
Adsorption of Biomineralization Protein Mms6 on Magnetite (Fe3O4) Nanoparticles

 and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Adsorption of Proteins on Magnetite Nanoparticles
2.2. Adsorption Affinity of Mms6 to Magnetite Nanoparticle
2.3. Electron Microscopic Observation of Protein-Nanoparticle Conjugates
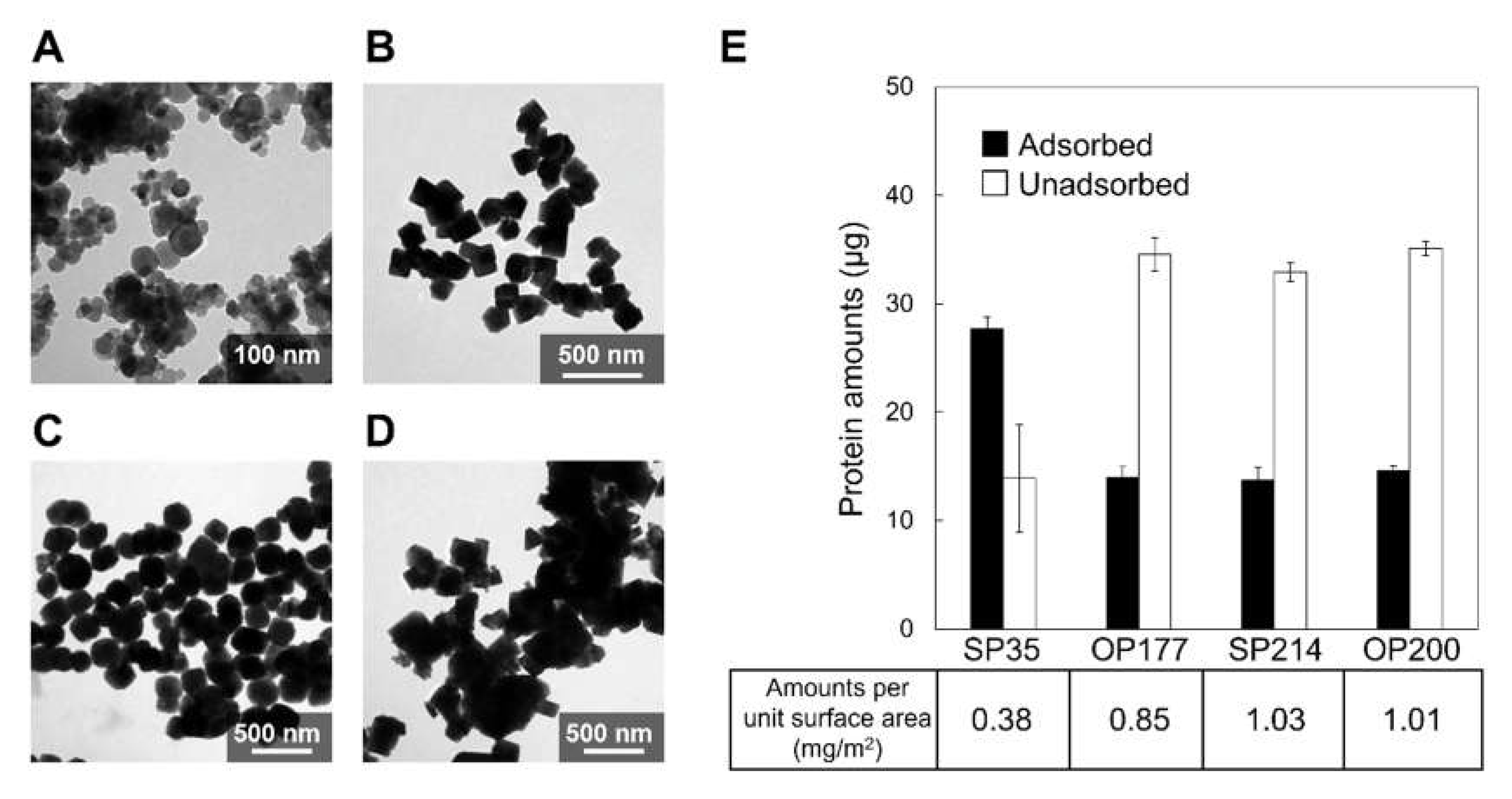
2.4. Effect of Crystal Faces on Adsorption of Mms6 on Magnetite Nanoparticle
2.5. Role of Adsorption in the Magnetite Biomineralization by Magnetotactic Bacteria
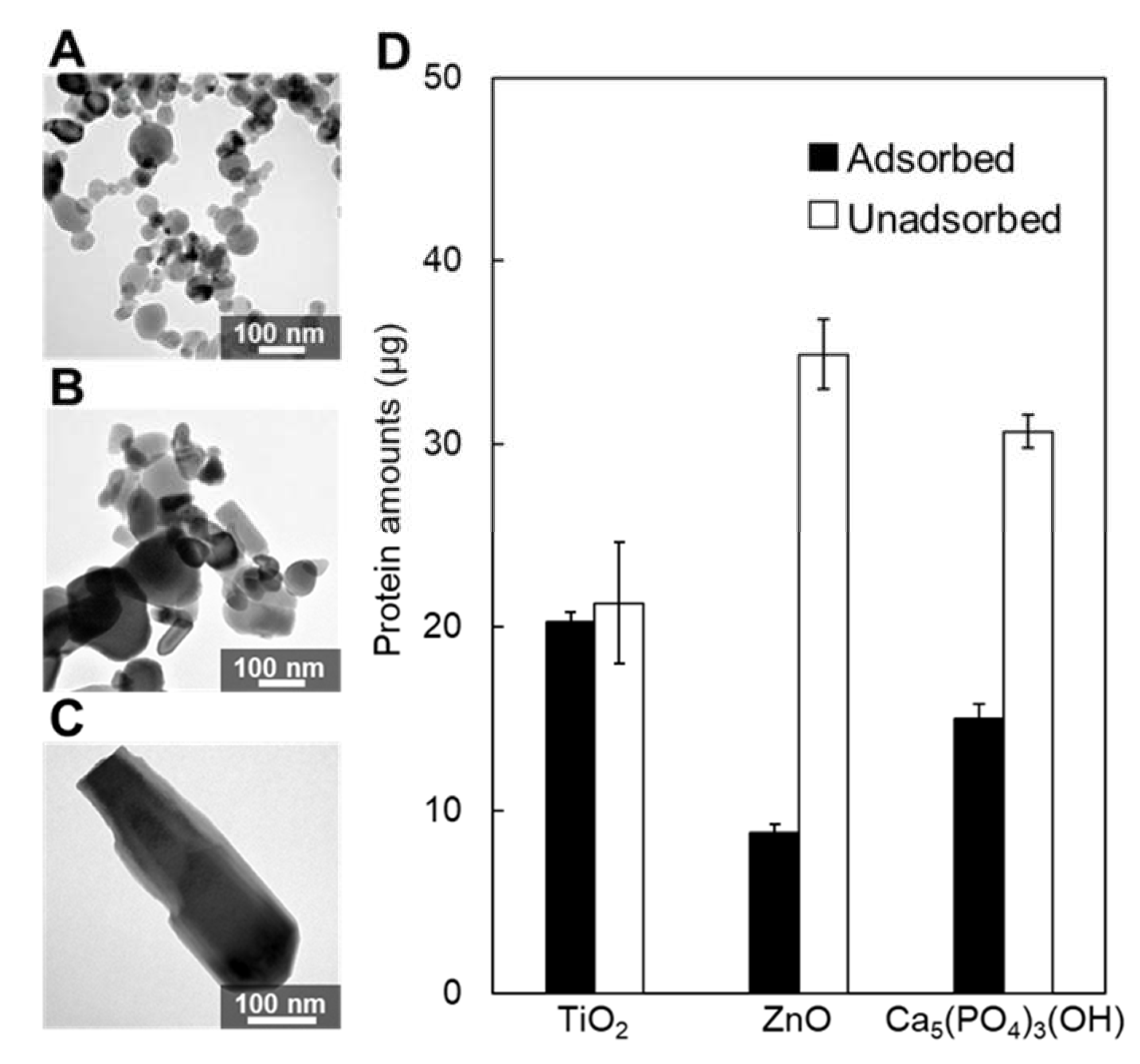
2.6. Adsorption on Inorganic Oxide Particles
3. Materials and Methods
3.1. Materials
3.2. Purification of Recombinant Proteins Expressed in Escherichia coli
3.3. In Vitro Protein Adsorption Assay Using Metal Oxide Nanoparticles
3.3.1. Preparation of Protein Solutions
3.3.2. Protein Adsorption on Metal Oxide Nanoparticles
3.3.3. Protein Desorption from Metal Oxide Nanoparticles
3.4. TEM Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mann, S.; Hannington, J.P.; Williams, R. Phospholipid vesicles as a model system for biomineralization. Nature 1986, 324, 565–567. [Google Scholar] [CrossRef] [PubMed]
- Aizenberg, J.; Black, A.J.; Whitesides, G.M. Control of crystal nucleation by patterned self-assembled monolayers. Nature 1999, 398, 495–498. [Google Scholar] [CrossRef]
- Weaver, J.C.; Wang, Q.; Miserez, A.; Tantuccio, A.; Stromberg, R.; Bozhilov, K.N.; Maxwell, P.; Nay, R.; Heier, S.T.; DiMasi, E.; et al. Analysis of an ultra hard magnetic biomineral in chiton radular teeth. Mater. Today 2010, 13, 42–52. [Google Scholar] [CrossRef]
- Weaver, J.C.; Milliron, G.W.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E.; et al. The stomatopod dactyl club: A formidable damage-tolerant biological hammer. Science 2012, 336, 1275–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemoto, M.; Wang, Q.; Li, D.; Pan, S.; Matsunaga, T.; Kisailus, D. Proteomic analysis from the mineralized radular teeth of the giant Pacific chiton, Cryptochiton stelleri (Mollusca). Proteomics 2012, 12, 2890–2894. [Google Scholar] [CrossRef]
- Wang, Q.; Nemoto, M.; Li, D.; Weaver, J.C.; Weden, B.; Stegemeier, J.; Bozhilov, K.N.; Wood, L.R.; Milliron, G.W.; Kim, C.S.; et al. Phase transformations and structural developments in the radular teeth of Cryptochiton stelleri. Adv. Funct. Mater. 2013, 23, 2908–2917. [Google Scholar] [CrossRef]
- Kisailus, D.; Nemoto, M. Structural and proteomic analyses of iron oxide biomineralization in chiton teeth. In Biological Magnetic Materials and Applications; Matsunaga, T., Tanaka, T., Kisailus, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; pp. 53–73. [Google Scholar]
