
Derivation and Characterization of Endothelial Cells from Porcine Induced Pluripotent Stem Cells


Abstract
:1. Introduction
2. Results
2.1. Generation and Characterization of piPSC-ECs
2.2. In Vitro and In Vivo Angiogenesis of the Derived piPSC-ECs
2.3. Genome-Wide Transcriptional Profile Analysis for the Regulation of piPSC-EC Differentiation
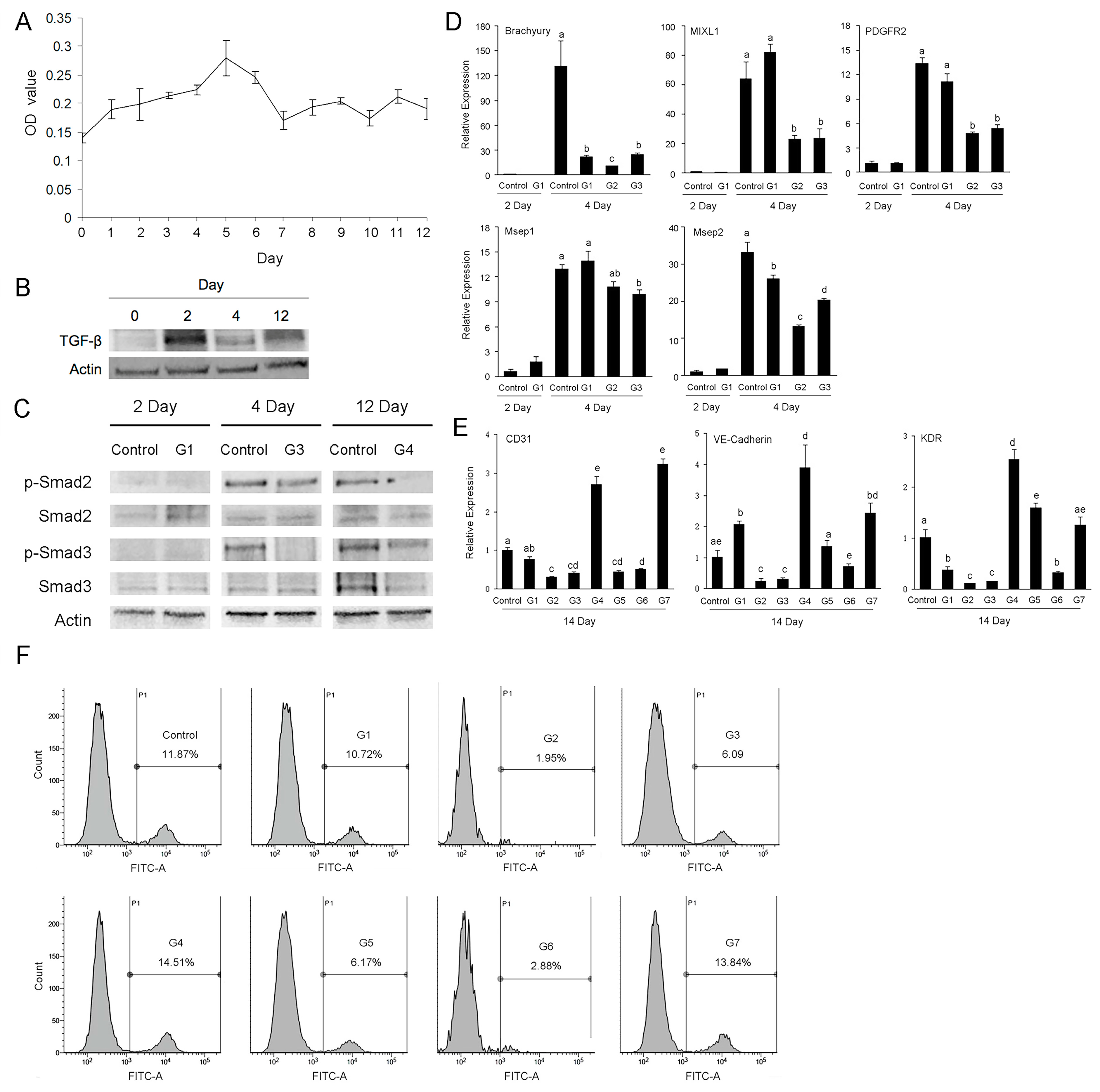
2.4. Effects of TGF-β on the Differentiation of Porcine iPSC-ECs
3. Discussion
4. Materials and Methods
4.1. Differentiation of ECs from piPSCs
4.2. In Vitro Tube Formation Assay
4.3. Uptake of Dil-Ac-LDL
4.4. Immunofluorescence Staining
4.5. Flow Cytometric Analysis
4.6. Cell Culture on 3D Scaffold
4.7. Mouse Ischemic Hindlimb Model and Cell Transplantation
4.8. Histological Analysis
4.9. RNA Sequencing
4.10. High-Throughput Data Analysis
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Western Blotting
4.13. Quantitative Real Time-PCR (qRT-PCR)
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990–2019: Update from the GBD 2019 Study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef] [PubMed]
- Furchgott, R.F.; Zawadzki, J.V. The obligatory role of endothelial cells in the relaxation of arterial smooth muscle by acetylcholine. Nature 1980, 288, 373–376. [Google Scholar] [CrossRef]
- Pober, J.S.; Sessa, W.C. Evolving functions of endothelial cells in inflammation. Nat. Rev. Immunol. 2007, 7, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef] [PubMed]
- Losordo, D.W.; Kibbe, M.R.; Mendelsohn, F.; Marston, W.; Driver, V.R.; Sharafuddin, M.; Teodorescu, V.; Wiechmann, B.N.; Thompson, C.; Kraiss, L.; et al. A Randomized, Controlled Pilot Study of Autologous CD34+ Cell Therapy for Critical Limb Ischemia. Circ. Cardiovasc. Interv. 2012, 5, 821–830. [Google Scholar] [CrossRef] [Green Version]
- Madeddu, P.; Emanueli, C.; Spillmann, F.; Meloni, M.; Bouby, N.; Richer, C.; Alhenc-Gelas, F.; Van Weel, V.; Eefting, D.; Quax, P.; et al. Murine models of myocardial and limb ischemia: Diagnostic end-points and relevance to clinical problems. Vasc. Pharmacol. 2006, 45, 281–301. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of Pluripotent Stem Cells from Mouse Embryonic and Adult Fibroblast Cultures by Defined Factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Taura, D.; Sone, M.; Homma, K.; Oyamada, N.; Takahashi, K.; Tamura, N.; Yamanaka, S.; Nakao, K. Induction and Isolation of Vascular Cells from Human Induced Pluripotent Stem Cells—Brief Report. Arter. Thromb. Vasc. Biol. 2009, 29, 1100–1103. [Google Scholar] [CrossRef] [Green Version]
