
Insight into the Mammalian Aquaporin Interactome

 ,
,
Abstract
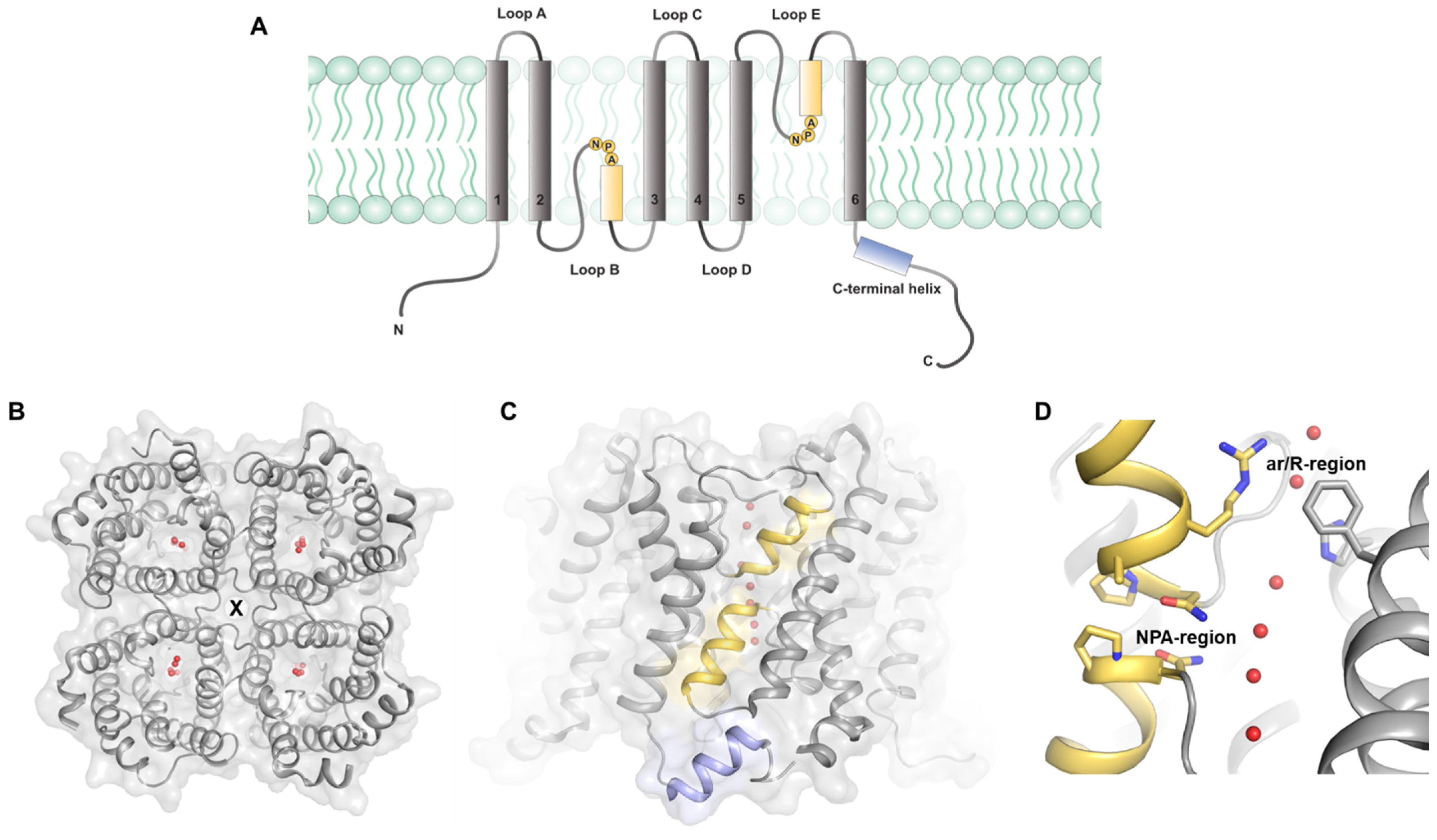
:1. Introduction
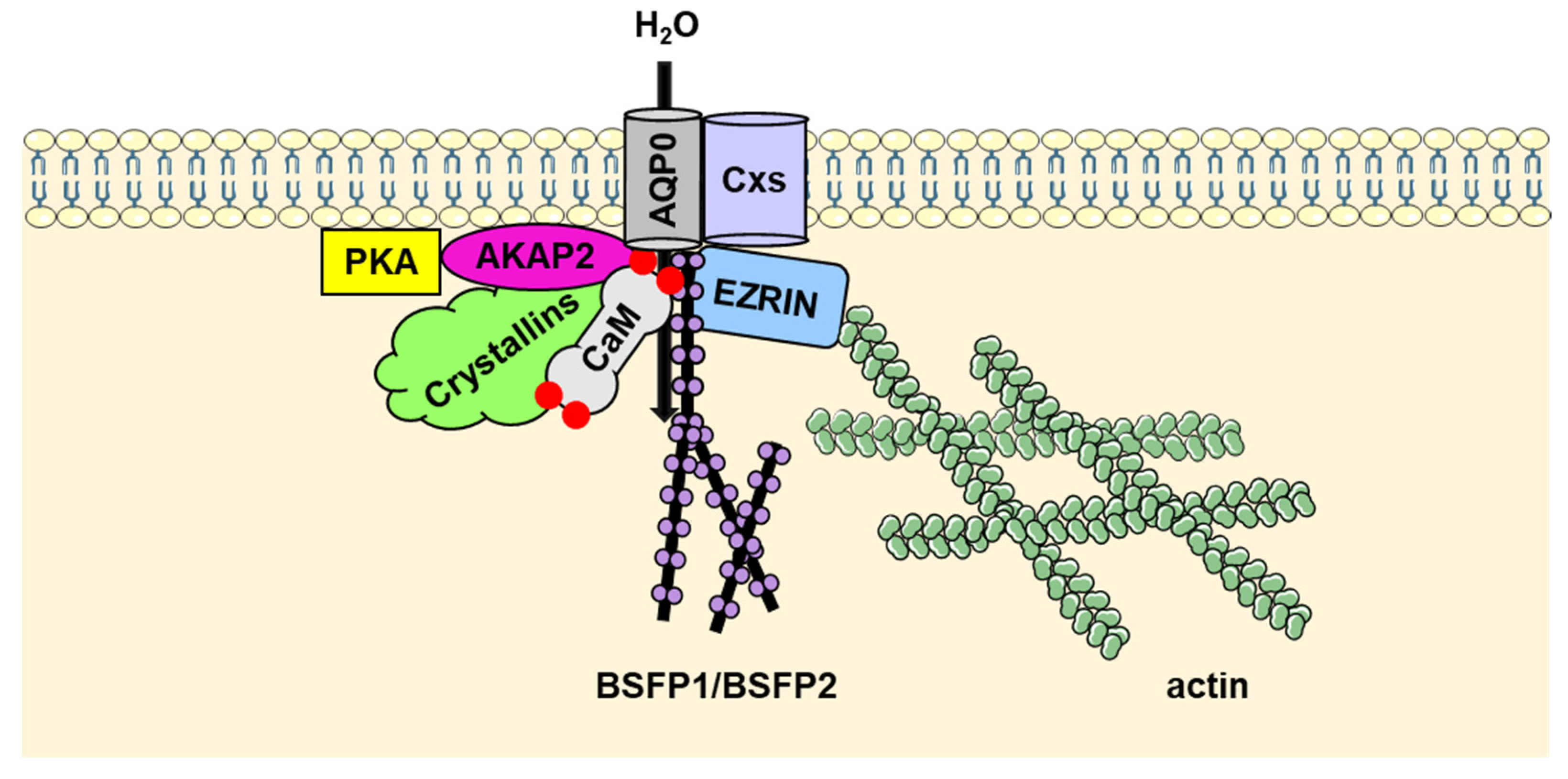
2. AQP0
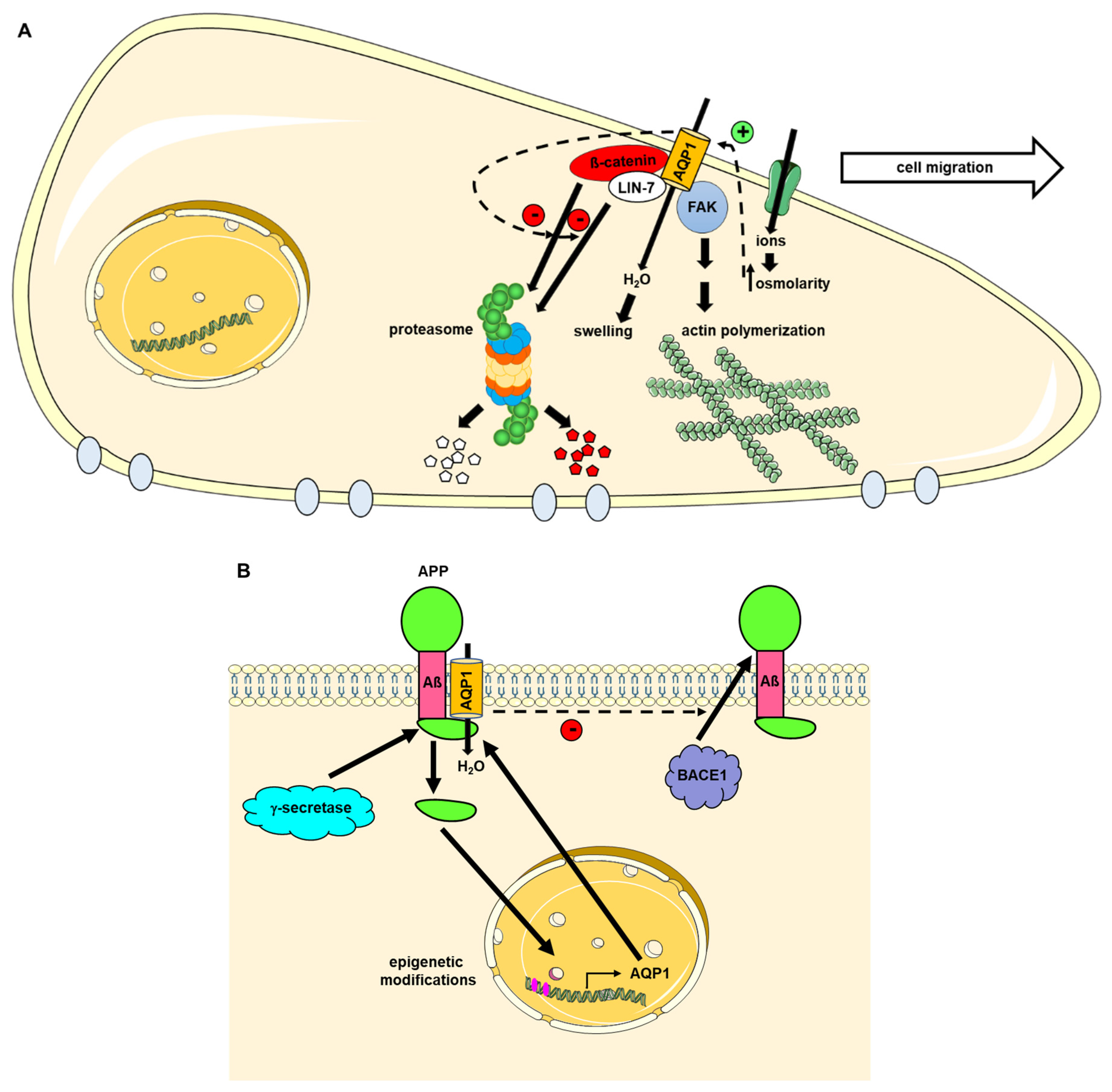
3. AQP1
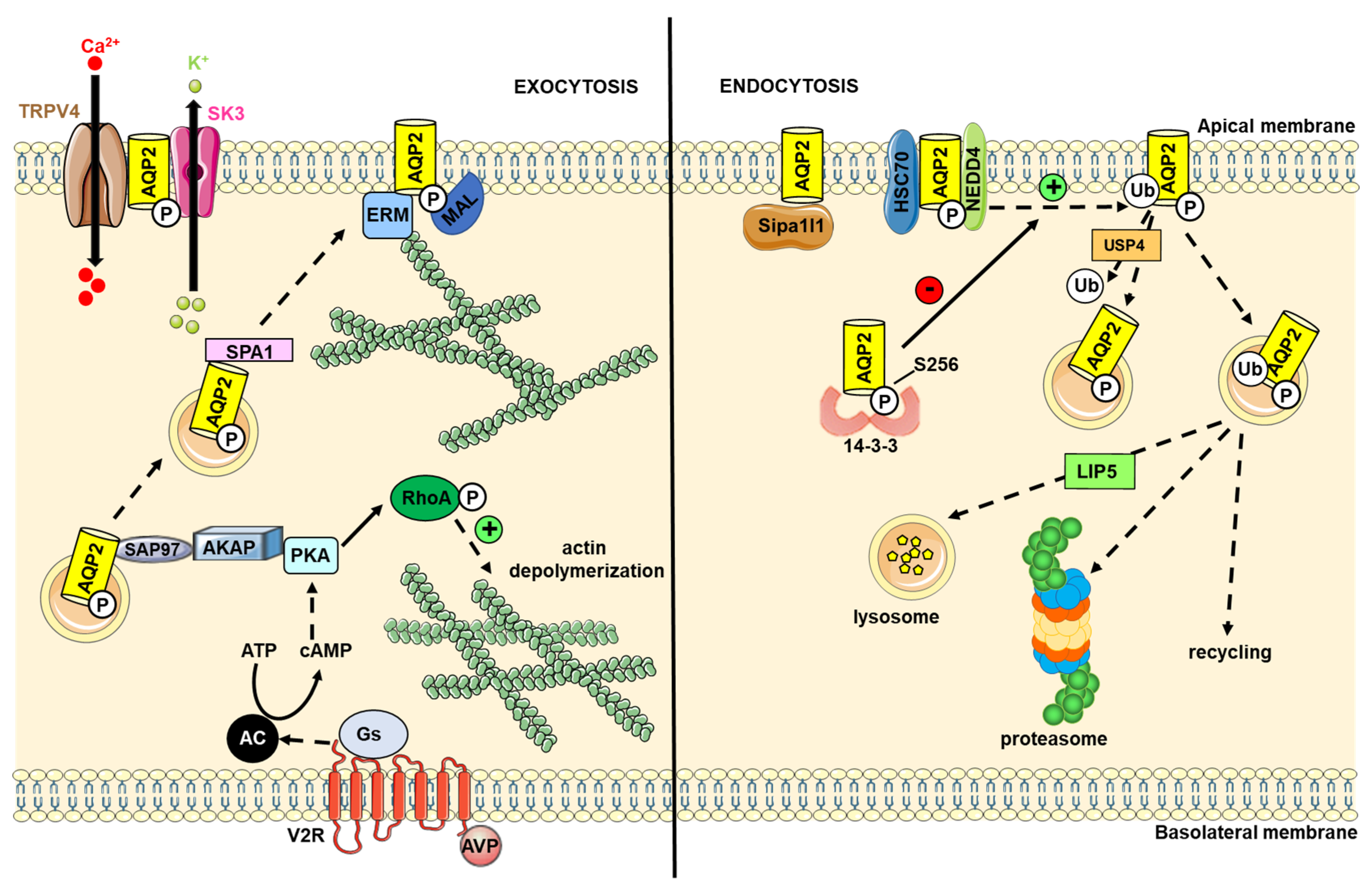
4. AQP2
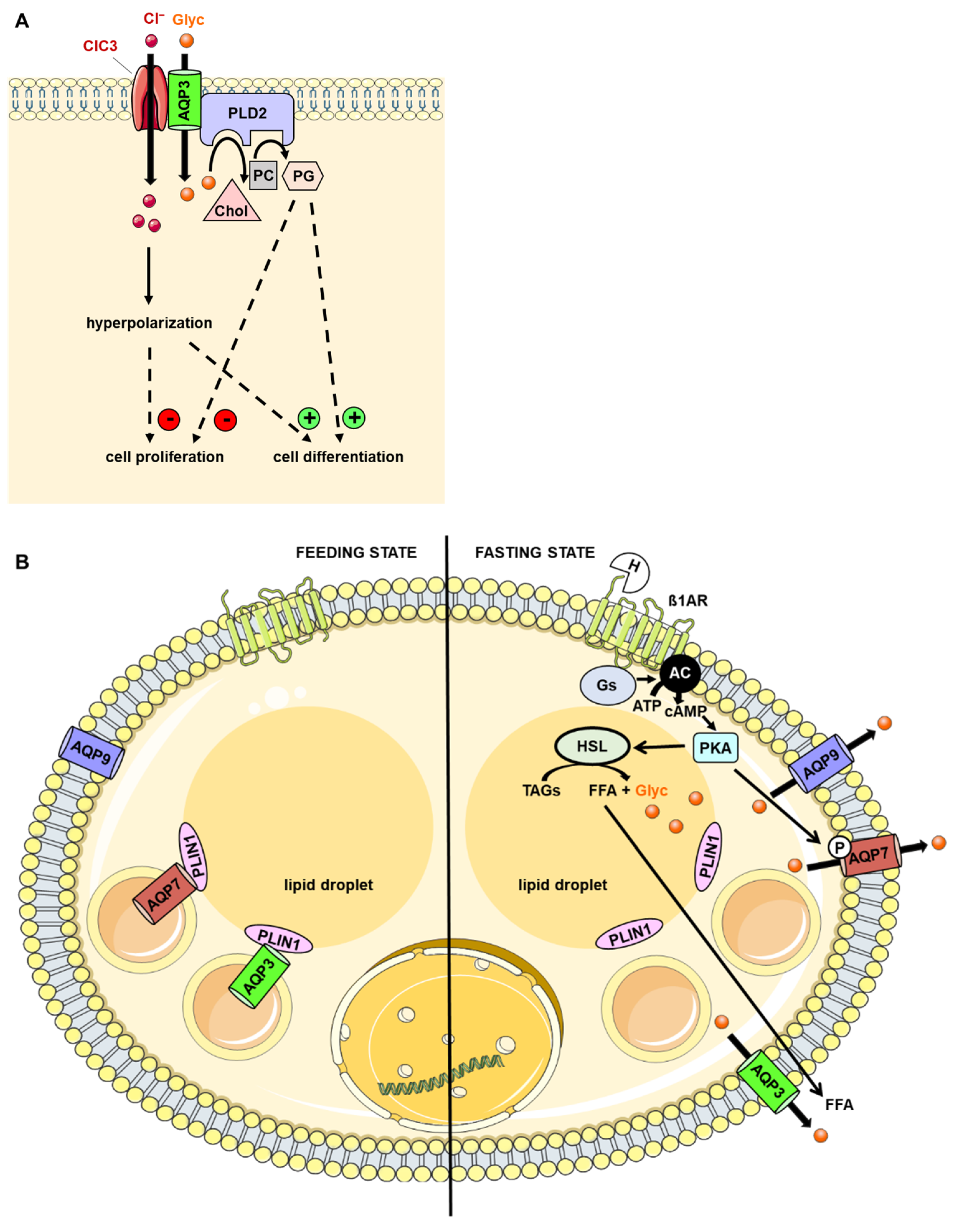
5. AQP3

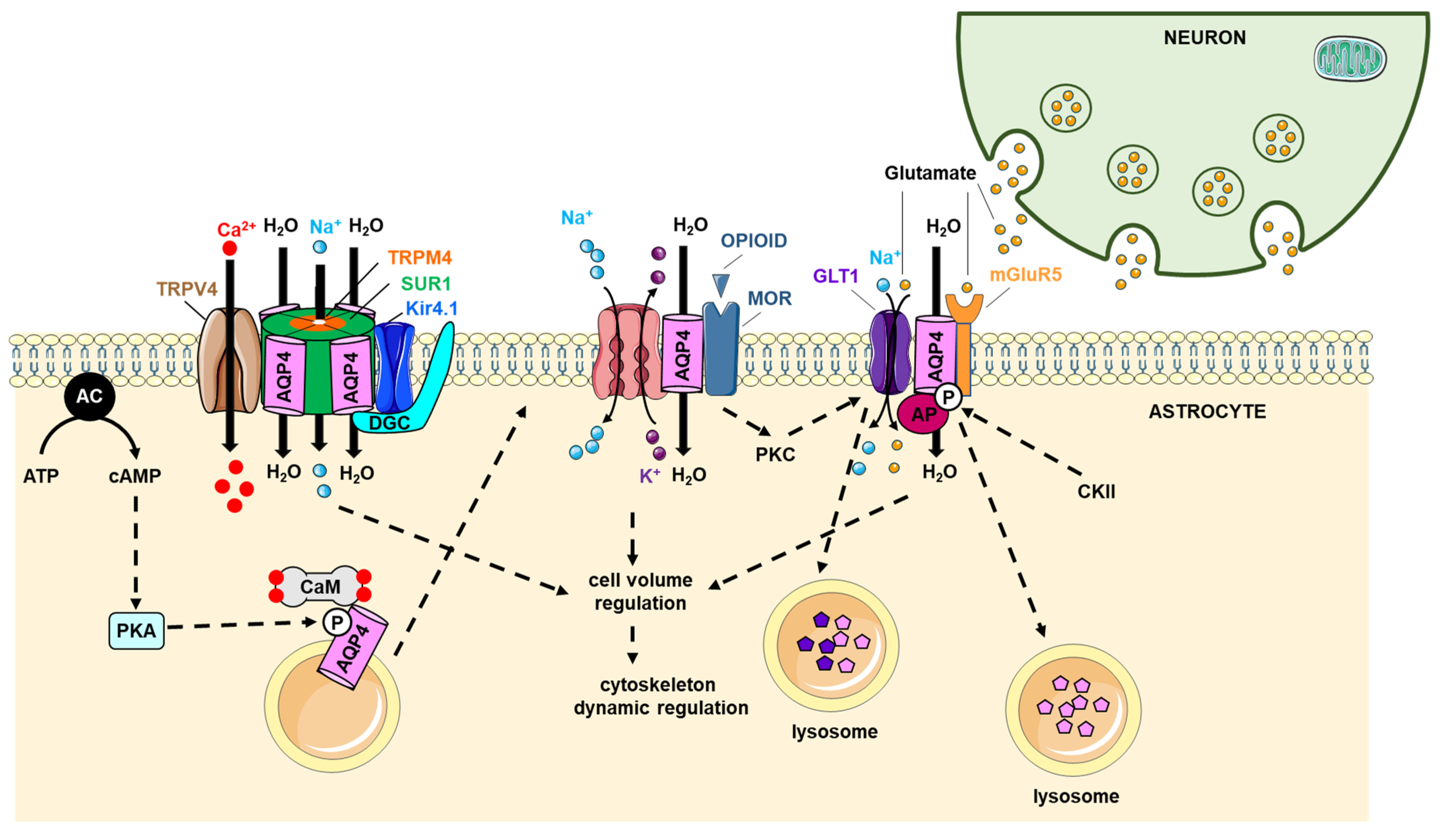
6. AQP4
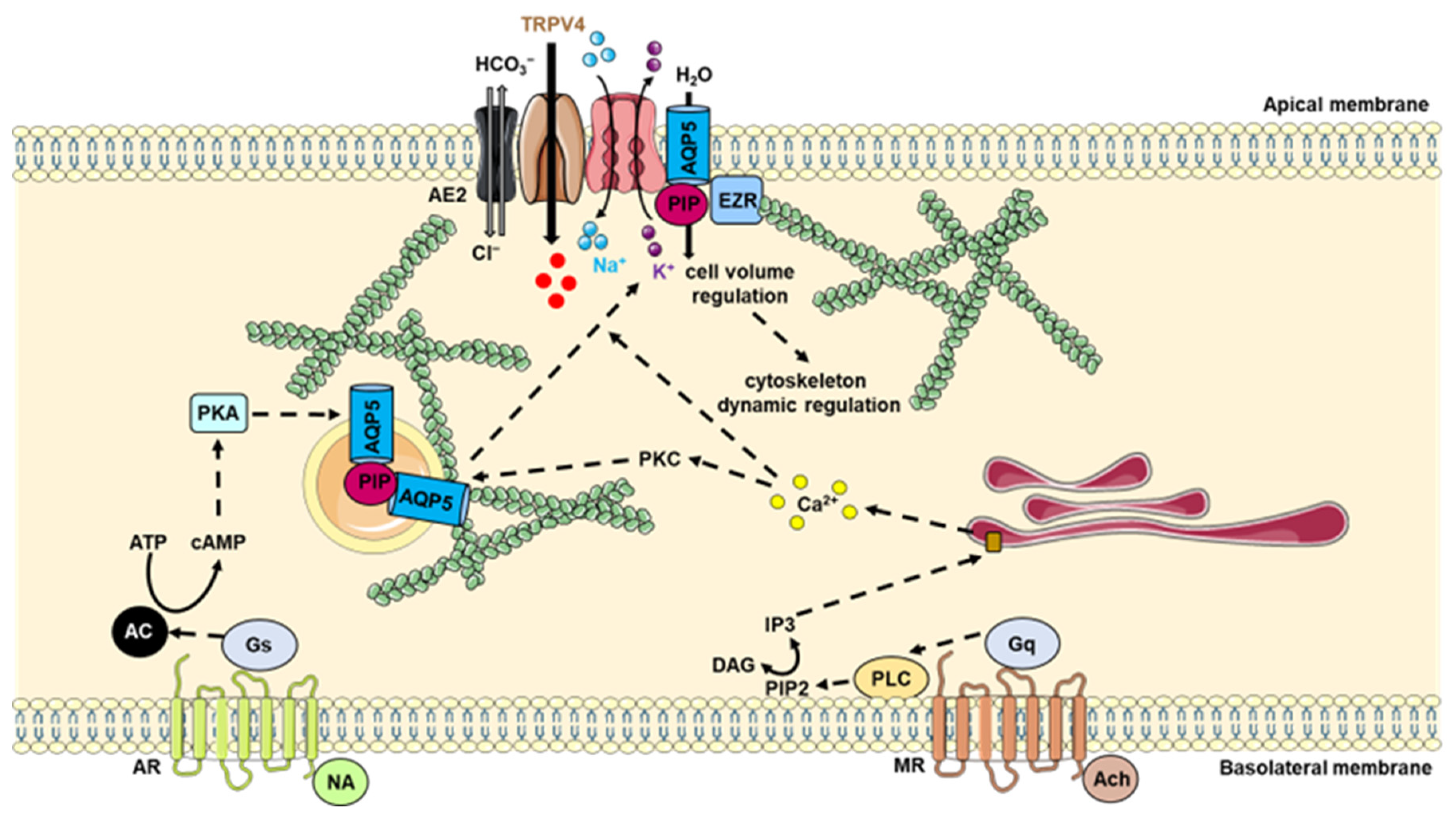
7. AQP5
8. AQP6
9. AQP7
10. AQP8
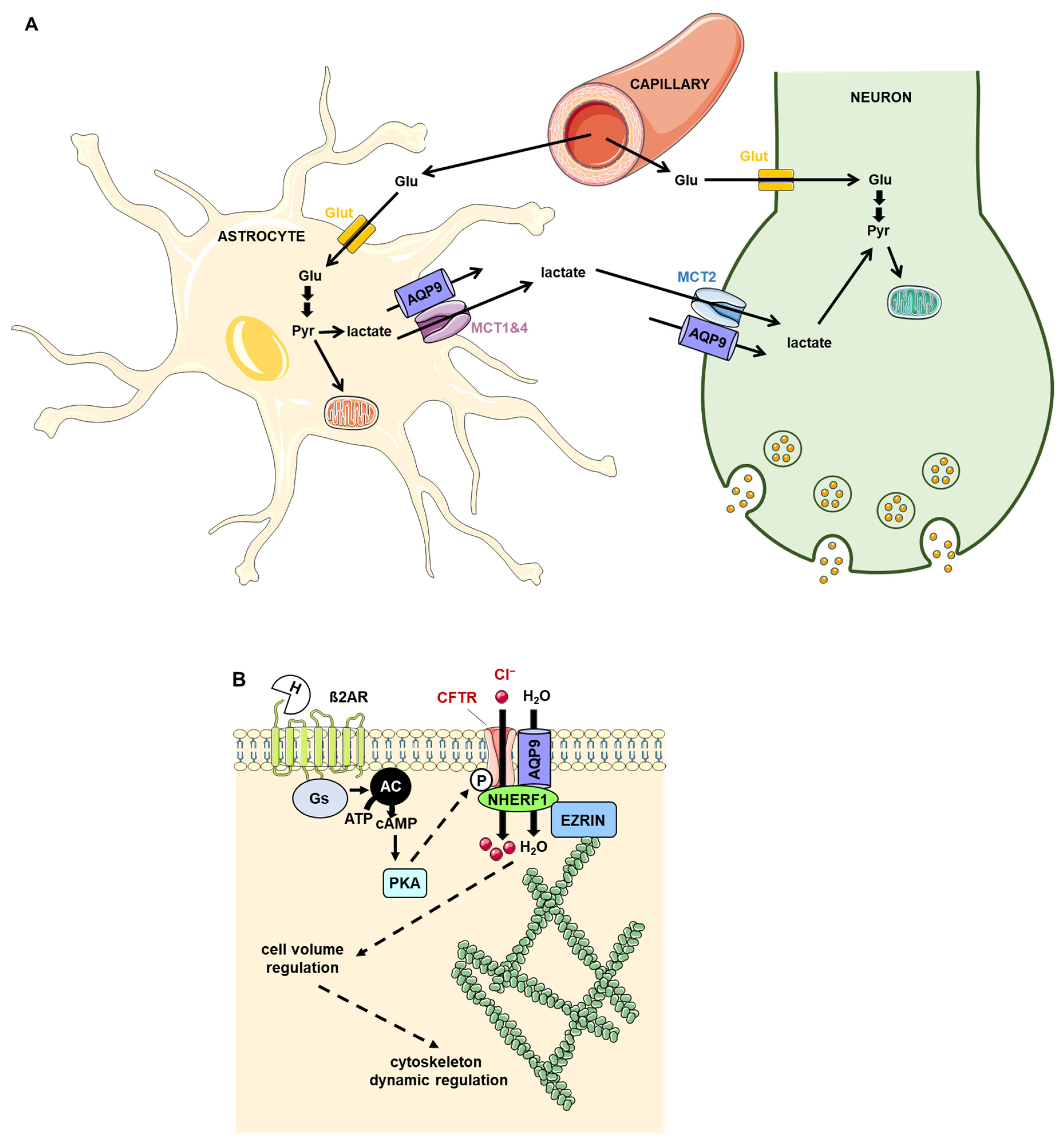
11. AQP9
12. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Agre, P. Aquaporin Water Channels (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2004, 43, 4278–4290. [Google Scholar] [CrossRef] [PubMed]
- Laloux, T.; Junqueira, B.; Maistriaux, L.C.; Ahmed, J.; Jurkiewicz, A.; Chaumont, F. Plant and Mammal Aquaporins: Same but Different. Int. J. Mol. Sci. 2018, 19, 521. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S.; Mitra, A.K. Structure and Function of Aquaporin Water Channels. Am. J. Physiol. Ren. Physiol. 2000, 278, F13–F28. [Google Scholar] [CrossRef]
- Madeira, A.; Fernández-Veledo, S.; Camps, M.; Zorzano, A.; Moura, T.F.; Ceperuelo-Mallafré, V.; Vendrell, J.; Soveral, G. Human Aquaporin-11 Is a Water and Glycerol Channel and Localizes in the Vicinity of Lipid Droplets in Human Adipocytes. Obesity 2014, 22, 2010–2017. [Google Scholar] [CrossRef]
- Sorrentino, I.; Galli, M.; Medraño-Fernandez, I.; Sitia, R. Transfer of H2O2 from Mitochondria to the Endoplasmic Reticulum via Aquaporin-11. Redox Biol. 2022, 55, 102410. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Conner, M.T.; Bill, R.M.; Conner, A.C. Structural Determinants of Oligomerization of the Aquaporin-4 Channel. J. Biol. Chem. 2016, 291, 6858–6871. [Google Scholar] [CrossRef] [PubMed]
- Sjöhamn, J.; Hedfalk, K. Unraveling Aquaporin Interaction Partners. Biochim. Biophys. Acta 2014, 1840, 1614–1623. [Google Scholar] [CrossRef]
- Roche, J.V.; Törnroth-Horsefield, S. Aquaporin Protein-Protein Interactions. Int J. Mol. Sci. 2017, 18, 2255. [Google Scholar] [CrossRef]
- Verkman, A.S. Role of Aquaporin Water Channels in Eye Function. Exp. Eye Res. 2003, 76, 137–143. [Google Scholar] [CrossRef]
- Schey, K.L.; Petrova, R.S.; Gletten, R.B.; Donaldson, P.J. The Role of Aquaporins in Ocular Lens Homeostasis. Int. J. Mol. Sci. 2017, 18, 2693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.D.; King, L.S.; Agre, P. The Aquaporin Family of Water Channel Proteins in Clinical Medicine. Medicine 1997, 76, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, K.; Tanaka, A.; Jung, K.; Matsuda, A.; Okamoto, N.; Oida, K.; Ohmori, K.; Matsuda, H. Patterns of Aquaporin Expression in the Canine Eye. Vet. J. 2011, 190, e72–e77. [Google Scholar] [CrossRef] [PubMed]
- Hermo, L.; Krzeczunowicz, D.; Ruz, R. Cell Specificity of Aquaporins 0, 3, and 10 Expressed in the Testis, Efferent Ducts, and Epididymis of Adult Rats. J. Androl. 2004, 25, 494–505. [Google Scholar] [CrossRef]
- Huebert, R.C.; Splinter, P.L.; Garcia, F.; Marinelli, R.A.; LaRusso, N.F. Expression and Localization of Aquaporin Water Channels in Rat Hepatocytes. Evidence for a Role in Canalicular Bile Secretion. J. Biol. Chem. 2002, 277, 22710–22717. [Google Scholar] [CrossRef] [PubMed]
- Varadaraj, K.; Kumari, S.S. Molecular Mechanism of Aquaporin 0-Induced Fiber Cell to Fiber Cell Adhesion in the Eye Lens. Biochem. Biophys. Res. Commun. 2018, 506, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Gonen, T.; Cheng, Y.; Kistler, J.; Walz, T. Aquaporin-0 Membrane Junctions Form upon Proteolytic Cleavage. J. Mol. Biol. 2004, 342, 1337–1345. [Google Scholar] [CrossRef]
- Gonen, T.; Cheng, Y.; Sliz, P.; Hiroaki, Y.; Fujiyoshi, Y.; Harrison, S.C.; Walz, T. Lipid-Protein Interactions in Double-Layered Two-Dimensional AQP0 Crystals. Nature 2005, 438, 633–638. [Google Scholar] [CrossRef]
- Gonen, T.; Sliz, P.; Kistler, J.; Cheng, Y.; Walz, T. Aquaporin-0 Membrane Junctions Reveal the Structure of a Closed Water Pore. Nature 2004, 429, 193–197. [Google Scholar] [CrossRef]
- Ball, L.E.; Little, M.; Nowak, M.W.; Garland, D.L.; Crouch, R.K.; Schey, K.L. Water Permeability of C-Terminally Truncated Aquaporin 0 (AQP0 1-243) Observed in the Aging Human Lens. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4820–4828. [Google Scholar] [CrossRef]
- Kumari, S.S.; Varadaraj, K. Intact and N- or C-terminal end truncated AQP0 function as open water channels and cell-to-cell adhesion proteins: End truncation could be a prelude for adjusting the refractive index of the lens to prevent spherical aberration. Biochim. Biophys. Acta. 2014, 1840, 2862–2877. [Google Scholar] [CrossRef] [Green Version]
- Villalobo, A.; Ishida, H.; Vogel, H.J.; Berchtold, M.W. Calmodulin as a Protein Linker and a Regulator of Adaptor/Scaffold Proteins. Biochim. Biophys. Acta Mol. Cell. Res. 2018, 1865, 507–521. [Google Scholar] [CrossRef] [PubMed]
- Girsch, S.J.; Peracchia, C. Lens Cell-to-Cell Channel Protein: I. Self-Assembly into Liposomes and Permeability Regulation by Calmodulin. J. Membr. Biol. 1985, 83, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Reichow, S.L.; Clemens, D.M.; Freites, J.A.; Németh-Cahalan, K.L.; Heyden, M.; Tobias, D.J.; Hall, J.E.; Gonen, T. Allosteric Mechanism of Water-Channel Gating by Ca2+-Calmodulin. Nat. Struct. Mol. Biol. 2013, 20, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Reichow, S.L.; Gonen, T. Noncanonical Binding of Calmodulin to Aquaporin-0: Implications for Channel Regulation. Structure 2008, 16, 1389–1398. [Google Scholar] [CrossRef]
