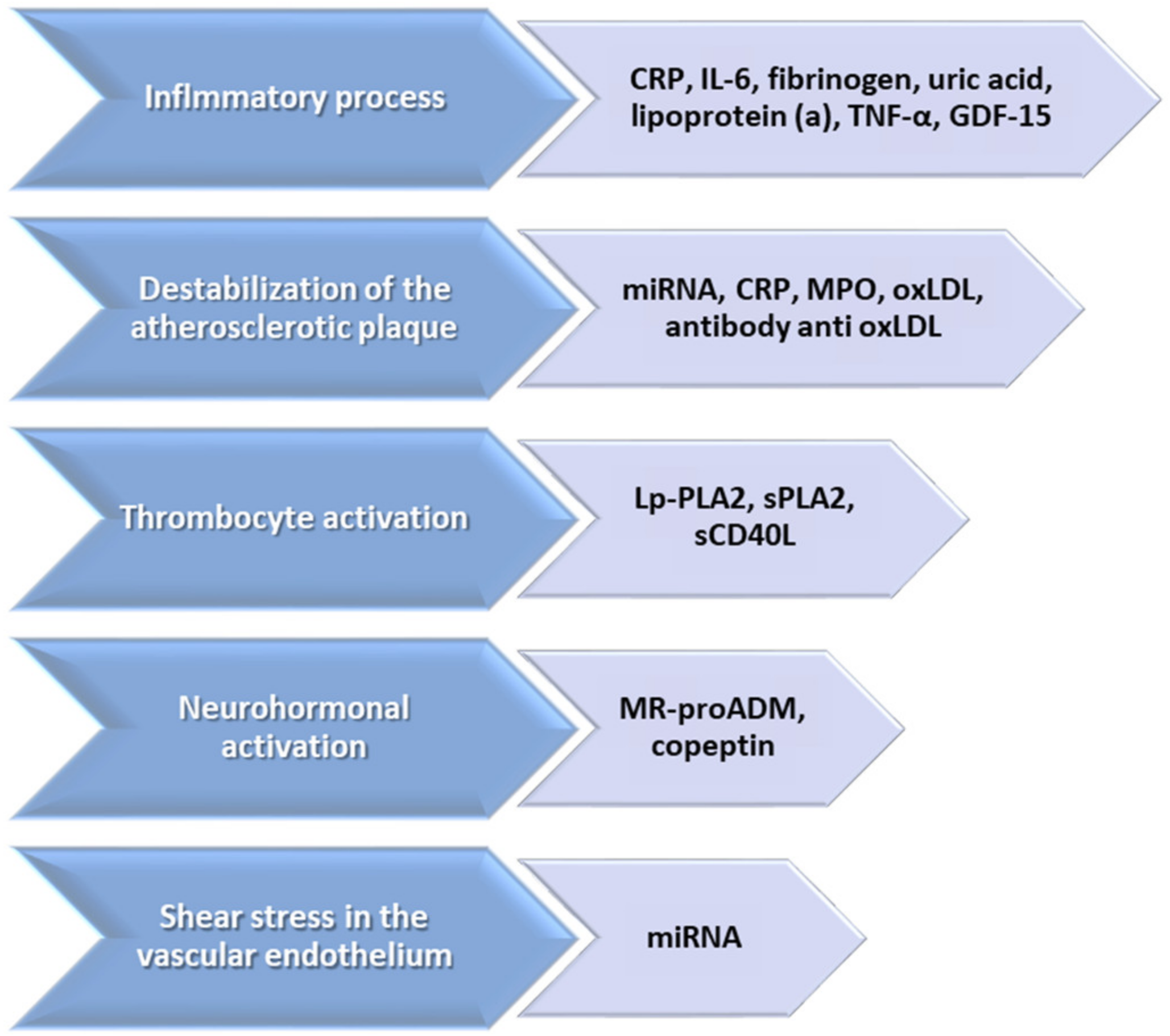
2. Biomarkers—Pathophysiological and Clinical Implications—An Arch in Time
3. Midkine—A Biomarker for All Stages of Atherosclerosis
4. Biomarkers of Destabilization of Atherosclerotic Plaque—microRNA
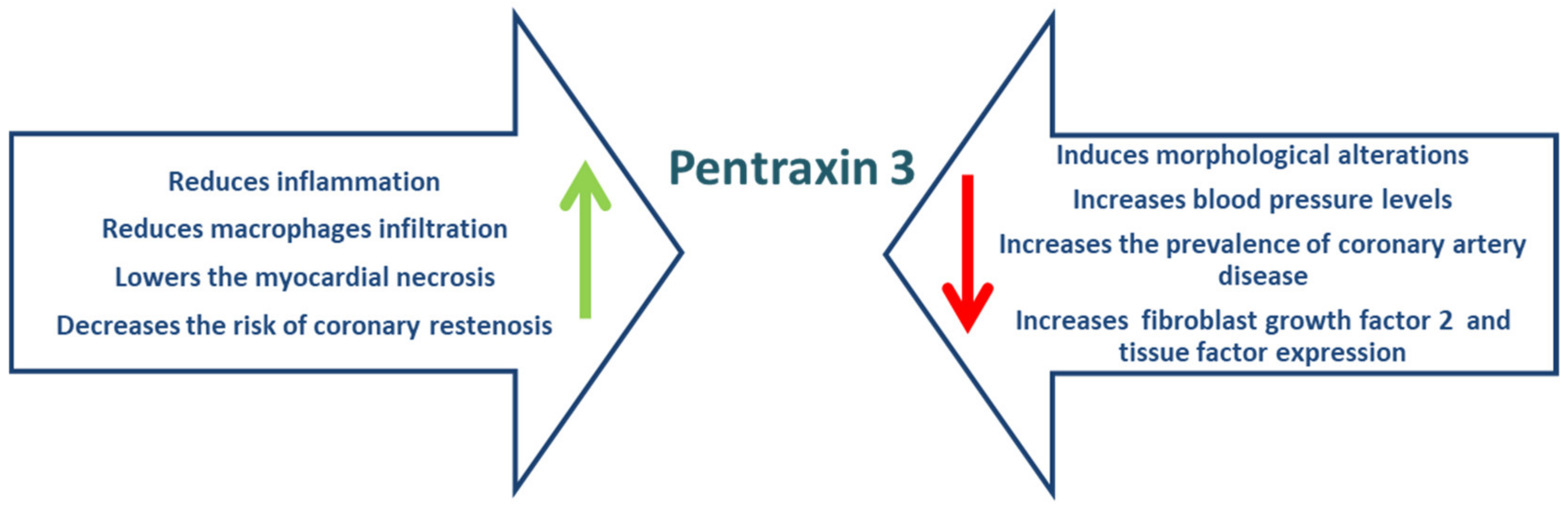
5. Pentraxin 3
6. Endothelial-Cell-Derived Extracellular Vesicles
7. Role of NLRP3 Inflammasome in Atherosclerosis
8. Biomarkers of Thrombocyte Activation—Midregional Proadrenomedullin
9. Stromelysin-2
10. Assessing Atherosclerosis through Artificial Intelligence
11. Conclusions—Atherosclerosis as a “Moving Target”
Author Contributions
Funding
Conflicts of Interest
References
- Botts, S.R.; Fish, J.E.; Howe, K.L. Dysfunctional Vascular Endothelium as a Driver of Atherosclerosis: Emerging Insights into Pathogenesis and Treatment. Front. Pharmacol. 2021, 12, 787541. [Google Scholar] [CrossRef] [PubMed]
- Buliga-Finis, O.N.; Ouatu, A.; Badescu, M.C.; Dima, N.; Tanase, D.M.; Richter, P.; Rezus, C. Beyond the Cardiorenal Syndrome: Pathophysiological Approaches and Biomarkers for Renal and Cardiac Crosstalk. Diagnostics 2022, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Much, A.A.; Maor, E.; Asher, E.; Younis, A.; Xu, Y.; Lu, Y.; Liu, X.; Shu, J.; Bragazzi, N.L. Global, Regional, and National Burden of Ischaemic Heart Disease and Its Attributable Risk Factors, 1990–2017: Results from the Global Burden of Disease Study 2017. Eur. Heart J.-Qual. Care Clin. Outcomes 2022, 8, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Piepoli, M.F.; Hoes, A.W.; Agewall, S.; Albus, C.; Brotons, C.; Catapano, A.L.; Cooney, M.-T.; Corrà, U.; Cosyns, B.; Deaton, C.; et al. 2016 European Guidelines on Cardiovascular Disease Prevention in Clinical Practice: The Sixth Joint Task Force of the European Society of Cardiology and Other Societies on Cardiovascular Disease Prevention in Clinical Practice (Constituted by Representatives of 10 Societies and by Invited Experts) Developed with the Special Contribution of the European Association for Cardiovascular Prevention & Rehabilitation (EACPR). Eur. Heart. J. 2016, 37, 2315–2381. [Google Scholar] [CrossRef]
- Olson, R.E. Discovery of the Lipoproteins, Their Role in Fat Transport and Their Significance as Risk Factors. J. Nutr. 1998, 128, 439S–443S. [Google Scholar] [CrossRef]
- Steinberg, D. Thematic Review Series: The Pathogenesis of Atherosclerosis. An Interpretive History of the Cholesterol Controversy: Part II:The Early Evidence Linking Hypercholesterolemia to Coronary Disease in Humans. J. Lipid Res. 2005, 46, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Konstantinov, I.E.; Jankovic, G.M. Alexander I. Ignatowski: A Pioneer in the Study of Atherosclerosis. Tex. Heart Inst. J. 2013, 40, 246–249. [Google Scholar]
- Seidman, M.A.; Mitchell, R.N.; Stone, J.R. Pathophysiology of Atherosclerosis. In Cellular and Molecular Pathobiology of Cardiovascular Disease; Elsevier: Amsterdam, The Netherlands, 2014; pp. 221–237. ISBN 978-0-12-405206-2. [Google Scholar]
- Mallika, V.; Goswami, B.; Rajappa, M. Atherosclerosis Pathophysiology and the Role of Novel Risk Factors: A Clinicobiochemical Perspective. Angiology 2007, 58, 513–522. [Google Scholar] [CrossRef]
- Cismaru, G.; Serban, T.; Tirpe, A. Ultrasound Methods in the Evaluation of Atherosclerosis: From Pathophysiology to Clinic. Biomedicines 2021, 9, 418. [Google Scholar] [CrossRef]
- Falk, E.; Shah, P.K.; Fuster, V. Coronary Plaque Disruption. Circulation 1995, 92, 657–671. [Google Scholar] [CrossRef]
- Surma, S.; Czober, T.; Lepich, T.; Sierka, O.; Bajor, G. Selected Biomarkers of Atherosclerosis—Clinical Aspects. Acta Angiol. 2020, 26, 28–39. [Google Scholar] [CrossRef]
- Stary, H.C.; Chandler, A.B.; Dinsmore, R.E.; Fuster, V.; Glagov, S.; Insull, W.; Rosenfeld, M.E.; Schwartz, C.J.; Wagner, W.D.; Wissler, R.W. A Definition of Advanced Types of Atherosclerotic Lesions and a Histological Classification of Atherosclerosis: A Report From the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1995, 92, 1355–1374. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J. Assessing the Role of Circulating, Genetic, and Imaging Biomarkers in Cardiovascular Risk Prediction. Circulation 2011, 123, 551–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yla-Herttuala, S.; Bentzon, J.F.; Daemen, M.; Falk, E.; Garcia-Garcia, H.M.; Herrmann, J.; Hoefer, I.; Jauhiainen, S.; Jukema, J.W.; Krams, R.; et al. Stabilization of Atherosclerotic Plaques: An Update. Eur. Heart J. 2013, 34, 3251–3258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biomarkers Definitions Working Group. Biomarkers and Surrogate Endpoints: Preferred Definitions and Conceptual Framework. Clin. Pharmacol. Ther. 2001, 69, 89–95. [Google Scholar] [CrossRef]
- Hoefer, I.E.; Steffens, S.; Ala-Korpela, M.; Bäck, M.; Badimon, L.; Bochaton-Piallat, M.-L.; Boulanger, C.M.; Caligiuri, G.; Dimmeler, S.; Egido, J.; et al. Novel Methodologies for Biomarker Discovery in Atherosclerosis. Eur. Heart J. 2015, 36, 2635–2642. [Google Scholar] [CrossRef] [Green Version]
- Wong, Y.-K.; Tse, H.-F. Circulating Biomarkers for Cardiovascular Disease Risk Prediction in Patients with Cardiovascular Disease. Front. Cardiovasc. Med. 2021, 8, 713191. [Google Scholar] [CrossRef]
- Wielkoszyński, T.; Zalejska-Fiolka, J.; Strzelczyk, J.K.; Owczarek, A.J.; Cholewka, A.; Furmański, M.; Stanek, A. Oxysterols Increase Inflammation, Lipid Marker Levels and Reflect Accelerated Endothelial Dysfunction in Experimental Animals. Mediat. Inflamm. 2018, 2018, 2784701. [Google Scholar] [CrossRef]
- Wang, J.; Tan, G.-J.; Han, L.-N.; Bai, Y.-Y.; He, M.; Liu, H.-B. Novel Biomarkers for Cardiovascular Risk Prediction. J. Geriatr. Cardiol. 2017, 14, 135–150. [Google Scholar] [CrossRef]
- Zhang, H.; Park, Y.; Wu, J.; Chen, X.; Lee, S.; Yang, J.; Dellsperger, K.C.; Zhang, C. Role of TNF-Alpha in Vascular Dysfunction. Clin. Sci. 2009, 116, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Mendall, M.A.; Patel, P.; Asante, M.; Ballam, L.; Morris, J.; Strachan, D.P.; Camm, A.J.; Northfield, T.C. Relation of Serum Cytokine Concentrations to Cardiovascular Risk Factors and Coronary Heart Disease. Heart 1997, 78, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Rifai, N.; Stampfer, M.J.; Hennekens, C.H. Plasma Concentration of Interleukin-6 and the Risk of Future Myocardial Infarction among Apparently Healthy Men. Circulation 2000, 101, 1767–1772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, A.B.; Siegart, N.M.; de Leon, J. Interleukin-6 in Atherosclerosis: Atherogenic or Atheroprotective? Clin. Lipidol. 2017, 12, 14–23. [Google Scholar]
- Biscetti, F.; Nardella, E.; Cecchini, A.L.; Flex, A.; Landolfi, R. Biomarkers of Vascular Disease in Diabetes: The Adipose-Immune System Cross Talk. Intern. Emerg. Med. 2020, 15, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.C.; Yi, T.C.; Weng, H.Y.; Zhang, L.; Li, Y.X.; Mohetaboer, M.; Zhang, Y.; Jiang, J.; Li, J.P.; Huo, Y. Prognostic value of myeloperoxidase concentration in patients with acute coronary syndrome. Zhonghua Xin Xue Guan Bing Za Zhi 2018, 46, 284–291. [Google Scholar] [CrossRef]
- Ley, K.; Huo, Y. VCAM-1 Is Critical in Atherosclerosis. J. Clin. Investig. 2001, 107, 1209–1210. [Google Scholar] [CrossRef]