- Choi, K.D.; Yu, J.; Smuga-Otto, K.; Salvagiotto, G.; Rehrauer, W.; Vodyanik, M.; Thomson, J.; Slukvin, I. Hematopoietic and endothelial differentiation of human induced pluripotent stem cells. Stem Cells 2009, 27, 559–567. [Google Scholar] [CrossRef]
- Risau, W.; Flamme, I. Vasculogenesis. Annu. Rev. Cell Dev. Biol. 1995, 11, 73–91. [Google Scholar] [CrossRef]
- Kimelman, D.; Griffin, K.J. Vertebrate mesendoderm induction and patterning. Curr. Opin. Genet. Dev. 2000, 10, 350–356. [Google Scholar] [CrossRef]
- Zorn, A.M.; Wells, J.M. Molecular Basis of Vertebrate Endoderm Development. Int. Rev. Cytol. 2007, 259, 49–111. [Google Scholar] [PubMed]
- Dyer, M.; Farrington, S.; Mohn, D.; Munday, J.; Baron, M. Indian hedgehog activates hematopoiesis and vasculogenesis and can respecify prospective neurectodermal cell fate in the mouse embryo. Development 2001, 128, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Flamme, I.; Breier, G.; Risau, W. Vascular Endothelial Growth Factor (VEGF) and VEGF Receptor 2(flk-1) Are Expressed during Vasculogenesis and Vascular Differentiation in the Quail Embryo. Dev. Biol. 1995, 169, 699–712. [Google Scholar] [CrossRef]
- Banerjee, S.; Dhara, S.K.; Bacanamwo, M. Endoglin is a novel endothelial cell specification gene. Stem Cell Res. 2012, 8, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.Y.; Sriram, G.; Rufaihah, A.J.; Neoh, K.G.; Cao, T. Efficient Derivation of Lateral Plate and Paraxial Mesoderm Subtypes from Human Embryonic Stem Cells Through GSKi-Mediated Differentiation. Stem Cells Dev. 2013, 22, 1893–1906. [Google Scholar] [CrossRef] [Green Version]
- Orlova, V.V.; Hil, F.E.V.D.; Petrus-Reurer, S.; Drabsch, Y.; Dijke, P.T.; Mummery, C.L. Generation, expansion and functional analysis of endothelial cells and pericytes derived from human pluripotent stem cells. Nat. Protoc. 2014, 9, 1514–1531. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.-Y.; Lin, B.; Kim, J.; Sullivan, M.; Tobita, K.; Salama, G.; Yang, L. Repopulation of decellularized mouse heart with human induced pluripotent stem cell-derived cardiovascular progenitor cells. Nat. Commun. 2013, 4, 2307. [Google Scholar] [CrossRef]
- James, D.; Nam, H.S.; Seandel, M.; Nolan, D.; Janovitz, T.; Tomishima, M.; Studer, L.; Lee, G.; Lyden, D.; Benezra, R.; et al. Expansion and maintenance of human embryonic stem cell-derived endothelial cells by TGFbeta inhibition is Id1 dependent. Nat. Biotechnol. 2010, 28, 161–166. [Google Scholar] [CrossRef]
- Liang, J.; Huang, W.; Cai, W.; Wang, L.; Guo, L.; Paul, C.; Yu, X.-Y.; Wang, Y. Inhibition of microRNA-495 Enhances Therapeutic Angiogenesis of Human Induced Pluripotent Stem Cells. Stem Cells 2016, 35, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.L.; Hartvigsen, M.L.; Hedemann, M.S.; Lærke, H.N.; Hermansen, K.; Knudsen, K.E.B. Similar metabolic responses in pigs and humans to breads with different contents and compositions of dietary fibers: A metabolomics study. Am. J. Clin. Nutr. 2014, 99, 941–949. [Google Scholar] [CrossRef]
