
Critical Roles of Circular RNA in Tumor Metastasis via Acting as a Sponge of miRNA/isomiR


{kind=link}
{kind=link}
{kind=link}
Abstract
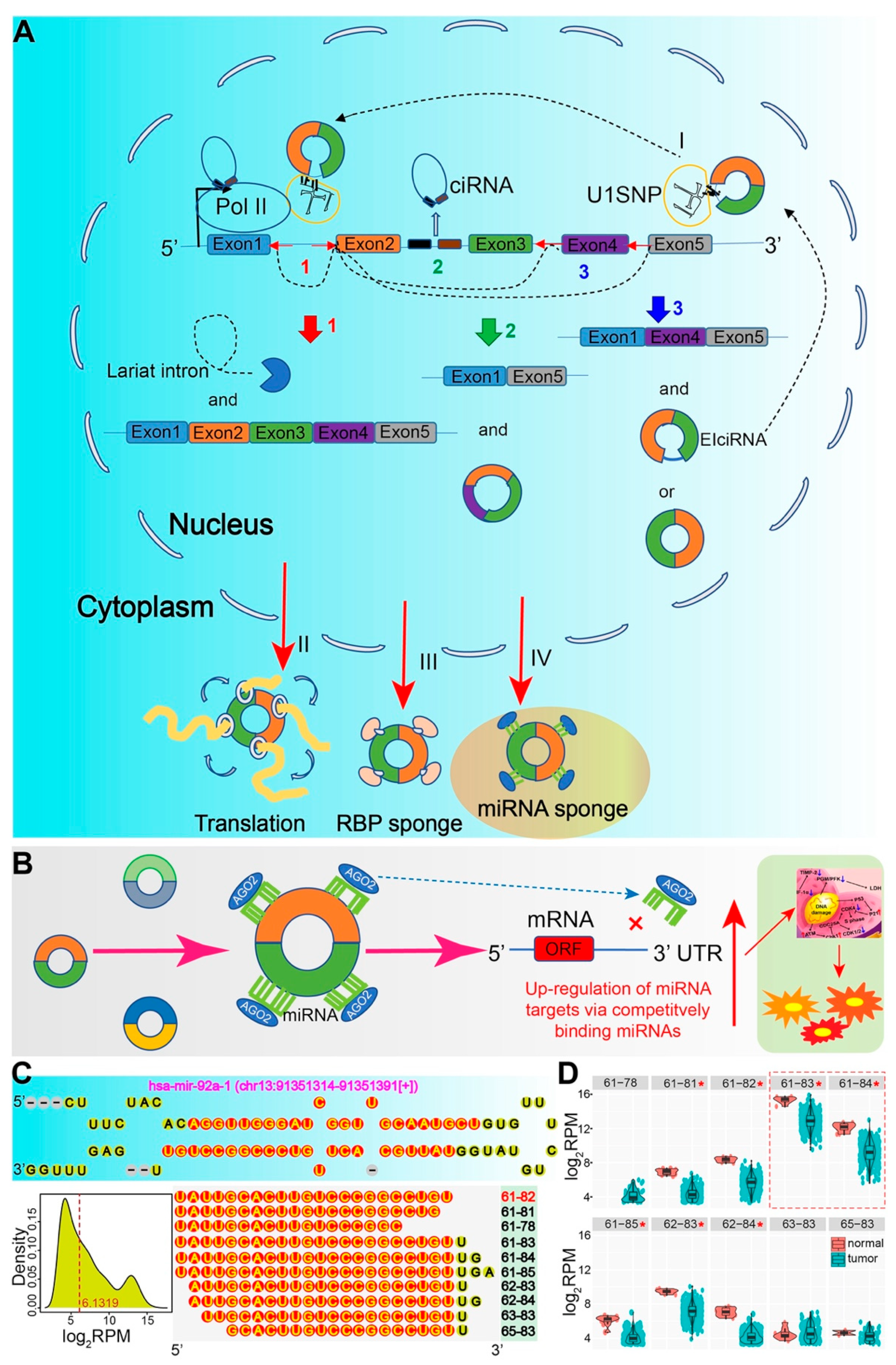
:1. Background

2. CircRNA Plays a Role in Tumorigenesis and Angiogenesis via Sponging miRNAs
2.1. CircRNA-Associated ceRNA Network in Lung Cancer
2.2. CircRNA-Associated ceRNA Network in Colorectal Cancer
2.3. CircRNA-Associated ceRNA Network in Gastric Carcinoma
2.4. CircRNA-Associated ceRNA Network in Breast Cancer
2.5. CircRNA-Associated ceRNA Network in Liver Cancer
2.6. CircRNA-Associated ceRNA Network in Esophageal Cancer
2.7. CircRNA-Associated ceRNA Network in Pancreatic Cancer
2.8. CircRNA-Associated ceRNA Network in Cervical Cancer
3. Potential Roles of circRNAs in Tumor Microenvironment through Interacting miRNAs
4. Widespread circRNA-Associated ceRNAs and Complexity of RNA Cross-Talks
5. Challenges and Future Perspectives
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Guan, X. Cancer metastases: Challenges and opportunities. Acta Pharm. Sin. B 2015, 5, 402–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Tan, G.C.; Chan, E.; Molnar, A.; Sarkar, R.; Alexieva, D.; Isa, I.M.; Robinson, S.; Zhang, S.C.; Ellis, P.; Langford, C.F.; et al. 5′ isomiR variation is of functional and evolutionary importance. Nucleic Acids Res. 2014, 42, 9424–9435. [Google Scholar] [CrossRef]
- Guo, L.; Liang, T. microRNAs and their variants in an RNA world: Implications for complex interactions and diverse roles in an RNA regulatory network. Brief. Bioinform. 2018, 19, 245–253. [Google Scholar] [CrossRef]
- Morin, R.D.; O’Connor, M.D.; Griffith, M.; Kuchenbauer, F.; Delaney, A.; Prabhu, A.L.; Zhao, Y.; McDonald, H.; Zeng, T.; Hirst, M.; et al. Application of massively parallel sequencing to microRNA profiling and discovery in human embryonic stem cells. Genome Res. 2008, 18, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Neilsen, C.T.; Goodall, G.J.; Bracken, C.P. IsomiRs—The overlooked repertoire in the dynamic microRNAome. Trends Genet. 2012, 28, 544–549. [Google Scholar] [CrossRef]
- Babapoor, S.; Fleming, E.; Wu, R.; Dadras, S.S. A novel miR-451a isomiR, associated with amelanotypic phenotype, acts as a tumor suppressor in melanoma by retarding cell migration and invasion. PLoS ONE 2014, 9, e107502. [Google Scholar] [CrossRef] [Green Version]
- Llorens, F.; Banez-Coronel, M.; Pantano, L.; del Rio, J.A.; Ferrer, I.; Estivill, X.; Marti, E. A highly expressed miR-101 isomiR is a functional silencing small RNA. BMC Genom. 2013, 14, 104. [Google Scholar] [CrossRef] [Green Version]
- Mercey, O.; Popa, A.; Cavard, A.; Paquet, A.; Chevalier, B.; Pons, N.; Magnone, V.; Zangari, J.; Brest, P.; Zaragosi, L.E.; et al. Characterizing isomiR variants within the microRNA-34/449 family. FEBS Lett. 2017, 591, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Van der Kwast, R.; Woudenberg, T.; Quax, P.H.A.; Nossent, A.Y. MicroRNA-411 and Its 5′-IsomiR Have Distinct Targets and Functions and Are Differentially Regulated in the Vasculature under Ischemia. Mol. Ther. 2020, 28, 157–170. [Google Scholar] [CrossRef]
- De Wit, E.; Linsen, S.E.; Cuppen, E.; Berezikov, E. Repertoire and evolution of miRNA genes in four divergent nematode species. Genome Res. 2009, 19, 2064–2074. [Google Scholar] [CrossRef] [Green Version]
- Nikolova, M.; Naydenov, M.; Glogovitis, I.; Apostolov, A.; Saare, M.; Boggavarapu, N.; Salumets, A.; Baev, V.; Yahubyan, G. Coupling miR/isomiR and mRNA Expression Signatures Unveils New Molecular Layers of Endometrial Receptivity. Life 2021, 11, 1391. [Google Scholar] [CrossRef]
- Guo, L.; Li, Y.; Cirillo, K.M.; Marick, R.A.; Su, Z.; Yin, X.; Hua, X.; Mills, G.B.; Sahni, N.; Yi, S.S. mi-IsoNet: Systems-scale microRNA landscape reveals rampant isoform-mediated gain of target interaction diversity and signaling specificity. Brief. Bioinform. 2021, 22, bbab091. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Annotating high confidence microRNAs using deep sequencing data. Nucleic Acids Res. 2014, 42, D68–D73. [Google Scholar] [CrossRef] [Green Version]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Gevertz, J.L.; Torquato, S. Modeling the effects of vasculature evolution on early brain tumor growth. J. Theor. Biol. 2006, 243, 517–531. [Google Scholar] [CrossRef]
