
The Role of Autophagy and Pyroptosis in Liver Disorders


1
Institute of Chronic Disease Risks Assessment, Henan University, Kaifeng 475004, China
2
School of Basic Medical Sciences, Henan University, Kaifeng 475004, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(11), 6208; https://doi.org/10.3390/ijms23116208
Submission received: 14 April 2022
/
Revised: 30 May 2022
/
Accepted: 31 May 2022
/
Published: 1 June 2022
(This article belongs to the Special Issue The Role of Autophagy Processes in Human Diseases)
Abstract
:Pyroptosis is a programmed cell death caused by inflammasomes, which can detect cell cytosolic contamination or disturbance. In pyroptosis, caspase-1 or caspase-11/4/5 is activated, cleaving gasdermin D to separate its N-terminal pore-forming domain (PFD). The oligomerization of PFD forms macropores in the membrane, resulting in swelling and membrane rupture. According to the different mechanisms, pyroptosis can be divided into three types: canonical pathway-mediated pyroptosis, non-canonical pathway-mediated pyroptosis, and caspase-3-induced pyroptosis. Pyroptosis has been reported to play an important role in many tissues and organs, including the liver. Autophagy is a highly conserved process of the eukaryotic cell cycle. It plays an important role in cell survival and maintenance by degrading organelles, proteins and macromolecules in the cytoplasm. Therefore, the dysfunction of this process is involved in a variety of pathological processes. In recent years, autophagy and pyroptosis and their interactions have been proven to play an important role in various physiological and pathological processes, and have gradually attracted more and more attention to become a research hotspot. Therefore, this review summarized the role of autophagy and pyroptosis in liver disorders, and analyzed the related mechanism to provide a basis for future research.
1. Introduction
In the 1990s, a caspase-1-dependent and bacteria-induced cell death appeared in macrophages infected with Salmonella typhimurium, named pyroptosis in 2000 [1,2,3]. Pyroptosis is a kind of gasdermin(GSDM)-mediated programmed cell death, characterized by the formation of holes in the cell membrane, cytolysis, and the release of the pro-inflammatory cytokines. Pyroptosis is an important innate immune mechanism and contributes to the inflammation through releasing interleukin 1β(IL-1β), IL-18 and other inflammatory substances [4,5]. More and more evidence indicates that pyroptosis contributes to many diseases, including liver diseases [6,7]. However, the relevant mechanisms have not been fully clarified. Autophagy is an important, closely coordinated and conserved cellular pathway. This process separates proteins and damaged or aged organelles into double-membrane vesicles named autophagosomes and finally fuses with lysosomes, resulting in the degradation of isolated components [8]. Autophagy plays an important role in maintaining the balance of cell component synthesis, decomposition and reuse, and participates in various physiological processes [9]. It has been reported that autophagy is involved in many diseases, such as cancer, neurodegenerative diseases and infection/immune diseases [10,11,12,13]. Evidence shows autophagy inhibition upregulates galangin-induced pyroptosis in human glioblastoma multiforme cells [14] and promotes pneumococcus-induced pyroptosis [15], indicating that autophagy and pyroptosis are closely related and play a vital role in a variety of physiological and pathological processes. Furthermore, in recent years, autophagy and pyroptosis and their interactions have been reported to play an important role in a variety of physiological and pathological processes, and have attracted increasing attention to become a research hotspot. However, the relevant mechanism is not completely clear [16]. Therefore, in this review, we reviewed the recent progress regarding the role and the mechanism of autophagy, pyroptosis and the relationship between them in liver disorders to provide theoretical reference for future related research.
2. Overview of Pyroptosis
2.1. Characteristics and Mechanism of Pyroptosis
In 1992, when A. Zychlinsky treated macrophages with Shigella flexneri, pyroptosis was first found [2]. However, this was regarded as a kind of apoptosis, and it was named pyroptosis until 2000 [17,18]. Pyroptosis is a programmed cell death, significantly different from apoptosis and autophagy in cell morphology and function. It is characterized by membrane perforation mediated by gasdermin(GSDM) protein family, inflammatory factors (including IL-1β and IL-18) release and cytolysis [19]. GSDM proteins include GSDMA, GSDMB, GSDMC, GSDMD, GSDME and DFNB59. Except for DFNB59, the other GSDM family proteins all have similar N-terminal parts, which is related to the formation of the pyrolytic pores in the cell membrane. It has been reported that GSDMB, GSDMC, GSDMD and GSDME are related to pyroptosis; whether other GSDM family proteins contribute to pyroptosis remains to be studied [20,21]. Many pathological factors play an important role in pyroptosis, including cholesterol, oxidative stress and inflammatory cytokines [4]. Cholesterol is one of the important structural components of the mammalian cell membrane. It can destroy the stability of the lysosomal membrane structure and lead to lysosomal damage and cause the outflow of lysosomal contents, resulting in NLRP3 inflammasome activation and pyroptosis [22]. Reactive oxygen species(ROS) produced by oxidative stress can activate NLRP3 inflammasome, then activate caspase-1 to initiate pyroptosis [23]. Pyroptosis is a highly inflammatory cell death pattern induced by inflammatory microsomes, which depends on the activation of caspase-1 [24]. Caspase-1 cleaves IL-1β precursors into active IL-1β which recruits and activates other immune cells, promotes the synthesis of chemokines (such as IL-18), inflammatory factors (such as IL-6) and adhesion factors, and finally leads to a “cascade effect”, thus amplifying the inflammatory response [25,26].
2.2. Classification of Pyroptosis
So far, there are three pathways leading to pyroptosis. One is the canonical pathway [27], the second is the non-canonical pathway [28], and the third is the recently discovered caspase-3-induced pyroptosis. After cells receive different stimuli, pyroptosis is initiated by different pathways but finally completed by GSDM protein [29]. Canonical pyroptosis is mediated by caspase-1 activated by the NLRP3 inflammasome. The active caspase-1 can be automatically cleaved into its CARD domain and P20/P10 dimers at the specific location. After that, the two P20/P10 dimers oligomerize to form a tetramer to cleave the specific site of GSDMD and accurately bind to the GSDMD-C domain [30]. GSDMD is cut into N-terminal fragments that can attach to the cell membrane and oligomerize to form the pyroptotic pore. Moreover, P20/P10 tetramers can cleave pro-IL-1β and pro-IL-18 induced by NF-κB signalling into their active forms. Furthermore, due to the water inflow caused by osmotic pressure, the pyrolysis pore can lead the cell to swell, and IL-1β and IL-18 can escape through GSDMD pores, thus inducing inflammation [28]. Unlike the canonical pathway, non-canonical pyroptosis depends on the activation of caspase-4/5/11 [31]. Human caspase 4/5 and mouse caspase 11 can bind to bacterial LPS and induce inflammation of cell necrosis. Like caspase-1, activated caspase-11 can also cleave GSDMD and release IL-1β and IL-18, then induce the formation of cell membrane pores. In addition, the activated caspase-11 can also promote K+ outflow, activate NLRP3/ASC/caspase-1 and induce cellular inflammatory response [30,32,33]. Recently, it has been found that there is also a caspase-3-dependent pyroptosis pathway. Unlike caspase-1/11/4/5, caspase-3 induces cell pore formation through cleaving GSDME and promoting the re-entry of the GSDME-N domain into the cell membrane, resulting in pyroptosis (Figure 1) [4]. The distribution and expression level of GSDME determine the cell death pattern through caspase-3 activation. When cells overexpress GSDME, activated caspase-3 will induce pyroptosis. For cells expressing a low level of GSDME, the active caspase-3 will induce apoptosis [29,34,35]. In recent years, pyroptosis has been reported to play a vital role in many diseases, including liver diseases [36,37]. However, the related mechanism is not completely clear.