- Wernersson, R.; Schierup, M.H.; Jørgensen, F.G.; Gorodkin, J.; Panitz, F.; Stærfeldt, H.-H.; Christensen, O.F.; Mailund, T.; Hornshøj, H.; Klein, A.; et al. Pigs in sequence space: A 0.66X coverage pig genome survey based on shotgun sequencing. BMC Genom. 2005, 6, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunney, J.K. Advances in Swine Biomedical Model Genomics. Int. J. Biol. Sci. 2007, 3, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Pankajakshan, D.; Kansal, V.; Agrawal, D.K. In vitro differentiation of bone marrow derived porcine mesenchymal stem cells to endothelial cells. J. Tissue Eng. Regen. Med. 2013, 7, 911–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colazzo, F.; Chester, A.H.; Taylor, P.M.; Yacoub, M.H. Induction of mesenchymal to endothelial transformation of adipose-derived stem cells. J. Heart Valve Dis. 2010, 19, 736–744. [Google Scholar] [PubMed]
- Gu, M.; Nguyen, P.K.; Lee, A.S.; Xu, D.; Hu, S.; Plews, J.R.; Han, L.; Huber, B.; Lee, W.H.; Gong, Y.; et al. Microfluidic Single-Cell Analysis Shows That Porcine Induced Pluripotent Stem Cell–Derived Endothelial Cells Improve Myocardial Function by Paracrine Activation. Circ. Res. 2012, 111, 882–893. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Lv, J.; Li, X.; Li, Y.; Xu, Q.; Jin, J.; Zhang, Y.; Liu, Z. Derivation of endothelial cells from porcine induced pluripotent stem cells by optimized single layer culture system. J. Veter-Sci. 2020, 21, e9. [Google Scholar] [CrossRef]
- Li, X.; Yu, Y.; Wei, R.; Li, Y.; Lv, J.; Liu, Z.; Zhang, Y. In vitro and in vivo study on angiogenesis of porcine induced pluripotent stem cell-derived endothelial cells. Differentiation 2021, 120, 10–18. [Google Scholar] [CrossRef]
- Noren, D.P.; Chou, W.H.; Lee, S.H.; Qutub, A.A.; Warmflash, A.; Wagner, D.S.; Popel, A.S.; Levchenko, A. Endothelial cells decode VEGF-mediated Ca2+ signaling patterns to produce distinct functional responses. Sci. Signal. 2016, 9, ra20. [Google Scholar] [CrossRef] [Green Version]
- Moccia, F.; Negri, S.; Shekha, M.; Faris, P.; Guerra, G. Endothelial Ca2+ Signaling, Angiogenesis and Vasculogenesis: Just What It Takes to Make a Blood Vessel. Int. J. Mol. Sci. 2019, 20, 3962. [Google Scholar] [CrossRef] [Green Version]
- Savage, A.M.; Kurusamy, S.; Chen, Y.; Jiang, Z.; Chhabria, K.; Macdonald, R.B.; Kim, H.R.; Wilson, H.L.; Van Eeden, F.J.M.; Armesilla, A.L.; et al. tmem33 is essential for VEGF-mediated endothelial calcium oscillations and angiogenesis. Nat. Commun. 2019, 10, 732. [Google Scholar] [CrossRef] [PubMed]