- Jakobisiak, M.; Lasek, W.; Golab, J. Natural mechanisms protecting against cancer. Immunol. Lett. 2003, 90, 103–122. [Google Scholar] [CrossRef]
- Garikipati, V.N.S.; Verma, S.K.; Cheng, Z.; Liang, D.; Truongcao, M.M.; Cimini, M.; Yue, Y.; Huang, G.; Wang, C.; Benedict, C.; et al. Circular RNA CircFndc3b modulates cardiac repair after myocardial infarction via FUS/VEGF-A axis. Nat. Commun. 2019, 10, 4317. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, D.; Caponnetto, A.; Brex, D.; Mirabella, F.; Barbagallo, C.; Lauretta, G.; Morrone, A.; Certo, F.; Broggi, G.; Caltabiano, R.; et al. CircSMARCA5 Regulates VEGFA mRNA Splicing and Angiogenesis in Glioblastoma Multiforme Through the Binding of SRSF1. Cancers 2019, 11, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Guo, Y.; Chen, C.; Fan, D.; Wu, X.; Zhao, L.; Shao, B.; Sun, Z.; Ji, Z. Circ3823 contributes to growth, metastasis and angiogenesis of colorectal cancer: Involvement of miR-30c-5p/TCF7 axis. Mol. Cancer 2021, 20, 93. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Zhang, J.Y.; Gong, L.P.; Feng, Z.Y.; Wang, D.; Pan, Y.H.; Sun, L.P.; Wen, J.Y.; Chen, G.F.; Liang, J.; et al. Hypoxia-induced ebv-circLMP2A promotes angiogenesis in EBV-associated gastric carcinoma through the KHSRP/VHL/HIF1alpha/VEGFA pathway. Cancer Lett. 2022, 526, 259–272. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, Z.; Wu, B.; Sun, F. Circular RNA circFOXP1 promotes angiogenesis by regulating microRNA -127-5p/CDKN2AIP signaling pathway in osteosarcoma. Bioengineered 2021, 12, 9991–9999. [Google Scholar] [CrossRef]
- Seimiya, T.; Otsuka, M.; Iwata, T.; Shibata, C.; Tanaka, E.; Suzuki, T.; Koike, K. Emerging Roles of Exosomal Circular RNAs in Cancer. Front. Cell Dev. Biol. 2020, 8, 568366. [Google Scholar] [CrossRef]
- Yeung, K.T.; Yang, J. Epithelial-mesenchymal transition in tumor metastasis. Mol. Oncol. 2017, 11, 28–39. [Google Scholar] [CrossRef] [Green Version]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [Green Version]
- Rajgopal, S.; Fredrick, S.J.; Parvathi, V.D. CircRNAs: Insights into Gastric Cancer. Gastrointest Tumors 2021, 8, 159–168. [Google Scholar] [CrossRef]
- Global Burden of Disease Cancer, C.; Kocarnik, J.M.; Compton, K.; Dean, F.E.; Fu, W.; Gaw, B.L.; Harvey, J.D.; Henrikson, H.J.; Lu, D.; Pennini, A.; et al. Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life Years for 29 Cancer Groups from 2010 to 2019: A Systematic Analysis for the Global Burden of Disease Study 2019. JAMA Oncol. 2022, 8, 420–444. [Google Scholar] [CrossRef]
- Wadowska, K.; Bil-Lula, I.; Trembecki, L.; Sliwinska-Mosson, M. Genetic Markers in Lung Cancer Diagnosis: A Review. Int. J. Mol. Sci. 2020, 21, 4569. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Wang, X.; Du, Q.; Wu, N.; Liu, X.; Chen, Y.; Wang, X. The circRNA circP4HB promotes NSCLC aggressiveness and metastasis by sponging miR-133a-5p. Biochem. Biophys. Res. Commun. 2019, 513, 904–911. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Zheng, Y.; Jiang, Y.; Li, X.; Geng, J.; Shen, Y.; Li, Q.; Wang, X.; Zhao, C.; Chen, Y.; et al. The circRNA circPTPRA suppresses epithelial-mesenchymal transitioning and metastasis of NSCLC cells by sponging miR-96-5p. EBioMedicine 2019, 44, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Ma, W.; Ma, C. Circ_0067934 promotes non-small cell lung cancer development by regulating miR-1182/KLF8 axis and activating Wnt/beta-catenin pathway. Biomed. Pharmacother. 2020, 129, 110461. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Si, J.; Xiao, B.; Xiong, Y.; Dai, C.; Yang, Y.; Li, S.; Ma, Y. circ_0000567/miR-421/TMEM100 Axis Promotes the Migration and Invasion of Lung Adenocarcinoma and Is Associated with Prognosis. J. Cancer 2022, 13, 1540–1552. [Google Scholar] [CrossRef]