- Varona, J.F.; Ortiz-Regalón, R.; Sánchez-Vera, I.; López-Melgar, B.; García-Durango, C.; Castellano Vázquez, J.M.; Solís, J.; Fernández-Friera, L.; Vidal-Vanaclocha, F. Soluble ICAM 1 and VCAM 1 Blood Levels Alert on Subclinical Atherosclerosis in Non Smokers with Asymptomatic Metabolic Syndrome. Arch. Med. Res. 2019, 50, 20–28. [Google Scholar] [CrossRef]
- Shrivastava, A.K.; Singh, H.V.; Raizada, A.; Singh, S.K. C-Reactive Protein, Inflammation and Coronary Heart Disease. Egypt. Heart J. 2015, 67, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Orsó, E.; Schmitz, G. Lipoprotein(a) and Its Role in Inflammation, Atherosclerosis and Malignancies. Clin. Res. Cardiol. Suppl. 2017, 12, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Stec, J.J.; Silbershatz, H.; Tofler, G.H.; Matheney, T.H.; Sutherland, P.; Lipinska, I.; Massaro, J.M.; Wilson, P.F.; Muller, J.E.; D’Agostino, R.B. Association of Fibrinogen with Cardiovascular Risk Factors and Cardiovascular Disease in the Framingham Offspring Population. Circulation 2000, 102, 1634–1638. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.-L.; Shi, Y.-H.; Hao, G.; Li, W.; Le, G.-W. Increasing Oxidative Stress with Progressive Hyperlipidemia in Human: Relation between Malondialdehyde and Atherogenic Index. J. Clin. Biochem. Nutr. 2008, 43, 154–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viigimaa, M.; Abina, J.; Zemtsovskaya, G.; Tikhaze, A.; Konovalova, G.; Kumskova, E.; Lankin, V. Malondialdehyde-Modified Low-Density Lipoproteins as Biomarker for Atherosclerosis. Blood Press. 2010, 19, 164–168. [Google Scholar] [CrossRef] [PubMed]
- Alghazeer, R.; Aboulmeedah, E.; Elgahmasi, S.; Alghazir, N.; Almukthar, Z.; Enaami, M.; Rhuma, A. Comparative Evaluation of Antioxidant Enzymes and Serum Selenium Levels in Libyan Atherosclerotic Patients. J. Biosci. Med. 2019, 7, 51–69. [Google Scholar] [CrossRef] [Green Version]
- Stanek, A.; Cholewka, A.; Wielkoszyński, T.; Romuk, E.; Sieroń, K.; Sieroń, A. Increased Levels of Oxidative Stress Markers, Soluble CD40 Ligand, and Carotid Intima-Media Thickness Reflect Acceleration of Atherosclerosis in Male Patients with Ankylosing Spondylitis in Active Phase and without the Classical Cardiovascular Risk Factors. Oxidative Med. Cell. Longev. 2017, 2017, 9712536. [Google Scholar] [CrossRef] [Green Version]
- Weckbach, L.T.; Groesser, L.; Borgolte, J.; Pagel, J.-I.; Pogoda, F.; Schymeinsky, J.; Müller-Höcker, J.; Shakibaei, M.; Muramatsu, T.; Deindl, E.; et al. Midkine Acts as Proangiogenic Cytokine in Hypoxia-Induced Angiogenesis. Am. J. Physiol.-Heart Circ. Physiol. 2012, 303, H429–H438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemoto, Y.; Horiba, M.; Harada, M.; Sakamoto, K.; Takeshita, K.; Murohara, T.; Kadomatsu, K.; Kamiya, K. Midkine Promotes Atherosclerotic Plaque Formation Through Its Pro-Inflammatory, Angiogenic and Anti-Apoptotic Functions in Apolipoprotein E-Knockout Mice. Circ. J. 2018, 82, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-Z.; Wang, G.; Yin, S.-H.; Yu, X.-H. Midkine: A Multifaceted Driver of Atherosclerosis. Clin. Chim. Acta 2021, 521, 251–257. [Google Scholar] [CrossRef]
- Guzel, S.; Cinemre, F.B.; Guzel, E.C.; Kucukyalcin, V.; Kiziler, A.R.; Cavusoglu, C.; Gulyasar, T.; Cinemre, H.; Aydemir, B. Midkine Levels and Its Relationship with Atherosclerotic Risk Factors in Essential Hypertensive Patients. Niger J. Clin. Pract. 2018, 21, 894–900. [Google Scholar] [CrossRef]
- Ou, H.-X.; Huang, Q.; Liu, C.-H.; Xiao, J.; Lv, Y.-C.; Li, X.; Lei, L.-P.; Mo, Z.-C. Midkine Inhibits Cholesterol Efflux by Decreasing ATP-Binding Membrane Cassette Transport Protein A1 via Adenosine Monophosphate-Activated Protein Kinase/Mammalian Target of Rapamycin Signaling in Macrophages. Circ. J. 2020, 84, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.-H.; Fu, Y.-C.; Zhang, D.-W.; Yin, K.; Tang, C.-K. Foam Cells in Atherosclerosis. Clin. Chim. Acta 2013, 424, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Grootaert, M.O.J.; Moulis, M.; Roth, L.; Martinet, W.; Vindis, C.; Bennett, M.R.; De Meyer, G.R.Y. Vascular Smooth Muscle Cell Death, Autophagy and Senescence in Atherosclerosis. Cardiovasc. Res. 2018, 114, 622–634. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Uryga, A.K.; Reinhold, J.; Figg, N.; Baker, L.; Finigan, A.; Gray, K.; Kumar, S.; Clarke, M.; Bennett, M. Vascular Smooth Muscle Cell Senescence Promotes Atherosclerosis and Features of Plaque Vulnerability. Circulation 2015, 132, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Allahverdian, S.; Chehroudi, A.C.; McManus, B.M.; Abraham, T.; Francis, G.A. Contribution of Intimal Smooth Muscle Cells to Cholesterol Accumulation and Macrophage-like Cells in Human Atherosclerosis. Circulation 2014, 129, 1551–1559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basatemur, G.L.; Jørgensen, H.F.; Clarke, M.C.H.; Bennett, M.R.; Mallat, Z. Vascular Smooth Muscle Cells in Atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef]
- Masuda, T.; Maeda, K.; Sato, W.; Kosugi, T.; Sato, Y.; Kojima, H.; Kato, N.; Ishimoto, T.; Tsuboi, N.; Uchimura, K.; et al. Growth Factor Midkine Promotes T-Cell Activation through Nuclear Factor of Activated T Cells Signaling and Th1 Cell Differentiation in Lupus Nephritis. Am. J. Pathol. 2017, 187, 740–751. [Google Scholar] [CrossRef] [Green Version]
- Kosugi, T.; Yuzawa, Y.; Sato, W.; Arata-Kawai, H.; Suzuki, N.; Kato, N.; Matsuo, S.; Kadomatsu, K. Midkine Is Involved in Tubulointerstitial Inflammation Associated with Diabetic Nephropathy. Lab. Investig 2007, 87, 903–913. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Thewke, D.P.; Su, Y.R.; Linton, M.F.; Fazio, S.; Sinensky, M.S. Reduced Macrophage Apoptosis Is Associated with Accelerated Atherosclerosis in Low-Density Lipoprotein Receptor-Null Mice. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 174–179. [Google Scholar] [CrossRef] [Green Version]
- Farahi, L.; Sinha, S.K.; Lusis, A.J. Roles of Macrophages in Atherogenesis. Front. Pharm. 2021, 12, 785220. [Google Scholar] [CrossRef]
- Erbilgin, A.; Seldin, M.M.; Wu, X.; Mehrabian, M.; Zhou, Z.; Qi, H.; Dabirian, K.S.; Sevag Packard, R.R.; Hsieh, W.; Bensinger, S.J.; et al. Transcription Factor Zhx2 Deficiency Reduces Atherosclerosis and Promotes Macrophage Apoptosis in Mice. Arter. Thromb. Vasc. Biol. 2018, 38, 2016–2027. [Google Scholar] [CrossRef] [Green Version]
- Harada, M.; Hojo, M.; Kamiya, K.; Kadomatsu, K.; Murohara, T.; Kodama, I.; Horiba, M. Exogenous Midkine Administration Prevents Cardiac Remodeling in Pacing-Induced Congestive Heart Failure of Rabbits. Heart Vessel. 2016, 31, 96–104. [Google Scholar] [CrossRef]
- Kitahara, T.; Shishido, T.; Suzuki, S.; Katoh, S.; Sasaki, T.; Ishino, M.; Nitobe, J.; Miyamoto, T.; Miyashita, T.; Watanabe, T.; et al. Serum Midkine as a Predictor of Cardiac Events in Patients with Chronic Heart Failure. J. Card Fail 2010, 16, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Przybylowski, P.; Malyszko, J.; Malyszko, J.S. Serum Midkine Is Related to NYHA Class and Cystatin C in Heart Transplant Recipients. Transpl. Proc. 2010, 42, 3704–3707. [Google Scholar] [CrossRef] [PubMed]
