
Salivary Effector Sm9723 of Grain Aphid Sitobion miscanthi Suppresses Plant Defense and Is Essential for Aphid Survival on Wheat

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis of Candidate Salivary Effector Sm9723
2.2. Expression Levels of Candidate Effector Sm9723 of S. miscanthi in Different Tissues and Various Aphid Feeding Time Points on Wheat
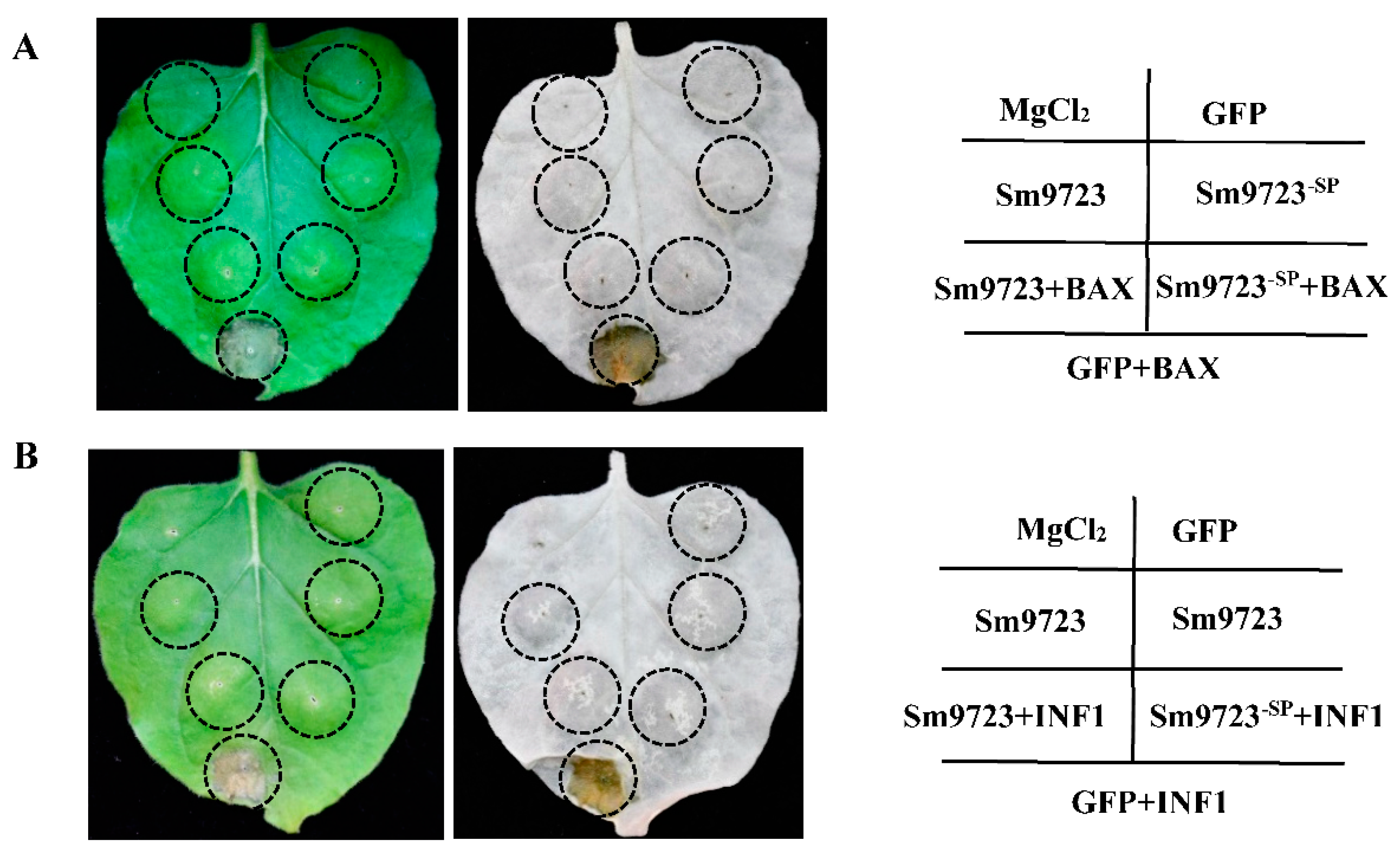
2.3. Overexpression of Sm9723 in Nicotiana benthamiana Suppresses Cell Death
2.4. Transient Overexpression of Sm9723 in N. benthamiana Suppresses PTI-Associated Callose Deposition and Plant Defense Responses
2.5. Silencing of Sm9723 by a Nanocarrier-Mediated dsRNA Delivery System
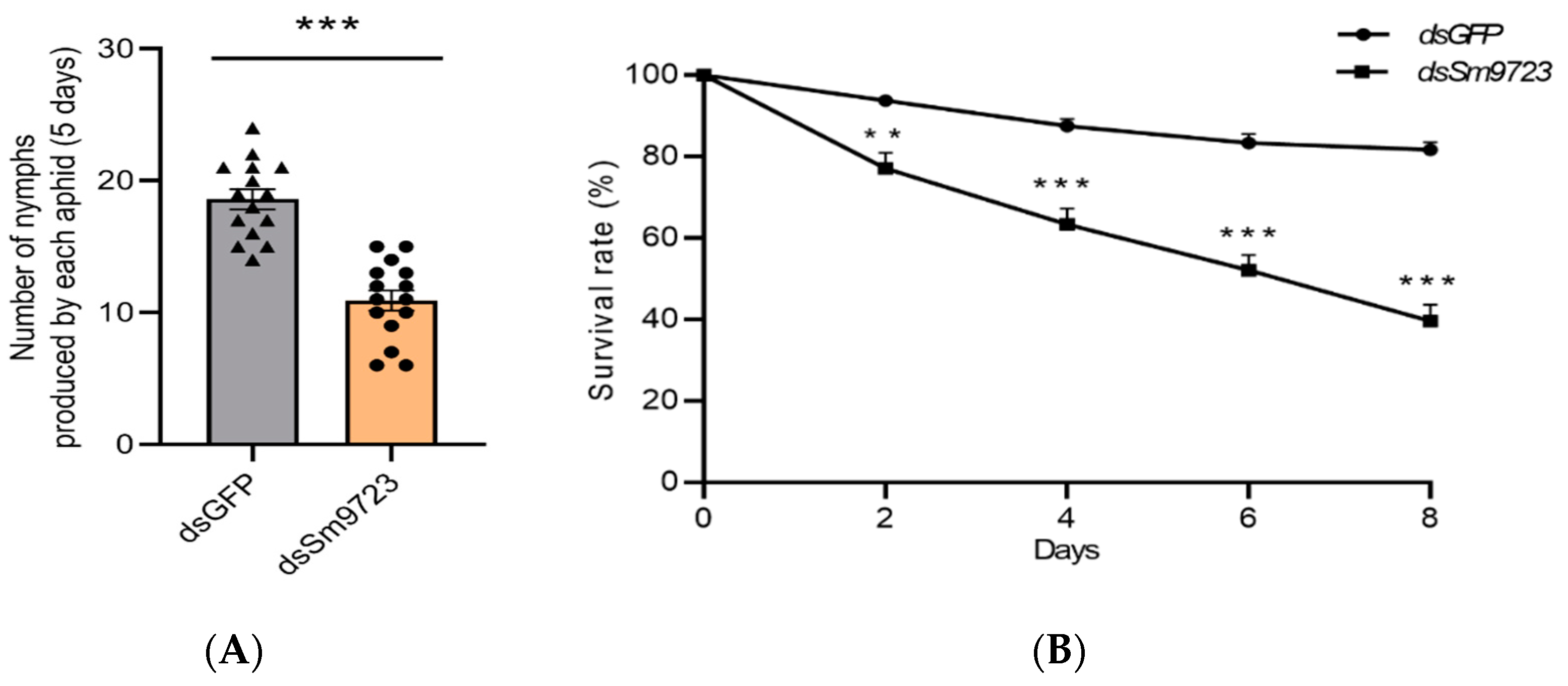
2.6. Effects of Sm9723 Silencing on Fecundity and Survival Rate of S. miscanthi
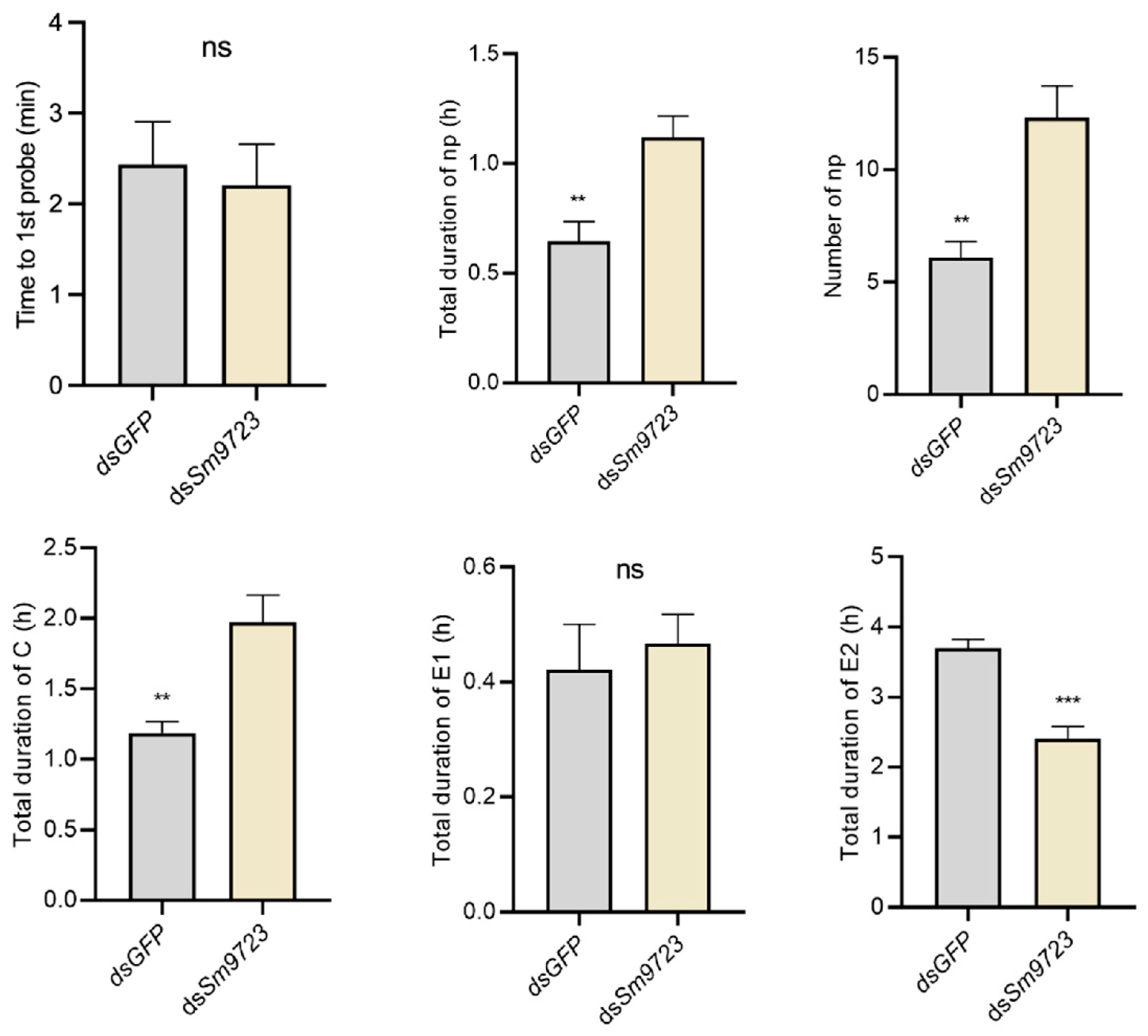
2.7. Effects of Sm9723 Silencing on S. miscanthi Feeding Behavior
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Aphids and Plants
5.2. Sequence Analysis
5.3. Agrobacterium tumefaciens Infiltration Assays to Suppress BAX/INF1-Induced Programmed Cell Death
5.4. Detecting Callose Deposition in N. benthamiana Leaves
5.5. RNAi Application Using the Nanocarrier-Mediated Transdermal dsRNA Delivery System
5.6. Aphid Performance
5.7. Aphid Feeding Behavior Monitoring by Electronic Penetration Graph (EPG)
5.8. RT-qPCR
5.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification and Information Guide; John Wiley & Sons Ltd.: New York, NY, USA, 2000. [Google Scholar]
- Guerrieri, E.; Digilio, M.C. Aphid-plant interactions: A review. J. Plant Interact. 2008, 3, 223–232. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Esch, T.H. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Miles, P.W. Aphid saliva. Biol. Rev. 1999, 74, 41–85. [Google Scholar] [CrossRef]
- Carolan, J.C.; Fitzroy, C.I.J.; Ashton, P.D.; Douglas, A.E.; Wilkinson, T.L. The secreted salivary proteome of the pea aphid Acyrthosiphon pisum characterized by mass spectrometry. Proteomics 2009, 9, 2457–2467. [Google Scholar] [CrossRef] [PubMed]