- Kreida, S.; Roche, J.V.; Olsson, C.; Linse, S.; Törnroth-Horsefield, S. Protein-Protein Interactions in AQP Regulation–Biophysical Characterization of AQP0-CaM and AQP2-LIP5 Complex Formation. Faraday Discuss. 2018, 209, 35–54. [Google Scholar] [CrossRef]
- Fields, J.B.; Németh-Cahalan, K.L.; Freites, J.A.; Vorontsova, I.; Hall, J.E.; Tobias, D.J. Calmodulin Gates Aquaporin 0 Permeability through a Positively Charged Cytoplasmic Loop. J. Biol. Chem. 2017, 292, 185–195. [Google Scholar] [CrossRef]
- Rose, K.M.L.; Wang, Z.; Magrath, G.N.; Hazard, E.S.; Hildebrandt, J.D.; Schey, K.L. Aquaporin 0-Calmodulin Interaction and the Effect of Aquaporin 0 Phosphorylation. Biochemistry 2008, 47, 339–347. [Google Scholar] [CrossRef]
- Gold, M.G.; Reichow, S.L.; O’Neill, S.E.; Weisbrod, C.R.; Langeberg, L.K.; Bruce, J.E.; Gonen, T.; Scott, J.D. AKAP2 Anchors PKA with Aquaporin-0 to Support Ocular Lens Transparency. EMBO Mol. Med. 2012, 4, 15–26. [Google Scholar] [CrossRef]
- Greenwald, E.C.; Saucerman, J.J. Bigger, Better, Faster: Principles and Models of AKAP Anchoring Protein Signaling. J. Cardiovasc. Pharmacol. 2011, 58, 462–469. [Google Scholar] [CrossRef]
- Ponuwei, G.A. A Glimpse of the ERM Proteins. J. Biomed. Sci. 2016, 23, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Schey, K.L. Aquaporin-0 Interacts with the FERM Domain of Ezrin/Radixin/Moesin Proteins in the Ocular Lens. Invest. Ophthalmol. Vis. Sci. 2011, 52, 5079–5087. [Google Scholar] [CrossRef] [PubMed]
- Straub, B.K.; Boda, J.; Kuhn, C.; Schnoelzer, M.; Korf, U.; Kempf, T.; Spring, H.; Hatzfeld, M.; Franke, W.W. A Novel Cell-Cell Junction System: The Cortex Adhaerens Mosaic of Lens Fiber Cells. J. Cell. Sci. 2003, 116, 4985–4995. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Landsbury, A.; Dahm, R.; Liu, Y.; Zhang, Q.; Quinlan, R.A. Functions of the Intermediate Filament Cytoskeleton in the Eye Lens. J. Clin. Investig. 2009, 119, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Hutcheson, A.M.; Quinlan, R.A. In Vitro Studies on the Assembly Properties of the Lens Proteins CP49, CP115: Coassembly with Alpha-Crystallin but Not with Vimentin. Exp. Eye Res. 1995, 60, 181–192. [Google Scholar] [CrossRef]
- Lindsey Rose, K.M.; Gourdie, R.G.; Prescott, A.R.; Quinlan, R.A.; Crouch, R.K.; Schey, K.L. The C Terminus of Lens Aquaporin 0 Interacts with the Cytoskeletal Proteins Filensin and CP49. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1562–1570. [Google Scholar] [CrossRef]
- Wang, Z.; Schey, K.L. Identification of a Direct Aquaporin-0 Binding Site in the Lens-Specific Cytoskeletal Protein Filensin. Exp. Eye Res. 2017, 159, 23–29. [Google Scholar] [CrossRef]
- Nakazawa, Y.; Oka, M.; Furuki, K.; Mitsuishi, A.; Nakashima, E.; Takehana, M. The Effect of the Interaction between Aquaporin 0 (AQP0) and the Filensin Tail Region on AQP0 Water Permeability. Mol. Vis. 2011, 17, 3191–3199. [Google Scholar] [PubMed]
- Liu, J.; Xu, J.; Gu, S.; Nicholson, B.J.; Jiang, J.X. Aquaporin 0 Enhances Gap Junction Coupling via Its Cell Adhesion Function and Interaction with Connexin 50. J. Cell. Sci. 2011, 124, 198–206. [Google Scholar] [CrossRef]
- Beyer, E.C.; Berthoud, V.M. Connexin Hemichannels in the Lens. Front. Physiol. 2014, 5, 20. [Google Scholar] [CrossRef]
- Yu, X.S.; Jiang, J.X. Interaction of Major Intrinsic Protein (Aquaporin-0) with Fiber Connexins in Lens Development. J. Cell. Sci. 2004, 117, 871–880. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.S.; Yin, X.; Lafer, E.M.; Jiang, J.X. Developmental Regulation of the Direct Interaction between the Intracellular Loop of Connexin 45.6 and the C Terminus of Major Intrinsic Protein (Aquaporin-0). J. Biol. Chem. 2005, 280, 22081–22090. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Biswas, S.; Rodriguez, L.; Li, Z.; Li, Y.; Riquelme, M.A.; Shi, W.; Wang, K.; White, T.W.; Reilly, M.; et al. Connexin 50 and AQP0 Are Essential in Maintaining Organization and Integrity of Lens Fibers. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4021–4032. [Google Scholar] [CrossRef] [PubMed]
- Sprague-Piercy, M.A.; Rocha, M.A.; Kwok, A.O.; Martin, R.W. α-Crystallins in the Vertebrate Eye Lens: Complex Oligomers and Molecular Chaperones. Annu. Rev. Phys. Chem. 2021, 72, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.-F.; Liang, J.J. Confocal Fluorescence Microscopy Study of Interaction between Lens MIP26/AQP0 and Crystallins in Living Cells. J. Cell. Biochem. 2008, 104, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Swamy-Mruthinti, S.; Srinivas, V.; Hansen, J.E.; Rao, C.M. Thermal Stress Induced Aggregation of Aquaporin 0 (AQP0) and Protection by α-Crystallin via Its Chaperone Function. PLoS ONE 2013, 8, e80404. [Google Scholar] [CrossRef]
- Fan, J.; Donovan, A.K.; Ledee, D.R.; Zelenka, P.S.; Fariss, R.N.; Chepelinsky, A.B. GammaE-Crystallin Recruitment to the Plasma Membrane by Specific Interaction between Lens MIP/Aquaporin-0 and GammaE-Crystallin. Investig. Ophthalmol. Vis. Sci. 2004, 45, 863–871. [Google Scholar] [CrossRef]
- Wang, Z.; Friedrich, M.G.; Truscott, R.J.W.; Schey, K.L. Cleavage C-Terminal to Asp Leads to Covalent Crosslinking of Long-Lived Human Proteins. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 831–839. [Google Scholar] [CrossRef]