- Woulfe, K.C.; Sucharov, C.C. Midkine’s Role in Cardiac Pathology. J. Cardiovasc. Dev. Dis. 2017, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadomatsu, K.; Bencsik, P.; Görbe, A.; Csonka, C.; Sakamoto, K.; Kishida, S.; Ferdinandy, P. Therapeutic Potential of Midkine in Cardiovascular Disease. Br. J. Pharm. 2014, 171, 936–944. [Google Scholar] [CrossRef] [Green Version]
- Becker, R.C.; Owens, A.P.; Sadayappan, S. Tissue-Level Inflammation and Ventricular Remodeling in Hypertrophic Cardiomyopathy. J. Thromb. Thrombolysis 2020, 49, 177–183. [Google Scholar] [CrossRef]
- Salaru, D.L.; Albert, C.; Königsmark, U.; Brandt, S.; Halloul, Z.; Heller, A.; Heidecke, H.; Dragun, D.; Mertens, P.R. Serum Levels for Midkine, a Heparin-Binding Growth Factor, Inversely Correlate with Angiotensin and Endothelin Receptor Autoantibody Titers in Patients with Macroangiopathy. Int. Angiol. 2014, 33, 372–378. [Google Scholar]
- Chen, Z.; Zhu, Y.; Zhang, L. Study of Three Novel Biomarkers, MR-ProADM, Midkine, and Stromelysin2, and Peripheral Atherosclerosis in a Chinese Han Population: A Case-Control Study. Eur. J. Inflamm. 2020, 18, 205873922096055. [Google Scholar] [CrossRef]
- Smolarz, B.; Durczyński, A.; Romanowicz, H.; Szyłło, K.; Hogendorf, P. MiRNAs in Cancer (Review of Literature). Int. J. Mol. Sci. 2022, 23, 2805. [Google Scholar] [CrossRef]
- Lee, L.W.; Zhang, S.; Etheridge, A.; Ma, L.; Martin, D.; Galas, D.; Wang, K. Complexity of the MicroRNA Repertoire Revealed by Next-Generation Sequencing. RNA 2010, 16, 2170–2180. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Su, B. Small but Influential: The Role of MicroRNAs on Gene Regulatory Network and 3′UTR Evolution. J. Genet. Genom. 2009, 36, 1–6. [Google Scholar] [CrossRef]
- Rossbach, M. Small Non-Coding RNAs as Novel Therapeutics. Curr. Mol. Med. 2010, 10, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Pozniak, T.; Shcharbin, D.; Bryszewska, M. Circulating MicroRNAs in Medicine. Int. J. Mol. Sci. 2022, 23, 3996. [Google Scholar] [CrossRef] [PubMed]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum MicroRNAs Are Promising Novel Biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733–1741. [Google Scholar] [CrossRef]
- Solly, E.L.; Dimasi, C.G.; Bursill, C.A.; Psaltis, P.J.; Tan, J.T.M. MicroRNAs as Therapeutic Targets and Clinical Biomarkers in Atherosclerosis. JCM 2019, 8, 2199. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs Are Transported in Plasma and Delivered to Recipient Cells by High-Density Lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Glass, C.K.; Witztum, J.L. Atherosclerosis: The Road Ahead. Cell 2001, 104, 503–516. [Google Scholar] [CrossRef] [Green Version]
- Pentikäinen, M.O.; Öörni, K.; Ala-Korpela, M.; Kovanen, P.T. Modified LDL – Trigger of Atherosclerosis and Inflammation in the Arterial Intima. J. Intern. Med. 2000, 247, 359–370. [Google Scholar] [CrossRef]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. MiR-122 Regulation of Lipid Metabolism Revealed by in Vivo Antisense Targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Belkin, N.; Feinberg, M.W. Endothelial MicroRNAs and Atherosclerosis. Curr. Atheroscler. Rep. 2013, 15, 372. [Google Scholar] [CrossRef]
- Sun, X.; He, S.; Wara, A.K.M.; Icli, B.; Shvartz, E.; Tesmenitsky, Y.; Belkin, N.; Li, D.; Blackwell, T.S.; Sukhova, G.K.; et al. Systemic Delivery of MicroRNA-181b Inhibits Nuclear Factor-ΚB Activation, Vascular Inflammation, and Atherosclerosis in Apolipoprotein E–Deficient Mice. Circ. Res. 2014, 114, 32–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Y.; Yuan, J.; Zhang, F.; Lei, Q.; Zhang, T.; Li, K.; Guo, J.; Hong, Y.; Bu, G.; Lv, X.; et al. MicroRNA-181a-5p and MicroRNA-181a-3p Cooperatively Restrict Vascular Inflammation and Atherosclerosis. Cell Death Dis. 2019, 10, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, Z.; Tan, J.; Miao, Y.; Zhang, Q. The Role of Microvesicles Containing MicroRNAs in Vascular Endothelial Dysfunction. J. Cell Mol. Med. 2019, 23, 7933–7945. [Google Scholar] [CrossRef] [PubMed]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, E.; Pap, E.; Kittel, A.; et al. Membrane Vesicles, Current State-of-the-Art: Emerging Role of Extracellular Vesicles. Cell Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral Sphingomyelinase 2 (NSMase2)-Dependent Exosomal Transfer of Angiogenic MicroRNAs Regulate Cancer Cell Metastasis. J. Biol. Chem. 2013, 288, 10849–10859. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory Mechanisms and Intercellular Transfer of MicroRNAs in Living Cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [Green Version]
- Jansen, F.; Yang, X.; Proebsting, S.; Hoelscher, M.; Przybilla, D.; Baumann, K.; Schmitz, T.; Dolf, A.; Endl, E.; Franklin, B.S.; et al. MicroRNA Expression in Circulating Microvesicles Predicts Cardiovascular Events in Patients with Coronary Artery Disease. J. Am. Heart Assoc. 2014, 3, e001249. [Google Scholar] [CrossRef] [Green Version]
- Peluso, I.; Morabito, G.; Urban, L.; Ioannone, F.; Serafini, M. Oxidative Stress in Atherosclerosis Development: The Central Role of LDL and Oxidative Burst. Endocr. Metab. Immune Disord. Drug Targets 2012, 12, 351–360. [Google Scholar] [CrossRef]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef]
- Joris, V.; Gomez, E.L.; Menchi, L.; Lobysheva, I.; Di Mauro, V.; Esfahani, H.; Condorelli, G.; Balligand, J.-L.; Catalucci, D.; Dessy, C. MicroRNA-199a-3p and MicroRNA-199a-5p Take Part to a Redundant Network of Regulation of the NOS (NO Synthase)/NO Pathway in the Endothelium. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2345–2357. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-Y.; Chen, Y.-T.; Su, Y.-W.; Chang, C.-C.; Huang, P.-H.; Lin, S.-J. Statin Therapy Reduces Future Risk of Lower-Limb Amputation in Patients with Diabetes and Peripheral Artery Disease. J. Clin. Endocrinol. Metab. 2017, 102, 2373–2381. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Mi, X.; Chen, Y.; Feng, C.; Hou, Z.; Hui, R.; Zhang, W. MicroRNA-216a Induces Endothelial Senescence and Inflammation via Smad3/IκBα Pathway. J. Cell. Mol. Med. 2018, 22, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, M.-A.; Karunakaran, D.; Geoffrion, M.; Cheng, H.S.; Tandoc, K.; Perisic Matic, L.; Hedin, U.; Maegdefessel, L.; Fish, J.E.; Rayner, K.J. Extracellular Vesicles Secreted by Atherogenic Macrophages Transfer MicroRNA to Inhibit Cell Migration. Arter. Thromb. Vasc. Biol. 2018, 38, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonyushkin, T.; Binder, C.J. Extracellular Vesicles Act as Messengers of Macrophages Sensing Atherogenic Stimuli. Arter. Thromb. Vasc. Biol. 2018, 38, 2–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Atanasov, A.G. The MicroRNAs Regulating Vascular Smooth Muscle Cell Proliferation: A Minireview. Int. J. Mol. Sci. 2019, 20, 324. [Google Scholar] [CrossRef] [Green Version]
- Alshanwani, A.R.; Riches-Suman, K.; O’Regan, D.J.; Wood, I.C.; Turner, N.A.; Porter, K.E. MicroRNA-21 Drives the Switch to a Synthetic Phenotype in Human Saphenous Vein Smooth Muscle Cells. IUBMB Life 2018, 70, 649–657. [Google Scholar] [CrossRef] [PubMed]
- MicroRNA-1 Inhibits Myocardin-Induced Contractility of Human Vascular Smooth Muscle Cells-Jiang-2010-Journal of Cellular Physiology-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/10.1002/jcp.22230 (accessed on 26 March 2022).