- Will, T.; Kornemann, S.R.; Furch, A.C.U.; Tjallingii, W.F.; van Bel, A.J.E. Aphid watery saliva counteracts sieve-tube occlusion: A universal phenomenon? J. Exp. Biol. 2009, 212, 3305–3312. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.R.; Dillwith, J.W.; Puterka, G.J. Comparison of salivary proteins from five aphid (Hemiptera: Aphididae) species. Environ. Entomol. 2011, 40, 151–156. [Google Scholar] [CrossRef]
- Will, T.; Steckbauer, K.; Hardt, M.; van Bel, A.J.E. Aphid gel saliva: Sheath structure, protein composition and secretory dependence on stylet-tip milieu. PLoS ONE 2012, 7, e46903. [Google Scholar] [CrossRef]
- Tjallingii, W.F. Salivary secretions by aphids interacting with proteins of phloem wound responses. J. Exp. Biol. 2006, 57, 739–745. [Google Scholar] [CrossRef]
- Furch, A.C.U.; Van Bel, A.J.E.; Torsten, W. Aphid salivary proteases are capable. of degrading sieve-tube proteins. J. Exp. Bot. 2015, 66, 533–539. [Google Scholar] [CrossRef] [Green Version]
- van Bel, A.J.E.; Torsten, W. Functional evaluation of proteins in watery and gel saliva of aphids. Front. Plant Sci. 2016, 7, 1840. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Rodriguez, P.A.; Bos, J.I.B. Toward understanding the role of aphid effectors in plant infestation. Mol. Plant Microbe Interact. 2013, 26, 25–30. [Google Scholar] [CrossRef] [Green Version]
- De Vos, M.; Jander, G. Myzus persicae (green peach aphid) salivary components induce defence responses in Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1548–1560. [Google Scholar] [CrossRef]
- Bos, J.I.; Prince, D.; Pitino, M.; Maffei, M.E.; Win, J.; Hogenhout, S.A. A functional genomics approach identifies candidate effectors from the aphid species Myzus persicae (green peach aphid). PLoS Genet. 2010, 6, e1001216. [Google Scholar] [CrossRef]
- Rodriguez, P.A.; Stam, R.; Warbroek, T.; Bos, J.I. Mp10 and Mp42 from the aphid species Myzus persicae trigger plant defenses in Nicotiana benthamiana through different activities. Mol. Plant Microbe Interact. 2014, 27, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Dai, H.; Zhang, Y.; Chandrasekar, R.; Luo, L.; Hiromasa, Y.; Sheng, C.; Peng, G.; Chen, S.; Tomich, J.M.; et al. Armet is an effector protein mediating aphid–plant interactions. FASEB J. 2019, 29, 2032–2045. [Google Scholar] [CrossRef]