- Nemoto, M.; Ren, D.; Herrera, S.; Pan, S.; Tamura, T.; Inagaki, K.; Kisailus, D. Integrated transcriptomic and proteomic analyses of a molecular mechanism of radular teeth biomineralization in Cryptochiton stelleri. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Shishehbor, M.; Guarín-Zapata, N.; Kirchhofer, N.D.; Li, J.; Cruz, L.; Wang, T.; Bhowmick, S.; Stauffer, D.; Manimunda, P.; et al. A natural impact-resistant bicontinuous composite nanoparticle coating. Nat. Mater. 2020, 19, 1236–1243. [Google Scholar] [CrossRef]
- Wang, T.; Huang, W.; Pham, C.H.; Murata, S.; Herrera, S.; Kirchhofer, N.D.; Arkook, B.; Stekovic, D.; Itkis, M.E.; Goldman, N.; et al. Mesocrystalline ordering and phase transformation of iron oxide biominerals in the ultrahard teeth of Cryptochiton stelleri. Small Struct. 2022, 3, 2100202. [Google Scholar] [CrossRef]
- Hoang, Q.Q.; Sicheri, F.; Howard, A.J.; Yang, D.S.C. Bone recognition mechanism of porcine osteocalcin from crystal structure. Nature 2003, 425, 977–980. [Google Scholar] [CrossRef]
- He, G.; Dahl, T.; Veis, A.; George, A. Nucleation of apatite crystals in vitro by self-assembled dentin matrix protein 1. Nat. Mater. 2003, 2, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayanan, R.; Kini, R.M.; Valiyaveettil, S. Investigation of the role of ansocalcin in the biomineralization in goose eggshell matrix. Proc. Natl. Acad. Sci. USA 2002, 99, 5155–5159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brutchey, R.L.; Yoo, E.S.; Morse, D.E. Biocatalytic synthesis of a nanostructured and crystalline bimetallic perovskite-like barium oxofluorotitanate at low temperature. J. Am. Chem. Soc. 2006, 128, 10288–10294. [Google Scholar] [CrossRef] [PubMed]
- Sumerel, J.L.; Yang, W.; Kisailus, D.; Weaver, J.C.; Choi, J.H.; Morse, D.E. Biocatalytically templated synthesis of titanium dioxide. Chem. Mater. 2003, 15, 4804–4809. [Google Scholar] [CrossRef]
- Kisailus, D.; Choi, J.H.; Weaver, J.C.; Yang, W.; Morse, D.E. Enzymatic synthesis and nanostructural control of gallium oxide at low temperature. Adv. Mater. 2005, 17, 314–318. [Google Scholar] [CrossRef]
- Curnow, P.; Bessette, P.H.; Kisailus, D.; Murr, M.M.; Daugherty, P.S.; Morse, D.E. Enzymatic synthesis of layered titanium phosphates at low temperature and neutral pH by cell-surface display of silicatein-α. J. Am. Chem. Soc. 2005, 127, 15749–15755. [Google Scholar] [CrossRef]