3. Overview of Autophagy
Autophagy is a self-degradation and self-sustaining process in eukaryotic cells, which plays a significant role in clearing damaged organelles, proteins or cell fragments from cells [38]. In this process, the abnormal proteins, organelles, and pathogens are wrapped in bilayers to form autophagosomes that are transferred to lysosomes for degradation [39]. At present, there are three kinds of autophagy based on the transmission pathway of proteins and organelles to lysosomes: macroautophagy, microautophagy, and chaperone-mediated autophagy [40]. Macroautophagy is responsible for the degradation of microorganisms and organelles and is the most studied autophagy. In this process, the substance to be degraded is wrapped by a double membrane vesicle to form an autophagosome then fused with the lysosome for degradation. Microautophagy does not form autophagosomes and mainly degrades cell components by invaginating and/or dividing the cytoplasm on the lysosomal membrane. Chaperone-mediated autophagy is a selective autophagy in which intracellular proteins are transported to the lysosomal chamber after binding with chaperones and then digested by lysosomal enzymes(Figure 2) [41]. As we all know, autophagy is caused by various environmental stresses, such as nutrient deficiency, hypoxia and growth factor deficiency, to eliminate the stress-induced damage and help cells to return to normal after stress relief [42]. Autophagy under physiological conditions is usually maintained at a basic level. Stress stimulation can significantly enhance autophagy, thereby eliminating abnormal proteins in cells to promote cell survival [43]. However, if autophagy is at a high level for a long time, cell death will be induced. Thus, the role of autophagy is a “double-edged sword” [44,45]. Increasing evidence indicates that dysfunctional autophagy is involved in many diseases, including liver diseases [46,47], although the relevant mechanism has not been fully studied.
4. The Role of Autophagy and Pyroptosis in Liver Disorders
4.1. The Role of Autophagy and Pyroptosis in Nonalcoholic Fatty Liver Disease
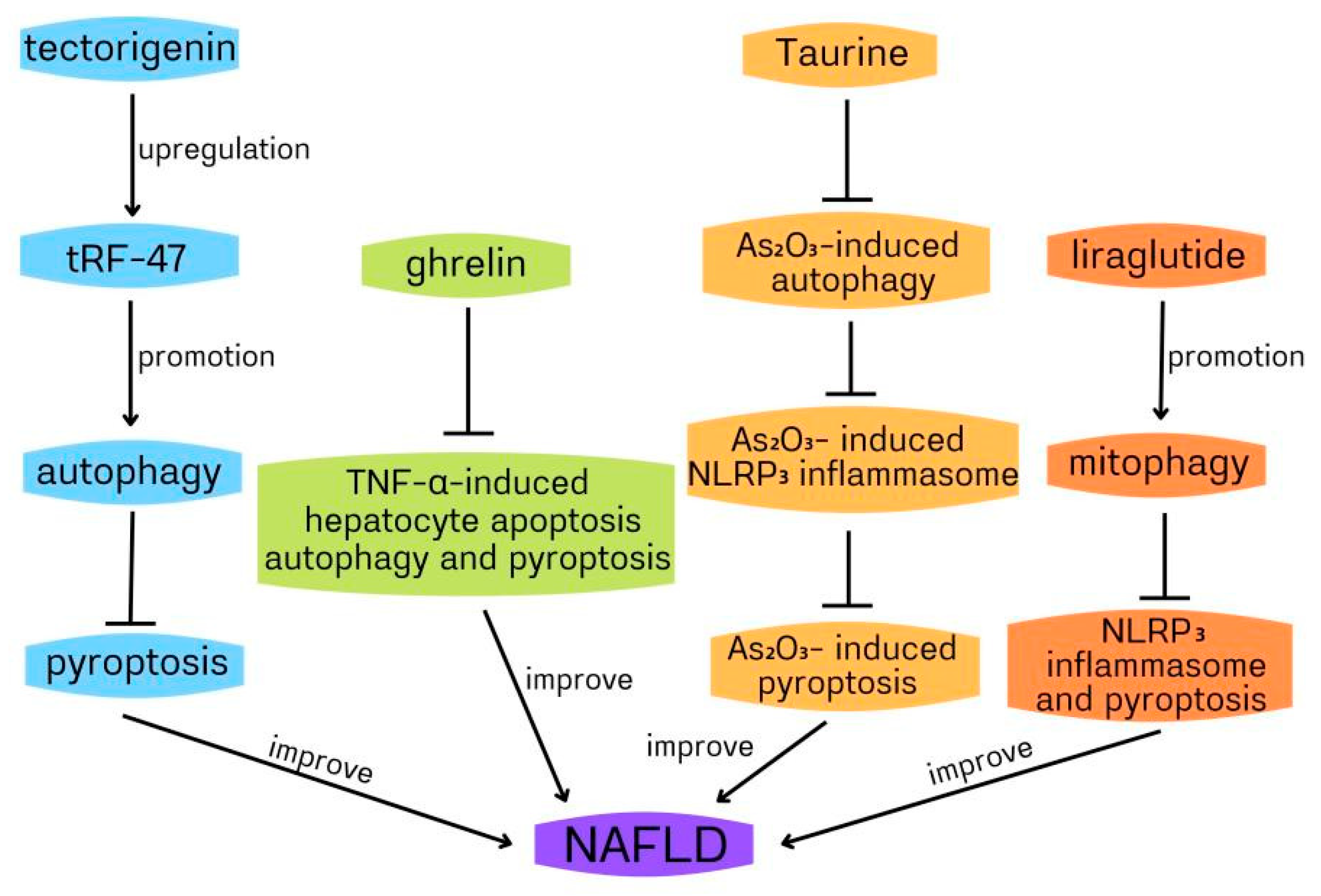
Non-alcoholic fatty liver (NAFLD) includes fatty liver, non-alcoholic steatohepatitis (NASH) and liver cirrhosis, excluding excessive drinking and viral infection. It is a clinicopathological syndrome characterized by liver fat accumulation. Because of its high incidence rate (about 20–30%) and the lack of effective clinical treatment, NAFLD has become a serious chronic disease globally [48,49,50]. NAFLD is associated with many factors, including type 2 diabetes mellitus (T2DM), insulin resistance, dyslipidaemia and hypertension, although the exact mechanism is not fully understood [51,52]. Ghrelin is a polypeptide containing 28 amino acids. It is synthesized from gastric mucosa and secreted into the blood. There are two main forms: acylated ghrelin and deacylated ghrelin [53,54]. The increasing evidence indicates that ghrelin plays an important role in NAFLD [55,56]. Silvia Ezquerro et al. showed that the circulating acylation/deacetylation ghrelin ratio and TNF-α in obese patients with NAFLD were upregulated, while the level of deacetylated ghrelin decreased. Six months after bariatric surgery, the liver function was significantly improved, and the circulating acylated/deacylated ghrelin ratio decreased. In obese patients with type 2 diabetes, ghrelin, and its acylase ghrelin O-acyltransferase(GOAT) increased, and pyroptosis, apoptosis, and compromised autophagy of liver cells increased [57]. It has been reported that TNF-α-induced hepatocyte cell death contributes to NAFLD development [58]. Thus, reducing TNF-α-induced hepatocyte cell death may become a new strategy to improve NAFLD. In HepG2 hepatocytes, the acylated and deacylated ghrelin decreased TNF-α- induced cleavage of caspase-3 and caspase-8, TUNEL positive cells, caspase-1 activation, and high-mobility group box 1(HMGB1) expression, indicating that ghrelin inhibited apoptosis and pyroptosis induced by TNF-α. In addition, the acylated ghrelin inhibited the basal and TNF-α-induced hepatocyte autophagy, which can be demonstrated by the downregulated LC3II/I ratio and upregulated p62 accumulation via AMPK/mTOR. Collectively, ghrelin reduces TNF-α-induced human hepatocyte apoptosis, autophagy, and HMGB1-mediated pyroptosis to play a protective role against hepatocyte cell death, thus preventing NAFLD progression to NASH [57]. In the above study, the impaired autophagy mediated the protective role of ghrelin in the human hepatocyte, which contradicted the previous report that ghrelin upregulated autophagy in rat hepatocytes to improve liver injury [59,60]. The reason may be related to the different stages of liver injury and remains to be clarified. It has been reported that TNF-α increases cytoplasmic HMGB1 expression to induce pyroptosis during liver failure [61]. Therefore, it can be deduced that ghrelin suppressed HMGB1-mediated pyroptosis by reducing TNF-α. In the ghrelin improvement of NAFLD, whether autophagy can regulate pyroptosis remains to be studied.