- Monzani, E.; Bazzotti, R.; Perego, C.; La Porta, C.A.M. AQP1 Is Not Only a Water Channel: It Contributes to Cell Migration through Lin7/Beta-Catenin. PLoS ONE 2009, 4, e6167. [Google Scholar] [CrossRef]
- Meng, F.; Rui, Y.; Xu, L.; Wan, C.; Jiang, X.; Li, G. Aqp1 Enhances Migration of Bone Marrow Mesenchymal Stem Cells through Regulation of FAK and β-Catenin. Stem Cells Dev. 2014, 23, 66–75. [Google Scholar] [CrossRef]
- Saadoun, S.; Papadopoulos, M.C.; Hara-Chikuma, M.; Verkman, A.S. Impairment of Angiogenesis and Cell Migration by Targeted Aquaporin-1 Gene Disruption. Nature 2005, 434, 786–792. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Saadoun, S.; Verkman, A.S. Aquaporins and Cell Migration. Pflug. Arch. 2008, 456, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y. Aquaporin-1 Activity of Plasma Membrane Affects HT20 Colon Cancer Cell Migration. IUBMB Life 2009, 61, 1001–1009. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Dorward, H.; Yool, A.J.; Smith, E.; Townsend, A.R.; Price, T.J.; Hardingham, J.E. Role of Aquaporin 1 Signalling in Cancer Development and Progression. Int. J. Mol. Sci. 2017, 18, 299. [Google Scholar] [CrossRef] [PubMed]
- Dajani, S.; Saripalli, A.; Sharma-Walia, N. Water Transport Proteins-Aquaporins (AQPs) in Cancer Biology. Oncotarget 2018, 9, 36392–36405. [Google Scholar] [CrossRef]
- Wang, W.; Li, Q.; Yang, T.; Li, D.; Ding, F.; Sun, H.; Bai, G. Anti-Cancer Effect of Aquaporin 5 Silencing in Colorectal Cancer Cells in Association with Inhibition of Wnt/β-Catenin Pathway. Cytotechnology 2018, 70, 615–624. [Google Scholar] [CrossRef]
- Misawa, T.; Arima, K.; Mizusawa, H.; Satoh, J. Close Association of Water Channel AQP1 with Amyloid-Beta Deposition in Alzheimer Disease Brains. Acta Neuropathol. 2008, 116, 247–260. [Google Scholar] [CrossRef]
- Huysseune, S.; Kienlen-Campard, P.; Hébert, S.; Tasiaux, B.; Leroy, K.; Devuyst, O.; Brion, J.-P.; De Strooper, B.; Octave, J.-N. Epigenetic Control of Aquaporin 1 Expression by the Amyloid Precursor Protein. FASEB J. 2009, 23, 4158–4167. [Google Scholar] [CrossRef]
- Park, J.; Madan, M.; Chigurupati, S.; Baek, S.H.; Cho, Y.; Mughal, M.R.; Yu, A.; Chan, S.L.; Pattisapu, J.V.; Mattson, M.P.; et al. Neuronal Aquaporin 1 Inhibits Amyloidogenesis by Suppressing the Interaction Between Beta-Secretase and Amyloid Precursor Protein. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76, 23–31. [Google Scholar] [CrossRef]
- Noda, Y.; Sasaki, S. Updates and Perspectives on Aquaporin-2 and Water Balance Disorders. Int. J. Mol. Sci. 2021, 22, 12950. [Google Scholar] [CrossRef]
- Fenton, R.A.; Murali, S.K.; Moeller, H.B. Advances in Aquaporin-2 Trafficking Mechanisms and Their Implications for Treatment of Water Balance Disorders. Am. J. Physiol. Cell Physiol. 2020, 319, C1–C10. [Google Scholar] [CrossRef]
- Isobe, K.; Jung, H.J.; Yang, C.-R.; Claxton, J.; Sandoval, P.; Burg, M.B.; Raghuram, V.; Knepper, M.A. Systems-Level Identification of PKA-Dependent Signaling in Epithelial Cells. Proc. Natl. Acad. Sci. USA 2017, 114, E8875–E8884. [Google Scholar] [CrossRef] [PubMed]
- Yui, N.; Ando, F.; Sasaki, S.; Uchida, S. Ser-261 Phospho-Regulation Is Involved in PS256 and PS269-Mediated Aquaporin-2 Apical Translocation. Biochem. Biophys. Res. Commun. 2017, 490, 1039–1044. [Google Scholar] [CrossRef] [PubMed]
- Kamsteeg, E.-J.; Hendriks, G.; Boone, M.; Konings, I.B.M.; Oorschot, V.; van der Sluijs, P.; Klumperman, J.; Deen, P.M.T. Short-Chain Ubiquitination Mediates the Regulated Endocytosis of the Aquaporin-2 Water Channel. Proc. Natl. Acad. Sci. USA 2006, 103, 18344–18349. [Google Scholar] [CrossRef] [PubMed]
- Chou, C.-L.; Hwang, G.; Hageman, D.J.; Han, L.; Agrawal, P.; Pisitkun, T.; Knepper, M.A. Identification of UT-A1- and AQP2-Interacting Proteins in Rat Inner Medullary Collecting Duct. Am. J. Physiol. Cell Physiol. 2018, 314, C99–C117. [Google Scholar] [CrossRef]
- Tamma, G.; Klussmann, E.; Procino, G.; Svelto, M.; Rosenthal, W.; Valenti, G. CAMP-Induced AQP2 Translocation Is Associated with RhoA Inhibition through RhoA Phosphorylation and Interaction with RhoGDI. J. Cell. Sci. 2003, 116, 1519–1525. [Google Scholar] [CrossRef]
- Lu, H.A.J.; Sun, T.-X.; Matsuzaki, T.; Yi, X.-H.; Eswara, J.; Bouley, R.; McKee, M.; Brown, D. Heat Shock Protein 70 Interacts with Aquaporin-2 and Regulates Its Trafficking. J. Biol. Chem. 2007, 282, 28721–28732. [Google Scholar] [CrossRef]
- Zwang, N.A.; Hoffert, J.D.; Pisitkun, T.; Moeller, H.B.; Fenton, R.A.; Knepper, M.A. Identification of Phosphorylation-Dependent Binding Partners of Aquaporin-2 Using Protein Mass Spectrometry. J. Proteome. Res. 2009, 8, 1540–1554. [Google Scholar] [CrossRef]
- Centrone, M.; Ranieri, M.; Di Mise, A.; Berlingerio, S.P.; Russo, A.; Deen, P.M.T.; Staub, O.; Valenti, G.; Tamma, G. AQP2 Abundance Is Regulated by the E3-Ligase CHIP Via HSP70. Cell. Physiol. Biochem. 2017, 44, 515–531. [Google Scholar] [CrossRef]
- Wang, P.-J.; Lin, S.-T.; Liu, S.-H.; Kuo, K.-T.; Hsu, C.-H.; Knepper, M.A.; Yu, M.-J. Vasopressin-Induced Serine 269 Phosphorylation Reduces Sipa1l1 (Signal-Induced Proliferation-Associated 1 like 1)-Mediated Aquaporin-2 Endocytosis. J. Biol. Chem. 2017, 292, 7984–7993. [Google Scholar] [CrossRef]