- Pereda, J.; Royo, D.; Gabasa, M.; Pujols, L.; Molina-Molina, M. Myofibroblast cell transition induced by TGF-b1 implies an altered arachidonic acid metabolism in human lung. In Proceedings of the American Thoracic Society 2010 International Conference, New Orleans, LA, USA, 14–19 May 2010. [Google Scholar]
- Luo, K. Signaling Cross Talk between TGF-β/Smad and Other Signaling Pathways. Cold Spring Harb. Perspect. Biol. 2016, 9, a022137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armulik, A.; Genové, G.; Betsholtz, C. Pericytes: Developmental, physiological, and pathological perspectives, problems, and promises. Dev. Cell 2011, 21, 193–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watabe, T.; Nishihara, A.; Mishima, K.; Yamashita, J.; Shimizu, K.; Miyazawa, K.; Nishikawa, S.; Miyazono, K. TGF-beta receptor kinase inhibitor enhances growth and integrity of embryonic stem cell-derived endothelial cells. J. Cell Biol. 2003, 163, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Aoki, H.; Yamashita, M.; Hashita, T.; Ogami, K.; Hoshino, S.; Iwao, T.; Matsunaga, T. Efficient differentiation and purification of human induced pluripotent stem cell-derived endothelial progenitor cells and expansion with the use of inhibitors of ROCK, TGF-β, and GSK3β. Heliyon 2020, 6, e03493. [Google Scholar] [CrossRef]
- Bai, H.; Xie, Y.-L.; Gao, Y.-X.; Cheng, T.; Wang, Z.Z. The Balance of Positive and Negative Effects of TGF-β Signaling Regulates the Development of Hematopoietic and Endothelial Progenitors in Human Pluripotent Stem Cells. Stem Cells Dev. 2013, 22, 2765–2776. [Google Scholar] [CrossRef] [Green Version]
- Inman, G.J.; Nicolás, F.J.; Callahan, J.F.; Harling, J.D.; Gaster, L.M.; Reith, A.D.; Laping, N.J.; Hill, C.S. SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol. Pharmacol. 2002, 62, 65–74. [Google Scholar] [CrossRef]
- Hockemeyer, D.; Jaenisch, R. Induced Pluripotent Stem Cells Meet Genome Editing. Cell Stem Cell 2016, 18, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Godini, R.; Lafta, H.Y.; Fallahi, H. Epigenetic modifications in the embryonic and induced pluripotent stem cells. Gene Expr. Patterns 2018, 29, 1–9. [Google Scholar] [CrossRef]