Taurine (Tau) is a sulfur-containing compound β-amino acids, which exists in many human and animal tissues, and it plays an important role in the prevention of NASH [62]. Tianming Qiu and colleagues found that arsenic trioxide(As2O3) could cause NASH, upregulated autophagy, activate NLRP3 inflammasome, increase lipid accumulation, and lead to the dysregulation of lipid-related genes. Tau dampened the inflammation, pyroptosis and autophagy induced by As2O3. In HepG2 cells, NLRP3 inflammasome activation, which was cathepsin B(CTSB)-dependent, mediated As2O3-induced pyroptosis. Moreover, the inhibition of autophagy by inhibitor suppressed As2O3-induced upregulated expression of cytosolic CTSB and subsequent LDH release, NLRP3 inflammasome activation and pyroptosis, suggesting that the increase of intracellular autophagy was related to the increase of cytoplasmic CTSB, the activation of NLRP3 inflammasome and its mediated pyroptosis induced by As2O3. In addition, Tau inhibited As2O3-induced autophagy, CTSB expression, NLRP3 inflammasome activation, and pyroptosis, and reduced LDH release. From the above, it could be deduced that Tau attenuated As2O3-induced pyroptosis through inhibiting CTSB-dependent NLRP3 inflammasome activation induced by As2O3 via suppressing As2O3-induced autophagy. Furthermore, the inhibition of NLRP3 inflammasome, autophagy, and CTSB, and Tau treatment did not reduce lipid accumulation induced by As2O3, indicating that Tau dampened As2O3-induced liver inflammation and pyroptosis by inhibiting the autophagic-CTSB-NLRP3 inflammasome pathway rather than reducing lipid accumulation [63]. Autophagic death often contributes to many liver disorders [64,65]. Similarly, in the above study, As2O3-induced autophagy leads to hepatocyte pyroptosis, which is involved in the development of NASH. In the above role of Tau in improving NASH, autophagy promotes pyroptosis through NLRP3 inflammasome. Many previous pieces of evidence have shown that autophagy is closely related to lipid metabolism [66,67,68]. Pyroptosis is also involved in lipid metabolism [69,70]; therefore, whether autophagy/pyroptosis is related to As2O3-induced liver lipid accumulation needs further study. Similar to Tau, liraglutide can also improve NASH. Liraglutide is an analogue of glucagon-like peptide-1 (GLP-1) and can improve NASH [71,72]. Xinyang Yu and colleagues used palmitic acid and lipopolysaccharide to stimulate HepG2 cells to establish a NASH model to assess the role of liraglutide, NLRP3 inflammasome and mitophagy in NASH. Liraglutide decreased lipid accumulation, suppressed NLRP3 inflammasome and pyroptosis activation, improved mitochondrial dysfunction, reduced reactive oxygen species (ROS) production, and enhanced hepatocyte mitophagy. The inhibition of mitophagy by 3-methyladenine (3-MA)/PINK1-directed siRNA dampened liraglutide suppression of NLRP3 inflammasome and pyroptosis activation, suggesting that liraglutide improved NASH by suppressing NLRP3 inflammasome and pyroptosis activation through promoting mitophagy [73]. In the above study, it can be deduced that enhancing mitophagy can alleviate NLRP3 inflammasome-mediated infammatory injury and pyroptosis of NASH. In liraglutide improvement of NASH, mitophagy inhibits pyroptosis by inhibiting NLRP3 inflammasome activation. Mitophagy can inhibit the activation of NLRP3 inflammasome by scavenging damaged mitochondria and reducing the ROS production, and then inhibit the activation of the canonical pathway of pyroptosis. Evidence indicates that LPS induces pyroptosis through a non-canonical process by activating caspase-11 [69,74]. Then, in the above study, whether mitophagy inhibits pyroptosis through caspase-11 remains to be clarified. Another study also confirmed that autophagy/pyroptosis is involved in NASH. Blueberries have been reported to improve NASH [75]; however, it is unclear which active ingredient in blueberries plays this role. Juanjuan Zhu et al. found that tectorigenin (TEC, one active ingredient in blueberries) could distinctly suppress lipid droplet formation, inflammatory mediators release, and promote cell proliferation in steatosis hepatocytes. Similarly, TEC also inhibited lipid damage and lipid accumulation induced by high-fat diets in vivo. In the NASH model of mice and cell, TEC promoted autophagy and suppressed pyroptosis and the release of inflammatory mediators. Moreover, 3-MA abolished TEC-mediated inhibition of the lipid deposition, NLRP3, and GSDEM (a marker of pyroptosis), indicating that TEC suppressed pyroptosis, NLRP3 inflammasome, and lipid deposition through promoting autophagy in the NASH model of cells. In addition, the expression of tRF-47 (a kind of tsRNAs) was upregulated by TEC. tRF-47 knockdown dampened TEC improvement of NASH in vitro through the inhibition of autophagy, activation of pyroptosis and promotion of infammatory factors release. Similarly, tRF-47 inhibition worsened the lipid deposition of NASH in vivo. Collectively, TEC ameliorated NASH by inhibiting pyroptosis through promoting autophagy via upregulating tRF-47 [76]. In the above study, enhancing autophagy suppressed pyroptosis by inhibiting NLRP3 inflammasome/GSDME in TEC improvement of NASH, which needed to be further confirmed.
In conclusion, autophagy and pyroptosis play a protective role against NAFLD (Figure 3), which will provide a new strategy for the treatment of NAFLD.
4.2. The Role of Autophagy and Pyroptosis in Hepatocellular Carcinoma
Hepatocellular carcinoma (HCC) is one of the most frequent primary liver cancers and the third leading cause of cancer death [77,78]. The evidence indicates that 17β-estradiol (E2) plays a protective role against HCC by activating the NLRP3 inflammasome [79]; however, the mechanism is unclear. The results of Qing Wei and colleagues showed that E2 induced NLRP3 inflammasome activation, evidenced by increased expression levels of caspase 1 and IL-1β in HCC cells. E2 also reduced the viability and increased the mortality rate of HepG2 cells. At the same time, caspase 1-specific inhibitor YVAD-cmk significantly reversed the E2 cytotoxic effect, indicating that E2 induced HCC cell death through activating the NLRP3 inflammasome. Further experiments showed that E2 notably downregulated autophagy in HCC cells, which was reversed by YVAD-cmk, indicating that E2 inhibition of autophagy was mediated by NLRP3 inflammasome. Additionally, the inhibition of autophagy by 3-MA significantly promoted E2-induced pyroptosis, which was reversed by YVAD-cmk, suggesting that autophagy negatively regulated caspase-1-dependent pyroptosis. Summarily, E2 induced NLRP3 inflammasome-caspase 1-dependent pyroptosis through inhibiting autophagy. Autophagy negatively regulates pyroptosis through NLRP3 inflammasome [80]. In the above study, autophagy inhibition promotes the pyroptosis of HCC cells, thus providing a new idea for HCC treatment by regulating autophagy.