- Noda, Y.; Horikawa, S.; Furukawa, T.; Hirai, K.; Katayama, Y.; Asai, T.; Kuwahara, M.; Katagiri, K.; Kinashi, T.; Hattori, M.; et al. Aquaporin-2 Trafficking Is Regulated by PDZ-Domain Containing Protein SPA-1. FEBS Lett. 2004, 568, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Nooh, M.M.; Kale, A.; Bahouth, S.W. Involve.ement of PDZ-SAP97 Interactions in Regulating AQP2 Translocation in Response to Vasopressin in LLC-PK1 Cells. Am. J. Physiol. Renal. Physiol. 2019, 317, F375–F387. [Google Scholar] [CrossRef]
- Okutsu, R.; Rai, T.; Kikuchi, A.; Ohno, M.; Uchida, K.; Sasaki, S.; Uchida, S. AKAP220 Colocalizes with AQP2 in the Inner Medullary Collecting Ducts. Kidney Int. 2008, 74, 1429–1433. [Google Scholar] [CrossRef]
- Klussmann, E.; Rosenthal, W. Role and Identification of Protein Kinase A Anchoring Proteins in Vasopressin-Mediated Aquaporin-2 Translocation. Kidney Int. 2001, 60, 446–449. [Google Scholar] [CrossRef]
- Moeller, H.B.; Slengerik-Hansen, J.; Aroankins, T.; Assentoft, M.; MacAulay, N.; Moestrup, S.K.; Bhalla, V.; Fenton, R.A. Regulation of the Water Channel Aquaporin-2 via 14-3-3θ and -ζ. J. Biol. Chem. 2016, 291, 2469–2484. [Google Scholar] [CrossRef]
- Murali, S.K.; Aroankins, T.S.; Moeller, H.B.; Fenton, R.A. The Deubiquitylase USP4 Interacts with the Water Channel AQP2 to Modulate Its Apical Membrane Accumulation and Cellular Abundance. Cells 2019, 8, 265. [Google Scholar] [CrossRef] [PubMed]
- Trimpert, C.; Wesche, D.; de Groot, T.; Pimentel Rodriguez, M.M.; Wong, V.; van den Berg, D.T.M.; Cheval, L.; Ariza, C.A.; Doucet, A.; Stagljar, I.; et al. NDFIP Allows NEDD4/NEDD4L-Induced AQP2 Ubiquitination and Degradation. PLoS ONE 2017, 12, e0183774. [Google Scholar] [CrossRef]
- van Balkom, B.W.M.; Boone, M.; Hendriks, G.; Kamsteeg, E.-J.; Robben, J.H.; Stronks, H.C.; van der Voorde, A.; van Herp, F.; van der Sluijs, P.; Deen, P.M.T. LIP5 Interacts with Aquaporin 2 and Facilitates Its Lysosomal Degradation. J. Am. Soc. Nephrol. 2009, 20, 990–1001. [Google Scholar] [CrossRef]
- Roche, J.V.; Survery, S.; Kreida, S.; Nesverova, V.; Ampah-Korsah, H.; Gourdon, M.; Deen, P.M.T.; Törnroth-Horsefield, S. Phosphorylation of Human Aquaporin 2 (AQP2) Allosterically Controls Its Interaction with the Lysosomal Trafficking Protein LIP5. J. Biol. Chem. 2017, 292, 14636–14648. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.V.; Nesverova, V.; Olsson, C.; Deen, P.M.; Törnroth-Horsefield, S. Structural Insights into AQP2 Targeting to Multivesicular Bodies. Int. J. Mol. Sci. 2019, 20, 5351. [Google Scholar] [CrossRef] [PubMed]
- Kamsteeg, E.-J.; Duffield, A.S.; Konings, I.B.M.; Spencer, J.; Pagel, P.; Deen, P.M.T.; Caplan, M.J. MAL Decreases the Internalization of the Aquaporin-2 Water Channel. Proc. Natl. Acad. Sci. USA 2007, 104, 16696–16701. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Suzuki, T.; Hagiwara, H.; Kuwahara, M.; Sasaki, S.; Takata, K.; Matsuzaki, T. Close Association of Aquaporin-2 Internalization with Caveolin-1. Acta Histochem. Cytochem. 2012, 45, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Noda, Y.; Horikawa, S.; Katayama, Y.; Sasaki, S. Water Channel Aquaporin-2 Directly Binds to Actin. Biochem. Biophys Res. Commun. 2004, 322, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Yui, N.; Noda, Y. Actin Directly Interacts with Different Membrane Channel Proteins and Influences Channel Activities: AQP2 as a Model. Biochim. Biophys Acta 2014, 1838, 514–520. [Google Scholar] [CrossRef] [PubMed]
- Moeller, H.B.; Praetorius, J.; Rützler, M.R.; Fenton, R.A. Phosphorylation of Aquaporin-2 Regulates Its Endocytosis and Protein-Protein Interactions. Proc. Natl. Acad. Sci. USA 2010, 107, 424–429. [Google Scholar] [CrossRef]
- Holst, M.R.; Jensen, L.G.; Aaron, J.; Login, F.H.; Rajkumar, S.; Hahn, U.; Nejsum, L.N. Regulated Exocytosis: Renal Aquaporin-2 3D Vesicular Network Organization and Association with F-Actin. Am. J. Physiol. Cell. Physiol. 2021, 321, C1060–C1069. [Google Scholar] [CrossRef]
- Noda, Y.; Horikawa, S.; Kanda, E.; Yamashita, M.; Meng, H.; Eto, K.; Li, Y.; Kuwahara, M.; Hirai, K.; Pack, C.; et al. Reciprocal Interaction with G-Actin and Tropomyosin Is Essential for Aquaporin-2 Trafficking. J. Cell. Biol. 2008, 182, 587–601. [Google Scholar] [CrossRef]
- Jang, K.-J.; Cho, H.S.; Kang, D.H.; Bae, W.G.; Kwon, T.-H.; Suh, K.-Y. Fluid-Shear-Stress-Induced Translocation of Aquaporin-2 and Reorganization of Actin Cytoskeleton in Renal Tubular Epithelial Cells. Integr. Biol. 2011, 3, 134–141. [Google Scholar] [CrossRef]
- Li, W.; Jin, W.W.; Tsuji, K.; Chen, Y.; Nomura, N.; Su, L.; Yui, N.; Arthur, J.; Cotecchia, S.; Paunescu, T.G.; et al. Ezrin Directly Interacts with AQP2 and Promotes Its Endocytosis. J. Cell. Sci. 2017, 130, 2914–2925. [Google Scholar] [CrossRef]
- Tamma, G.; Procino, G.; Mola, M.G.; Svelto, M.; Valenti, G. Functional Involvement of Annexin-2 in CAMP Induced AQP2 Trafficking. Pflugers. Arch. 2008, 456, 729–736. [Google Scholar] [CrossRef]
- Pizzoni, A.; López González, M.; Di Giusto, G.; Rivarola, V.; Capurro, C.; Ford, P. AQP2 Can Modulate the Pattern of Ca2+ Transients Induced by Store-Operated Ca2+ Entry under TRPV4 Activation. J. Cell. Biochem. 2018, 119, 4120–4133. [Google Scholar] [CrossRef]
