
Conjugation of the 9-kDa Isoform of Granulysin with Liposomes Potentiates Its Cytotoxicity

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
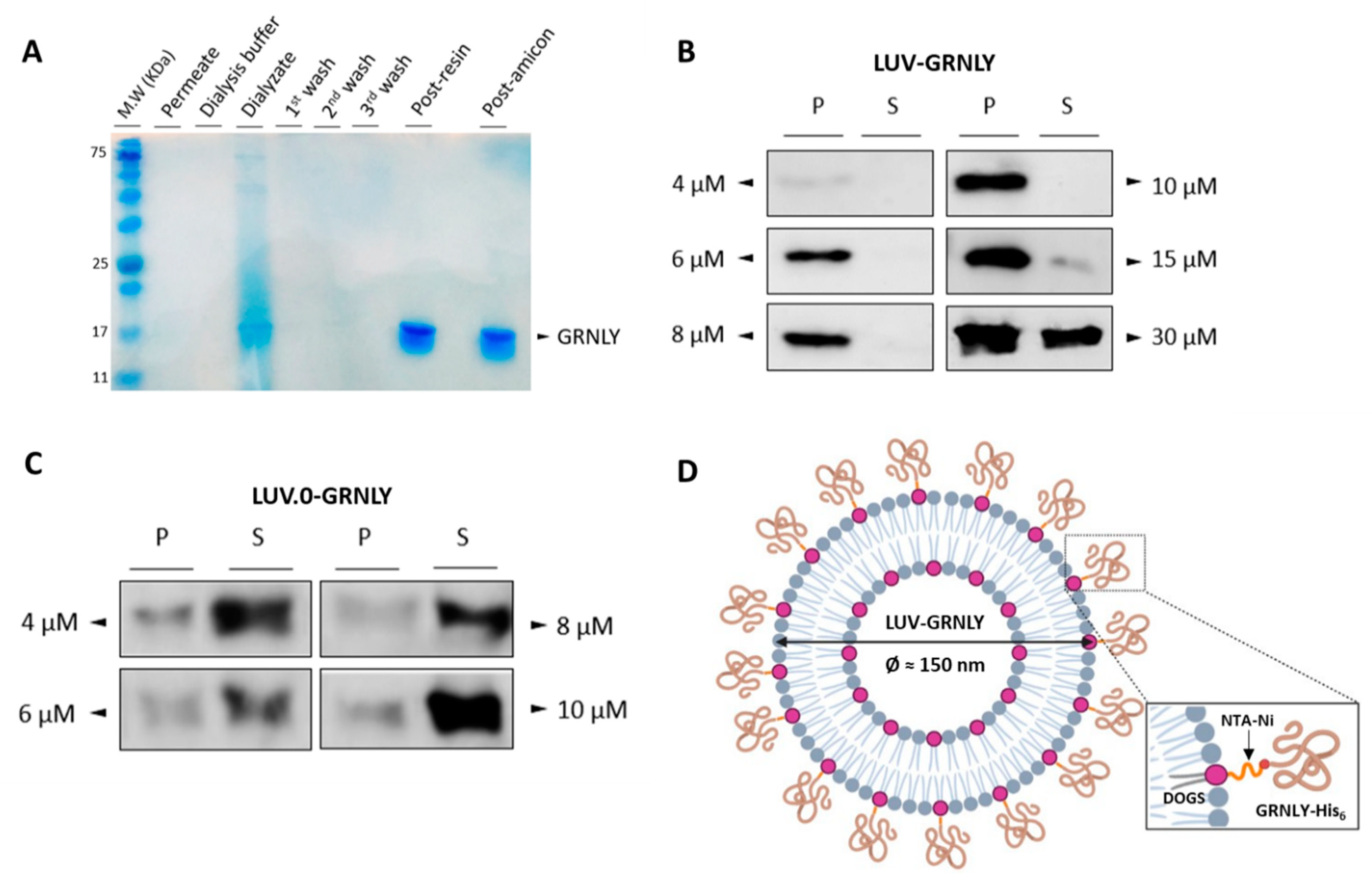
2.1. Production and Purification of the Recombinant 9 kDa GRNLY in Pichia Pastoris
2.2. GRNLY Binds Efficiently to LUV Surface through the DOGS-NTA-Ni Lipid
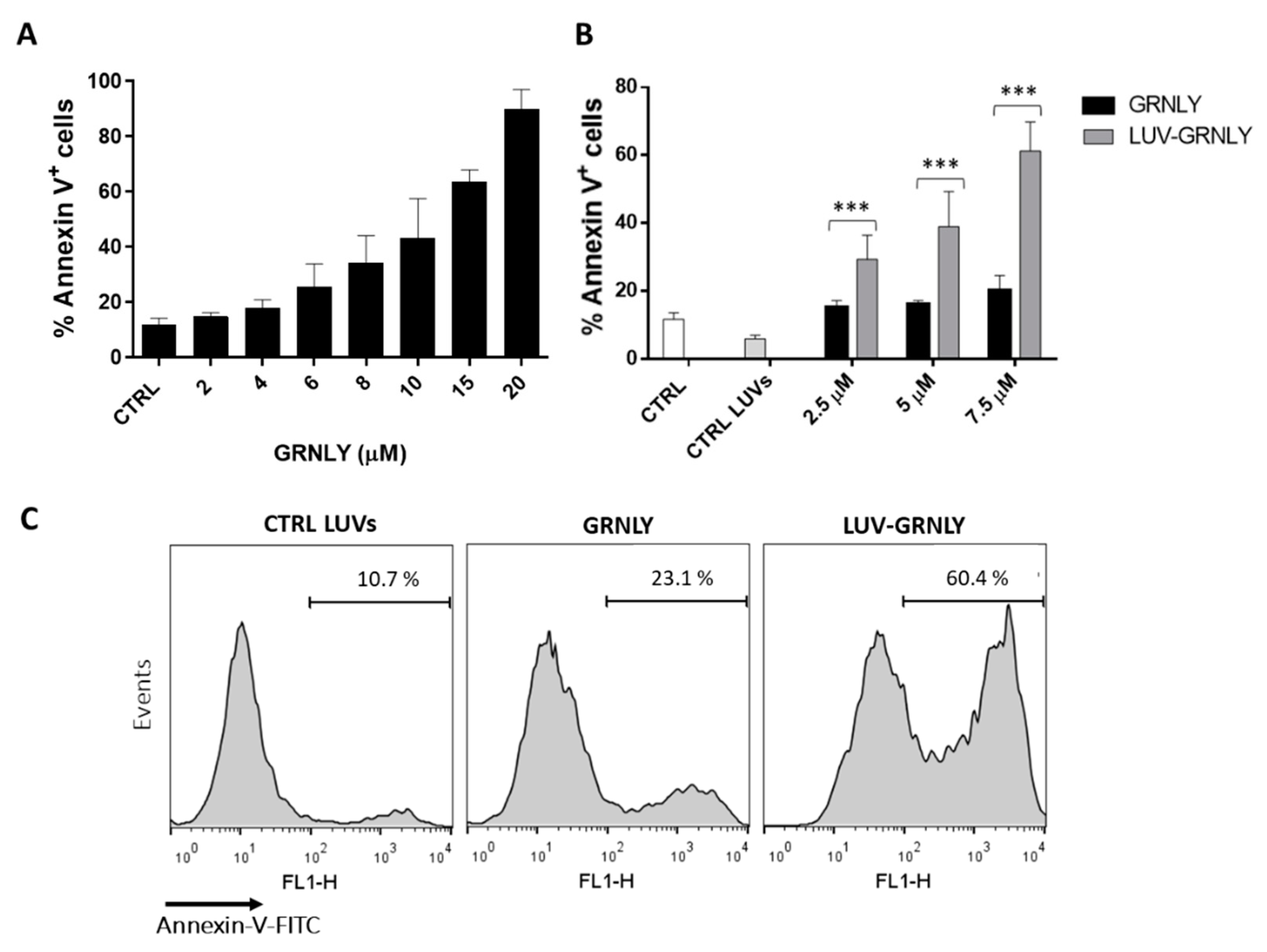
2.3. In Vitro Cytotoxic Effect of LUV-GRNLY on the Jurkat Cell Line
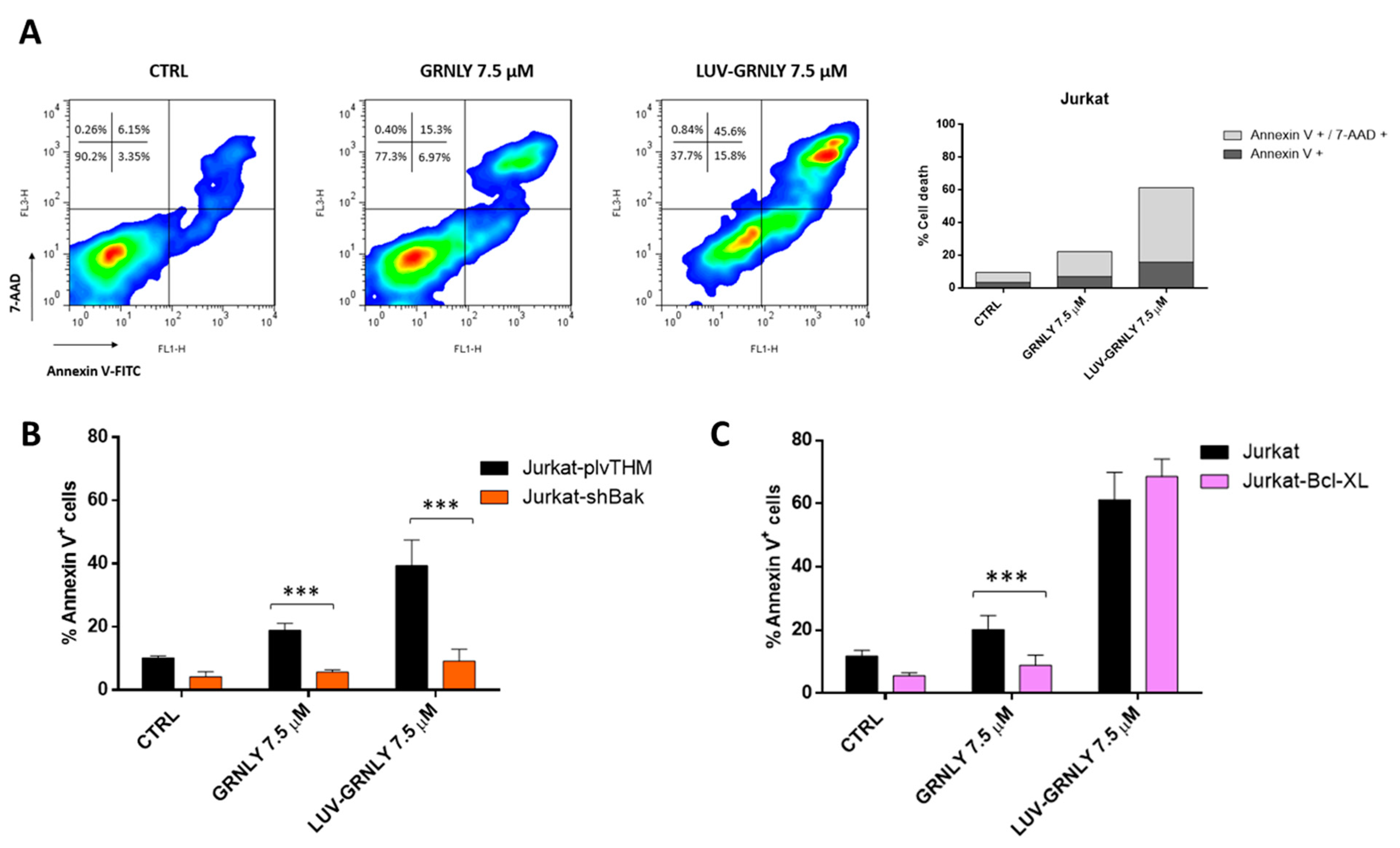
2.4. Analysis of the Mechanism of Cell Death Induced by GRNLY and LUV-GRNLY in Jurkat Cells
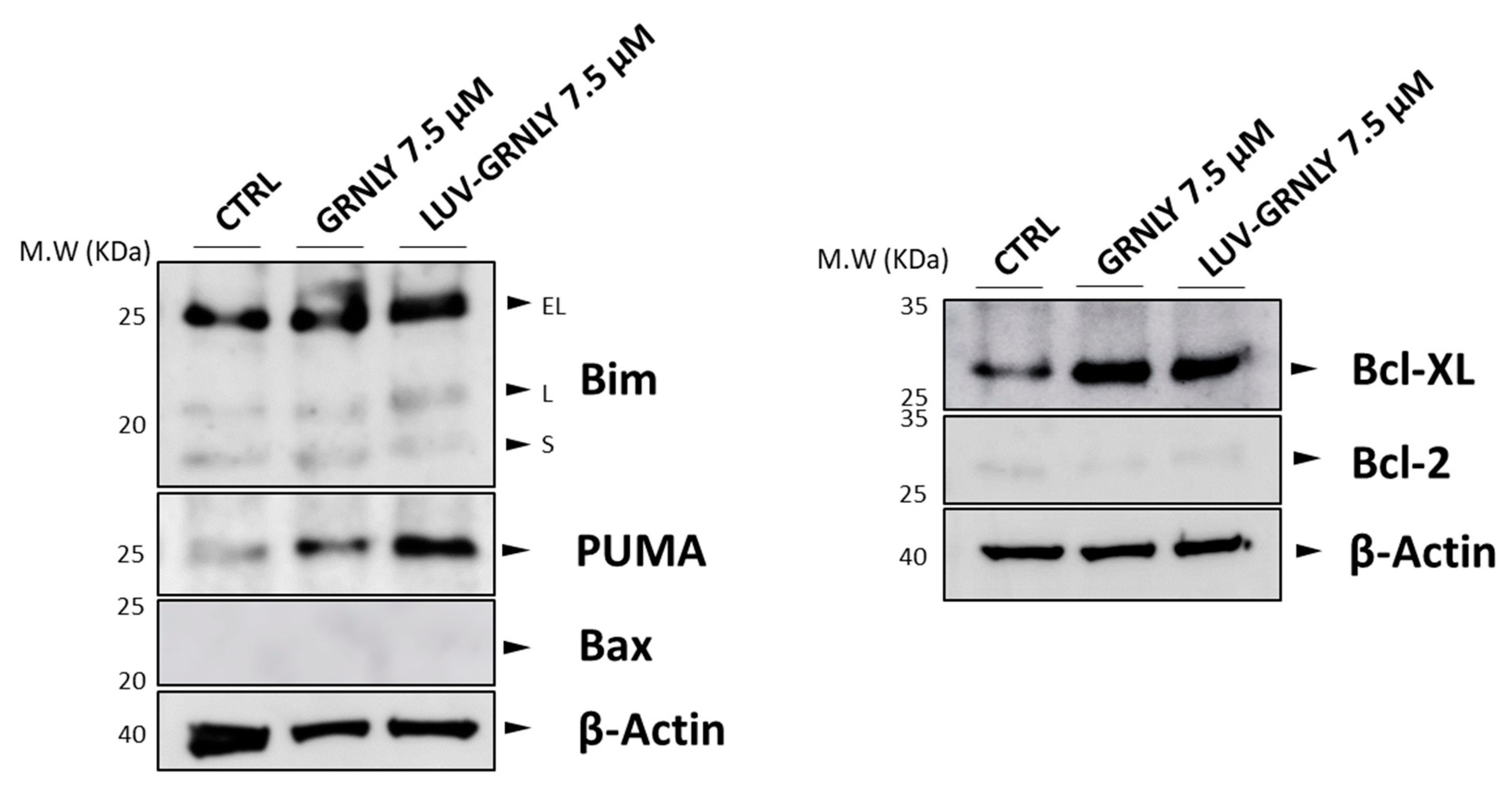
2.5. GRNLY and LUV-GRNLY Treatments Increase PUMA, Bim and Bcl-xL Expression
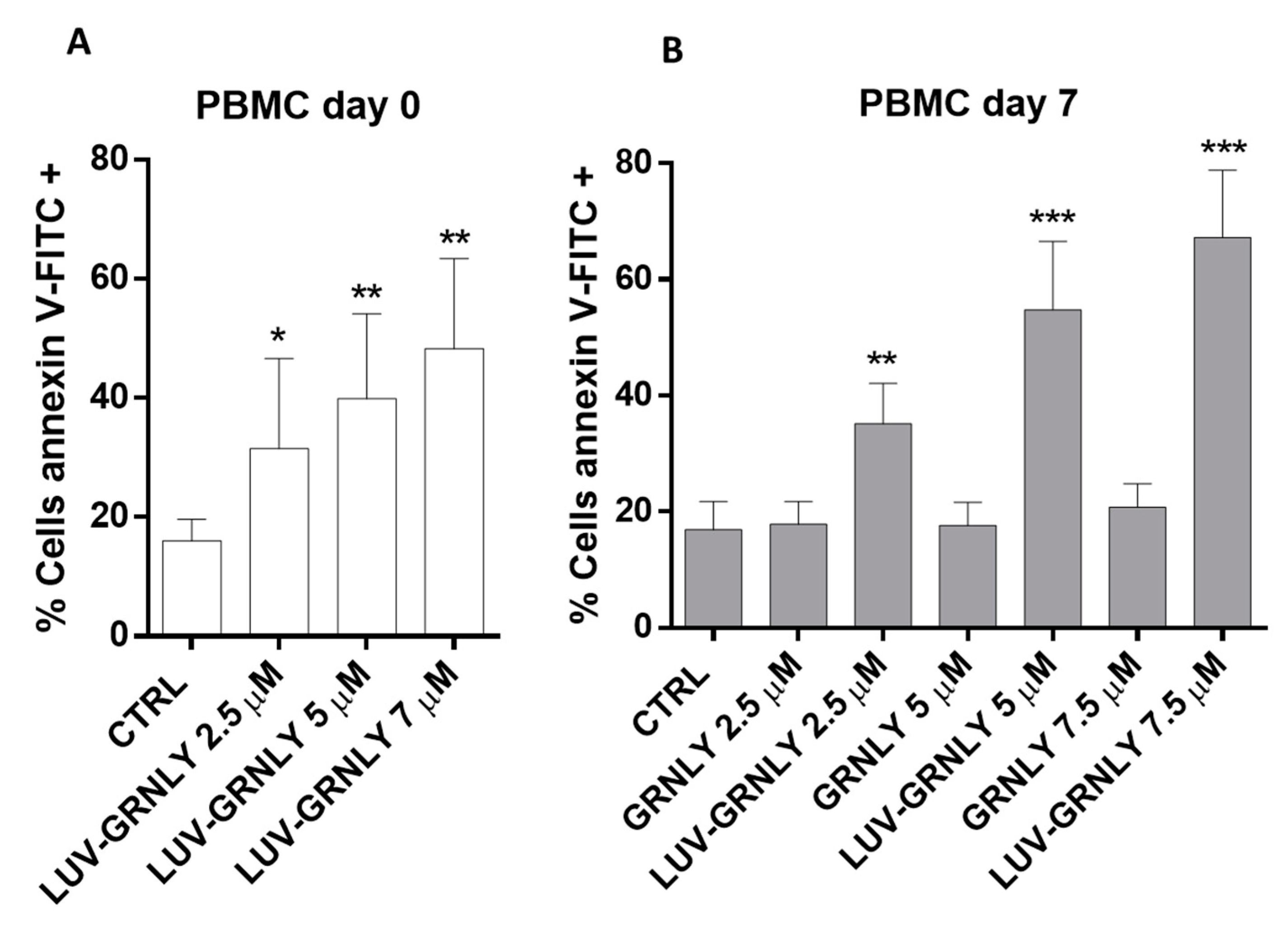
2.6. Toxicity against PBMC and T Cell Blasts from Healthy Donors
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Expression and Purification of 9 kDa Granulysin
4.3. Large Unilamellar Vesicle (LUV) Synthesis and Preparation of Lipid Nanoparticles Coated with 9 kDa Recombinant Granulysin (LUV-GRNLY)
4.4. Optimization Procedure for LUV-GRNLY Synthesis; Binding and Affinity Assays
4.5. Estimation of the Number of GRNLY Molecules per Liposome
4.6. In Vitro Cytotoxicity Assays—Apoptosis Quantification by Flow Cytometry
4.7. Western-Blot Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clayberger, C.; Finn, M.W.; Wang, T.; Saini, R.; Wilson, C.; Barr, V.A.; Sabatino, M.; Castiello, L.; Stroncek, D.; Krensky, A.M. 15 kDa Granulysin Causes Differentiation of Monocytes to Dendritic Cells but Lacks Cytotoxic Activity. J. Immunol. 2012, 188, 6119–6126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clayberger, C.; Krensky, A.M. Granulysin. Curr. Opin. Immunol. 2003, 15, 560–565. [Google Scholar] [CrossRef]