- Wang, K.; Wang, X.; Li, H.; Zheng, S.; Ren, Q.; Wang, Y.; Niu, Y.; Li, W.; Zhou, X.; Zhang, L. A statherin-derived peptide promotes hydroxyapatite crystallization and in situ remineralization of artificial enamel caries. RSC Adv. 2018, 8, 1647–1655. [Google Scholar] [CrossRef] [Green Version]
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef]
- Sakaguchi, T.; Burgess, J.G.; Matsunaga, T. Magnetite formation by a sulphate-reducing bacterium. Nature 1993, 365, 47–49. [Google Scholar] [CrossRef]
- Islam, T.; Peng, C.; Ali, I. Morphological and cellular diversity of magnetotactic bacteria: A review. J. Basic Microbiol. 2018, 58, 378–389. [Google Scholar] [CrossRef]
- Ozawa, E. Microwave-assisted magnetization reversal in dispersed nanosized barium ferrite particles for high-density magnetic recording tape. IEEE Trans. Magn. 2019, 55, 1–4. [Google Scholar] [CrossRef]
- Zhang, H.-w.; Liu, Y.; Sun, S.-h. Synthesis and assembly of magnetic nanoparticles for information and energy storage applications. Front. Phys. China 2010, 5, 347–356. [Google Scholar] [CrossRef]
- Matsunaga, T.; Sakaguchi, T.; Tadakoro, F. Magnetite formation by a magnetic bacterium capable of growing aerobically. Appl. Microbiol. Biotechnol. 1991, 35, 651–655. [Google Scholar] [CrossRef]
- Arakaki, A.; Yamagishi, A.; Fukuyo, A.; Tanaka, M.; Matsunaga, T. Co-ordinated functions of Mms proteins define the surface structure of cubo-octahedral magnetite crystals in magnetotactic bacteria. Mol. Microbiol. 2014, 93, 554–567. [Google Scholar] [CrossRef]
- Arakaki, A.; Webb, J.; Matsunaga, T. A novel protein tightly bound to bacterial magnetic particles in Magnetospirillum magneticum strain AMB-1. J. Biol. Chem. 2003, 278, 8745–8750. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Mazuyama, E.; Arakaki, A.; Matsunaga, T. Mms6 protein regulates crystal morphology during nano-sized magnetite biomineralization in vivo. J. Biol. Chem. 2011, 286, 6386–6392. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, A.E.; Bramble, J.P.; Hounslow, A.M.; Williamson, M.P.; Monnington, A.E.; Cooke, D.J.; Staniland, S.S. Ferrous iron binding key to Mms6 magnetite biomineralisation: A mechanistic study to understand magnetite formation using pH titration and NMR spectroscopy. Chem. Eur. J. 2016, 22, 7885. [Google Scholar] [CrossRef] [Green Version]
- Feng, S.; Wang, L.; Palo, P.; Liu, X.; Mallapragada, S.K.; Nilsen-Hamilton, M. Integrated self-assembly of the Mms6 magnetosome protein to form an iron-responsive structure. Int. J. Mol. Sci. 2013, 14, 14594–14606. [Google Scholar] [CrossRef]