- Mamenko, M.; Dhande, I.; Tomilin, V.; Zaika, O.; Boukelmoune, N.; Zhu, Y.; Gonzalez-Garay, M.L.; Pochynyuk, O.; Doris, P.A. Defective Store-Operated Calcium Entry Causes Partial Nephrogenic Diabetes Insipidus. J. Am. Soc. Nephrol. 2016, 27, 2035–2048. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Chen, L.; Zhang, X.; Zhou, Q.; Li, J.J.-M.; Berger, S.; Borok, Z.; Zhou, B.; Xiao, Z.; Yin, H.; et al. Aqp5 Is a New Transcriptional Target of Dot1a and a Regulator of Aqp2. PLoS ONE 2013, 8, e53342. [Google Scholar] [CrossRef] [PubMed]
- Rojek, A.; Praetorius, J.; Frøkiaer, J.; Nielsen, S.; Fenton, R.A. A Current View of the Mammalian Aquaglyceroporins. Annu. Rev. Physiol. 2008, 70, 301–327. [Google Scholar] [CrossRef]
- Calamita, G.; Delporte, C. Involvement of Aquaglyceroporins in Energy Metabolism in Health and Disease. Biochimie 2021, 188, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Verkman, A.S. Role of Aquaporins in Lung Liquid Physiology. Respir. Physiol. Neurobiol. 2007, 159, 324–330. [Google Scholar] [CrossRef]
- Yadav, E.; Yadav, N.; Hus, A.; Yadav, J.S. Aquaporins in Lung Health and Disease: Emerging Roles, Regulation, and Clinical Implications. Respir. Med. 2020, 174, 106193. [Google Scholar] [CrossRef] [PubMed]
- Yde, J.; Keely, S.J.; Moeller, H.B. Expression, Regulation and Function of Aquaporin-3 in Colonic Epithelial Cells. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183619. [Google Scholar] [CrossRef] [PubMed]
- Kortenoeven, M.L.A.; Fenton, R.A. Renal Aquaporins and Water Balance Disorders. Biochim. Biophys. Acta 2014, 1840, 1533–1549. [Google Scholar] [CrossRef]
- Bollag, W.B.; Aitkens, L.; White, J.; Hyndman, K.A. Aquaporin-3 in the Epidermis: More than Skin Deep. Am. J. Physiol. Cell. Physiol. 2020, 318, C1144–C1153. [Google Scholar] [CrossRef]
- Tricarico, P.M.; Mentino, D.; De Marco, A.; Del Vecchio, C.; Garra, S.; Cazzato, G.; Foti, C.; Crovella, S.; Calamita, G. Aquaporins Are One of the Critical Factors in the Disruption of the Skin Barrier in Inflammatory Skin Diseases. Int. J. Mol. Sci. 2022, 23, 4020. [Google Scholar] [CrossRef]
- Chiadak, J.D.; Gena, P.; Gregoire, F.; Bolaky, N.; Delforge, V.; Perret, J.; Calamita, G.; Delporte, C. Lipopolysaccharide Modifies Glycerol Permeability and Metabolism in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2017, 18, 2566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, H.; Liu, E.; Guang, Y.; Yang, L.; Mao, J.; Zhu, L.; Chen, L.; Wang, L. The AQP-3 Water Channel and the ClC-3 Chloride Channel Coordinate the Hypotonicity-Induced Swelling Volume in Nasopharyngeal Carcinoma Cells. Int. J. Biochem. Cell. Biol. 2014, 57, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Deng, Z.; Yang, L.; Luo, H.; Liu, S.; Li, Y.; Wei, Y.; Peng, S.; Zhu, L.; Wang, L.; et al. The AQP-3 Water Channel Is a Pivotal Modulator of Glycerol-Induced Chloride Channel Activation in Nasopharyngeal Carcinoma Cells. Int. J. Biochem. Cell. Biol. 2016, 72, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Sundelacruz, S.; Levin, M.; Kaplan, D.L. Role of Membrane Potential in the Regulation of Cell Proliferation and Differentiation. Stem. Cell. Rev. Rep. 2009, 5, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Bollinger Bollag, W. Aquaporin 3 Colocates with Phospholipase D2 in Caveolin-Rich Membrane Microdomains and Is Downregulated upon Keratinocyte Differentiation. J. Investig. Dermatol. 2003, 121, 1487–1495. [Google Scholar] [CrossRef]
- Reppetti, J.; Reca, A.; Seyahian, E.A.; Medina, Y.; Martínez, N.; Szpilbarg, N.; Damiano, A.E. Intact Caveolae Are Required for Proper Extravillous Trophoblast Migration and Differentiation. J. Cell. Physiol. 2020, 235, 3382–3392. [Google Scholar] [CrossRef]
- Huang, P.; Hansen, J.S.; Saba, K.H.; Bergman, A.; Negoita, F.; Gourdon, P.; Hagström-Andersson, A.; Lindkvist-Petersson, K. Aquaglyceroporins and Orthodox Aquaporins in Human Adipocytes. Biochim. Biophys. Acta. Biomembr. 2022, 1864, 183795. [Google Scholar] [CrossRef]
- Hansen, J.S.; Krintel, C.; Hernebring, M.; Haataja, T.J.K.; de Marè, S.; Wasserstrom, S.; Kosinska-Eriksson, U.; Palmgren, M.; Holm, C.; Stenkula, K.G.; et al. Perilipin 1 Binds to Aquaporin 7 in Human Adipocytes and Controls Its Mobility via Protein Kinase A Mediated Phosphorylation. Metabolism 2016, 65, 1731–1742. [Google Scholar] [CrossRef]
- Iena, F.M.; Kalucka, J.; Nielsen, L.; Søndergaard, E.; Nielsen, S.; Lebeck, J. Localization of Aquaglyceroporins in Human and Murine White Adipose Tissue. Histochem. Cell. Biol. 2022, 157, 623–639. [Google Scholar] [CrossRef]
- Takata, K.; Matsuzaki, T.; Tajika, Y. Aquaporins: Water Channel Proteins of the Cell Membrane. Prog. Histochem. Cytochem. 2004, 39, 1–83. [Google Scholar] [CrossRef]
- Kitchen, P.; Salman, M.M.; Halsey, A.M.; Clarke-Bland, C.; MacDonald, J.A.; Ishida, H.; Vogel, H.J.; Almutiri, S.; Logan, A.; Kreida, S.; et al. Targeting Aquaporin-4 Subcellular Localization to Treat Central Nervous System Edema. Cell 2020, 181, 784–799.e19. [Google Scholar] [CrossRef] [PubMed]
- Plog, B.A.; Nedergaard, M. The Glymphatic System in Central Nervous System Health and Disease: Past, Present, and Future. Annu. Rev. Pathol. 2018, 13, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.J.; Jin, B.-J.; Ratelade, J.; Verkman, A.S. Aggregation State Determines the Localization and Function of M1- and M23-Aquaporin-4 in Astrocytes. J. Cell. Biol. 2014, 204, 559–573. [Google Scholar] [CrossRef] [PubMed]