- Lin, Y.; Liu, X.; Cheng, Y.; Yang, J.; Huo, Y.; Zhang, C. Involvement of MicroRNAs in Hydrogen Peroxide-Mediated Gene Regulation and Cellular Injury Response in Vascular Smooth Muscle Cells *. J. Biol. Chem. 2009, 284, 7903–7913. [Google Scholar] [CrossRef] [Green Version]
- Jin, H.; Li, D.Y.; Chernogubova, E.; Sun, C.; Busch, A.; Eken, S.M.; Saliba-Gustafsson, P.; Winter, H.; Winski, G.; Raaz, U.; et al. Local Delivery of MiR-21 Stabilizes Fibrous Caps in Vulnerable Atherosclerotic Lesions. Mol. Ther. 2018, 26, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Eken, S.M.; Jin, H.; Chernogubova, E.; Li, Y.; Simon, N.; Sun, C.; Korzunowicz, G.; Busch, A.; Bäcklund, A.; Österholm, C.; et al. MicroRNA-210 Enhances Fibrous Cap Stability in Advanced Atherosclerotic Lesions. Circ. Res. 2017, 120, 633–644. [Google Scholar] [CrossRef]
- Sedding, D.G.; Boyle, E.C.; Demandt, J.A.F.; Sluimer, J.C.; Dutzmann, J.; Haverich, A.; Bauersachs, J. Vasa Vasorum Angiogenesis: Key Player in the Initiation and Progression of Atherosclerosis and Potential Target for the Treatment of Cardiovascular Disease. Front. Immunol. 2018, 9, 706. [Google Scholar] [CrossRef] [Green Version]
- Boyle, E.C.; Sedding, D.G.; Haverich, A. Targeting Vasa Vasorum Dysfunction to Prevent Atherosclerosis. Vasc. Pharm. 2017, 96–98, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Urbich, C.; Kuehbacher, A.; Dimmeler, S. Role of MicroRNAs in Vascular Diseases, Inflammation, and Angiogenesis. Cardiovasc. Res. 2008, 79, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Xun, M.; Chen, Y. Circular RNAs: Regulators of Vascular Smooth Muscle Cells in Cardiovascular Diseases. J. Mol. Med. 2022, 100, 519–535. [Google Scholar] [CrossRef]
- Parahuleva, M.S.; Lipps, C.; Parviz, B.; Hölschermann, H.; Schieffer, B.; Schulz, R.; Euler, G. MicroRNA Expression Profile of Human Advanced Coronary Atherosclerotic Plaques. Sci. Rep. 2018, 8, 7823. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Silva, D.; Carneiro, F.; Almeida, K.; Fernandes-Santos, C. Role of MiRNAs on the Pathophysiology of Cardiovascular Diseases. Arq. Bras. Cardiol. 2018, 11, 738–746. [Google Scholar] [CrossRef]
- Zhang, X.; Shao, S.; Geng, H.; Yu, Y.; Wang, C.; Liu, Z.; Yu, C.; Jiang, X.; Deng, Y.; Gao, L.; et al. Expression Profiles of Six Circulating MicroRNAs Critical to Atherosclerosis in Patients with Subclinical Hypothyroidism: A Clinical Study. J. Clin. Endocrinol. Metab. 2014, 99, E766–E774. [Google Scholar] [CrossRef] [Green Version]
- Pereira-da-Silva, T.; Coutinho Cruz, M.; Carrusca, C.; Cruz Ferreira, R.; Napoleão, P.; Mota Carmo, M. Circulating MicroRNA Profiles in Different Arterial Territories of Stable Atherosclerotic Disease: A Systematic Review. Am. J. Cardiovasc. Dis. 2018, 8, 1–13. [Google Scholar]
- Fichtlscherer, S.; De Rosa, S.; Fox, H.; Schwietz, T.; Fischer, A.; Liebetrau, C.; Weber, M.; Hamm, C.W.; Röxe, T.; Müller-Ardogan, M.; et al. Circulating MicroRNAs in Patients with Coronary Artery Disease. Circ. Res. 2010, 107, 677–684. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lian, Y.; Wen, X.; Guo, J.; Wang, Z.; Jiang, S.; Hu, Y. Expression of MiR-126 and Its Potential Function in Coronary Artery Disease. Afr. Health Sci. 2017, 17, 474–480. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.; Baker, M.B.; Patel, R.S.; Quyyumi, A.A.; Bao, G.; Searles, C.D. MicroRNA Expression Profile in CAD Patients and the Impact of ACEI/ARB. Cardiol. Res. Pract. 2011, 2011, 532915. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Wang, Q.; Zhu, R.; Yang, J. Circulating Non-Coding RNAs as Potential Biomarkers for Ischemic Stroke: A Systematic Review. J. Mol. Neurosci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Akella, A.; Bhattarai, S.; Dharap, A. Long Noncoding RNAs in the Pathophysiology of Ischemic Stroke. Neuromolecular Med. 2019, 21, 474–483. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, P.; Dong, X.; Jin, J.; Xu, Y. The Role of LncRNAs in Ischemic Stroke. Neurochem. Int. 2021, 147, 105019. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Zhan, X.; He, M.; Wang, J.; Qiu, X. MiR-135b Levels in the Peripheral Blood Serve as a Marker Associated with Acute Ischemic Stroke. Exp. Med. 2020. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Kan, P.; Ma, Z.; Wang, Y.; Song, W.; Huang, C.; Zhang, B. Exploring the Potential Value of MiR-148b-3p, MiR-151b and MiR-27b-3p as Biomarkers in Acute Ischemic Stroke. Biosci. Rep. 2018, 38, BSR20181033. [Google Scholar] [CrossRef] [Green Version]
- Mahjoob, G.; Ahmadi, Y.; Fatima Rajani, H.; Khanbabaei, N.; Abolhasani, S. Circulating MicroRNAs as Predictive Biomarkers of Coronary Artery Diseases in Type 2 Diabetes Patients. J. Clin. Lab. Anal. 2022, e24380. [Google Scholar] [CrossRef]
- Du, Y.; Yang, S.H.; Li, S.; Cui, C.J.; Zhang, Y.; Zhu, C.G.; Guo, Y.L.; Wu, N.Q.; Gao, Y.; Sun, J.; et al. Circulating MicroRNAs as Novel Diagnostic Biomarkers for Very Early-Onset (≤40 Years) Coronary Artery Disease. Biomed. Environ. Sci. 2016, 29, 545–554. [Google Scholar] [CrossRef]
- Zhang, P.; Liang, T.; Chen, Y.; Wang, X.; Wu, T.; Xie, Z.; Luo, J.; Yu, Y.; Yu, H. Circulating Exosomal MiRNAs as Novel Biomarkers for Stable Coronary Artery Disease. Biomed. Res. Int. 2020, 2020, 3593962. [Google Scholar] [CrossRef]
- Šatrauskienė, A.; Navickas, R.; Laucevičius, A.; Krilavičius, T.; Užupytė, R.; Zdanytė, M.; Ryliškytė, L.; Jucevičienė, A.; Holvoet, P. Mir-1, MiR-122, MiR-132, and MiR-133 Are Related to Subclinical Aortic Atherosclerosis Associated with Metabolic Syndrome. Int. J. Environ. Res. Public Health 2021, 18, 1483. [Google Scholar] [CrossRef]
- Xu, K.; Chen, C.; Wu, Y.; Wu, M.; Lin, L. Advances in MiR-132-Based Biomarker and Therapeutic Potential in the Cardiovascular System. Front. Pharm. 2021, 12, 751487. [Google Scholar] [CrossRef]
- Li, H.; Zhang, P.; Li, F.; Yuan, G.; Wang, X.; Zhang, A.; Li, F. Plasma MiR-22-5p, MiR-132-5p, and MiR-150-3p Are Associated with Acute Myocardial Infarction. BioMed Res. Int. 2019, 2019, 5012648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; de Ronde, M.W.J.; Kok, M.G.M.; Beijk, M.A.; De Winter, R.J.; van der Wal, A.C.; Sondermeijer, B.M.; Meijers, J.C.M.; Creemers, E.E.; Pinto-Sietsma, S.-J. MiR-223-3p and MiR-122-5p as Circulating Biomarkers for Plaque Instability. Open Heart 2020, 7, e001223. [Google Scholar] [CrossRef] [PubMed]
- Hromadka, M.; Motovska, Z.; Hlinomaz, O.; Kala, P.; Tousek, F.; Jarkovsky, J.; Beranova, M.; Jansky, P.; Svoboda, M.; Krepelkova, I.; et al. MiR-126-3p and MiR-223-3p as Biomarkers for Prediction of Thrombotic Risk in Patients with Acute Myocardial Infarction and Primary Angioplasty. J. Pers. Med. 2021, 11, 508. [Google Scholar] [CrossRef]
- Jebari-Benslaiman, S.; Galicia-García, U.; Larrea-Sebal, A.; Olaetxea, J.R.; Alloza, I.; Vandenbroeck, K.; Benito-Vicente, A.; Martín, C. Pathophysiology of Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 3346. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Zhu, Q.; Zhao, Z.; Zu, X.; Liu, J. Magic and Mystery of MicroRNA-32. J. Cell Mol. Med. 2021, 25, 8588–8601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cai, H.; Zhu, M.; Qian, Y.; Lin, S.; Li, X. Circulating MicroRNAs as Biomarkers for Severe Coronary Artery Disease. Medicine 2020, 99, e19971. [Google Scholar] [CrossRef] [PubMed]
- Rozhkova, A.V.; Dmitrieva, V.G.; Nosova, E.V.; Dergunov, A.D.; Limborska, S.A.; Dergunova, L.V. Genomic Variants and Multilevel Regulation of ABCA1, ABCG1, and SCARB1 Expression in Atherogenesis. J. Cardiovasc. Dev. Dis. 2021, 8, 170. [Google Scholar] [CrossRef]
- Schumacher, D.; Curaj, A.; Simsekyilmaz, S.; Schober, A.; Liehn, E.A.; Mause, S.F. MiR155 Deficiency Reduces Myofibroblast Density but Fails to Improve Cardiac Function after Myocardial Infarction in Dyslipidemic Mouse Model. Int. J. Mol. Sci. 2021, 22, 5480. [Google Scholar] [CrossRef]
- Li, J.; Cai, S.X.; He, Q.; Zhang, H.; Friedberg, D.; Wang, F.; Redington, A.N. Intravenous MiR-144 Reduces Left Ventricular Remodeling after Myocardial Infarction. Basic Res. Cardiol. 2018, 113, 36. [Google Scholar] [CrossRef]
- Casula, M.; Montecucco, F.; Bonaventura, A.; Liberale, L.; Vecchié, A.; Dallegri, F.; Carbone, F. Update on the Role of Pentraxin 3 in Atherosclerosis and Cardiovascular Diseases. Vasc. Pharmacol. 2017, 99, 1–12. [Google Scholar] [CrossRef]
- Pepys, M.B.; Baltz, M.L. Acute Phase Proteins with Special Reference to C-Reactive Protein and Related Proteins (Pentaxins) and Serum Amyloid A Protein. In Advances in Immunology; Dixon, F.J., Kunkel, H.G., Eds.; Academic Press: Cambridge, MA, USA, 1983; Volume 34, pp. 141–212. [Google Scholar]
- Richter, P.; Burlui, A.; Bratoiu, I.; Cardoneanu, A.; Rezus, C.; Rezus, E. A Review of Anti-C Reactive Protein Antibodies in Systemic Lupus Erythematosus. J. Interdiscip. Med. 2021, 6, 60–66. [Google Scholar] [CrossRef]
- Vilahur, G.; Badimon, L. Biological Actions of Pentraxins. Vasc. Pharmacol. 2015, 73, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Kunes, P.; Holubcova, Z.; Kolackova, M.; Krejsek, J. Pentraxin 3(PTX 3): An Endogenous Modulator of the Inflammatory Response. Mediat. Inflamm. 2012, 2012, 920517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deban, L.; Russo, R.C.; Sironi, M.; Moalli, F.; Scanziani, M.; Zambelli, V.; Cuccovillo, I.; Bastone, A.; Gobbi, M.; Valentino, S.; et al. Regulation of Leukocyte Recruitment by the Long Pentraxin PTX3. Nat. Immunol. 2010, 11, 328–334. [Google Scholar] [CrossRef]