- Mutti, N.S.; Park, Y.; Reese, J.C.; Reeck, G.R. RNAi knockdown of a salivary transcript leading to lethality in the pea aphid, Acyrthosiphon pisum. J. Insect Sci. 2006, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Mutti, N.S.; Louis, J.; Pappan, L.K.; Pappan, K.; Begum, K.; Chen, M.S.; Park, Y.; Dittmer, N.; Marshall, J.; Reese, J.C.; et al. A protein from the salivary glands of the pea aphid, Acyrthosiphon pisum, is essential in feeding on a host plant. Proc. Natl. Acad. Sci. USA 2008, 105, 9965–9969. [Google Scholar] [CrossRef] [Green Version]
- Atamian, H.S.; Chaudhary, R.; Cin, V.D.; Bao, E.; Girke, T.; Kaloshian, I. In planta expression or delivery of potato aphid Macrosiphum euphorbiae effectors Me10 and Me23 enhances aphid fecundity. Mol. Plant Microbe Interact. 2013, 26, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Elzinga, D.A.; De Vos, M.; Jander, G. Suppression of plant defenses by a Myzus persicae (green peach aphid) salivary effector protein. Mol. Plant Microbe Interact. 2014, 27, 747–756. [Google Scholar] [CrossRef] [Green Version]
- Kettles, G.J.; Kaloshian, I. The potato aphid salivary effector Me47 is a glutathione-S-transferase involved in modifying plant responses to aphid infestation. Front. Plant Sci. 2016, 7, e1142. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, R.; Peng, H.C.; He, J.; MacWilliams, J.; Teixeira, M.; Tsuchiya, T.; Chesnais, Q.; Mudgett, M.B.; Kaloshian, I. Aphid effector Me10 interacts with tomato TFT7, a 14-3-3 isoform involved in aphid resistance. New Phytol. 2019, 221, 1518–1528. [Google Scholar] [CrossRef] [Green Version]
- Escudero-Martinez, C.; Rodriguez, P.A.; Liu, S.; Santos, P.A.; Stephens, J.; Bos, J.I.B. An aphid effector promotes barley susceptibility through suppression of defence gene expression. J. Exp. Bot. 2020, 71, 2796–2807. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Zhang, Q.; Qin, Y.; Yin, H.; Zhang, S.; Li, Q.; Zhang, Y.; Fan, J.; Chen, J. A chromosome-level draft genome of the grain aphid Sitobion miscanthi. Giga Sci. 2019, 8, giz101. [Google Scholar] [CrossRef] [Green Version]