- Wang, L.; Prozorov, T.; Palo, P.E.; Liu, X.; Vaknin, D.; Prozorov, R.; Mallapragada, S.; Nilsen-Hamilton, M. Self-assembly and biphasic iron-binding characteristics of Mms6, a bacterial protein that promotes the formation of superparamagnetic magnetite nanoparticles of uniform size and shape. Biomacromolecules 2012, 13, 98–105. [Google Scholar] [CrossRef]
- Prozorov, T.; Mallapragada, S.K.; Narasimhan, B.; Wang, L.; Palo, P.; Nilsen-Hamilton, M.; Williams, T.J.; Bazylinski, D.A.; Prozorov, R.; Canfield, P.C. Protein-mediated synthesis of uniform superparamagnetic magnetite nanocrystals. Adv. Funct. Mater. 2007, 17, 951–957. [Google Scholar] [CrossRef]
- Amemiya, Y.; Arakaki, A.; Staniland, S.S.; Tanaka, T.; Matsunaga, T. Controlled formation of magnetite crystal by partial oxidation of ferrous hydroxide in the presence of recombinant magnetotactic bacterial protein Mms6. Biomater. 2007, 28, 5381–5389. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.; Masuda, F.; Amemiya, Y.; Tanaka, T.; Matsunaga, T. Control of the morphology and size of magnetite particles with peptides mimicking the Mms6 protein from magnetotactic bacteria. J. Colloid Interface Sci. 2010, 343, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.M.; Rawlings, A.E.; Galloway, J.M.; Staniland, S.S. Using a biomimetic membrane surface experiment to investigate the activity of the magnetite biomineralisation protein Mms6. RSC Adv. 2016, 6, 7356–7363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Källberg, M.; Wang, H.; Wang, S.; Peng, J.; Wang, Z.; Lu, H.; Xu, J. Template-based protein structure modeling using the RaptorX web server. Nat. Protoc. 2012, 7, 1511–1522. [Google Scholar] [CrossRef] [Green Version]
- Pyles, H.; Zhang, S.; De Yoreo, J.J.; Baker, D. Controlling protein assembly on inorganic crystals through designed protein interfaces. Nature 2019, 571, 251–256. [Google Scholar] [CrossRef]
- Eguchi, Y.; Fukumori, Y.; Taoka, A. Measuring magnetosomal pH of the magnetotactic bacterium Magnetospirillum magneticum AMB-1 using pH-sensitive fluorescent proteins. Biosci. Biotechnol. Biochem. 2018, 82, 1243–1251. [Google Scholar] [CrossRef]
- Imamura, K.; Kawasaki, Y.; Awadzu, T.; Sakiyama, T.; Nakanishi, K. Contribution of acidic amino residues to the adsorption of peptides onto a stainless steel surface. J. Colloid Interface Sci. 2003, 267, 294–301. [Google Scholar] [CrossRef]
- Magro, M.; Cozza, G.; Molinari, S.; Venerando, A.; Baratella, D.; Miotto, G.; Zennaro, L.; Rossetto, M.; Frömmel, J.; Kopečná, M. Role of carboxylic group pattern on protein surface in the recognition of iron oxide nanoparticles: A key for protein corona formation. Int. J. Biol. Macromol. 2020, 164, 1715–1728. [Google Scholar] [CrossRef]