- de la Torre, Y.M.; Fabbri, M.; Jaillon, S.; Bastone, A.; Nebuloni, M.; Vecchi, A.; Mantovani, A.; Garlanda, C. Evolution of the Pentraxin Family: The New Entry PTX4. J. Immunol. 2010, 184, 5055–5064. [Google Scholar] [CrossRef] [Green Version]
- Klouche, M.; Peri, G.; Knabbe, C.; Eckstein, H.-H.; Schmid, F.-X.; Schmitz, G.; Mantovani, A. Modified Atherogenic Lipoproteins Induce Expression of Pentraxin-3 by Human Vascular Smooth Muscle Cells. Atherosclerosis 2004, 175, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Napoleone, E.; Di Santo, A.; Bastone, A.; Peri, G.; Mantovani, A.; de Gaetano, G.; Donati, M.B.; Lorenzet, R. Long Pentraxin PTX3 Upregulates Tissue Factor Expression in Human Endothelial Cells: A Novel Link between Vascular Inflammation and Clotting Activation. Arter. Thromb. Vasc. Biol. 2002, 22, 782–787. [Google Scholar] [CrossRef] [Green Version]
- Napoleone, E.; di Santo, A.; Peri, G.; Mantovani, A.; de Gaetano, G.; Donati, M.B.; Lorenzet, R. The Long Pentraxin PTX3 Up-Regulates Tissue Factor in Activated Monocytes: Another Link between Inflammation and Clotting Activation. J. Leukoc. Biol. 2004, 76, 203–209. [Google Scholar] [CrossRef]
- Zlibut, A.; Bocsan, I.C.; Agoston-Coldea, L. Pentraxin-3 and Endothelial Dysfunction. In Advances in Clinical Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; Volume 91, pp. 163–179. ISBN 978-0-12-817471-5. [Google Scholar]
- O’Neill, C.L.; Guduric-Fuchs, J.; Chambers, S.E.J.; O’Doherty, M.; Bottazzi, B.; Stitt, A.W.; Medina, R.J. Endothelial Cell-Derived Pentraxin 3 Limits the Vasoreparative Therapeutic Potential of Circulating Angiogenic Cells. Cardiovasc. Res. 2016, 112, 677–688. [Google Scholar] [CrossRef] [Green Version]
- Jenny, N.S.; Arnold, A.M.; Kuller, L.H.; Tracy, R.P.; Psaty, B.M. Associations of Pentraxin 3 with Cardiovascular Disease and All-Cause Death: The Cardiovascular Health Study. Arter. Thromb. Vasc. Biol. 2009, 29, 594–599. [Google Scholar] [CrossRef]
- Jenny, N.S.; Blumenthal, R.S.; Kronmal, R.A.; Rotter, J.I.; Siscovick, D.S.; Psaty, B.M. Associations of Pentraxin 3 with Cardiovascular Disease: The Multi-Ethnic Study of Atherosclerosis. J. Thromb. Haemost. 2014, 12, 999–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Kawano, Y.; Imamura, T.; Kawakita, K.; Sagara, M.; Matsuo, T.; Kakitsubata, Y.; Ishikawa, T.; Kitamura, K.; Hatakeyama, K.; et al. Reciprocal Contribution of Pentraxin 3 and C-Reactive Protein to Obesity and Metabolic Syndrome. Obesity 2010, 18, 1871–1874. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Sugiyama, A.; Reid, P.C.; Ito, Y.; Miyauchi, K.; Mukai, S.; Sagara, M.; Miyamoto, K.; Satoh, H.; Kohno, I.; et al. Establishment of a High Sensitivity Plasma Assay for Human Pentraxin3 as a Marker for Unstable Angina Pectoris. Arter. Thromb. Vasc. Biol. 2007, 27, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Kotooka, N.; Inoue, T.; Aoki, S.; Anan, M.; Komoda, H.; Node, K. Prognostic Value of Pentraxin 3 in Patients with Chronic Heart Failure. Int. J. Cardiol. 2008, 130, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Savchenko, A.; Imamura, M.; Ohashi, R.; Jiang, S.; Kawasaki, T.; Hasegawa, G.; Emura, I.; Iwanari, H.; Sagara, M.; Tanaka, T.; et al. Expression of Pentraxin 3 (PTX3) in Human Atherosclerotic Lesions. J. Pathol. 2008, 215, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Nerkiz, P.; Doganer, Y.C.; Aydogan, U.; Akbulut, H.; Parlak, A.; Aydogdu, A.; Sari, O.; Cayci, T.; Barcin, C.; Koc, B. Serum Pentraxin-3 Level in Patients Who Underwent Coronary Angiography and Relationship with Coronary Atherosclerosis. Med. Princ. Pract. 2015, 24, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Ristagno, G.; Fumagalli, F.; Bottazzi, B.; Mantovani, A.; Olivari, D.; Novelli, D.; Latini, R. Pentraxin 3 in Cardiovascular Disease. Front. Immunol. 2019, 10, 823. [Google Scholar] [CrossRef] [PubMed]
- Salio, M.; Chimenti, S.; De Angelis, N.; Molla, F.; Maina, V.; Nebuloni, M.; Pasqualini, F.; Latini, R.; Garlanda, C.; Mantovani, A. Cardioprotective Function of the Long Pentraxin PTX3 in Acute Myocardial Infarction. Circulation 2008, 117, 1055–1064. [Google Scholar] [CrossRef]
- Norata, G.D.; Garlanda, C.; Catapano, A.L. The Long Pentraxin PTX3: A Modulator of the Immunoinflammatory Response in Atherosclerosis and Cardiovascular Diseases. Trends Cardiovasc. Med. 2010, 20, 35–40. [Google Scholar] [CrossRef]
- Nauta, A.J.; Bottazzi, B.; Mantovani, A.; Salvatori, G.; Kishore, U.; Schwaeble, W.J.; Gingras, A.R.; Tzima, S.; Vivanco, F.; Egido, J.; et al. Biochemical and Functional Characterization of the Interaction between Pentraxin 3 and C1q. Eur. J. Immunol. 2003, 33, 465–473. [Google Scholar] [CrossRef]
- Yasunaga, T.; Ikeda, S.; Koga, S.; Nakata, T.; Yoshida, T.; Masuda, N.; Kohno, S.; Maemura, K. Plasma Pentraxin 3 Is a More Potent Predictor of Endothelial Dysfunction than High-Sensitive C-Reactive Protein. Int. Heart J. 2014, 55, 160–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrizzo, A.; Lenzi, P.; Procaccini, C.; Damato, A.; Biagioni, F.; Ambrosio, M.; Amodio, G.; Remondelli, P.; Del Giudice, C.; Izzo, R.; et al. Pentraxin 3 Induces Vascular Endothelial Dysfunction Through a P-Selectin/Matrix Metalloproteinase-1 Pathway. Circulation 2015, 131, 1495–1505; discussion 1505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latini, R.; Maggioni, A.P.; Peri, G.; Gonzini, L.; Lucci, D.; Mocarelli, P.; Vago, L.; Pasqualini, F.; Signorini, S.; Soldateschi, D.; et al. Prognostic Significance of the Long Pentraxin PTX3 in Acute Myocardial Infarction. Circulation 2004, 110, 2349–2354. [Google Scholar] [CrossRef] [PubMed]
- Agrotis, A.; Kanellakis, P.; Kostolias, G.; Di Vitto, G.; Wei, C.; Hannan, R.; Jennings, G.; Bobik, A. Proliferation of Neointimal Smooth Muscle Cells after Arterial Injury: Dependence on Interactions Between Fibroblast Growth Factor Receptor-2 and Fibroblast Growth Factor-9. J. Biol. Chem. 2004, 279, 42221–42229. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.L.; Reidy, M.A. Basic Fibroblast Growth Factor: Its Role in the Control of Smooth Muscle Cell Migration. Am. J. Pathol. 1993, 143, 1024–1031. [Google Scholar]
- Fornai, F.; Carrizzo, A.; Forte, M.; Ambrosio, M.; Damato, A.; Ferrucci, M.; Biagioni, F.; Busceti, C.; Puca, A.A.; Vecchione, C. The Inflammatory Protein Pentraxin 3 in Cardiovascular Disease. Immun. Ageing 2016, 13, 25. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, M.; Yuksel, N.; Takmaz, T.; Dincel, A.S.; Bilgihan, A.; Altınkaynak, H. Aqueous Humor Pentraxin-3 Levels in Patients with Diabetes Mellitus. Eye 2017, 31, 1463–1467. [Google Scholar] [CrossRef] [Green Version]
- Akgul, O.; Baycan, O.F.; Bulut, U.; Somuncu, M.U.; Pusuroglu, H.; Ozyilmaz, S.; Gul, M.; Demir, A.R.; Yılmaz, E.; Yazan, S.; et al. Long-Term Prognostic Value of Elevated Pentraxin 3 in Patients Undergoing Primary Angioplasty for ST-Elevation Myocardial Infarction. Coron. Artery Dis. 2015, 26, 592–597. [Google Scholar] [CrossRef]
- Kimura, S.; Inagaki, H.; Haraguchi, G.; Sugiyama, T.; Miyazaki, T.; Hatano, Y.; Yoshikawa, S.; Ashikaga, T.; Isobe, M. Relationships of Elevated Systemic Pentraxin-3 Levels with High-Risk Coronary Plaque Components and Impaired Myocardial Perfusion After Percutaneous Coronary Intervention in Patients With ST-Elevation Acute Myocardial Infarction. Circ. J. 2014, 78, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Li, Y.; Wen, J.; Li, W.; Xu, Y. Elevated Plasma Level of Pentraxin-3 Predicts In-Hospital and 30-Day Clinical Outcomes in Patients with Non-ST-Segment Elevation Myocardial Infarction Who Have Undergone Percutaneous Coronary Intervention. CRD 2014, 129, 178–188. [Google Scholar] [CrossRef]
- Matsubara, J.; Sugiyama, S.; Nozaki, T.; Sugamura, K.; Konishi, M.; Ohba, K.; Matsuzawa, Y.; Akiyama, E.; Yamamoto, E.; Sakamoto, K.; et al. Pentraxin 3 Is a New Inflammatory Marker Correlated with Left Ventricular Diastolic Dysfunction and Heart Failure with Normal Ejection Fraction. J. Am. Coll. Cardiol. 2011, 57, 861–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubin, R.; Li, Y.; Ix, J.H.; Shlipak, M.G.; Whooley, M.; Peralta, C.A. Associations of Pentraxin-3 with Cardiovascular Events, Incident Heart Failure, and Mortality among Persons with Coronary Heart Disease: Data from the Heart and Soul Study. Am. Heart J. 2012, 163, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charla, E.; Mercer, J.; Maffia, P.; Nicklin, S.A. Extracellular Vesicle Signalling in Atherosclerosis. Cell Signal 2020, 75, 109751. [Google Scholar] [CrossRef] [PubMed]
- Yates, A.G.; Pink, R.C.; Erdbrügger, U.; Siljander, P.R.-M.; Dellar, E.R.; Pantazi, P.; Akbar, N.; Cooke, W.R.; Vatish, M.; Dias-Neto, E.; et al. In Sickness and in Health: The Functional Role of Extracellular Vesicles in Physiology and Pathology in Vivo: Part II: Pathology: Part II: Pathology. J. Extracell Vesicles 2022, 11, e12190. [Google Scholar] [CrossRef] [PubMed]
- Dorobantu, M.; Simionescu, M.; Popa-Fotea, N.-M. Molecular Research in Cardiovascular Disease. Int. J. Mol. Sci. 2021, 22, 7199. [Google Scholar] [CrossRef]
- Coly, P.-M.; Boulanger, C.M. Role of Extracellular Vesicles in Atherosclerosis: An Update. J. Leukoc. Biol. 2022, 111, 51–62. [Google Scholar] [CrossRef]