- Peña, S.V.; Hanson, D.A.; Carr, B.A.; Goralski, T.J.; Krensky, A.M. Processing, subcellular localization, and function of 519 (granulysin), a human late T cell activation molecule with homology to small, lytic, granule proteins. J. Immunol. 1997, 158, 2680–2688. [Google Scholar] [PubMed]
- Crespo, Â.; Mulik, S.; Dotiwala, F.; Ansara, J.; Sen Santara, S.; Ingersoll, K.; Ovies, C.; Junqueira, C.; Tilburgs, T.; Strominger, J.; et al. Decidual NK Cells Transfer Granulysin to Selectively Kill Bacteria in Trophoblasts. Cell 2020, 182, 1125–1139. [Google Scholar] [CrossRef]
- Dotiwala, F.; Mulik, S.; Polidoro, R.; Ansara, J.; Burleigh, B.; Walch, M.; Gazzinelli, R.; Lieberman, J. Killer lymphocytes use granulysin, perforin and granzymes to kill intracellular parasites. Nat. Med. 2016, 22, 210–216. [Google Scholar] [CrossRef]
- Stenger, S.; Hanson, D.A.; Teitelbaum, R.; Dewan, P.; Niazi, K.R.; Froelich, C.J.; Ganz, T.; Thoma-Uszynski, S.; Melián, A.; Bogdan, C.; et al. An antimicrobial activity of cytolytic T cells mediated by granulysin. Science 1998, 282, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Aporta, A.; Catalán, E.; Galán-Malo, P.; Ramírez-Labrada, A.; Pérez, M.; Azaceta, G.; Palomera, L.; Naval, J.; Marzo, I.; Pardo, J.; et al. Granulysin induces apoptotic cell death and cleavage of the autophagy regulator Atg5 in human hematological tumors. Biochem. Pharmacol. 2014, 87, 410–423. [Google Scholar] [CrossRef]