4.3. The Role of Autophagy and Pyroptosis in Hepatotoxicity
Patulin is a mycotoxin produced by many common fungi in fruit and vegetable products. It has been reported that patulin induces hepatotoxicity [81,82,83]. Qian Chu et al. found that patulin promoted pyroptosis and NLRP3 inflammasome-mediated inflammation, evidenced by the upregulated expression levels of NLRP3, IL-1β, IL-18, pro-caspase-1, cleaved caspase-1 GSDMD, and cleaved GSDMD in mouse livers. Similarly, in HepG2 cells, patulin also induced pyroptosis and NLRP3 inflammasome activation, while treatment with NLRP3 inhibitor MCC950 or cathepsin B inhibitor downregulated the levels of NLRP3, caspase-1 and IL-1β, indicating that NLRP3 inflammasome/cathepsin B mediated patulin-induced pyroptosis. Caspase-1 inhibitor Ac-YVAD-cmk reduced the levels of GSDMD and IL-1β in HepG2 cells, which confirmed that patulin-induced pyroptosis was dependent on NLRP3 inflammasome. Furthermore, autophagy inhibitor 3-MA dampened patulin-induced induction of cytoplasmic cathepsin B expression, NLRP3 inflammasome activation, pyroptosis and inflammation. Collectively, patulin promoted pyroptosis perhaps through upregulating autophagy/NLRP3 inflammasome/cathepsin B in the liver. The above study showed that patulin could induce autophagy, reduce the stability of the lysosomal membrane, activate cathepsin B and then activate NLRP3 inflammasome, thus finally causing pyroptosis. That is to say, autophagy positively regulates pyroptosis through the NLRP3 inflammasome [84].
Benzo[a]pyrene (BaP) is a common polycyclic aromatic compound which is easy to be produced in the processing of petroleum and fatty food. It is a strong carcinogen and has strong immunotoxicity and reproductive toxicity [85,86,87]. As the main metabolic organ of BaP intake, the ability of the liver to metabolize BAP is much stronger than that of other organs, so the hepatotoxicity caused by BAP is also stronger than that of other organs [88,89,90]. Li Yuan and colleagues showed that BaP promoted HL-7702 cell death, upregulated the intracellular levels of ROS and inhibited HL-7702 cell growth by blocking the cell cycle in the S phase. BaP induced pyroptosis is evidenced by the increase of LDH and NO release, and the electrical conductivity of HL-7702 cells. Meanwhile, BaP also upregulated the protein expression levels of procaspase-1, caspase-1, IL-1β and IL-18. Since caspase-1 and inflammatory factors are important markers of pyroptosis activation, it could be deduced that BaP induced cell death by promoting pyroptosis. Moreover, in HL-7702 cell, BaP enhanced autophagy, and the inhibition of autophagy by 3-MA notably suppressed the release of NO and LDH, the upregulation of electrical conductivity, and the expression levels of pyroptotic marker proteins (caspase-1, IL-1β, IL-18), indicating that BaP induced pyroptosis by promoting autophagy. Futhermore, the pyroptosis inhibitor Ac-YVAD-CM also significantly abolished BaP-promoted autophagic cell death, evidenced by the increase of autophagic vacuoles and the upregulated expression of LC3-II and Beclin-1. Summarily, BaP induced HL-7702 cell death by promoting autophagy and pyroptosis simultaneously. In addition, Autophagy and pyroptosis promote each other in HL-7702 cells [91]. ROS has been involved in autophagy and pyroptosis [92]. In the above study, BaP increased ROS level and induced autophagic cell death and pyroptosis in HL-7702 cells, ROS may mediate the positive relationship between autophagy and pyroptosis.
5. Conclusions
In this review, we summarized the role of autophagy and pyroptosis in liver disorders as follows: (1) ghrelin plays a protective role against NAFLD by decreasing TNF-α-induced human hepatocyte autophagy and HMGB1-mediated pyroptosis; (2) Tau inhibits As2O3-induced pyroptosis through inhibiting CTSB-dependent NLRP3 inflammasome activation via suppressing autophagy in NASH; (3) liraglutide ameliorates NASH through inhibition of NLRP3 inflammasome and pyroptosis activation via promoting mitophagy; (4) tectorigenin improves NASH through inhibition of pyroptosis by promoting autophagy via upregulating tRF-47; (5) 17β-estradiol promotes NLRP3 inflammasome-caspase 1-dependent pyroptosis by inhibiting autophagy; (6) patulin induces pyroptosis perhaps through autophagy/NLRP3 inflammasome/cathepsin B in the liver; (7) BaP induces HL-7702 cell death by promoting autophagy and pyroptosis simultaneously (Table 1). These results indicate that autophagic death and pyroptosis can lead to hepatocyte death, play an important role in liver disorders, and may be important targets for treating various liver diseases.
It has been reported that autophagy negatively regulates pyroptosis, and the mechanisms can be summarized as follows. One is that autophagy inhibits pyroptosis by eliminating damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs). Another is that autophagy inhibits pyroptosis by inhibiting the basic components in pyroptosis [16]. In this review, autophagy regulates caspase-1-mediated canonical pyroptosis through NLRP3 inflammasome and ROS in the liver, which belongs to the second mechanism mentioned above. Whether autophagy can regulate pyroptosis through other mechanisms needs to be further studied.
It can be seen from this review that in the liver, autophagy negatively regulates pyroptosis in some cases, and autophagy and pyroptosis exist in a mutually exclusive manner. In other cases, they promote each other. This may be due to different types of cell stimulation or stimulation time because different stimuli or stimuli exposure time may lead to different autophagy and pyroptosis. The above reasons need to be further studied. At present, most existing studies on the role of autophagy and pyroptosis in the liver are in vitro. Still, there is a lack of corresponding in vivo research, which makes the basis of the research results vulnerable. Therefore, many in vivo experiments are required in the future to further verify the existing results. In addition, the role of autophagy and pyroptosis in the liver and the relationship between them have not been clearly studied and need to be further clarified in the future. Moreover, autophagy, apoptosis, NLRP3 inflammasome, and pyroptosis are closely related, so the role and relationship of the above four in liver diseases is a topic worthy of study in the future.
It is believed that with the in-depth development of relevant research, autophagy/pyroptosis will provide a new strategy for the treatment of various liver diseases.