- Xin, J.; Shang, Q.; Desneux, N.; Gao, X. Genetic diversity of Sitobion avenae (Homoptera: Aphididae) populations from different geographic regions in China. PLoS ONE 2014, 9, e109349. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Liu, D.; Zhang, R.; Shi, X. Transcriptional responses in defense-related genes of Sitobion avenae (Hemiptera: Aphididae) feeding on wheat and barley. J. Econ. Entomol. 2019, 112, 382–395. [Google Scholar] [CrossRef]
- Zhang, Y.; Fu, Y.; Fan, J.; Li, Q.; Francis, F.; Chen, J. Comparative transcriptome and histological analyses of wheat in response to phytotoxic aphid Schizaphis graminum and non-phytotoxic aphid Sitobion avenae feeding. BMC Plant Biol. 2019, 19, 547. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, J.; Francis, F.; Chen, J. Watery saliva secreted by the grain aphid Sitobion avenae stimulates aphid resistance in wheat. J. Agric. Food Chem. 2017, 65, 8798–8805. [Google Scholar] [CrossRef]
- Zhang, Y.; Fan, J.; Sun, J.; Francis, F.; Chen, J. Transcriptome analysis of the salivary glands of the grain aphid, Sitobion avenae. Sci. Rep. 2017, 7, 15911. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fu, Y.; Francis, F.; Liu, X.; Chen, J. Insight into watery saliva proteomes of the grain aphid, Sitobion avenae. Arch. Insect Biochem. Physiol. 2021, 106, e21752. [Google Scholar] [CrossRef]
- Kamoun, S.; van West, P.; Vleeshouwers, V.G.A.A.; de Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Song, T.; Zhu, L.; Ye, W.; Wang, Y.; Shao, Y.; Dong, S.; Zhang, Z.; Dou, D.; Zheng, X.; et al. A Phytophthora sojae Glycoside Hydrolase 12 Protein Is a Major Virulence Factor during Soybean Infection and Is Recognized as a PAMP. Plant Cell 2015, 27, 2057–2072. [Google Scholar] [CrossRef] [Green Version]
- Felix, G.; Duran, J.D.; Volko, S.; Boller, T. Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J. 1999, 18, 265–276. [Google Scholar] [CrossRef]
- Chisholm, S.T.; Coaker, G.; Day, B.; Staskawicz, B.J. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 2006, 124, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Will, T.; van Bel, A.J.E. Physical and chemical interactions between aphids and plants. J. Exp. Bot. 2006, 57, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Yates, A.D.; Michel, A. Mechanisms of aphid adaptation to host plant resistance. Curr. Opin. Insect Sci. 2018, 26, 41–49. [Google Scholar] [CrossRef]