- Hiroaki, Y.; Tani, K.; Kamegawa, A.; Gyobu, N.; Nishikawa, K.; Suzuki, H.; Walz, T.; Sasaki, S.; Mitsuoka, K.; Kimura, K.; et al. Implications of the Aquaporin-4 Structure on Array Formation and Cell Adhesion. J. Mol. Biol. 2006, 355, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Neely, J.D.; Amiry-Moghaddam, M.; Ottersen, O.P.; Froehner, S.C.; Agre, P.; Adams, M.E. Syntrophin-Dependent Expression and Localization of Aquaporin-4 Water Channel Protein. Proc. Natl. Acad. Sci. USA 2001, 98, 14108–14113. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Rossi, A.; Nudel, U.; Svelto, M.; Frigeri, A. Dystrophin-Dependent and -Independent AQP4 Pools Are Expressed in the Mouse Brain. Glia 2008, 56, 869–876. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Rossi, A.; Mola, M.G.; Procino, G.; Frigeri, A.; Svelto, M. Actin Cytoskeleton Remodeling Governs Aquaporin-4 Localization in Astrocytes. Glia 2008, 56, 1755–1766. [Google Scholar] [CrossRef]
- Nicchia, G.P.; Srinivas, M.; Li, W.; Brosnan, C.F.; Frigeri, A.; Spray, D.C. New Possible Roles for Aquaporin-4 in Astrocytes: Cell Cytoskeleton and Functional Relationship with Connexin43. FASEB J. 2005, 19, 1674–1676. [Google Scholar] [CrossRef]
- Benfenati, V.; Caprini, M.; Dovizio, M.; Mylonakou, M.N.; Ferroni, S.; Ottersen, O.P.; Amiry-Moghaddam, M. An Aquaporin-4/Transient Receptor Potential Vanilloid 4 (AQP4/TRPV4) Complex Is Essential for Cell-Volume Control in Astrocytes. Proc. Natl. Acad. Sci. USA 2011, 108, 2563–2568. [Google Scholar] [CrossRef]
- Jo, A.O.; Ryskamp, D.A.; Phuong, T.T.T.; Verkman, A.S.; Yarishkin, O.; MacAulay, N.; Križaj, D. TRPV4 and AQP4 Channels Synergistically Regulate Cell Volume and Calcium Homeostasis in Retinal Müller Glia. J. Neurosci. 2015, 35, 13525–13537. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.M.; Blazer-Yost, B. Channels and Transporters in Astrocyte Volume Regulation in Health and Disease. Cell. Physiol. Biochem. 2022, 56, 12–30. [Google Scholar] [CrossRef] [PubMed]
- Vennekens, R.; Nilius, B. Insights into TRPM4 Function, Regulation and Physiological Role. Handb. Exp. Pharmacol. 2007, 269–285. [Google Scholar] [CrossRef]
- Stokum, J.A.; Kwon, M.S.; Woo, S.K.; Tsymbalyuk, O.; Vennekens, R.; Gerzanich, V.; Simard, J.M. SUR1-TRPM4 and AQP4 Form a Heteromultimeric Complex That Amplifies Ion/Water Osmotic Coupling and Drives Astrocyte Swelling. Glia 2018, 66, 108–125. [Google Scholar] [CrossRef] [PubMed]
- Jha, R.M.; Rani, A.; Desai, S.M.; Raikwar, S.; Mihaljevic, S.; Munoz-Casabella, A.; Kochanek, P.M.; Catapano, J.; Winkler, E.; Citerio, G.; et al. Sulfonylurea Receptor 1 in Central Nervous System Injury: An Updated Review. Int J. Mol. Sci. 2021, 22, 11899. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, P.; Öberg, F.; Sjöhamn, J.; Hedfalk, K.; Bill, R.M.; Conner, A.C.; Conner, M.T.; Törnroth-Horsefield, S. Plasma Membrane Abundance of Human Aquaporin 5 Is Dynamically Regulated by Multiple Pathways. PLoS ONE 2015, 10, e0143027. [Google Scholar] [CrossRef]
- Ishida, H.; Vogel, H.J.; Conner, A.C.; Kitchen, P.; Bill, R.M.; MacDonald, J.A. Simultaneous Binding of the N- and C-Terminal Cytoplasmic Domains of Aquaporin 4 to Calmodulin. Biochim. Biophys. Acta. Biomembr. 2022, 1864, 183837. [Google Scholar] [CrossRef]
- Michele, D.E.; Campbell, K.P. Dystrophin-Glycoprotein Complex: Post-Translational Processing and Dystroglycan Function. J. Biol. Chem. 2003, 278, 15457–15460. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Williamson, A.; Palomba, M.; Eid, T.; de Lanerolle, N.C.; Nagelhus, E.A.; Adams, M.E.; Froehner, S.C.; Agre, P.; Ottersen, O.P. Delayed K+ Clearance Associated with Aquaporin-4 Mislocalization: Phenotypic Defects in Brains of Alpha-Syntrophin-Null Mice. Proc. Natl. Acad. Sci. USA 2003, 100, 13615–13620. [Google Scholar] [CrossRef]
- Amiry-Moghaddam, M.; Xue, R.; Haug, F.-M.; Neely, J.D.; Bhardwaj, A.; Agre, P.; Adams, M.E.; Froehner, S.C.; Mori, S.; Ottersen, O.P. Alpha-Syntrophin Deletion Removes the Perivascular but Not Endothelial Pool of Aquaporin-4 at the Blood-Brain Barrier and Delays the Development of Brain Edema in an Experimental Model of Acute Hyponatremia. FASEB J. 2004, 18, 542–544. [Google Scholar] [CrossRef]
- Pannicke, T.; Iandiev, I.; Uckermann, O.; Biedermann, B.; Kutzera, F.; Wiedemann, P.; Wolburg, H.; Reichenbach, A.; Bringmann, A. A Potassium Channel-Linked Mechanism of Glial Cell Swelling in the Postischemic Retina. Mol. Cell. Neurosci. 2004, 26, 493–502. [Google Scholar] [CrossRef]
- Ruiz-Ederra, J.; Zhang, H.; Verkman, A.S. Evidence against Functional Interaction between Aquaporin-4 Water Channels and Kir4.1 Potassium Channels in Retinal Müller Cells. J. Biol. Chem. 2007, 282, 21866–21872. [Google Scholar] [CrossRef] [PubMed]