- Gamen, S.; Hanson, D.A.; Kaspar, A.; Naval, J.; Krensky, A.M.; Anel, A. Granulysin-induced apoptosis. I. Involvement of at least two distinct pathways. J. Immunol. 1998, 161, 1758–1764. [Google Scholar]
- Martinez-Lostao, L.; de Miguel, D.; Al-Wasaby, S.; Gallego-Lleyda, A.; Anel, A. Death ligands and granulysin: Mechanisms of tumor cell death induction and therapeutic opportunities. Immunotherapy 2015, 7, 883–892. [Google Scholar] [CrossRef]
- Kishi, A.; Takamori, Y.; Ogawa, K.; Takano, S.; Tomita, S.; Tanigawa, M.; Niman, M.; Kishida, T.; Fujita, S. Differential expression of granulysin and perforin by NK cells in cancer patients and correlation of impaired granulysin expression with progression of cancer. Cancer Immunol. Immunother. 2002, 50, 604–614. [Google Scholar] [CrossRef]
- Pagès, F.; Berger, A.; Camus, M.; Sanchez-Cabo, F.; Costes, A.; Molidor, R.; Mlecnik, B.; Kirilovsky, A.; Nilsson, M.; Damotte, D.; et al. Effector Memory T Cells, Early Metastasis, and Survival in Colorectal Cancer. N. Eng. J. Med. 2005, 353, 2654–2666. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, E.; Bodman-Smith, M. Granulysin: The attractive side of a natural born killer. Immunol. Lett. 2020, 217, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Tong, X.; Qu, X.; Wang, M. A Four-Gene-Based Prognostic Model Predicts Overall Survival in Patients with Cutaneous Melanoma. Front. Oncol. 2021, 11, 639874. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.P.; Lyu, S.C.; Clayberger, C.; Krensky, A.M. Granulysin-Mediated Tumor Rejection in Transgenic Mice. J. Immunol. 2007, 178, 77–84. [Google Scholar] [CrossRef]