- Züst, T.; Agrawal, A.A. Mechanisms and evolution of plant resistance to aphids. Nat. Plants 2016, 2, 15206. [Google Scholar] [CrossRef]
- Yin, C.; Hulbert, S. Prospects for functional analysis of effectors from cereal rust fungi. Euphytica 2011, 179, 57–67. [Google Scholar] [CrossRef]
- Xu, Q.; Tang, C.L.; Wang, X.D.; Sun, S.T.; Zhao, J.R.; Kang, Z.S.; Wang, X.J. An effector protein of the wheat stripe rust fungus targets chloroplasts and suppresses chloroplast function. Nat. Commun. 2019, 10, 5571. [Google Scholar] [CrossRef] [Green Version]
- Pitino, M.; Hogenhout, S.A. Aphid protein effectors promote aphid colonization in a plant species-specific manner. Mol. Plant Microbe Interact. 2013, 26, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.X.; Qian, L.X.; Wang, X.W.; Shao, R.X.; Hong, Y.; Liu, S.S.; Wang, X.W. A salivary effector enables whitefly to feed on host plants by eliciting salicylic acid-signaling pathway. Proc. Natl. Acad. Sci. USA 2019, 116, 490–495. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Qian, J.; Cai, C.; Ma, Z.Z.; Li, J.H.; Yin, M.Z.; Ren, B.Y.; Shen, J. Spray method application of transdermal dsRNA delivery system for efficient gene silencing and pest control on soybean aphid Aphis glycines. J. Pest Sci. 2020, 93, 449–459. [Google Scholar] [CrossRef]
- Niu, D.; Hamby, R.; Sanchez, J.N.; Cai, Q.; Yan, Q.; Jin, H. RNAs—A new frontier in crop protection. Curr. Opin. Biotech. 2021, 70, 204–212. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, Y.H.; Li, M.S.; Chao, Z.J.; Du, X.G.; Yan, S.; Shen, J. A first greenhouse application of bacteria-expressed and nanocarrier-delivered RNA pesticide for Myzus persicae control. J. Pest Sci. 2022, 694. [Google Scholar] [CrossRef]
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Nino Sanchez, J.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 2021, 19, 1756–1768. [Google Scholar] [CrossRef]
- Abdellatef, E.; Will, T.; Koch, A.; Imani, J.; Vilcinskas, A.; Kogel, K.H. Silencing the expression of the salivary sheath protein causes transgenerational feeding suppression in the aphid Sitobion avenae. Plant Biotechnol. J. 2015, 13, 849–857. [Google Scholar] [CrossRef]