- Wei, W.; Wang, C.; Wang, L.; Zhang, J. circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8. Open Med. 2022, 17, 536–549. [Google Scholar] [CrossRef]
- Lv, C.; Hu, Y.; Zhou, X.; Zhu, Y.; Wang, J.; Zhou, F. CircRNA SOD2 motivates non-small cell lungs cancer advancement with EMT via acting as microRNA-2355-5p′s competing endogenous RNA to mediate calmodulin regulated spectrin associated proteins-2. Bioengineered 2022, 13, 5756–5768. [Google Scholar] [CrossRef]
- Ren, T.; Liu, C.; Hou, J.; Shan, F. Hsa_circ_0043265 Suppresses Proliferation, Metastasis, EMT and Promotes Apoptosis in Non-Small Cell Lung Cancer Through miR-25-3p/FOXP2 Pathway. Oncol. Targets Ther. 2020, 13, 3867–3880. [Google Scholar] [CrossRef]
- Xue, Y.B.; Ding, M.Q.; Xue, L.; Luo, J.H. CircAGFG1 sponges miR-203 to promote EMT and metastasis of non-small-cell lung cancer by upregulating ZNF281 expression. Thorac. Cancer 2019, 10, 1692–1701. [Google Scholar] [CrossRef]
- Shen, Z.; Sun, S. CircPTCH1 Promotes Migration in Lung Cancer by Regulating MYCN Expression Through miR-34c-5p. Oncol. Targets Ther. 2021, 14, 4779–4789. [Google Scholar] [CrossRef]
- Chen, C.; Yuan, W.; Zhou, Q.; Shao, B.; Guo, Y.; Wang, W.; Yang, S.; Guo, Y.; Zhao, L.; Dang, Q.; et al. N6-methyladenosine-induced circ1662 promotes metastasis of colorectal cancer by accelerating YAP1 nuclear localization. Theranostics 2021, 11, 4298–4315. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Kesharwani, P.; Alhakamy, N.A.; Siddique, H.R. Accentuating CircRNA-miRNA-Transcription Factors Axis: A Conundrum in Cancer Research. Front. Pharmacol. 2021, 12, 784801. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Liu, Y.; Li, W.T.; Li, Y.; Zhu, J.F. CircFNDC3B sequestrates miR-937-5p to derepress TIMP3 and inhibit colorectal cancer progression. Mol. Oncol. 2020, 14, 2960–2984. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, A.; Banerjee, S.; Sharma, V.; Khandelia, P. Circular RNAs: Emerging Role in Cancer Diagnostics and Therapeutics. Front. Mol. Biosci. 2020, 7, 577938. [Google Scholar] [CrossRef]
- Ameli-Mojarad, M.; Ameli-Mojarad, M.; Hadizadeh, M.; Young, C.; Babini, H.; Nazemalhosseini-Mojarad, E.; Bonab, M.A. The effective function of circular RNA in colorectal cancer. Cancer Cell Int. 2021, 21, 496. [Google Scholar] [CrossRef]
- Papatsirou, M.; Artemaki, P.I.; Karousi, P.; Scorilas, A.; Kontos, C.K. Circular RNAs: Emerging Regulators of the Major Signaling Pathways Involved in Cancer Progression. Cancers 2021, 13, 2744. [Google Scholar] [CrossRef]
- Hong, J.Y.; Zapata, J.; Blackburn, A.; Baumert, R.; Bae, S.M.; Ji, H.; Nam, H.J.; Miller, R.K.; McCrea, P.D. A catenin of the plakophilin-subfamily, Pkp3, responds to canonical-Wnt pathway components and signals. Biochem. Biophys. Res. Commun. 2021, 563, 31–39. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, J.; Shi, H.; Gao, B.; Zhou, H.; Zhang, Y.; Zhao, D.; Gao, S.; Wang, C.; Zhang, L. Hsa_circ_0026628 promotes the development of colorectal cancer by targeting SP1 to activate the Wnt/beta-catenin pathway. Cell Death Dis. 2021, 12, 802. [Google Scholar] [CrossRef]
- Yang, T.; Sun, J.; Wang, W.; Li, D.; Yang, X.; Jia, A.; Ma, Y.; Fan, Z. Hsa_circ_0006732 regulates colorectal cancer cell proliferation, invasion and EMT by miR-127-5p/RAB3D axis. Mol. Cell Biochem. 2022, 1–10. [Google Scholar] [CrossRef]
- Luo, Y.; Yao, Q. Circ_0085315 promotes cell proliferation, invasion, and migration in colon cancer through miR-1200/MAP3K1 signaling pathway. Cell Cycle 2022, 21, 1194–1211. [Google Scholar] [CrossRef]