- Strohschein, S.; Hüttmann, K.; Gabriel, S.; Binder, D.K.; Heinemann, U.; Steinhäuser, C. Impact of Aquaporin-4 Channels on K+ Buffering and Gap Junction Coupling in the Hippocampus. Glia 2011, 59, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, S.; Zhang, K.; Wang, H.; Lan, L.; Ma, X.; Liu, X.; Zhang, S.; Zheng, J.; Wei, X.; et al. Aquaporin 4 Forms a Macromolecular Complex with Glutamate Transporter 1 and Mu Opioid Receptor in Astrocytes and Participates in Morphine Dependence. J. Mol. Neurosci. 2017, 62, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Illarionova, N.B.; Gunnarson, E.; Li, Y.; Brismar, H.; Bondar, A.; Zelenin, S.; Aperia, A. Functional and Molecular Interactions between Aquaporins and Na,K-ATPase. Neuroscience 2010, 168, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Jesus, T.T.; Bernardino, R.L.; Martins, A.D.; Sá, R.; Sousa, M.; Alves, M.G.; Oliveira, P.F. Aquaporin-4 as a Molecular Partner of Cystic Fibrosis Transmembrane Conductance Regulator in Rat Sertoli Cells. Biochem. Biophys. Res. Commun. 2014, 446, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Madrid, R.; Le Maout, S.; Barrault, M.B.; Janvier, K.; Benichou, S.; Mérot, J. Polarized Trafficking and Surface Expression of the AQP4 Water Channel Are Coordinated by Serial and Regulated Interactions with Different Clathrin-Adaptor Complexes. EMBO J. 2001, 20, 7008–7021. [Google Scholar] [CrossRef]
- Wallisch, J.S.; Janesko-Feldman, K.; Alexander, H.; Jha, R.M.; Farr, G.W.; McGuirk, P.R.; Kline, A.E.; Jackson, T.C.; Pelletier, M.F.; Clark, R.S.B.; et al. The Aquaporin-4 Inhibitor AER-271 Blocks Acute Cerebral Edema and Improves Early Outcome in a Pediatric Model of Asphyxial Cardiac Arrest. Pediatr. Res. 2019, 85, 511–517. [Google Scholar] [CrossRef]
- Verkman, A.S.; Smith, A.J.; Phuan, P.-W.; Tradtrantip, L.; Anderson, M.O. The Aquaporin-4 Water Channel as a Potential Drug Target in Neurological Disorders. Expert. Opin. Ther. Targets 2017, 21, 1161–1170. [Google Scholar] [CrossRef]
- D’Agostino, C.; Elkashty, O.A.; Chivasso, C.; Perret, J.; Tran, S.D.; Delporte, C. Insight into Salivary Gland Aquaporins. Cells 2020, 9, 1547. [Google Scholar] [CrossRef]
- Mobasheri, A.; Barrett-Jolley, R. Aquaporin Water Channels in the Mammary Gland: From Physiology to Pathophysiology and Neoplasia. J. Mammary Gland. Biol. Neoplasia 2014, 19, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Verkman, A.S. Aquaporin-5 Dependent Fluid Secretion in Airway Submucosal Glands. J. Biol. Chem. 2001, 276, 41288–41292. [Google Scholar] [CrossRef] [PubMed]
- Grey, A.C.; Walker, K.L.; Petrova, R.S.; Han, J.; Wilmarth, P.A.; David, L.L.; Donaldson, P.J.; Schey, K.L. Verification and Spatial Localization of Aquaporin-5 in the Ocular Lens. Exp. Eye Res. 2013, 108, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Petrova, R.S.; Schey, K.L.; Donaldson, P.J.; Grey, A.C. Spatial Distributions of AQP5 and AQP0 in Embryonic and Postnatal Mouse Lens Development. Exp. Eye Res. 2015, 132, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Matsui, M.; Uchida, K.; Futatsugi, A.; Kusakawa, S.; Matsumoto, N.; Nakamura, K.; Manabe, T.; Taketo, M.M.; Mikoshiba, K. M(3) Muscarinic Acetylcholine Receptor Plays a Critical Role in Parasympathetic Control of Salivation in Mice. J. Physiol. 2004, 558, 561–575. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Yuan, Z.; Inoue, N.; Skowronski, M.T.; Nakae, Y.; Shono, M.; Cho, G.; Yasui, M.; Agre, P.; Nielsen, S. Identification of AQP5 in Lipid Rafts and Its Translocation to Apical Membranes by Activation of M3 MAChRs in Interlobular Ducts of Rat Parotid Gland. Am. J. Physiol. Cell. Physiol. 2005, 289, C1303–C1311. [Google Scholar] [CrossRef]
- Hasegawa, T.; Azlina, A.; Javkhlan, P.; Yao, C.; Akamatsu, T.; Hosoi, K. Novel Phosphorylation of Aquaporin-5 at Its Threonine 259 through CAMP Signaling in Salivary Gland Cells. Am. J. Physiol. Cell. Physiol. 2011, 301, C667–C678. [Google Scholar] [CrossRef]
- Hwang, S.; Kang, J.Y.; Kim, M.J.; Shin, D.M.; Hong, J.H. Carbonic Anhydrase 12 Mutation Modulates Membrane Stability and Volume Regulation of Aquaporin 5. J. Enzym. Inhib. Med. Chem. 2019, 34, 179–188. [Google Scholar] [CrossRef]
- Liu, X.; Bandyopadhyay, B.C.; Bandyopadhyay, B.; Nakamoto, T.; Singh, B.; Liedtke, W.; Melvin, J.E.; Ambudkar, I. A Role for AQP5 in Activation of TRPV4 by Hypotonicity: Concerted Involvement of AQP5 and TRPV4 in Regulation of Cell Volume Recovery. J. Biol. Chem. 2006, 281, 15485–15495. [Google Scholar] [CrossRef]
- Bhattacharya, D.; Yu, L.; Wang, M. Expression Patterns of Conjunctival Mucin 5AC and Aquaporin 5 in Response to Acute Dry Eye Stress. PLoS ONE 2017, 12, e0187188. [Google Scholar] [CrossRef]
- Ohashi, Y.; Tsuzaka, K.; Takeuchi, T.; Sasaki, Y.; Tsubota, K. Altered Distribution of Aquaporin 5 and Its C-Terminal Binding Protein in the Lacrimal Glands of a Mouse Model for Sjögren’s Syndrome. Curr. Eye Res. 2008, 33, 621–629. [Google Scholar] [CrossRef]
- Chivasso, C.; Nesverova, V.; Järvå, M.; Blanchard, A.; Rose, K.L.; Öberg, F.K.; Wang, Z.; Martin, M.; Lhotellerie, F.; Zindy, E.; et al. Unraveling Human AQP5-PIP Molecular Interaction and Effect on AQP5 Salivary Glands Localization in SS Patients. Cells 2021, 10, 2108. [Google Scholar] [CrossRef]
- Chivasso, C.; Hagströmer, C.J.; Rose, K.L.; Lhotellerie, F.; Leblanc, L.; Wang, Z.; Moscato, S.; Chevalier, C.; Zindy, E.; Martin, M.; et al. Ezrin Is a Novel Protein Partner of Aquaporin-5 in Human Salivary Glands and Shows Altered Expression and Cellular Localization in Sjögren’s Syndrome. Int. J. Mol. Sci. 2021, 22, 9213. [Google Scholar] [CrossRef] [PubMed]
- Terawaki, S.; Maesaki, R.; Hakoshima, T. Structural Basis for NHERF Recognition by ERM Proteins. Structure 2006, 14, 777–789. [Google Scholar] [CrossRef]
- Muroi, S.-I.; Isohama, Y. Ezrin Regulates Ca2+ Ionophore-Induced Plasma Membrane Translocation of Aquaporin-5. Int. J. Mol. Sci. 2021, 22, 13505. [Google Scholar] [CrossRef]