- Al-Wasaby, S.; de Miguel, D.; Aporta, A.; Naval, J.; Conde, B.; Martínez-Lostao, L.; Anel, A. In vivo potential of recombinant granulysin against human tumors. Oncoimmunology 2015, 4, e1036213. [Google Scholar] [CrossRef] [Green Version]
- Al-Wasaby, S.; Guerrero-Ochoa, P.; Ibáñez-Pérez, R.; Soler, R.; Conde, B.; Martínez-Lostao, L.; Anel, A. In vivo potential of recombinant granulysin against human melanoma. Cancer Treat. Res. Commun. 2021, 27, 100355. [Google Scholar] [CrossRef]
- Ibáñez-Pérez, R.; Guerrero-Ochoa, P.; Al-Wasaby, S.; Navarro, R.; Tapia-Galisteo, A.; De Miguel, D.; Gonzalo, O.; Conde, B.; Martínez-Lostao, L.; Hurtado-Guerrero, R.; et al. Anti-tumoral potential of a human granulysinbased, CEA-targeted cytolytic immunotoxin. OncoImmunology 2019, 8, 1641392. [Google Scholar] [CrossRef] [Green Version]
- Guerrero-Ochoa, P.; Aguilar-Machado, D.; Ibáñez-Pérez, R.; Macías-León, J.; Hurtado-Guerrero, R.; Raso, J.; Anel, A. Production of a Granulysin-Based, Tn-Targeted Cytolytic Immunotoxin Using Pulsed Electric Field Technology. Int. J. Mol. Sci. 2020, 21, 6165. [Google Scholar] [CrossRef]
- Guerrero-Ochoa, P.; Ibáñez-Pérez, R.; Berbegal-Pinilla, G.; Aguilar, D.; Marzo, I.; Corzana, F.; Minjárez-Sáenz, M.; Macías-León, J.; Conde, B.; Raso, J.; et al. Preclinical Studies of Granulysin-Based Anti-MUC1-Tn Immunotoxins as a New Antitumoral Treatment. Biomedicines 2022, 10, 1223. [Google Scholar] [CrossRef]