Author Contributions
H.W. devised, wrote and funded the manuscript; H.Z. and H.L. wrote the manuscript; Y.Y. drew the figure. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from key scientific and technological projects in Henan Province, China (Grant No. 202102310153), and grants from the Construction of first-class medical disciplines in the Medical College of Henan University from 2020 to 2021 for Hongggang Wang.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Monack, D.M.; Raupach, B.; Hromockyj, A.E.; Falkow, S. Salmonella typhimurium invasion induces apoptosis in infected macrophages. Proc. Natl. Acad. Sci. USA 1996, 93, 9833–9838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zychlinsky, A.; Prevost, M.C.; Sansonetti, P.J. Shigella flexneri induces apoptosis in infected macrophages. Nature 1992, 358, 167–169. [Google Scholar] [CrossRef] [PubMed]
- Hilbi, H.; Moss, J.E.; Hersh, D.; Chen, Y.; Arondel, J.; Banerjee, S.; Flavell, R.A.; Yuan, J.; Sansonetti, P.J.; Zychlinsky, A. Shigella-induced apoptosis is dependent on caspase-1 which binds to IpaB. J. Biol. Chem. 1998, 273, 32895–32900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, N.; Qi, Z.; Wang, Y.; Yang, X.; Yan, Z.; Li, M.; Ge, Q.; Zhang, J. Pyroptosis: A New Regulating Mechanism in Cardiovascular Disease. J. Inflamm. Res. 2021, 14, 2647–2666. [Google Scholar] [CrossRef]
- Li, L.; Jiang, M.; Qi, L.; Wu, Y.; Song, D.; Gan, J.; Li, Y.; Bai, Y. Pyroptosis, a new bridge to tumor immunity. Cancer Sci. 2021, 112, 3979–3994. [Google Scholar] [CrossRef]
- Gaul, S.; Leszczynska, A.; Alegre, F.; Kaufmann, B.; Johnson, C.D.; Adams, L.A.; Wree, A.; Damm, G.; Seehofer, D.; Calvente, C.J.; et al. Hepatocyte pyroptosis and release of inflammasome particles induce stellate cell activation and liver fibrosis. J. Hepatol. 2021, 74, 156–167. [Google Scholar] [CrossRef]
- Kadono, K.; Kageyama, S.; Nakamura, K.; Hirao, H.; Ito, T.; Kojima, H.; Dery, K.J.; Li, X.; Kupiec-Weglinski, J.W. Myeloid Ikaros-SIRT1 signaling axis regulates hepatic inflammation and pyroptosis in ischemia-stressed mouse and human liver. J. Hepatol. 2022, 76, 896–909. [Google Scholar] [CrossRef]
- Behrends, C.; Sowa, M.E.; Gygi, S.P.; Harper, J.W. Network organization of the human autophagy system. Nature 2010, 466, 68–76. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Gao, J.; Li, M.; Li, H.; Wang, Y.; Li, H.; Xu, C. Dopamine D2 receptors contribute to cardioprotection of ischemic post-conditioning via activating autophagy in isolated rat hearts. Int. J. Cardiol. 2016, 203, 837–839. [Google Scholar] [CrossRef]
- Kuma, A.; Komatsu, M.; Mizushima, N. Autophagy-monitoring and autophagy-deficient mice. Autophagy 2017, 13, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Shirakabe, A.; Ikeda, Y.; Sciarretta, S.; Zablocki, D.K.; Sadoshima, J. Aging and Autophagy in the Heart. Circ. Res. 2016, 118, 1563–1576. [Google Scholar] [CrossRef] [Green Version]
- Menzies, F.M.; Fleming, A.; Rubinsztein, D.C. Compromised autophagy and neurodegenerative diseases. Nat. Rev. Neurosci. 2015, 16, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Marsh, T.; Debnath, J. Autophagy suppresses breast cancer metastasis by degrading NBR1. Autophagy 2020, 16, 1164–1165. [Google Scholar] [CrossRef]
- Kong, Y.; Feng, Z.; Chen, A.; Qi, Q.; Han, M.; Wang, S.; Zhang, Y.; Zhang, X.; Yang, N.; Wang, J.; et al. The Natural Flavonoid Galangin Elicits Apoptosis, Pyroptosis, and Autophagy in Glioblastoma. Front. Oncol. 2019, 9, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.Y.; Paton, J.C.; Briles, D.E.; Rhee, D.K.; Pyo, S. Streptococcus pneumoniae induces pyroptosis through the regulation of autophagy in murine microglia. Oncotarget 2015, 6, 44161–44178. [Google Scholar] [CrossRef] [Green Version]
- Guo, R.; Wang, H.; Cui, N. Autophagy Regulation on Pyroptosis: Mechanism and Medical Implication in Sepsis. Mediat. Inflamm. 2021, 2021, 9925059. [Google Scholar] [CrossRef]
- Cookson, B.T.; Brennan, M.A. Pro-inflammatory programmed cell death. Trends Microbiol. 2001, 9, 113–114. [Google Scholar] [CrossRef]
- Boise, L.H.; Collins, C.M. Salmonella-induced cell death: Apoptosis, necrosis or programmed cell death? Trends Microbiol. 2001, 9, 64–67. [Google Scholar] [CrossRef]
- Zhang, K.J.; Wu, Q.; Jiang, S.M.; Ding, L.; Liu, C.X.; Xu, M.; Wang, Y.; Zhou, Y.; Li, L. Pyroptosis: A New Frontier in Kidney Diseases. Oxid. Med. Cell Longev. 2021, 2021, 6686617. [Google Scholar] [CrossRef]
- Feng, S.; Fox, D.; Man, S.M. Mechanisms of Gasdermin Family Members in Inflammasome Signaling and Cell Death. J. Mol. Biol. 2018, 430, 3068–3080. [Google Scholar] [CrossRef]
- Ding, J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.; Sun, H.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Dang, E.V.; McDonald, J.G.; Russell, D.W.; Cyster, J.G. Oxysterol Restraint of Cholesterol Synthesis Prevents AIM2 Inflammasome Activation. Cell 2017, 171, 1057–1071.e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minutoli, L.; Puzzolo, D.; Rinaldi, M.; Irrera, N.; Marini, H.; Arcoraci, V.; Bitto, A.; Crea, G.; Pisani, A.; Squadrito, F.; et al. ROS-Mediated NLRP3 Inflammasome Activation in Brain, Heart, Kidney, and Testis Ischemia/Reperfusion Injury. Oxid. Med. Cell. Longev. 2016, 2016, 2183026. [Google Scholar] [CrossRef] [PubMed]
- Tavakoli Dargani, Z.; Singla, R.; Johnson, T.; Kukreja, R.; Singla, D.K. Exosomes derived from embryonic stem cells inhibit doxorubicin and inflammation-induced pyroptosis in muscle cells. Can. J. Physiol. Pharm. 2018, 96, 304–307. [Google Scholar] [CrossRef] [Green Version]
- Zeng, C.; Wang, R.; Tan, H. Role of Pyroptosis in Cardiovascular Diseases and its Therapeutic Implications. Int. J. Biol. Sci. 2019, 15, 1345–1357. [Google Scholar] [CrossRef] [Green Version]
- Que, Y.Y.; Zhu, T.; Zhang, F.X.; Peng, J. Neuroprotective effect of DUSP14 overexpression against isoflurane-induced inflammatory response, pyroptosis and cognitive impairment in aged rats through inhibiting the NLRP3 inflammasome. Eur. Rev. Med. Pharm. Sci. 2020, 24, 7101–7113. [Google Scholar]