- Dignat-George, F.; Boulanger, C.M. The Many Faces of Endothelial Microparticles. ATVB 2011, 31, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Deng, W.; Tang, T.; Hou, Y.; Zeng, Q.; Wang, Y.; Fan, W.; Qu, S. Extracellular Vesicles in Atherosclerosis. Clin. Chim. Acta 2019, 495, 109–117. [Google Scholar] [CrossRef]
- Xu, H.; Ni, Y.-Q.; Liu, Y.-S. Mechanisms of Action of MiRNAs and LncRNAs in Extracellular Vesicle in Atherosclerosis. Front Cardiovasc. Med. 2021, 8, 733985. [Google Scholar] [CrossRef]
- Bobryshev, Y.V.; Killingsworth, M.C.; Orekhov, A.N. Increased Shedding of Microvesicles from Intimal Smooth Muscle Cells in Athero-Prone Areas of the Human Aorta: Implications for Understanding of the Predisease Stage. Pathobiology 2013, 80, 24–31. [Google Scholar] [CrossRef]
- Perrotta, I.; Aquila, S. Exosomes in Human Atherosclerosis: An Ultrastructural Analysis Study. Ultrastruct. Pathol. 2016, 40, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, R.B.; Chirinos, J.A.; Soriano, A.O.; Jy, W.; Horstman, L.; Jimenez, J.J.; Mendez, A.; Ferreira, A.; de Marchena, E.; Ahn, Y.S. Endothelial Microparticles and Platelet and Leukocyte Activation in Patients with the Metabolic Syndrome. Am. J. Cardiol. 2006, 98, 70–74. [Google Scholar] [CrossRef]
- Amabile, N.; Cheng, S.; Renard, J.M.; Larson, M.G.; Ghorbani, A.; McCabe, E.; Griffin, G.; Guerin, C.; Ho, J.E.; Shaw, S.Y.; et al. Association of Circulating Endothelial Microparticles with Cardiometabolic Risk Factors in the Framingham Heart Study. Eur. Heart J. 2014, 35, 2972–2979. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-J.; Liu, Y.; Chen, Y.; Yu, D.; Williams, K.J.; Liu, M.-L. Novel Proteolytic Microvesicles Released from Human Macrophages after Exposure to Tobacco Smoke. Am. J. Pathol. 2013, 182, 1552–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, A.C.; Peter, A.A.; Mendez, A.J.; Jimenez, J.J.; Mauro, L.M.; Chirinos, J.A.; Ghany, R.; Virani, S.; Garcia, S.; Horstman, L.L.; et al. Postprandial Hypertriglyceridemia Increases Circulating Levels of Endothelial Cell Microparticles. Circulation 2004, 110, 3599–3603. [Google Scholar] [CrossRef] [Green Version]
- Nomura, S.; Shouzu, A.; Omoto, S.; Nishikawa, M.; Iwasaka, T. Effects of Losartan and Simvastatin on Monocyte-Derived Microparticles in Hypertensive Patients with and without Type 2 Diabetes Mellitus. Clin. Appl. Thromb. Hemost. 2004, 10, 133–141. [Google Scholar] [CrossRef]
- Konkoth, A.; Saraswat, R.; Dubrou, C.; Sabatier, F.; Leroyer, A.S.; Lacroix, R.; Duchez, A.-C.; Dignat-George, F. Multifaceted Role of Extracellular Vesicles in Atherosclerosis. Atherosclerosis 2021, 319, 121–131. [Google Scholar] [CrossRef]
- de Freitas, R.C.C.; Hirata, R.D.C.; Hirata, M.H.; Aikawa, E. Circulating Extracellular Vesicles as Biomarkers and Drug Delivery Vehicles in Cardiovascular Diseases. Biomolecules 2021, 11, 388. [Google Scholar] [CrossRef]
- Nozaki, T.; Sugiyama, S.; Koga, H.; Sugamura, K.; Ohba, K.; Matsuzawa, Y.; Sumida, H.; Matsui, K.; Jinnouchi, H.; Ogawa, H. Significance of a Multiple Biomarkers Strategy Including Endothelial Dysfunction to Improve Risk Stratification for Cardiovascular Events in Patients at High Risk for Coronary Heart Disease. J. Am. Coll. Cardiol. 2009, 54, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Lässer, C.; Seyed Alikhani, V.; Ekström, K.; Eldh, M.; Torregrosa Paredes, P.; Bossios, A.; Sjöstrand, M.; Gabrielsson, S.; Lötvall, J.; Valadi, H. Human Saliva, Plasma and Breast Milk Exosomes Contain RNA: Uptake by Macrophages. J. Transl. Med. 2011, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Bernal-Mizrachi, L.; Jy, W.; Jimenez, J.J.; Pastor, J.; Mauro, L.M.; Horstman, L.L.; de Marchena, E.; Ahn, Y.S. High Levels of Circulating Endothelial Microparticles in Patients with Acute Coronary Syndromes. Am. Heart J. 2003, 145, 962–970. [Google Scholar] [CrossRef]
- Leroyer, A.S.; Isobe, H.; Lesèche, G.; Castier, Y.; Wassef, M.; Mallat, Z.; Binder, B.R.; Tedgui, A.; Boulanger, C.M. Cellular Origins and Thrombogenic Activity of Microparticles Isolated from Human Atherosclerotic Plaques. J. Am. Coll. Cardiol. 2007, 49, 772–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berezin, A.E.; Berezin, A.A. Extracellular Vesicles and Thrombogenicity in Atrial Fibrillation. Int. J. Mol. Sci. 2022, 23, 1774. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Bai, H.; Chen, X.; Gong, J.; Li, D.; Sun, Z. Anticoagulant Treatment Is Associated with Decreased Mortality in Severe Coronavirus Disease 2019 Patients with Coagulopathy. J. Thromb. Haemost. 2020, 18, 1094–1099. [Google Scholar] [CrossRef]
- Aharon, A.; Tamari, T.; Brenner, B. Monocyte-Derived Microparticles and Exosomes Induce Procoagulant and Apoptotic Effects on Endothelial Cells. Thromb. Haemost. 2008, 100, 878–885. [Google Scholar] [CrossRef]
- Wang, J.-G.; Williams, J.C.; Davis, B.K.; Jacobson, K.; Doerschuk, C.M.; Ting, J.P.-Y.; Mackman, N. Monocytic Microparticles Activate Endothelial Cells in an IL-1β–Dependent Manner. Blood 2011, 118, 2366–2374. [Google Scholar] [CrossRef] [Green Version]
- Saheera, S.; Jani, V.P.; Witwer, K.W.; Kutty, S. Extracellular Vesicle Interplay in Cardiovascular Pathophysiology. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1749–H1761. [Google Scholar] [CrossRef]
- Buffolo, F.; Monticone, S.; Camussi, G.; Aikawa, E. Role of Extracellular Vesicles in the Pathogenesis of Vascular Damage. Hypertension 2022, 79, 863–873. [Google Scholar] [CrossRef]
- Ramírez, R.; Ceprian, N.; Figuer, A.; Valera, G.; Bodega, G.; Alique, M.; Carracedo, J. Endothelial Senescence and the Chronic Vascular Diseases: Challenges and Therapeutic Opportunities in Atherosclerosis. J. Pers. Med. 2022, 12, 215. [Google Scholar] [CrossRef]
- Sun, X.; Feinberg, M.W. Vascular Endothelial Senescence: Pathobiological Insights, Emerging Long Noncoding RNA Targets, Challenges and Therapeutic Opportunities. Front. Physiol. 2021, 12, 693067. [Google Scholar] [CrossRef]
- Lenasi, H. Endothelial Dysfunction: Old Concepts and New Challenges; BoD–Books on Demand; IntechOpen: London, UK, 2018; ISBN 978-1-78984-253-1. [Google Scholar]
- Wu, C.-M.; Zheng, L.; Wang, Q.; Hu, Y.-W. The Emerging Role of Cell Senescence in Atherosclerosis. Clin. Chem. Lab. Med. (CCLM) 2021, 59, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Buendía, P.; Montes de Oca, A.; Madueño, J.A.; Merino, A.; Martín-Malo, A.; Aljama, P.; Ramírez, R.; Rodríguez, M.; Carracedo, J. Endothelial Microparticles Mediate Inflammation-Induced Vascular Calcification. FASEB J. 2015, 29, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular Smooth Muscle Cells in Atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef]
- Goetzl, E.J.; Schwartz, J.B.; Mustapic, M.; Lobach, I.V.; Daneman, R.; Abner, E.L.; Jicha, G.A. Altered Cargo Proteins of Human Plasma Endothelial Cell–Derived Exosomes in Atherosclerotic Cerebrovascular Disease. FASEB J. 2017, 31, 3689–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, A.; Wang, H.; Su, Y.; Zhang, C.; Qiu, Y.; Zhou, Y.; Wan, Y.; Hu, B.; Li, Y. Exosomes: Biomarkers and Therapeutic Targets of Diabetic Vascular Complications. Front. Endocrinol. 2021, 12, 720466. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, Q.; Liu, D.; Liu, Z. Exosomes: Advances, Development and Potential Therapeutic Strategies in Diabetic Nephropathy. Metabolism 2021, 122, 154834. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, J.; Qu, D.; Wang, L.; Wong, C.M.; Lau, C.-W.; Huang, Y.; Wang, Y.F.; Huang, H.; Xia, Y.; et al. Serum Exosomes Mediate Delivery of Arginase 1 as a Novel Mechanism for Endothelial Dysfunction in Diabetes. Proc. Natl. Acad. Sci. USA 2018, 115, E6927–E6936. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Song, X.; Yu, H.; Sun, J.; Zhao, Y. Therapeutic Potentials of Extracellular Vesicles for the Treatment of Diabetes and Diabetic Complications. Int. J. Mol. Sci. 2020, 21, 5163. [Google Scholar] [CrossRef]
- Heo, J.; Kang, H. Exosome-Based Treatment for Atherosclerosis. Int. J. Mol. Sci. 2022, 23, 1002. [Google Scholar] [CrossRef]
- Libby, P.; Tabas, I.; Fredman, G.; Fisher, E.A. Inflammation and Its Resolution as Determinants of Acute Coronary Syndromes. Circ. Res. 2014, 114, 1867–1879. [Google Scholar] [CrossRef] [Green Version]
- Schroder, K.; Tschopp, J. The Inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 Inflammasomes Are Required for Atherogenesis and Activated by Cholesterol Crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altaf, A.; Qu, P.; Zhao, Y.; Wang, H.; Lou, D.; Niu, N. NLRP3 Inflammasome in Peripheral Blood Monocytes of Acute Coronary Syndrome Patients and Its Relationship with Statins. Coron. Artery Dis. 2015, 26, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Xie, W.-L.; Kong, W.-W.; Chen, D.; Qu, P. Expression of the NLRP3 Inflammasome in Carotid Atherosclerosis. J. Stroke Cerebrovasc. Dis. 2015, 24, 2455–2466. [Google Scholar] [CrossRef]
- Xiao, H.; Lu, M.; Lin, T.Y.; Chen, Z.; Chen, G.; Wang, W.-C.; Marin, T.; Shentu, T.; Wen, L.; Gongol, B.; et al. Sterol Regulatory Element Binding Protein 2 Activation of NLRP3 Inflammasome in Endothelium Mediates Hemodynamic-Induced Atherosclerosis Susceptibility. Circulation 2013, 128, 632–642. [Google Scholar] [CrossRef]
- Zheng, F.; Xing, S.; Gong, Z.; Xing, Q. NLRP3 Inflammasomes Show High Expression in Aorta of Patients with Atherosclerosis. Heart Lung Circ. 2013, 22, 746–750. [Google Scholar] [CrossRef]