- He, J.H.; Han, Z.P.; Luo, J.G.; Jiang, J.W.; Zhou, J.B.; Chen, W.M.; Lv, Y.B.; He, M.L.; Zheng, L.; Li, Y.G.; et al. Hsa_Circ_0007843 Acts as a mIR-518c-5p Sponge to Regulate the Migration and Invasion of Colon Cancer SW480 Cells. Front. Genet. 2020, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.Y.; Sun, D.; Xin, Y. Hsa_circ_0005230 is up-regulated and promotes gastric cancer cell invasion and migration via regulating the miR-1299/RHOT1 axis. Bioengineered 2022, 13, 5046–5063. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Sun, D.; Li, W.; Xin, Y. CircRNA_100290 promotes GC cell proliferation and invasion via the miR-29b-3p/ITGA11 axis and is regulated by EIF4A3. Cancer Cell Int. 2021, 21, 324. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chi, K.; Xiang, H.; Yang, Y. Circ_0032821 Facilitates Gastric Cancer Cell Proliferation, Migration, Invasion and Glycolysis by Regulating MiR-1236-3p/HMGB1 Axis. Cancer Manag. Res. 2020, 12, 9965–9976. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, H.; Jia, Y.; Xu, Z.; Zhang, L.; Sun, M.; Fu, J. circRNA_0005529 facilitates growth and metastasis of gastric cancer via regulating miR-527/Sp1 axis. BMC Mol. Cell Biol. 2021, 22, 6. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dai, X.; Guo, X.; Cheng, A.; Mac, S.M.; Wang, Z. Circ-OXCT1 Suppresses Gastric Cancer EMT and Metastasis by Attenuating TGF-beta Pathway Through the Circ-OXCT1/miR-136/SMAD4 Axis. Oncol. Targets Ther. 2020, 13, 3987–3998. [Google Scholar] [CrossRef]
- Wu, J.; Chen, Z.; Song, Y.; Zhu, Y.; Dou, G.; Shen, X.; Zhou, Y.; Jiang, H.; Li, J.; Peng, Y. CircRNA_0005075 suppresses carcinogenesis via regulating miR-431/p53/epithelial-mesenchymal transition axis in gastric cancer. Cell Biochem. Funct. 2020, 38, 932–942. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, Q.; Liao, B.; Qiu, X.; Hu, S.; Xu, Q. circ_0006089 promotes gastric cancer growth, metastasis, glycolysis, and angiogenesis by regulating miR-361-3p/TGFB1. Cancer Sci. 2022, 113, 2044–2055. [Google Scholar] [CrossRef]
- Zha, Q.; Wu, X.; Zhang, J.; Xu, T.; Shi, Y.; Sun, Y.; Fang, Y.; Gu, Y.; Ma, P.; Shu, Y.; et al. Hsa_circ_0007967 promotes gastric cancer proliferation through the miR-411-5p/MAML3 axis. Cell Death Discov. 2022, 8, 144. [Google Scholar] [CrossRef]
- Wang, X.; Song, Z.; Meng, Q.; Xia, S.; Wang, C.; Huang, X. Circular RNA circ_0006089 regulates the IGF1R expression by targeting miR-143-3p to promote gastric cancer proliferation, migration and invasion. Cell Cycle 2022, 1–14. [Google Scholar] [CrossRef]
- AmeliMojarad, M.; AmeliMojarad, M.; Pourmahdian, A. Circular RNA circ_0051620 sponges miR-338-3p and regulates ADAM17 to promote the gastric cancer progression. Pathol. Res. Pract. 2022, 233, 153887. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Liu, J.; Guo, X.; Cheng, A.; Deng, X.; Guo, L.; Wang, Z. Circular RNA circFGD4 suppresses gastric cancer progression via modulating miR-532-3p/APC/beta-catenin signalling pathway. Clin. Sci. 2020, 134, 1821–1839. [Google Scholar] [CrossRef] [PubMed]
- Youn, H.J.; Han, W. A Review of the Epidemiology of Breast Cancer in Asia: Focus on Risk Factors. Asian Pac. J Cancer Prev. 2020, 21, 867–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darbeheshti, F.; Mahdiannasser, M.; Noroozi, Z.; Firoozi, Z.; Mansoori, B.; Daraei, A.; Bastami, M.; Nariman-Saleh-Fam, Z.; Valipour, E.; Mansoori, Y. Circular RNA-associated ceRNA network involved in HIF-1 signalling in triple-negative breast cancer: Circ_0047303 as a potential key regulator. J. Cell Mol. Med. 2021, 25, 11322–11332. [Google Scholar] [CrossRef] [PubMed]
- Shujuan, K.; Zhongxin, L.; Jingfang, M.; Zhili, C.; Wei, W.; Liu, Q.; Li, Y. Circular RNA circ_0000518 promotes breast cancer progression through the microRNA-1225-3p/SRY-box transcription factor 4 pathway. Bioengineered 2022, 13, 2611–2622. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.L.; Fan, Y.; Liu, X.D.; Liu, W.; Zhao, M. circ_0089153 exacerbates breast cancer cells proliferation and metastasis via sponging miR-2467-3p/E2F6. Environ. Toxicol. 2022, 37, 1458–1471. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Li, J.; Li, L.; Chen, Z.; Wang, N.; Zhu, M.; Mi, H.; Xiong, Y.; Guo, G.; Gu, Y. Circular RNA circ_IRAK3 contributes to tumor growth through upregulating KIF2A via adsorbing miR-603 in breast cancer. Cancer Cell Int. 2022, 22, 81. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Lv, M.; Cao, W.; Liu, X.; Cui, J.; Wang, Y.; Wang, Y.; Nie, G.; Liu, X.; Wang, H. Circular RNA 000554 represses epithelial-mesenchymal transition in breast cancer by regulating microRNA-182/ZFP36 axis. FASEB J. 2020, 34, 11405–11420. [Google Scholar] [CrossRef]