- Goobes, G.; Goobes, R.; Shaw, W.J.; Gibson, J.M.; Long, J.R.; Raghunathan, V.; Schueler-Furman, O.; Popham, J.M.; Baker, D.; Campbell, C.T. The structure, dynamics, and energetics of protein adsorption—lessons learned from adsorption of statherin to hydroxyapatite. Magn. Reason. Chem. 2007, 45, S32–S47. [Google Scholar] [CrossRef]
- Kosmulski, M. Chemical Properties of Material Surfaces; CRC Press: Boca Raton, FL, USA, 2001; Volume 102. [Google Scholar]
- Peng, Z.; Hidajat, K.; Uddin, M. Adsorption of bovine serum albumin on nanosized magnetic particles. J. Colloid Interface Sci. 2004, 271, 277–283. [Google Scholar] [CrossRef]
- De Sousa, M.E.; Fernandez van Raap, M.B.; Rivas, P.C.; Mendoza Zélis, P.; Girardin, P.; Pasquevich, G.A.; Alessandrini, J.L.; Muraca, D.; Sánchez, F.H. Stability and relaxation mechanisms of citric acid coated magnetite nanoparticles for magnetic hyperthermia. J. Phys. Chem. C 2013, 117, 5436–5445. [Google Scholar] [CrossRef] [Green Version]
- Dheyab, M.A.; Aziz, A.A.; Jameel, M.S.; Noqta, O.A.; Khaniabadi, P.M.; Mehrdel, B. Simple rapid stabilization method through citric acid modification for magnetite nanoparticles. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rehana, D.; Haleel, A.K.; Rahiman, A.K. Hydroxy, carboxylic and amino acid functionalized superparamagnetic iron oxide nanoparticles: Synthesis, characterization and in vitro anti-cancer studies. J. Chem. Sci. 2015, 127, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Hiradate, S.; Takagi, S. Interaction of mugineic acid with synthetically produced iron oxides. Soil Sci. Soc. Am. J. 1993, 57, 1254–1260. [Google Scholar] [CrossRef]
- Puddu, V.; Perry, C.C. Peptide adsorption on silica nanoparticles: Evidence of hydrophobic interactions. ACS Nano 2012, 6, 6356–6363. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, X.; Feng, S.; Wang, W.; Schmidt-Rohr, K.; Akinc, M.; Nilsen-Hamilton, M.; Vaknin, D.; Mallapragada, S. Morphological transformations in the magnetite biomineralizing protein Mms6 in iron solutions: A small-angle X-ray scattering study. Langmuir 2015, 31, 2818–2825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fincham, A.; Moradian-Oldak, J.; Simmer, J.; Sarte, P.; Lau, E.; Diekwisch, T.; Slavkin, H. Self-assembly of a recombinant amelogenin protein generates supramolecular structures. J. Struct. Biol. 1994, 112, 103–109. [Google Scholar] [CrossRef]
- Fincham, A.; Moradian-Oldak, J.; Diekwisch, T.; Lyaruu, D.; Wright, J.; Bringas, P., Jr.; Slavkin, H. Evidence for amelogenin" nanospheres" as functional components of secretory-stage enamel matrix. J. Struct. Biol. 1995, 115, 50–59. [Google Scholar] [CrossRef]