- Adams, W.; Zhang, Y.; Cloutier, J.; Kuchimanchi, P.; Newton, G.; Sehrawat, S.; Aird, W.C.; Mayadas, T.N.; Luscinskas, F.; García-Cardeña, G. Functional Vascular Endothelium Derived from Human Induced Pluripotent Stem Cells. Stem Cell Rep. 2013, 1, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.-J.; Lee, N.; Lee, J.Y.; Choi, Y.J.; Ii, M.; Wecker, A.; Jeong, J.-O.; Curry, C.; Qin, G.; Yoon, Y.-S. Role of host tissues for sustained humoral effects after endothelial progenitor cell transplantation into the ischemic heart. J. Exp. Med. 2007, 204, 3257–3269. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Wang, Q.-L.; Zhou, P.; Wang, G.-D.; Tan, Y.-Z.; Wang, H.-J. Thymosin β4 promotes the survival and angiogenesis of transplanted endothelial progenitor cells in the infarcted myocardium. Int. J. Mol. Med. 2017, 39, 1347–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, T.S.; Bhutto, I.; Zimmerlin, L.; Huo, J.S.; Nagaria, P.; Miller, D.; Rufaihah, A.J.; Talbot, C.; Aguilar, J.; Grebe, R.; et al. Vascular progenitors from cord blood-derived induced pluripotent stem cells possess augmented capacity for regenerating ischemic retinal vasculature. Circulation 2014, 129, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayton, Z.E.; Yuen, G.S.; Sadeghipour, S.; Hywood, J.D.; Wong, J.W.; Huang, N.F.; Ng, M.K.; Cooke, J.P.; Patel, S. A comparison of the pro-angiogenic potential of human induced pluripotent stem cell derived endothelial cells and induced endothelial cells in a murine model of peripheral arterial disease. Int. J. Cardiol. 2017, 234, 81–89. [Google Scholar] [CrossRef]
- Lai, R.C.; Tan, S.S.; Teh, B.J.; Sze, S.K.; Arslan, F.; De Kleijn, D.P.; Choo, A.; Lim, S.K. Proteolytic Potential of the MSC Exosome Proteome: Implications for an Exosome-Mediated Delivery of Therapeutic Proteasome. Int. J. Proteom. 2012, 2012, 971907. [Google Scholar] [CrossRef] [Green Version]
- Umezu, T.; Tadokoro, H.; Azuma, K.; Yoshizawa, S.; Ohyashiki, K.; Ohyashiki, J.H. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1. Blood 2014, 124, 3748–3757. [Google Scholar] [CrossRef]
- David, C.J.; Massagué, J. Contextual determinants of TGFβ action in development, immunity and cancer. Nat. Rev. Mol. Cell Biol. 2018, 19, 419–435. [Google Scholar] [CrossRef]
- Poon, E.; Clermont, F.; Firpo, M.T.; Akhurst, R.J. TGFbeta inhibition of yolk-sac-like differentiation of human embryonic stem-cell-derived embryoid bodies illustrates differences between early mouse and human development. J. Cell Sci. 2006, 119, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Dickson, M.; Martin, J.; Cousins, F.; Kulkarni, A.; Karlsson, S.; Akhurst, R. Defective haematopoiesis and vasculogenesis in transforming growth factor-beta 1 knock out mice. Development 1995, 121, 1845–1854. [Google Scholar] [CrossRef]