- Sanz, L.; Ibáñez-Pérez, R.; Guerrero-Ochoa, P.; Lacadena, J.; Anel, A. Antibody-Based Immunotoxins for Colorectal Cancer Therapy. Biomedicines 2021, 9, 1729. [Google Scholar] [CrossRef]
- Shah, P.; Shende, P. Biomacromolecule-Functionalized Nanoparticle-Based Conjugates for Potentiation of Anticancer Therapy. Curr. Cancer Drug Targets 2022, 22, 31–48. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, D.; Basáñez, G.; Sánchez, D.; Galán, P.; Marzo, I.; Larrad, L.; Naval, J.; Pardo, J.; Anel, A.; Martinez-Lostao, L. Thethering Apo2L/TRAIL to liposomes overcomes chemoresistance of human hematological tumor cells. Mol. Pharm. 2013, 10, 893–904. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, D.; Lemke, J.; Anel, A.; Walczak, H.; Martinez-Lostao, L. Onto better TRAILs for cancer treatment. Cell Death Differ. 2016, 23, 733–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego-Lleyda, A.; De Miguel, D.; Anel, A.; Martinez-Lostao, L. Lipid Nanoparticles Decorated with TNF-Related Aptosis-Inducing Ligand (TRAIL) Are More Cytotoxic than Soluble Recombinant TRAIL in Sarcoma. Int. J. Mol. Sci. 2018, 19, 1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Miguel, D.; Gallego-Lleyda, A.; Martinez-Ara, M.; Plou, J.; Anel, A.; Martinez-Lostao, L. Double-Edged Lipid Nanoparticles Combining Liposome-Bound TRAIL and Encapsulated Doxorubicin Showing an Extraordinary Synergistic Pro-Apoptotic Potential. Cancers 2019, 11, 1948. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Luan, G.; Shen, G.; Wu, L.; Jia, H.; Zhong, Y.; Li, R.; Li, G.; Shen, Y.; Sun, J.; et al. Production and characterization of recombinant 9 and 15 kDa granulysin by fed-batch fermentation in Pichia pastoris. Appl. Microbiol. Biotechnol. 2013, 97, 7669–7677. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef]
- Cheong, I.; Huang, X.; Thornton, K.; Diaz, L.; Zhou, S. Targeting cancer with bugs and liposomes: Ready, aim, fire. Cancer Res. 2007, 67, 9605–9608. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.; Abuwatfa, W.; Awad, N.; Sabouni, R.; Husseini, G. Encapsulation, Release, and Cytotoxicity of Doxorubicin Loaded in Liposomes, Micelles, and Metal-Organic Frameworks: A Review. Pharmaceutics 2022, 14, 254. [Google Scholar] [CrossRef]
- Tarner, I.; Muller-Ladner, U. Drug delivery systems for the treatment of rheumatoid arthritis. Expert Opin. Drug Deliv. 2008, 5, 1027–1037. [Google Scholar] [CrossRef]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Anel, A.; Gallego-Lleyda, A.; de Miguel, D.; Naval, J.; Martinez-Lostao, L. Role of Exosomes in the Regulation of T-cell Mediated Immune Responses and in Autoimmune Disease. Cells 2019, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Lorenzo, M.J.; Anel, A.; Gamen, S.; Monleón, I.; Lasierra, P.; Larrad, L.; Piñeiro, A.; Alava, M.A.; Naval, J. Activated human T cells release bioactive Fas ligand and APO2 ligand in microvesicles. J. Immunol. 1999, 163, 1274–1281. [Google Scholar] [PubMed]