- Jorgensen, I.; Rayamajhi, M.; Miao, E.A. Programmed cell death as a defence against infection. Nat. Rev. Immunol. 2017, 17, 151–164. [Google Scholar] [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O’Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666–671. [Google Scholar] [CrossRef]
- Wang, Y.; Gao, W.; Shi, X.; Ding, J.; Liu, W.; He, H.; Wang, K.; Shao, F. Chemotherapy drugs induce pyroptosis through caspase-3 cleavage of a gasdermin. Nature 2017, 547, 99–103. [Google Scholar] [CrossRef]
- Wang, K.; Sun, Q.; Zhong, X.; Zeng, M.; Zeng, H.; Shi, X.; Li, Z.; Wang, Y.; Zhao, Q.; Shao, F.; et al. Structural Mechanism for GSDMD Targeting by Autoprocessed Caspases in Pyroptosis. Cell 2020, 180, 941–955.e20. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, Y.; Wang, Y.; Gao, W.; Ding, J.; Li, P.; Hu, L.; Shao, F. Inflammatory caspases are innate immune receptors for intracellular LPS. Nature 2014, 514, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, X.; Zheng, Y.; Jiang, J.; Hu, J. What role does pyroptosis play in microbial infection? J. Cell Physiol. 2019, 234, 7885–7892. [Google Scholar] [CrossRef] [PubMed]
- Wandel, M.P.; Kim, B.H.; Park, E.S.; Boyle, K.B.; Nayak, K.; Lagrange, B.; Herod, A.; Henry, T.; Zilbauer, M.; Rohde, J.; et al. Guanylate-binding proteins convert cytosolic bacteria into caspase-4 signaling platforms. Nat. Immunol. 2020, 21, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhong, T.; Ma, Y.; Wan, X.; Qin, A.; Yao, B.; Zou, H.; Song, Y.; Yin, D. Bnip3 mediates doxorubicin-induced cardiomyocyte pyroptosis via caspase-3/GSDME. Life Sci. 2020, 242, 117186. [Google Scholar] [CrossRef]
- Zhang, C.C.; Li, C.G.; Wang, Y.F.; Xu, L.H.; He, X.H.; Zeng, Q.Z.; Zeng, C.Y.; Mai, F.Y.; Hu, B.; Ouyang, D.Y. Chemotherapeutic paclitaxel and cisplatin differentially induce pyroptosis in A549 lung cancer cells via caspase-3/GSDME activation. Apoptosis 2019, 24, 312–325. [Google Scholar] [CrossRef]
- Oh, S.; Son, M.; Byun, K.A.; Jang, J.T.; Choi, C.H.; Son, K.H.; Byun, K. Attenuating Effects of Dieckol on High-Fat Diet-Induced Nonalcoholic Fatty Liver Disease by Decreasing the NLRP3 Inflammasome and Pyroptosis. Mar. Drugs 2021, 19, 318. [Google Scholar] [CrossRef]
- Zhang, X.; Shang, X.; Jin, S.; Ma, Z.; Wang, H.; Ao, N.; Yang, J.; Du, J. Vitamin D ameliorates high-fat-diet-induced hepatic injury via inhibiting pyroptosis and alters gut microbiota in rats. Arch. Biochem. Biophys. 2021, 705, 108894. [Google Scholar] [CrossRef]
- Buzun, K.; Gornowicz, A.; Lesyk, R.; Bielawski, K.; Bielawska, A. Autophagy Modulators in Cancer Therapy. Int. J. Mol. Sci. 2021, 22, 5804. [Google Scholar] [CrossRef]
- Wang, J.; Wu, D.; Wang, H. Hydrogen sulfide plays an important protective role by influencing autophagy in diseases. Physiol. Res. 2019, 68, 335–345. [Google Scholar] [CrossRef]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Wang, H.; Li, X. The Role of the Interplay Between Autophagy and NLRP3 Inflammasome in Metabolic Disorders. Front. Cell Dev. Biol. 2021, 9, 634118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H. Regulation of Autophagy by mTOR Signaling Pathway. Adv. Exp. Med. Biol. 2019, 1206, 67–83. [Google Scholar] [PubMed]
- Glick, D.; Barth, S.; Macleod, K.F. Autophagy: Cellular and molecular mechanisms. J. Pathol. 2010, 221, 3–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Huerta, P.; Troncoso-Escudero, P.; Jerez, C.; Hetz, C.; Vidal, R.L. The intersection between growth factors, autophagy and ER stress: A new target to treat neurodegenerative diseases? Brain Res. 2016, 1649, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Nasiri-Ansari, N.; Nikolopoulou, C.; Papoutsi, K.; Kyrou, I.; Mantzoros, C.S.; Kyriakopoulos, G.; Chatzigeorgiou, A.; Kalotychou, V.; Randeva, M.S.; Chatha, K.; et al. Empagliflozin Attenuates Non-Alcoholic Fatty Liver Disease (NAFLD) in High Fat Diet Fed ApoE((-/-)) Mice by Activating Autophagy and Reducing ER Stress and Apoptosis. Int. J. Mol. Sci. 2021, 22, 818. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, A.; Grossi, I.; Garcia-Gomez, R.; Patel, G.A.; Salvi, A.; Lavazza, A.; De Petro, G.; Monsalve, M.; Rezzani, R. Melatonin Effects on Non-Alcoholic Fatty Liver Disease Are Related to MicroRNA-34a-5p/Sirt1 Axis and Autophagy. Cells 2019, 8, 1053. [Google Scholar] [CrossRef] [Green Version]
- Santhekadur, P.K.; Kumar, D.P.; Sanyal, A.J. Preclinical models of non-alcoholic fatty liver disease. J. Hepatol. 2018, 68, 230–237. [Google Scholar] [CrossRef]
- Shen, X.; Jin, C.; Wu, Y.; Zhang, Y.; Wang, X.; Huang, W.; Li, J.; Wu, S.; Gao, X. Prospective study of perceived dietary salt intake and the risk of non-alcoholic fatty liver disease. J. Hum. Nutr. Diet. 2019, 32, 802–809. [Google Scholar] [CrossRef]
- DeWeerdt, S. Disease progression: Divergent paths. Nature 2017, 551, S92–S93. [Google Scholar] [CrossRef] [Green Version]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Charlton, M.; Cusi, K.; Rinella, M.; Harrison, S.A.; Brunt, E.M.; Sanyal, A.J. The diagnosis and management of nonalcoholic fatty liver disease: Practice guidance from the American Association for the Study of Liver Diseases. Hepatology 2018, 67, 328–357. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.M.; Stepanova, M.; Younossi, Y.; Golabi, P.; Mishra, A.; Rafiq, N.; Henry, L. Epidemiology of chronic liver diseases in the USA in the past three decades. Gut 2020, 69, 564–568. [Google Scholar] [CrossRef] [PubMed]
- Yamada, C.; Hattori, T.; Ohnishi, S.; Takeda, H. Ghrelin Enhancer, the Latest Evidence of Rikkunshito. Front. Nutr. 2021, 8, 761631. [Google Scholar] [CrossRef] [PubMed]
- Ginter, G.; Ceranowicz, P.; Warzecha, Z. Protective and Healing Effects of Ghrelin and Risk of Cancer in the Digestive System. Int. J. Mol. Sci. 2021, 22, 10571. [Google Scholar] [CrossRef]
- Chen, M.; Xing, J.; Pan, D.; Gao, P. Effect of Chinese Herbal Medicine Mixture 919 Syrup on Regulation of the Ghrelin Pathway and Intestinal Microbiota in Rats with Non-alcoholic Fatty Liver Disease. Front. Microbiol. 2021, 12, 793854. [Google Scholar] [CrossRef]