- Cunha, S.I.; Pardali, E.; Thorikay, M.; Anderberg, C.; Hawinkels, L.; Goumans, M.-J.; Seehra, J.; Heldin, C.-H.; Dijke, P.T.; Pietras, K. Genetic and pharmacological targeting of activin receptor-like kinase 1 impairs tumor growth and angiogenesis. J. Exp. Med. 2010, 207, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.-J.; Kim, Y.H.; Choe, S.-W.; Tak, Y.G.; Garrido-Martin, E.M.; Chang, M.; Lee, Y.J.; Oh, S.P. Enhanced Responses to Angiogenic Cues Underlie the Pathogenesis of Hereditary Hemorrhagic Telangiectasia 2. PLoS ONE 2013, 8, e63138. [Google Scholar]
- Crick, S.J.; Sheppard, M.N.; Ho, S.Y.; Gebstein, L.; Anderson, R.H. Anatomy of the pig heart: Comparisons with normal human cardiac structure. J. Anat. 1998, 193, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Lingmei, Q.; Shen, Y.; Qian, L.; Kong, X.; Chen, M.; Cao, K.; Zhang, F. Transplantation of iPSc Restores Cardiac Function by Promoting Angiogenesis and Ameliorating Cardiac Remodeling in a Post-infarcted Swine Model. Cell Biophys. 2014, 71, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Liu, H.; Yang, M.; Bai, Y.; Zhang, B.; Wang, C.; Yan, Z.; Niu, G.; Zou, Y.; Li, Y. Bone marrow mesenchymal stem cell-derived endothelial cells increase capillary density and accelerate angiogenesis in mouse hindlimb ischemia model. Stem Cell Res. Ther. 2020, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Blankenberg, D.; Von Kuster, G.; Coraor, N.; Ananda, G.; Lazarus, R.; Mangan, M.; Nekrutenko, A.; Taylor, J. Galaxy: A Web-Based Genome Analysis Tool for Experimentalists. Curr. Protoc. Mol. Biol. 2010, 89, 19.10.1–19.10.21. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; A Lempicki, R. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2008, 4, 44–57. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group Name | Treatment Period | ||
|---|---|---|---|
| 0–2 Day | 3–4 Day | 5–12 Day | |
| Control | − | − | − |
| G1 | + | − | − |
| G2 | + | + | − |
| G3 | − | + | − |
| G4 | − | − | + |
| G5 | − | + | + |
| G6 | + | + | + |
| G7 | + | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Li, X.; Li, Y.; Wei, R.; Li, H.; Liu, Z.; Zhang, Y. Derivation and Characterization of Endothelial Cells from Porcine Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2022, 23, 7029. https://doi.org/10.3390/ijms23137029
Yu Y, Li X, Li Y, Wei R, Li H, Liu Z, Zhang Y. Derivation and Characterization of Endothelial Cells from Porcine Induced Pluripotent Stem Cells. International Journal of Molecular Sciences. 2022; 23(13):7029. https://doi.org/10.3390/ijms23137029
Chicago/Turabian StyleYu, Yang, Xuechun Li, Yimei Li, Renyue Wei, Hai Li, Zhonghua Liu, and Yu Zhang. 2022. "Derivation and Characterization of Endothelial Cells from Porcine Induced Pluripotent Stem Cells" International Journal of Molecular Sciences 23, no. 13: 7029. https://doi.org/10.3390/ijms23137029