- Qi, L.; Sun, B.; Yang, B.; Lu, S. circRNA RPPH1 Facilitates the Aggravation of Breast Cancer Development by Regulating miR-542-3p/ARHGAP1 Pathway. Cancer Biother. Radiopharm. 2021. [Google Scholar] [CrossRef]
- Tian, X.; Yang, H.; Fang, Q.; Quan, H.; Lu, H.; Wang, X. Circ_ZFR affects FABP7 expression to regulate breast cancer progression by acting as a sponge for miR-223-3p. Thorac. Cancer 2022, 13, 1369–1380. [Google Scholar] [CrossRef]
- Pan, G.; Mao, A.; Liu, J.; Lu, J.; Ding, J.; Liu, W. Circular RNA hsa_circ_0061825 (circ-TFF1) contributes to breast cancer progression through targeting miR-326/TFF1 signalling. Cell Prolif. 2020, 53, e12720. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Shen, H.; Gu, B.; Zhu, N. Circular RNA CNOT2 knockdown regulates twist family BHLH transcription factor via targeting microRNA 409-3p to prevent breast cancer invasion, migration and epithelial-mesenchymal transition. Bioengineered 2021, 12, 9058–9069. [Google Scholar] [CrossRef]
- Wang, F.; Wang, X.; Li, J.; Lv, P.; Han, M.; Li, L.; Chen, Z.; Dong, L.; Wang, N.; Gu, Y. Correction to: CircNOL10 suppresses breast cancer progression by sponging miR-767-5p to regulate SOCS2/JAK/STAT signaling. J. Biomed. Sci. 2021, 28, 31. [Google Scholar] [CrossRef]
- McGlynn, K.A.; Petrick, J.L.; El-Serag, H.B. Epidemiology of Hepatocellular Carcinoma. Hepatology 2021, 73 (Suppl. S1), 4–13. [Google Scholar] [CrossRef]
- Yang, G.; Xu, Q.; Wan, Y.; Zhang, L.; Wang, L.; Meng, F. Circ-CSPP1 knockdown suppresses hepatocellular carcinoma progression through miR-493-5p releasing-mediated HMGB1 downregulation. Cell Signal 2021, 86, 110065. [Google Scholar] [CrossRef]
- Zheng, S.; Hou, J.; Chang, Y.; Zhao, D.; Yang, H.; Yang, J. CircRNA Circ-CCND1 Aggravates Hepatocellular Carcinoma Tumorigenesis by Regulating the miR-497-5p/HMGA2 Axis. Mol. Biotechnol. 2022, 64, 178–186. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, D.; Zhu, T.; Yu, J.; Wu, X.; Lin, W.; Zhu, M.; Dai, Y.; Zhu, J. CircPUM1 promotes hepatocellular carcinoma progression through the miR-1208/MAP3K2 axis. J. Cell Mol. Med. 2021, 25, 600–612. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, Y.; Huang, Z.; Kong, Y.; Hu, X.; Xiao, W.; Quan, J.; Fan, X. CircRNA-5692 inhibits the progression of hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP expression. Cell Death Dis. 2019, 10, 900. [Google Scholar] [CrossRef]
- Zhang, B.; Li, F.; Zhu, Z.; Ding, A.; Luo, J. CircRNA CDR1as/miR-1287/Raf1 Axis Modulates Hepatocellular Carcinoma Progression Through MEK/ERK Pathway. Cancer Manag. Res. 2020, 12, 8951–8964. [Google Scholar] [CrossRef]
- Mao, Y.; Ding, Z.; Jiang, M.; Yuan, B.; Zhang, Y.; Zhang, X. Circ_0091579 exerts an oncogenic role in hepatocellular carcinoma via mediating miR-136-5p/TRIM27. Biomed. J. 2021. [Google Scholar] [CrossRef]
- Xu, G.; Zhang, P.; Liang, H.; Xu, Y.; Shen, J.; Wang, W.; Li, M.; Huang, J.; Ni, C.; Zhang, X.; et al. Circular RNA hsa_circ_0003288 induces EMT and invasion by regulating hsa_circ_0003288/miR-145/PD-L1 axis in hepatocellular carcinoma. Cancer Cell Int. 2021, 21, 212. [Google Scholar] [CrossRef]
- Liu, W.; Pan, Y.; Zhu, H.; Zhou, Y.; Zhang, H.; Liu, L.; Liu, Q.; Ji, G. CircRNA_0008194 functions as a ceRNA to promote invasion of hepatocellular carcinoma via inhibiting miR-190a/AHNAK signaling pathway. J. Clin. Lab. Anal. 2022, 36, e24286. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lei, K.; Lyu, Y.; Tan, B.; Liang, R.; Wu, D.; Wang, K.; Wang, W.; Lin, H.; Wang, M. hsa_circ_0001741 promotes esophageal squamous cell carcinoma stemness, invasion and migration by sponging miR-491-5p to upregulate NOTCH3 expression. Am. J. Cancer Res. 2022, 12, 2012–2031. [Google Scholar]
- Liang, W.; Wang, C.; Wang, J.; Zhang, M. Hsa_circ_0023984 Regulates Cell Proliferation, Migration, and Invasion in Esophageal Squamous Cancer via Regulating miR-1294/PI3K/Akt/c-Myc Pathway. Appl. Biochem. Biotechnol. 2022, 1–16. [Google Scholar] [CrossRef]