- Jin, Y.; Fu, J. Novel Insights into the NLRP3 Inflammasome in Atherosclerosis. J. Am. Heart Assoc. 2019, 8, e012219. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, J.; Evans, D.; King, A.; Dewberry, R.; Dower, S.; Crossman, D.; Francis, S. Interleukin-1β and Signaling of Interleukin-1 in Vascular Wall and Circulating Cells Modulates the Extent of Neointima Formation in Mice. Am. J. Pathol. 2006, 168, 1396–1403. [Google Scholar] [CrossRef] [Green Version]
- Menu, P.; Pellegrin, M.; Aubert, J.-F.; Bouzourene, K.; Tardivel, A.; Mazzolai, L.; Tschopp, J. Atherosclerosis in ApoE-Deficient Mice Progresses Independently of the NLRP3 Inflammasome. Cell Death Dis. 2011, 2, e137. [Google Scholar] [CrossRef] [Green Version]
- Kirii, H.; Niwa, T.; Yamada, Y.; Wada, H.; Saito, K.; Iwakura, Y.; Asano, M.; Moriwaki, H.; Seishima, M. Lack of Interleukin-1β Decreases the Severity of Atherosclerosis in ApoE-Deficient Mice. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 656–660. [Google Scholar] [CrossRef] [Green Version]
- Parathath, S.; Mick, S.L.; Feig, J.E.; Joaquin, V.; Grauer, L.; Habiel, D.M.; Gassmann, M.; Gardner, L.B.; Fisher, E.A. Hypoxia Is Present in Murine Atherosclerotic Plaques and Has Multiple Adverse Effects on Macrophage Lipid Metabolism. Circ. Res. 2011, 109, 1141–1152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folco, E.J.; Sheikine, Y.; Rocha, V.Z.; Christen, T.; Shvartz, E.; Sukhova, G.K.; Di Carli, M.F.; Libby, P. Hypoxia but Not Inflammation Augments Glucose Uptake in Human Macrophages: Implications for Imaging Atherosclerosis with 18Fluorine-Labeled 2-Deoxy-D-Glucose Positron Emission Tomography. J. Am. Coll. Cardiol. 2011, 58, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folco, E.J.; Sukhova, G.K.; Quillard, T.; Libby, P. Moderate Hypoxia Potentiates Interleukin-1β Production in Activated Human Macrophages. Circ. Res. 2014, 115, 875–883. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Xia, M.; Gulbins, E.; Boini, K.M.; Li, P.-L. Activation of Nlrp3 Inflammasomes Enhances Macrophage Lipid-Deposition and Migration: Implication of a Novel Role of Inflammasome in Atherogenesis. PLoS ONE 2014, 9, e87552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host Cell Death and Inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Grebe, A.; Hoss, F.; Latz, E. NLRP3 Inflammasome and the IL-1 Pathway in Atherosclerosis. Circ. Res. 2018, 122, 1722–1740. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Rane, M. Interleukin-6 Signaling and Anti-Interleukin-6 Therapeutics in Cardiovascular Disease. Circ. Res. 2021, 128, 1728–1746. [Google Scholar] [CrossRef]
- Abderrazak, A.; Couchie, D.; Mahmood, D.F.D.; Elhage, R.; Vindis, C.; Laffargue, M.; Matéo, V.; Büchele, B.; Ayala, M.R.; El Gaafary, M.; et al. Anti-Inflammatory and Antiatherogenic Effects of the NLRP3 Inflammasome Inhibitor Arglabin in ApoE2.Ki Mice Fed a High-Fat Diet. Circulation 2015, 131, 1061–1070. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Long, Y.; Chen, Y. Roles of Inflammasome in Cigarette Smoke-Related Diseases and Physiopathological Disorders: Mechanisms and Therapeutic Opportunities. Front. Immunol. 2021, 12, 720049. [Google Scholar] [CrossRef]
- Mehta, S.; Dhawan, V. Molecular Insights of Cigarette Smoke Condensate-Activated NLRP3 Inflammasome in THP-1 Cells in a Stage-Specific Atherogenesis. Int. Immunopharmacol. 2020, 88, 107013. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Mao, J.; Xu, S.; Zhao, L.; Long, L.; Chen, L.; Li, D.; Lu, S. Rosmarinic Acid Inhibits Nicotine-Induced C-Reactive Protein Generation by Inhibiting NLRP3 Inflammasome Activation in Smooth Muscle Cells. J. Cell Physiol. 2019, 234, 1758–1767. [Google Scholar] [CrossRef] [PubMed]
- Ozkok, A. Cholesterol-Embolization Syndrome: Current Perspectives. Vasc. Health Risk Manag. 2019, 15, 209–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saric, M.; Kronzon, I. Cholesterol Embolization Syndrome. Curr. Opin. Cardiol. 2011, 26, 472–479. [Google Scholar] [CrossRef]
- Satish, M.; Agrawal, D.K. Atherothrombosis and the NLRP3 Inflammasome—Endogenous Mechanisms of Inhibition. Transl. Res. 2020, 215, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.; Tsuruda, T.; Kita, T.; Kitamura, K.; Eto, T. Adrenomedullin. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 2480–2487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgenthaler, N.G.; Struck, J.; Alonso, C.; Bergmann, A. Measurement of Midregional Proadrenomedullin in Plasma with an Immunoluminometric Assay. Clin. Chem. 2005, 51, 1823–1829. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.Q.; O’Brien, R.J.; Struck, J.; Quinn, P.; Morgenthaler, N.; Squire, I.; Davies, J.; Bergmann, A.; Ng, L.L. Prognostic Value of Midregional Pro-Adrenomedullin in Patients with Acute Myocardial Infarction: The LAMP (Leicester Acute Myocardial Infarction Peptide) Study. J. Am. Coll. Cardiol. 2007, 49, 1525–1532. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Kuriyama, N.; Suzuki, Y.; Saito, S.; Tanaka, R.; Iwao, M.; Tanaka, M.; Maki, T.; Itoh, H.; Ihara, M.; et al. Mid-Regional pro-Adrenomedullin Is a Novel Biomarker for Arterial Stiffness as the Criterion for Vascular Failure in a Cross-Sectional Study. Sci. Rep. 2021, 11, 305. [Google Scholar] [CrossRef]
- Melander, O.; Newton-Cheh, C.; Almgren, P.; Hedblad, B.; Berglund, G.; Engström, G.; Persson, M.; Smith, J.G.; Magnusson, M.; Christensson, A.; et al. Novel and Conventional Biomarkers for the Prediction of Incident Cardiovascular Events in the Community. JAMA 2009, 302, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Roos, M.; Schuster, T.; Ndrepepa, G.; Baumann, M.; Lutz, J.; Braun, S.; Martinof, S.; Schömig, A.; Heemann, U.; Kastrati, A.; et al. Association of Midregional Proadrenomedullin with Coronary Artery Stenoses, Soft Atherosclerotic Plaques and Coronary Artery Calcium. Heart Vessel. 2012, 27, 352–359. [Google Scholar] [CrossRef]
- Yoshihara, F.; Ernst, A.; Morgenthaler, N.G.; Horio, T.; Nakamura, S.; Nakahama, H.; Nakata, H.; Bergmann, A.; Kangawa, K.; Kawano, Y. Midregional Proadrenomedullin Reflects Cardiac Dysfunction in Haemodialysis Patients with Cardiovascular Disease. Nephrol. Dial. Transplant. 2007, 22, 2263–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brouwers, F.P.; de Boer, R.A.; van der Harst, P.; Struck, J.; de Jong, P.E.; de Zeeuw, D.; Gans, R.O.; Gansevoort, R.T.; Hillege, H.L.; van Gilst, W.H.; et al. Influence of Age on the Prognostic Value of Mid-Regional pro-Adrenomedullin in the General Population. Heart 2012, 98, 1348–1353. [Google Scholar] [CrossRef] [PubMed]
- Neumann, J.T.; Tzikas, S.; Funke-Kaiser, A.; Wilde, S.; Appelbaum, S.; Keller, T.; Ojeda-Echevarria, F.; Zeller, T.; Zwiener, I.; Sinning, C.R.; et al. Association of MR-Proadrenomedullin with Cardiovascular Risk Factors and Subclinical Cardiovascular Disease. Atherosclerosis 2013, 228, 451–459. [Google Scholar] [CrossRef]
- Sawada, S. Cystatin C as a Promising Biomarker of Atherosclerotic Plaque. J. Atheroscler. Thromb. 2021, 28, 474–476. [Google Scholar] [CrossRef]
- Shi, G.P.; Sukhova, G.K.; Grubb, A.; Ducharme, A.; Rhode, L.H.; Lee, R.T.; Ridker, P.M.; Libby, P.; Chapman, H.A. Cystatin C Deficiency in Human Atherosclerosis and Aortic Aneurysms. J. Clin. Investig. 1999, 104, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Honda, K.; Yuzaki, M.; Tajima, K.; Nakamura, R.; Nakanishi, Y.; Kaneko, M.; Agematsu, K.; Nagashima, M. Serum Cystatin C Level as a Biomarker of Aortic Plaque in Patients with an Aortic Arch Aneurysm. J. Atheroscler. Thromb. 2021, 28, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, H.-P.; Wang, Y.-C.; Ren, T.-T.; Li, J.; Xu, M.-L.; Wang, X.-Q.; Liu, F.-C.; Lau, A.; Wen, Y.-F. Serum Cystatin C Level Is Associated with Carotid Intima-Media Thickening and Plaque. Scand. J. Clin. Lab. Investig. 2015, 75, 265–272. [Google Scholar] [CrossRef]
- Correa, S.; Morrow, D.A.; Braunwald, E.; Davies, R.Y.; Goodrich, E.L.; Murphy, S.A.; Cannon, C.P.; O’Donoghue, M.L. Cystatin C for Risk Stratification in Patients After an Acute Coronary Syndrome. JAHA 2018, 7, e009077. [Google Scholar] [CrossRef] [Green Version]
- Colley, K.J.; Wolfert, R.L.; Cobble, M.E. Lipoprotein Associated Phospholipase A2: Role in Atherosclerosis and Utility as a Biomarker for Cardiovascular Risk. EPMA J. 2011, 2, 27–38. [Google Scholar] [CrossRef] [Green Version]
- Ballantyne, C.M.; Hoogeveen, R.C.; Bang, H.; Coresh, J.; Folsom, A.R.; Chambless, L.E.; Myerson, M.; Wu, K.K.; Sharrett, A.R.; Boerwinkle, E. Lipoprotein-Associated Phospholipase A2, High-Sensitivity C-Reactive Protein, and Risk for Incident Ischemic Stroke in Middle-Aged Men and Women in the Atherosclerosis Risk in Communities (ARIC) Study. Arch. Intern. Med. 2005, 165, 2479–2484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballantyne, C.M.; Hoogeveen, R.C.; Bang, H.; Coresh, J.; Folsom, A.R.; Heiss, G.; Sharrett, A.R. Lipoprotein-Associated Phospholipase A2, High-Sensitivity C-Reactive Protein, and Risk for Incident Coronary Heart Disease in Middle-Aged Men and Women in the Atherosclerosis Risk in Communities (ARIC) Study. Circulation 2004, 109, 837–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, A.; Zheng, D.; Qiu, R.; Mai, W.; Zhou, Y. Lipoprotein-Associated Phospholipase A2 (Lp-PLA_2): A Novel and Promising Biomarker for Cardiovascular Risks Assessment. Dis. Markers 2013, 34, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Papapanagiotou, A.; Siasos, G.; Kassi, E.; Gargalionis, A.N.; Papavassiliou, A.G. Novel Inflammatory Markers in Hyperlipidemia: Clinical Implications. Curr. Med. Chem. 2015, 22, 2727–2743. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D. La Lp-PLA2, marqueur d’inflammation vasculaire et de vulnérabilité de la plaque d’athérosclérose. Ann. Pharm. Françaises 2016, 74, 190–197. [Google Scholar] [CrossRef]