- Raj, P.A.; Johnsson, M.; Levine, M.J.; Nancollas, G.H. Salivary statherin. Dependence on sequence, charge, hydrogen bonding potency, and helical conformation for adsorption to hydroxyapatite and inhibition of mineralization. J. Biol. Chem. 1992, 267, 5968–5976. [Google Scholar] [CrossRef]
- Bouropoulos, N.; Moradian–Oldak, J. Analysis of hydroxyapatite surface coverage by amelogenin nanospheres following the Langmuir model for protein adsorption. Calcif. Tissue Int. 2003, 72, 599–603. [Google Scholar] [CrossRef]
- Del Pino, P.; Pelaz, B.; Zhang, Q.; Maffre, P.; Nienhaus, G.U.; Parak, W.J. Protein corona formation around nanoparticles–from the past to the future. Mater. Horizons 2014, 1, 301–313. [Google Scholar] [CrossRef]
- Arakaki, A.; Kikuchi, D.; Tanaka, M.; Yamagishi, A.; Yoda, T.; Matsunaga, T. Comparative subcellular localization analysis of magnetosome proteins reveals a unique localization behavior of Mms6 protein onto magnetite crystals. J. Bacteriol. 2016, 198, 2794–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, W.; Saccardo, A.; Roccatano, D.; Aboagye-Mensah, D.; Alkaseem, M.; Jewkes, M.; Di Nezza, F.; Baron, M.; Soloviev, M.; Ferrari, E. Modular assembly of proteins on nanoparticles. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Gao, J.; Ai, H.; Chen, X. Applications and potential toxicity of magnetic iron oxide nanoparticles. Small 2013, 9, 1533–1545. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Size (kDa) | pI | Origin |
|---|---|---|---|
| His-Mms6 | 6.9 | 6.0 | M. magneticum AMB-1 |
| His-Mms7 | 6.5 | 6.8 | M. magneticum AMB-1 |
| His-α synuclein | 15.5 | 5.2 | Human |
| BSA | 66.3 | 4.9 | Bovine serum |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arai, K.; Murata, S.; Wang, T.; Yoshimura, W.; Oda-Tokuhisa, M.; Matsunaga, T.; Kisailus, D.; Arakaki, A. Adsorption of Biomineralization Protein Mms6 on Magnetite (Fe3O4) Nanoparticles. Int. J. Mol. Sci. 2022, 23, 5554. https://doi.org/10.3390/ijms23105554
Arai K, Murata S, Wang T, Yoshimura W, Oda-Tokuhisa M, Matsunaga T, Kisailus D, Arakaki A. Adsorption of Biomineralization Protein Mms6 on Magnetite (Fe3O4) Nanoparticles. International Journal of Molecular Sciences. 2022; 23(10):5554. https://doi.org/10.3390/ijms23105554
Chicago/Turabian StyleArai, Kosuke, Satoshi Murata, Taifeng Wang, Wataru Yoshimura, Mayumi Oda-Tokuhisa, Tadashi Matsunaga, David Kisailus, and Atsushi Arakaki. 2022. "Adsorption of Biomineralization Protein Mms6 on Magnetite (Fe3O4) Nanoparticles" International Journal of Molecular Sciences 23, no. 10: 5554. https://doi.org/10.3390/ijms23105554