- Meng, L.; Zheng, Y.; Liu, S.; Ju, Y.; Ren, S.; Sang, Y.; Zhu, Y.; Gu, L.; Liu, F.; Zhao, Y.; et al. ZEB1 represses biogenesis of circ-DOCK5 to facilitate metastasis in esophageal squamous cell carcinoma via a positive feedback loop with TGF-beta. Cancer Lett. 2021, 519, 117–129. [Google Scholar] [CrossRef]
- Weidle, U.H.; Sela, T.; Brinkmann, U.; Niewoehner, J. Circular RNAs With Efficacy in Preclinical In Vitro and In Vivo Models of Esophageal Squamous Cell Carcinoma. Cancer Genom. Proteom. 2022, 19, 283–298. [Google Scholar] [CrossRef]
- Pan, Z.; Lin, J.; Wu, D.; He, X.; Wang, W.; Hu, X.; Zhang, L.; Wang, M. Correction for: Hsa_circ_0006948 enhances cancer progression and epithelial-mesenchymal transition through the miR-490-3p/HMGA2 axis in esophageal squamous cell carcinoma. Aging 2020, 12, 4041–4044. [Google Scholar] [CrossRef]
- Song, H.; Tian, D.; Sun, J.; Mao, X.; Kong, W.; Xu, D.; Ji, Y.; Qiu, B.; Zhan, M.; Wang, J. circFAM120B functions as a tumor suppressor in esophageal squamous cell carcinoma via the miR-661/PPM1L axis and the PKR/p38 MAPK/EMT pathway. Cell Death Dis. 2022, 13, 361. [Google Scholar] [CrossRef] [PubMed]
- Lakiotaki, E.; Kanakoglou, D.S.; Pampalou, A.; Karatrasoglou, E.A.; Piperi, C.; Korkolopoulou, P. Dissecting the Role of Circular RNAs in Sarcomas with Emphasis on Osteosarcomas. Biomedicines 2021, 9, 1642. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Chen, Y.; Chen, Y.; Xie, X.; Liang, H.; Peng, F.; Che, W. Circ_0001273 downregulation inhibits the growth, migration and glutamine metabolism of esophageal cancer cells via targeting the miR-622/SLC1A5 signaling axis. Thorac. Cancer 2022, 13, 1795–1805. [Google Scholar] [CrossRef] [PubMed]
- Menini, S.; Iacobini, C.; Vitale, M.; Pesce, C.; Pugliese, G. Diabetes and Pancreatic Cancer-A Dangerous Liaison Relying on Carbonyl Stress. Cancers 2021, 13, 313. [Google Scholar] [CrossRef] [PubMed]
- Yarmishyn, A.A.; Ishola, A.A.; Chen, C.Y.; Verusingam, N.D.; Rengganaten, V.; Mustapha, H.A.; Chuang, H.K.; Teng, Y.C.; Phung, V.L.; Hsu, P.K.; et al. Circular RNAs Modulate Cancer Hallmark and Molecular Pathways to Support Cancer Progression and Metastasis. Cancers 2022, 14, 862. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Lou, U.K.; Fung, F.K.; Tong, J.H.M.; Zhang, C.H.; To, K.F.; Chan, S.L.; Chen, Y. CircRTN4 promotes pancreatic cancer progression through a novel CircRNA-miRNA-lncRNA pathway and stabilizing epithelial-mesenchymal transition protein. Mol. Cancer 2022, 21, 10. [Google Scholar] [CrossRef]
- Ma, G.; Li, G.; Fan, W.; Xu, Y.; Song, S.; Guo, K.; Liu, Z. Circ-0005105 activates COL11A1 by targeting miR-20a-3p to promote pancreatic ductal adenocarcinoma progression. Cell Death Dis. 2021, 12, 656. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Majidpoor, J.; Shoorei, H.; Hussen, B.M.; Hadayat Jamal, H.; Baniahmad, A.; Taheri, M.; Mokhtari, M. The Interaction between Non-Coding RNAs and Calcium Binding Proteins. Front. Oncol. 2022, 12, 848376. [Google Scholar] [CrossRef]
- Shen, P.; Yang, T.; Chen, Q.; Yuan, H.; Wu, P.; Cai, B.; Meng, L.; Huang, X.; Liu, J.; Zhang, Y.; et al. CircNEIL3 regulatory loop promotes pancreatic ductal adenocarcinoma progression via miRNA sponging and A-to-I RNA-editing. Mol. Cancer 2021, 20, 51. [Google Scholar] [CrossRef]
- Xu, H.; Chen, R.; Shen, Q.; Yang, D.; Peng, H.; Tong, J.; Fu, Q. Overexpression of Circular RNA circ_0013587 Reverses Erlotinib Resistance in Pancreatic Cancer Cells Through Regulating the miR-1227/E-Cadherin Pathway. Front. Oncol. 2021, 11, 754146. [Google Scholar] [CrossRef]
- Yu, S.; Wang, M.; Zhang, H.; Guo, X.; Qin, R. Circ_0092367 Inhibits EMT and Gemcitabine Resistance in Pancreatic Cancer via Regulating the miR-1206/ESRP1 Axis. Genes 2021, 12, 1701. [Google Scholar] [CrossRef]
- Sun, Z.; Niu, S.; Xu, F.; Zhao, W.; Ma, R.; Chen, M. CircAMOTL1 Promotes Tumorigenesis Through miR-526b/SIK2 Axis in Cervical Cancer. Front. Cell Dev. Biol. 2020, 8, 568190. [Google Scholar] [CrossRef]