- Naval, J.; de Miguel, D.; Gallego-Lleyda, A.; Anel, A.; Martinez-Lostao, L. Importance of TRAIL Molecular Anatomy in Receptor Oligomerization and Signaling. Implications for Cancer Therapy. Cancers 2019, 11, 444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejigah, V.; Owoseni, O.; Bataille-Backer, P.; Ogundipe, O.; Fisusi, F.; Adesina, S. Approaches to Improve Macromolecule and Nanoparticle Accumulation in the Tumor Microenvironment by the Enhanced Permeability and Retention Effect. Polymers 2022, 14, 2601. [Google Scholar] [CrossRef] [PubMed]
- Stylianopoulos, T. EPR-effect: Utilizing size-dependent nanoparticle delivery to solid tumors. Ther. Deliv. 2013, 4, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.; Vaux, D. Alterations in the apoptotic machinery and their potential role in anticancer drug resistance. Oncogene 2003, 22, 7414–7430. [Google Scholar] [CrossRef] [Green Version]
- Merino, D.; Kelly, G.; Lessene, G.; Wei, A.; Roberts, A.; Strasser, A. BH3-Mimetic Drugs: Blazing the Trail for New Cancer Medicines. Cancer Cell 2018, 34, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Radha, G.; Raghavan, S. BCL2: A promising cancer therapeutic target. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 309–314. [Google Scholar] [CrossRef]
- Barman, H.; Walch, M.; Latinovic-Golic, S.; Dumrese, C.; Dolder, M.; Groscurth, P.; Ziegler, U. Cholesterol in Negatively Charged Lipid Bilayers Modulates the Effect of the Antimicrobial Protein Granulysin. J. Membr. Biol. 2006, 212, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Bosque, A.; Pardo, J.; Martínez-Lorenzo, M.J.; Iturralde, M.; Marzo, I.; Piñeiro, A.; Alava, M.A.; Naval, J.; Anel, A. Down-regulation