- Yin, Y.; Wang, Q.; Qi, M.; Zhang, C.; Li, Z.; Zhang, W. Ghrelin ameliorates nonalcoholic steatohepatitis induced by chronic low-grade inflammation via blockade of Kupffer cell M1 polarization. J. Cell Physiol. 2021, 236, 5121–5133. [Google Scholar] [CrossRef]
- Ezquerro, S.; Mocha, F.; Fruhbeck, G.; Guzman-Ruiz, R.; Valenti, V.; Mugueta, C.; Becerril, S.; Catalan, V.; Gomez-Ambrosi, J.; Silva, C.; et al. Ghrelin Reduces TNF-alpha-Induced Human Hepatocyte Apoptosis, Autophagy, and Pyroptosis: Role in Obesity-Associated NAFLD. J. Clin. Endocrinol. Metab. 2019, 104, 21–37. [Google Scholar]
- Komiya, C.; Tanaka, M.; Tsuchiya, K.; Shimazu, N.; Mori, K.; Furuke, S.; Miyachi, Y.; Shiba, K.; Yamaguchi, S.; Ikeda, K.; et al. Antifibrotic effect of pirfenidone in a mouse model of human nonalcoholic steatohepatitis. Sci. Rep. 2017, 7, 44754. [Google Scholar] [CrossRef] [Green Version]
- Ezquerro, S.; Mendez-Gimenez, L.; Becerril, S.; Moncada, R.; Valenti, V.; Catalan, V.; Gomez-Ambrosi, J.; Fruhbeck, G.; Rodriguez, A. Acylated and desacyl ghrelin are associated with hepatic lipogenesis, beta-oxidation and autophagy: Role in NAFLD amelioration after sleeve gastrectomy in obese rats. Sci. Rep. 2016, 6, 39942. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Cheng, J.; Yu, F.; Li, H.; Guo, C.; Fan, X. Ghrelin Attenuated Lipotoxicity via Autophagy Induction and Nuclear Factor-kappaB Inhibition. Cell Physiol. Biochem. 2015, 37, 563–576. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, H.; Chen, Q.; Jiao, F.; Shi, C.; Pei, M.; Lv, J.; Zhang, H.; Wang, L.; Gong, Z. TNF-alpha/HMGB1 inflammation signalling pathway regulates pyroptosis during liver failure and acute kidney injury. Cell Prolif. 2020, 53, e12829. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Chen, S.; Chen, R.; Peng, Z.; Wan, J.; Wu, B. Taurine and tea polyphenols combination ameliorate nonalcoholic steatohepatitis in rats. BMC Complement. Altern. Med. 2017, 17, 455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, T.; Pei, P.; Yao, X.; Jiang, L.; Wei, S.; Wang, Z.; Bai, J.; Yang, G.; Gao, N.; Yang, L.; et al. Taurine attenuates arsenic-induced pyroptosis and nonalcoholic steatohepatitis by inhibiting the autophagic-inflammasomal pathway. Cell Death Dis. 2018, 9, 946. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Xu, X.; Che, D.; Fan, R.; Gao, M.; Cao, Y.; Ge, C.; Feng, Y.; Li, J.; Xie, S.; et al. Reactive Oxygen Species Mediate 6c-Induced Mitochondrial and Lysosomal Dysfunction, Autophagic Cell Death, and DNA Damage in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 10987. [Google Scholar] [CrossRef] [PubMed]
- Pramanick, A.; Chakraborti, S.; Mahata, T.; Basak, M.; Das, K.; Verma, S.K.; Sengar, A.S.; Singh, P.K.; Kumar, P.; Bhattacharya, B.; et al. G protein beta5-ATM complexes drive acetaminophen-induced hepatotoxicity. Redox. Biol. 2021, 43, 101965. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Wang, L.; Zhao, R.; Yan, F.; Sha, S.; Cui, C.; Song, J.; Hu, H.; Guo, X.; Yang, M.; et al. Mesenchymal stem cell-derived exosomes exert ameliorative effects in type 2 diabetes by improving hepatic glucose and lipid metabolism via enhancing autophagy. Stem. Cell Res. 2020, 11, 223. [Google Scholar] [CrossRef]
- Park, H.S.; Song, J.W.; Park, J.H.; Lim, B.K.; Moon, O.S.; Son, H.Y.; Lee, J.H.; Gao, B.; Won, Y.S.; Kwon, H.J. TXNIP/VDUP1 attenuates steatohepatitis via autophagy and fatty acid oxidation. Autophagy 2021, 17, 2549–2564. [Google Scholar] [CrossRef]
- Tanaka, S.; Hikita, H.; Tatsumi, T.; Sakamori, R.; Nozaki, Y.; Sakane, S.; Shiode, Y.; Nakabori, T.; Saito, Y.; Hiramatsu, N.; et al. Rubicon inhibits autophagy and accelerates hepatocyte apoptosis and lipid accumulation in nonalcoholic fatty liver disease in mice. Hepatology 2016, 64, 1994–2014. [Google Scholar] [CrossRef]
- Qiu, Z.; He, Y.; Ming, H.; Lei, S.; Leng, Y.; Xia, Z.Y. Lipopolysaccharide (LPS) Aggravates High Glucose- and Hypoxia/Reoxygenation-Induced Injury through Activating ROS-Dependent NLRP3 Inflammasome-Mediated Pyroptosis in H9C2 Cardiomyocytes. J. Diabetes Res. 2019, 2019, 8151836. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.; Zhu, M.; Liu, X.; Chen, X.; Yuan, Y.; Li, L.; Liu, J.; Lu, Y.; Cheng, J.; Chen, Y. Oleic acid ameliorates palmitic acid induced hepatocellular lipotoxicity by inhibition of ER stress and pyroptosis. Nutr. Metab. 2020, 17, 11. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, M.J.; Hull, D.; Guo, K.; Barton, D.; Hazlehurst, J.M.; Gathercole, L.L.; Nasiri, M.; Yu, J.; Gough, S.C.; Newsome, P.N.; et al. Glucagon-like peptide 1 decreases lipotoxicity in non-alcoholic steatohepatitis. J. Hepatol. 2016, 64, 399–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perakakis, N.; Stefanakis, K.; Feigh, M.; Veidal, S.S.; Mantzoros, C.S. Elafibranor and liraglutide improve differentially liver health and metabolism in a mouse model of non-alcoholic steatohepatitis. Liver Int. 2021, 41, 1853–1866. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Hao, M.; Liu, Y.; Ma, X.; Lin, W.; Xu, Q.; Zhou, H.; Shao, N.; Kuang, H. Liraglutide ameliorates non-alcoholic steatohepatitis by inhibiting NLRP3 inflammasome and pyroptosis activation via mitophagy. Eur. J. Pharm. 2019, 864, 172715. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.R.; Xing, R.L.; Wang, P.M.; Zhang, N.S.; Yin, S.J.; Li, X.C.; Zhang, L. NLRP1 and NLRP3 inflammasomes mediate LPS/ATPinduced pyroptosis in knee osteoarthritis. Mol. Med. Rep. 2018, 17, 5463–5469. [Google Scholar]
- Ren, T.; Zhu, J.; Zhu, L.; Cheng, M. The Combination of Blueberry Juice and Probiotics Ameliorate Non-Alcoholic Steatohepatitis (NASH) by Affecting SREBP-1c/PNPLA-3 Pathway via PPAR-alpha. Nutrients 2017, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Wen, Y.; Zhang, Q.; Nie, F.; Cheng, M.; Zhao, X. The monomer TEC of blueberry improves NASH by augmenting tRF-47-mediated autophagy/pyroptosis signaling pathway. J. Transl. Med. 2022, 20, 128. [Google Scholar] [CrossRef]
- Sugawara, Y.; Hibi, T. Surgical treatment of hepatocellular carcinoma. Biosci. Trends 2021, 15, 138–141. [Google Scholar] [CrossRef]