- Traversa, D.; Simonetti, G.; Tolomeo, D.; Visci, G.; Macchia, G.; Ghetti, M.; Martinelli, G.; Kristensen, L.S.; Storlazzi, C.T. Unravelling similarities and differences in the role of circular and linear PVT1 in cancer and human disease. Br. J. Cancer 2022, 126, 835–850. [Google Scholar] [CrossRef]
- Garlapati, P.; Ling, J.; Chiao, P.J.; Fu, J. Circular RNAs regulate cancer-related signaling pathways and serve as potential diagnostic biomarkers for human cancers. Cancer Cell Int. 2021, 21, 317. [Google Scholar] [CrossRef]
- Wise, P.M.; Neviani, P.; Riwaldt, S.; Corydon, T.J.; Wehland, M.; Braun, M.; Kruger, M.; Infanger, M.; Grimm, D. Changes in Exosomal miRNA Composition in Thyroid Cancer Cells after Prolonged Exposure to Real Microgravity in Space. Int. J. Mol. Sci. 2021, 22, 12841. [Google Scholar] [CrossRef]
- Yuan, D.D.; Jia, C.D.; Yan, M.Y.; Wang, J. Circular RNA hsa_circ_0000730 restrains cell proliferation, migration, and invasion in cervical cancer through miR-942-5p/PTEN axis. Kaohsiung J. Med. Sci. 2021, 37, 964–972. [Google Scholar] [CrossRef]
- Chi, X.; Gu, X.; Chen, S.; Shen, X. Circ_0003221 Downregulation Restrains Cervical Cancer Cell Growth, Metastasis and Angiogenesis by Governing the miR-139-3p/S100A14 Pathway. Reprod. Sci. 2022, 29, 1822–1835. [Google Scholar] [CrossRef]
- Ye, X.; Zhu, B.; Han, J.; Huang, J.; Wu, Y. Circ-0036602 Acts As a Sponge of MiR-34a-5p and MiR-431-5p to Promote Cervical Cancer Proliferation and Invasion. J. Genom. 2022, 10, 16–25. [Google Scholar] [CrossRef]
- Huang, D.; Li, C. circ-ACACA promotes proliferation, invasion, migration and glycolysis of cervical cancer cells by targeting the miR-582-5p/ERO1A signaling axis. Oncol. Lett. 2021, 22, 795. [Google Scholar] [CrossRef]
- Shen, L.; Dang, J.; Liu, S.; Xian, B.; Deng, Y.; Qu, D. CircRNA VPRBP inhibits tumorigenicity of cervical cancer via miR-93-5p/FRMD6 axis. Reprod. Sci. 2022, 1–14. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Sarfaraz, S.; Taheri, M.; Ayatollahi, S.A. Circ_CDR1as: A circular RNA with roles in the carcinogenesis. Pathol. Res. Pract. 2022, 236, 153968. [Google Scholar] [CrossRef]
- Forder, A.; Hsing, C.Y.; Trejo Vazquez, J.; Garnis, C. Emerging Role of Extracellular Vesicles and Cellular Communication in Metastasis. Cells 2021, 10, 3429. [Google Scholar] [CrossRef]
- Kinoshita, C.; Kubota, N.; Aoyama, K. Interplay of RNA-Binding Proteins and microRNAs in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 5292. [Google Scholar] [CrossRef]
- Wong, A.C.H.; Rasko, J.E.J. Splice and Dice: Intronic microRNAs, Splicing and Cancer. Biomedicines 2021, 9, 1268. [Google Scholar] [CrossRef]
- Taheri, M.; Barth, D.A.; Kargl, J.; Rezaei, O.; Ghafouri-Fard, S.; Pichler, M. Emerging Role of Non-Coding RNAs in Regulation of T-Lymphocyte Function. Front. Immunol. 2021, 12, 756042. [Google Scholar] [CrossRef]
- Wang, X.; Sheng, W.; Xu, T.; Xu, J.; Gao, R.; Zhang, Z. CircRNA hsa_circ_0110102 inhibited macrophage activation and hepatocellular carcinoma progression via miR-580-5p/PPARalpha/CCL2 pathway. Aging 2021, 13, 11969–11987. [Google Scholar] [CrossRef]
- Belanger, E.; Laprise, C. Could the Epigenetics of Eosinophils in Asthma and Allergy Solve Parts of the Puzzle? Int. J. Mol. Sci. 2021, 22, 8921. [Google Scholar] [CrossRef]
- Sas, Z.; Cendrowicz, E.; Weinhauser, I.; Rygiel, T.P. Tumor Microenvironment of Hepatocellular Carcinoma: Challenges and Opportunities for New Treatment Options. Int. J. Mol. Sci. 2022, 23, 3778. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, S.; Liao, X.; Li, M.; Chen, S.; Li, X.; Wu, X.; Yang, M.; Tang, M.; Hu, Y.; et al. Circular RNA circIKBKB promotes breast cancer bone metastasis through sustaining NF-kappaB/bone remodeling factors signaling. Mol. Cancer 2021, 20, 98. [Google Scholar] [CrossRef]
- Bolha, L.; Ravnik-Glavac, M.; Glavac, D. Circular RNAs: Biogenesis, Function, and a Role as Possible Cancer Biomarkers. Int. J. Genom. 2017, 2017, 6218353. [Google Scholar] [CrossRef]