of normal human T cell blast activation: Roles of APO2L/TRAIL, FasL and c- FLIP, Bim or Bcl-x isoform expression. J. Leukoc. Biol. 2005, 77, 568–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Lostao, L.; García-Alvarez, F.; Basáñez, G.; Alegre-Aguarón, E.; Desportes, P.; Larrad, L.; Naval, J.; Martínez-Lorenzo, M.J.; Anel, A. Liposome-Bound APO2L/TRAIL Is an Effective Treatment in a Rabbit Model of Rheumatoid Arthritis. Arthrit. Rheum. 2010, 62, 2272–2282. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soler-Agesta, R.; Guerrero-Ochoa, P.; Marco-Brualla, J.; Ibáñez-Pérez, R.; Marzo, I.; Martínez-Lostao, L.; Anel, A. Conjugation of the 9-kDa Isoform of Granulysin with Liposomes Potentiates Its Cytotoxicity. Int. J. Mol. Sci. 2022, 23, 8705. https://doi.org/10.3390/ijms23158705
Soler-Agesta R, Guerrero-Ochoa P, Marco-Brualla J, Ibáñez-Pérez R, Marzo I, Martínez-Lostao L, Anel A. Conjugation of the 9-kDa Isoform of Granulysin with Liposomes Potentiates Its Cytotoxicity. International Journal of Molecular Sciences. 2022; 23(15):8705. https://doi.org/10.3390/ijms23158705
Chicago/Turabian StyleSoler-Agesta, Ruth, Patricia Guerrero-Ochoa, Joaquín Marco-Brualla, Raquel Ibáñez-Pérez, Isabel Marzo, Luis Martínez-Lostao, and Alberto Anel. 2022. "Conjugation of the 9-kDa Isoform of Granulysin with Liposomes Potentiates Its Cytotoxicity" International Journal of Molecular Sciences 23, no. 15: 8705. https://doi.org/10.3390/ijms23158705