- El-Khoueiry, A.B.; Hanna, D.L.; Llovet, J.; Kelley, R.K. Cabozantinib: An evolving therapy for hepatocellular carcinoma. Cancer Treat. Rev. 2021, 98, 102221. [Google Scholar] [CrossRef]
- Wei, Q.; Guo, P.; Mu, K.; Zhang, Y.; Zhao, W.; Huai, W.; Qiu, Y.; Li, T.; Ma, X.; Liu, Y.; et al. Estrogen suppresses hepatocellular carcinoma cells through ERbeta-mediated upregulation of the NLRP3 inflammasome. Lab. Investig. 2015, 95, 804–816. [Google Scholar] [CrossRef]
- Wei, Q.; Zhu, R.; Zhu, J.; Zhao, R.; Li, M. E2-Induced Activation of the NLRP3 Inflammasome Triggers Pyroptosis and Inhibits Autophagy in HCC Cells. Oncol. Res. 2019, 27, 827–834. [Google Scholar] [CrossRef]
- Cui, J.; Yin, S.; Zhao, C.; Fan, L.; Hu, H. Combining Patulin with Cadmium Induces Enhanced Hepatotoxicity and Nephrotoxicity In Vitro and In Vivo. Toxins 2021, 13, 221. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Liu, S.; Cui, J.; Liu, X.; Zhao, C.; Fan, L.; Yin, S.; Hu, H. Combination of Patulin and Chlorpyrifos Synergistically Induces Hepatotoxicity via Inhibition of Catalase Activity and Generation of Reactive Oxygen Species. J. Agric. Food Chem. 2019, 67, 11474–11480. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Huang, C.; Lu, Q.; Liang, H.; Li, J.; Xu, D. Involvement of caspase in patulin-induced hepatotoxicity in vitro and in vivo. Toxicon 2022, 206, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Chu, Q.; Wang, S.; Jiang, L.; Jiao, Y.; Sun, X.; Li, J.; Yang, L.; Hou, Y.; Wang, N.; Yao, X.; et al. Patulin induces pyroptosis through the autophagic-inflammasomal pathway in liver. Food Chem. Toxicol. 2021, 147, 111867. [Google Scholar] [CrossRef]
- Quintanilla-Mena, M.; Vega-Arreguin, J.; Del Rio-Garcia, M.; Patino-Suarez, V.; Peraza-Echeverria, S.; Puch-Hau, C. The effect of benzo[a]pyrene on the gut microbiota of Nile tilapia (Oreochromis niloticus). Appl. Microbiol. Biotechnol. 2021, 105, 7935–7947. [Google Scholar] [CrossRef]
- Yanagisawa, R.; Koike, E.; Takano, H. Benzo[a]pyrene aggravates atopic dermatitis-like skin lesions in mice. Hum. Exp. Toxicol. 2021, 40, S269–S277. [Google Scholar] [CrossRef]
- Guarnieri, G.; Becatti, M.; Comeglio, P.; Vignozzi, L.; Maggi, M.; Vannelli, G.B.; Morelli, A. Benzo[a]pyrene impairs the migratory pattern of human gonadotropin-releasing-hormone-secreting neuroblasts. Eur. J. Histochem. 2021, 65, 3282. [Google Scholar] [CrossRef]
- Owumi, S.E.; Adeniyi, G.; Oyelere, A.K. The modulatory effect of taurine on benzo (a) pyrene-induced hepatorenal toxicity. Toxicol. Res. 2021, 10, 389–398. [Google Scholar] [CrossRef]
- Mantey, J.A.; Rekhadevi, P.V.; Diggs, D.L.; Ramesh, A. Metabolism of benzo(a)pyrene by subcellular fractions of gastrointestinal (GI) tract and liver in Apc(Min) mouse model of colon cancer. Tumour Biol. 2014, 35, 4929–4935. [Google Scholar] [CrossRef]
- Lee, S.C.; Jee, S.C.; Kim, M.; Kim, S.; Shin, M.K.; Kim, Y.; Sung, J.S. Curcumin Suppresses the Lipid Accumulation and Oxidative Stress Induced by Benzo[a]pyrene Toxicity in HepG2 Cells. Antioxidants 2021, 10, 1314. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, J.; Deng, H.; Gao, C. Benzo[a]pyrene Induces Autophagic and Pyroptotic Death Simultaneously in HL-7702 Human Normal Liver Cells. J. Agric. Food Chem. 2017, 65, 9763–9773. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Zhong, G.; Wu, S.; Jiang, X.; Liao, J.; Zhang, X.; Zhang, H.; Mehmood, K.; Tang, Z.; Hu, L. Arsenic and antimony co-induced nephrotoxicity via autophagy and pyroptosis through ROS-mediated pathway in vivo and in vitro. Ecotoxicol. Environ. Saf. 2021, 221, 112442. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The schematic diagram of the process of three types of pyroptosis.

Figure 2.
The Diagram of the process of three kinds of autophagy.

Figure 3.
The protective role of autophagy and pyroptosis against non-alcoholic fatty liver disease (NAFLD).
Figure 3.
The protective role of autophagy and pyroptosis against non-alcoholic fatty liver disease (NAFLD).


{kind=link}
{kind=link}
{kind=link}
Table 1.
The summary of the role of autophagy and pyroptosis in liver disorders.
| The Type of Liver Disorder | The Role of Autophagy and Pyroptosis | Experimental Model | Reference |
|---|---|---|---|
| non-alcoholic fatty liver disease (NAFLD) | ghrelin plays a protective role against NAFLD by decreasing TNF-α-induced human hepatocyte autophagy and HMGB1-mediated pyroptosis | liver biopsies of NAFLD patients and human hepG2 hepatocytes | [57] |
| non-alcoholic steatohepatitis(NASH) | Tau inhibits As2O3-induced pyroptosis by inhibiting CTSB-dependent NLRP3 inflammasome activation via suppressing autophagy | NASH model of mice/human hepG2 hepatocytes | [63] |
| NASH | liraglutide ameliorates NASH through inhibition of NLRP3 inflammasome and pyroptosis activation via promoting mitophagy | NASH model of mice/human hepG2 cells | [73] |
| NASH | tectorigenin improves NASH through inhibition of pyroptosis by promoting autophagy via upregulating tRF-47 | NASH model of mice/human hepG2 cells | [76] |
| hepatocellular carcinoma(HCC) | 17β-estradiol promotes NLRP3 inflammasome-caspase 1-dependent pyroptosis by inhibiting autophagyd | HCC cells | [80] |
| hepatotoxicity | patulin induces pyroptosis perhaps through autophagy/NLRP3 inflammasome/cathepsin B in liver | mice/human HepG2 cells and L02 cells | [84] |
| liver injury | BaP induces HL-7702 cell death by promoting autophagy and pyroptosis simultaneously | HL-7702 cells | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, H.; Liu, H.; Yang, Y.; Wang, H. The Role of Autophagy and Pyroptosis in Liver Disorders. Int. J. Mol. Sci. 2022, 23, 6208. https://doi.org/10.3390/ijms23116208
AMA Style
Zhao H, Liu H, Yang Y, Wang H. The Role of Autophagy and Pyroptosis in Liver Disorders. International Journal of Molecular Sciences. 2022; 23(11):6208. https://doi.org/10.3390/ijms23116208
Chicago/Turabian StyleZhao, Huijie, Huiyang Liu, Yihan Yang, and Honggang Wang. 2022. "The Role of Autophagy and Pyroptosis in Liver Disorders" International Journal of Molecular Sciences 23, no. 11: 6208. https://doi.org/10.3390/ijms23116208
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.