- Wortzel, I.; Dror, S.; Kenific, C.M.; Lyden, D. Exosome-Mediated Metastasis: Communication from a Distance. Dev. Cell 2019, 49, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Harper, K.L.; Mottram, T.J.; Whitehouse, A. Insights into the Evolving Roles of Circular RNAs in Cancer. Cancers 2021, 13, 4180. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wang, Q.; Shen, J.; Yang, B.B.; Ding, X. Circbank: A comprehensive database for circRNA with standard nomenclature. RNA Biol. 2019, 16, 899–905. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zheng, L.; Zhao, Y.; Wang, Q. Profiling and Bioinformatic Analyses Indicate Differential circRNA and miRNA/isomiR Expression and Interactions. Biomed. Res. Int. 2018, 2018, 8518563. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Han, L.; Guo, L. Rewired functional regulatory networks among miRNA isoforms (isomiRs) from let-7 and miR-10 gene families in cancer. Comput. Struct. Biotechnol. J. 2020, 18, 1238–1248. [Google Scholar] [CrossRef] [PubMed]
- Zhi, Q.; Wan, D.; Ren, R.; Xu, Z.; Guo, X.; Han, Y.; Liu, F.; Xu, Y.; Qin, L.; Wang, Y. Circular RNA profiling identifies circ102049 as a key regulator of colorectal liver metastasis. Mol. Oncol. 2021, 15, 623–641. [Google Scholar] [CrossRef]
- Yang, J.H.; Li, J.H.; Shao, P.; Zhou, H.; Chen, Y.Q.; Qu, L.H. starBase: A database for exploring microRNA-mRNA interaction maps from Argonaute CLIP-Seq and Degradome-Seq data. Nucleic Acids Res. 2011, 39, D202–D209. [Google Scholar] [CrossRef] [Green Version]
- Li, J.H.; Liu, S.; Zhou, H.; Qu, L.H.; Yang, J.H. starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014, 42, D92–D97. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- Henry, N.L.; Hayes, D.F. Cancer biomarkers. Mol. Oncol. 2012, 6, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Expert consensus document: Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting circular RNAs as a therapeutic approach: Current strategies and challenges. Signal Transduct. Target Ther. 2021, 6, 185. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Xue, W.; Zhang, L.; Yang, L.Z.; Cao, S.M.; Lei, Y.N.; Liu, C.X.; Guo, S.K.; Shan, L.; et al. Screening for functional circular RNAs using the CRISPR-Cas13 system. Nat. Methods 2021, 18, 51–59. [Google Scholar] [CrossRef]
- Karedath, T.; Al-Dasim, F.M.; Ahmed, I.; Al-Qurashi, A.; Raza, A.; Andrews, S.S.; Ahmed, A.A.; Ali Mohamoud, Y.; Dermime, S.; Malek, J.A. Regulation of Circular RNA CircNFATC3 in Cancer Cells Alters Proliferation, Migration, and Oxidative Phosphorylation. Front. Cell Dev. Biol. 2021, 9, 595156. [Google Scholar] [CrossRef]
- Muller, S.; Appel, B. In vitro circularization of RNA. RNA Biol. 2017, 14, 1018–1027. [Google Scholar] [CrossRef]
- Mavros, M.N.; Economopoulos, K.P.; Alexiou, V.G.; Pawlik, T.M. Treatment and Prognosis for Patients With Intrahepatic Cholangiocarcinoma: Systematic Review and Meta-analysis. JAMA Surg. 2014, 149, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.G.; Awan, F.M.; Du, W.W.; Zeng, Y.; Lyu, J.; Wu, G.S.; Yang, W.; Yang, B.B. The Circular RNA Interacts with STAT3, Increasing Its Nuclear Translocation and Wound Repair by Modulating Dnmt3a and miR-17 Function. Mol. Ther. 2017, 25, 2062–2074. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Jia, L.; Luo, L.; Xu, X.; Xiang, Y.; Ren, Y.; Ren, D.; Shen, L.; Liang, T. Critical Roles of Circular RNA in Tumor Metastasis via Acting as a Sponge of miRNA/isomiR. Int. J. Mol. Sci. 2022, 23, 7024. https://doi.org/10.3390/ijms23137024
Guo L, Jia L, Luo L, Xu X, Xiang Y, Ren Y, Ren D, Shen L, Liang T. Critical Roles of Circular RNA in Tumor Metastasis via Acting as a Sponge of miRNA/isomiR. International Journal of Molecular Sciences. 2022; 23(13):7024. https://doi.org/10.3390/ijms23137024
Chicago/Turabian StyleGuo, Li, Lin Jia, Lulu Luo, Xinru Xu, Yangyang Xiang, Yujie Ren, Dekang Ren, Lulu Shen, and Tingming Liang. 2022. "Critical Roles of Circular RNA in Tumor Metastasis via Acting as a Sponge of miRNA/isomiR" International Journal of Molecular Sciences 23, no. 13: 7024. https://doi.org/10.3390/ijms23137024