- Hassan, M. STABILITY and SOLID-TIMI 52: Lipoprotein Associated Phospholipase A2 (Lp-PLA2) as a Biomarker or Risk Factor for Cardiovascular Diseases. Glob. Cardiol. Sci. Pract. 2015, 2015, 6. [Google Scholar] [CrossRef]
- Maiolino, G.; Bisogni, V.; Rossitto, G.; Rossi, G.P. Lipoprotein-Associated Phospholipase A2 Prognostic Role in Atherosclerotic Complications. World. J. Cardiol. 2015, 7, 609–620. [Google Scholar] [CrossRef]
- Kolodgie, F.D.; Burke, A.P.; Skorija, K.S.; Ladich, E.; Kutys, R.; Makuria, A.T.; Virmani, R. Lipoprotein-Associated Phospholipase A2 Protein Expression in the Natural Progression of Human Coronary Atherosclerosis. Arter. Thromb. Vasc. Biol. 2006, 26, 2523–2529. [Google Scholar] [CrossRef] [Green Version]
- Mannheim, D.; Herrmann, J.; Versari, D.; Gössl, M.; Meyer, F.B.; McConnell, J.P.; Lerman, L.O.; Lerman, A. Enhanced Expression of Lp-PLA2 and Lysophosphatidylcholine in Symptomatic Carotid Atherosclerotic Plaques. Stroke 2008, 39, 1448–1455. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Maguire, C.T.; Wolfert, R.; Burns, A.R.; Reardon, M.; Geis, R.; Holvoet, P.; Morrisett, J.D. Relationship of Lipoprotein-Associated Phospholipase A2 and Oxidized Low Density Lipoprotein in Carotid Atherosclerosis. J. Lipid Res. 2009, 50, 1735–1743. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Gao, Y.; Wu, D.; Zhang, D. The Relationship of Lipoprotein-Associated Phospholipase A2 Activity with the Seriousness of Coronary Artery Disease. BMC Cardiovasc. Disord. 2020, 20, 295. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Visse, R.; Murphy, G. Structure and Function of Matrix Metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, J.A.; Orbe, J.; Martinez de Lizarrondo, S.; Calvayrac, O.; Rodriguez, C.; Martinez-Gonzalez, J.; Paramo, J.A. Metalloproteinases and Atherothrombosis: MMP-10 Mediates Vascular Remodeling Promoted by Inflammatory Stimuli. Front. Biosci. 2008, 13, 2916–2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myasoedova, V.A.; Chistiakov, D.A.; Grechko, A.V.; Orekhov, A.N. Matrix Metalloproteinases in Pro-Atherosclerotic Arterial Remodeling. J. Mol. Cell. Cardiol. 2018, 123, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Olejarz, W.; Łacheta, D.; Kubiak-Tomaszewska, G. Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability. Int. J. Mol. Sci. 2020, 21, 3946. [Google Scholar] [CrossRef]
- MacColl, E.; Khalil, R.A. Matrix Metalloproteinases as Regulators of Vein Structure and Function: Implications in Chronic Venous Disease. J. Pharm. Exp. 2015, 355, 410–428. [Google Scholar] [CrossRef] [Green Version]
- Purroy, A.; Roncal, C.; Orbe, J.; Meilhac, O.; Belzunce, M.; Zalba, G.; Villa-Bellosta, R.; Andrés, V.; Parks, W.C.; Páramo, J.A.; et al. Matrix Metalloproteinase-10 Deficiency Delays Atherosclerosis Progression and Plaque Calcification. Atherosclerosis 2018, 278, 124–134. [Google Scholar] [CrossRef]
- Matilla, L.; Roncal, C.; Ibarrola, J.; Arrieta, V.; García-Peña, A.; Fernández-Celis, A.; Navarro, A.; Álvarez, V.; Gainza, A.; Orbe, J.; et al. A Role for MMP-10 (Matrix Metalloproteinase-10) in Calcific Aortic Valve Stenosis. ATVB 2020, 40, 1370–1382. [Google Scholar] [CrossRef]
- Mohler, E.R.; Gannon, F.; Reynolds, C.; Zimmerman, R.; Keane, M.G.; Kaplan, F.S. Bone Formation and Inflammation in Cardiac Valves. Circulation 2001, 103, 1522–1528. [Google Scholar] [CrossRef]
- Bossé, Y.; Miqdad, A.; Fournier, D.; Pépin, A.; Pibarot, P.; Mathieu, P. Refining Molecular Pathways Leading to Calcific Aortic Valve Stenosis by Studying Gene Expression Profile of Normal and Calcified Stenotic Human Aortic Valves. Circ. Cardiovasc. Genet. 2009, 2, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Jung, J.-J.; Razavian, M.; Challa, A.A.; Nie, L.; Golestani, R.; Zhang, J.; Ye, Y.; Russell, K.S.; Robinson, S.P.; Heistad, D.D.; et al. Multimodality and Molecular Imaging of Matrix Metalloproteinase Activation in Calcific Aortic Valve Disease. J. Nucl. Med. 2015, 56, 933–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Zhang, J.-W.; Zang, G.-Y.; Pu, J. The Primary Use of Artificial Intelligence in Cardiovascular Diseases: What Kind of Potential Role Does Artificial Intelligence Play in Future Medicine? J. Geriatr. Cardiol. 2019, 16, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Turing, A.M. Computing Machinery and Intelligence. In Parsing the Turing Test: Philosophical and Methodological Issues in the Quest for the Thinking Computer; Epstein, R., Roberts, G., Beber, G., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 23–65. ISBN 978-1-4020-6710-5. [Google Scholar]
- Ghahramani, Z. Probabilistic Machine Learning and Artificial Intelligence. Nature 2015, 521, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Krittanawong, C.; Zhang, H.; Wang, Z.; Aydar, M.; Kitai, T. Artificial Intelligence in Precision Cardiovascular Medicine. J. Am. Coll. Cardiol. 2017, 69, 2657–2664. [Google Scholar] [CrossRef] [PubMed]
- Munger, E.; Hickey, J.W.; Dey, A.K.; Jafri, M.S.; Kinser, J.M.; Mehta, N.N. Application of Machine Learning in Understanding Atherosclerosis: Emerging Insights. APL Bioeng. 2021, 5, 011505. [Google Scholar] [CrossRef]
- Huynh-Thu, V.A.; Saeys, Y.; Wehenkel, L.; Geurts, P. Statistical Interpretation of Machine Learning-Based Feature Importance Scores for Biomarker Discovery. Bioinformatics 2012, 28, 1766–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, P.K.; Sharma, N.; Saba, L.; Paraskevas, K.I.; Kalra, M.K.; Johri, A.; Laird, J.R.; Nicolaides, A.N.; Suri, J.S. Unseen Artificial Intelligence-Deep Learning Paradigm for Segmentation of Low Atherosclerotic Plaque in Carotid Ultrasound: A Multicenter Cardiovascular Study. Diagnostics 2021, 11, 2257. [Google Scholar] [CrossRef]
- Hampe, N.; Wolterink, J.M.; van Velzen, S.G.M.; Leiner, T.; Išgum, I. Machine Learning for Assessment of Coronary Artery Disease in Cardiac CT: A Survey. Front. Cardiovasc. Med. 2019, 6, 172. [Google Scholar] [CrossRef] [Green Version]
- Boi, A.; Jamthikar, A.D.; Saba, L.; Gupta, D.; Sharma, A.; Loi, B.; Laird, J.R.; Khanna, N.N.; Suri, J.S. A Survey on Coronary Atherosclerotic Plaque Tissue Characterization in Intravascular Optical Coherence Tomography. Curr. Atheroscler. Rep. 2018, 20, 33. [Google Scholar] [CrossRef]
- Munger, E.; Choi, H.; Dey, A.K.; Elnabawi, Y.A.; Groenendyk, J.W.; Rodante, J.; Keel, A.; Aksentijevich, M.; Reddy, A.S.; Khalil, N.; et al. Application of Machine Learning to Determine Top Predictors of Noncalcified Coronary Burden in Psoriasis: An Observational Cohort Study. J. Am. Acad. Derm. 2020, 83, 1647–1653. [Google Scholar] [CrossRef]
- Forné, C.; Cambray, S.; Bermudez-Lopez, M.; Fernandez, E.; Bozic, M.; Valdivielso, J.M. The NEFRONA investigators Machine Learning Analysis of Serum Biomarkers for Cardiovascular Risk Assessment in Chronic Kidney Disease. Clin. Kidney J. 2020, 13, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.G.; Shah, N.H.; Dalman, R.L.; Nead, K.T.; Cooke, J.P.; Leeper, N.J. The Use of Machine Learning for the Identification of Peripheral Artery Disease and Future Mortality Risk. J. Vasc. Surg. 2016, 64, 1515–1522.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, A.M.; Demsas, F.; Leeper, N.J.; Ross, E.G. Leveraging Machine Learning and Artificial Intelligence to Improve Peripheral Artery Disease Detection, Treatment, and Outcomes. Circ. Res. 2021, 128, 1833–1850. [Google Scholar] [CrossRef]
- Qian, Y.; Zhang, L.; Sun, Z.; Zang, G.; Li, Y.; Wang, Z.; Li, L. Biomarkers of Blood from Patients with Atherosclerosis Based on Bioinformatics Analysis. Evol. Bioinform. Online 2021, 17, 11769343211046020. [Google Scholar] [CrossRef] [PubMed]
- Correia, M.; Kagenaar, E.; van Schalkwijk, D.B.; Bourbon, M.; Gama-Carvalho, M. Machine Learning Modelling of Blood Lipid Biomarkers in Familial Hypercholesterolaemia versus Polygenic/Environmental Dyslipidaemia. Sci. Rep. 2021, 11, 3801. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adam, C.A.; Șalaru, D.L.; Prisacariu, C.; Marcu, D.T.M.; Sascău, R.A.; Stătescu, C. Novel Biomarkers of Atherosclerotic Vascular Disease—Latest Insights in the Research Field. Int. J. Mol. Sci. 2022, 23, 4998. https://doi.org/10.3390/ijms23094998
Adam CA, Șalaru DL, Prisacariu C, Marcu DTM, Sascău RA, Stătescu C. Novel Biomarkers of Atherosclerotic Vascular Disease—Latest Insights in the Research Field. International Journal of Molecular Sciences. 2022; 23(9):4998. https://doi.org/10.3390/ijms23094998
Chicago/Turabian StyleAdam, Cristina Andreea, Delia Lidia Șalaru, Cristina Prisacariu, Dragoș Traian Marius Marcu, Radu Andy Sascău, and Cristian Stătescu. 2022. "Novel Biomarkers of Atherosclerotic Vascular Disease—Latest Insights in the Research Field" International Journal of Molecular Sciences 23, no. 9: 4998. https://doi.org/10.3390/ijms23094998