- Sun, Y.; Sparks, C.; Jones, H.; Riley, M.; Francis, F.; Du, W.; Xia, L. Silencing an essential gene involved in infestation and digestion in grain aphid through plant-mediated RNA interference generates aphid-resistant wheat plants. Plant Biotechnol. J. 2019, 17, 852–854. [Google Scholar] [CrossRef] [Green Version]
- Hood, M.; Shew, H.D. Applications of KOH-aniline blue fluorescence in the study of plant-fungal interactions. Phytopathology 1996, 86, 704–708. [Google Scholar] [CrossRef]
- Li, J.; Qian, J.; Xu, Y.; Yan, S.; Shen, J.; Yin, M. A facile-synthesized star polycation constructed as a highly efficient gene vector in pest management. ACS Sustain. Chem. Eng. 2019, 7, 6316–6322. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Aphid activities during sieve element punctures. Entomol. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Yang, C.; Pan, H.; Liu, Y.; Zhou, X. Selection of Reference Genes for Expression Analysis Using Quantitative Real-Time PCR in the Pea Aphid, Acyrthosiphon pisum (Harris) (Hemiptera, Aphidiae). PLoS ONE 2014, 9, e110454. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, X.; Fu, Y.; Crespo-Herrera, L.; Liu, H.; Wang, Q.; Zhang, Y.; Chen, J. Salivary Effector Sm9723 of Grain Aphid Sitobion miscanthi Suppresses Plant Defense and Is Essential for Aphid Survival on Wheat. Int. J. Mol. Sci. 2022, 23, 6909. https://doi.org/10.3390/ijms23136909
Zhang Y, Liu X, Fu Y, Crespo-Herrera L, Liu H, Wang Q, Zhang Y, Chen J. Salivary Effector Sm9723 of Grain Aphid Sitobion miscanthi Suppresses Plant Defense and Is Essential for Aphid Survival on Wheat. International Journal of Molecular Sciences. 2022; 23(13):6909. https://doi.org/10.3390/ijms23136909
Chicago/Turabian StyleZhang, Yong, Xiaobei Liu, Yu Fu, Leonardo Crespo-Herrera, Huan Liu, Qian Wang, Yumeng Zhang, and Julian Chen. 2022. "Salivary Effector Sm9723 of Grain Aphid Sitobion miscanthi Suppresses Plant Defense and Is Essential for Aphid Survival on Wheat" International Journal of Molecular Sciences 23, no. 13: 6909. https://doi.org/10.3390/ijms23136909