- Gletten, R.B.; Cantrell, L.S.; Bhattacharya, S.; Schey, K.L. Lens Aquaporin-5 Inserts Into Bovine Fiber Cell Plasma Membranes Via Unconventional Protein Secretion. Invest. Ophthalmol. Vis. Sci. 2022, 63, 5. [Google Scholar] [CrossRef]
- Petrova, R.S.; Bavana, N.; Zhao, R.; Schey, K.L.; Donaldson, P.J. Changes to Zonular Tension Alters the Subcellular Distribution of AQP5 in Regions of Influx and Efflux of Water in the Rat Lens. Invest. Ophthalmol. Vis. Sci. 2020, 61, 36. [Google Scholar] [CrossRef] [PubMed]
- Yasui, M.; Kwon, T.H.; Knepper, M.A.; Nielsen, S.; Agre, P. Aquaporin-6: An Intracellular Vesicle Water Channel Protein in Renal Epithelia. Proc. Natl. Acad. Sci. USA 1999, 96, 5808–5813. [Google Scholar] [CrossRef]
- Yasui, M.; Hazama, A.; Kwon, T.H.; Nielsen, S.; Guggino, W.B.; Agre, P. Rapid Gating and Anion Permeability of an Intracellular Aquaporin. Nature 1999, 402, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Rabaud, N.E.; Song, L.; Wang, Y.; Agre, P.; Yasui, M.; Carbrey, J.M. Aquaporin 6 Binds Calmodulin in a Calcium-Dependent Manner. Biochem. Biophys. Res. Commun. 2009, 383, 54–57. [Google Scholar] [CrossRef]
- Beitz, E.; Liu, K.; Ikeda, M.; Guggino, W.B.; Agre, P.; Yasui, M. Determinants of AQP6 Trafficking to Intracellular Sites versus the Plasma Membrane in Transfected Mammalian Cells. Biol. Cell. 2006, 98, 101–109. [Google Scholar] [CrossRef]
- Zhang, J.; Hupfeld, C.J.; Taylor, S.S.; Olefsky, J.M.; Tsien, R.Y. Insulin Disrupts Beta-Adrenergic Signalling to Protein Kinase A in Adipocytes. Nature 2005, 437, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Kuwahara, M.; Gu, Y.; Kageyama, Y.; Tohsaka, A.; Suzuki, F.; Marumo, F.; Sasaki, S. Cloning and Functional Expression of a New Water Channel Abundantly Expressed in the Testis Permeable to Water, Glycerol, and Urea. J. Biol. Chem. 1997, 272, 20782–20786. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Yamamoto, T.; Kondo, D.; Funaki, H.; Yaoita, E.; Kawasaki, K.; Sato, N.; Hatakeyama, K.; Kihara, I. Molecular Cloning of a New Aquaporin from Rat Pancreas and Liver. J. Biol. Chem. 1997, 272, 30329–30333. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Yang, B.; Verkman, A.S. Cloning of a Novel Water and Urea-Permeable Aquaporin from Mouse Expressed Strongly in Colon, Placenta, Liver, and Heart. Biochem. Biophys. Res. Commun. 1997, 240, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Calamita, G.; Ferri, D.; Gena, P.; Liquori, G.E.; Cavalier, A.; Thomas, D.; Svelto, M. The Inner Mitochondrial Membrane Has Aquaporin-8 Water Channels and Is Highly Permeable to Water. J. Biol. Chem. 2005, 280, 17149–17153. [Google Scholar] [CrossRef]
- Ikaga, R.; Namekata, I.; Kotiadis, V.N.; Ogawa, H.; Duchen, M.R.; Tanaka, H.; Iida-Tanaka, N. Knockdown of Aquaporin-8 Induces Mitochondrial Dysfunction in 3T3-L1 Cells. Biochem. Biophys. Rep. 2015, 4, 187–195. [Google Scholar] [CrossRef]
- Varadaraj, K.; Kumari, S.S. Lens Aquaporins Function as Peroxiporins to Facilitate Membrane Transport of Hydrogen Peroxide. Biochem. Biophys. Res. Commun. 2020, 524, 1025–1029. [Google Scholar] [CrossRef]
- Wang, H.; Schoebel, S.; Schmitz, F.; Dong, H.; Hedfalk, K. Characterization of Aquaporin-Driven Hydrogen Peroxide Transport. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183065. [Google Scholar] [CrossRef]
- Miller, E.W.; Dickinson, B.C.; Chang, C.J. Aquaporin-3 Mediates Hydrogen Peroxide Uptake to Regulate Downstream Intracellular Signaling. Proc. Natl. Acad. Sci. USA 2010, 107, 15681–15686. [Google Scholar] [CrossRef]
- Rodrigues, C.; Mósca, A.F.; Martins, A.P.; Nobre, T.; Prista, C.; Antunes, F.; Cipak Gasparovic, A.; Soveral, G. Rat Aquaporin-5 Is PH-Gated Induced by Phosphorylation and Is Implicated in Oxidative Stress. Int. J. Mol. Sci. 2016, 17, 2090. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Moniaga, C.S.; Nielsen, S.; Hara-Chikuma, M. Aquaporin-9 Facilitates Membrane Transport of Hydrogen Peroxide in Mammalian Cells. Biochem. Biophys. Res. Commun. 2016, 471, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Bestetti, S.; Galli, M.; Sorrentino, I.; Pinton, P.; Rimessi, A.; Sitia, R.; Medraño-Fernandez, I. Human Aquaporin-11 Guarantees Efficient Transport of H2O2 across the Endoplasmic Reticulum Membrane. Redox. Biol. 2019, 28, 101326. [Google Scholar] [CrossRef]
- Bienert, G.P.; Møller, A.L.B.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific Aquaporins Facilitate the Diffusion of Hydrogen Peroxide across Membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Agemark, M.; Kowal, J.; Kukulski, W.; Nordén, K.; Gustavsson, N.; Johanson, U.; Engel, A.; Kjellbom, P. Reconstitution of Water Channel Function and 2D-Crystallization of Human Aquaporin 8. Biochim. Biophys. Acta 2012, 1818, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Almasalmeh, A.; Krenc, D.; Wu, B.; Beitz, E. Structural Determinants of the Hydrogen Peroxide Permeability of Aquaporins. FEBS J. 2014, 281, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Soria, L.R.; Fanelli, E.; Altamura, N.; Svelto, M.; Marinelli, R.A.; Calamita, G. Aquaporin-8-Facilitated Mitochondrial Ammonia Transport. Biochem. Biophys. Res. Commun. 2010, 393, 217–221. [Google Scholar] [CrossRef]
- Saparov, S.M.; Liu, K.; Agre, P.; Pohl, P. Fast and Selective Ammonia Transport by Aquaporin-8. J. Biol. Chem. 2007, 282, 5296–5301. [Google Scholar] [CrossRef] [PubMed]
- Pellavio, G.; Todaro, F.; Alberizzi, P.; Scotti, C.; Gastaldi, G.; Lolicato, M.; Omes, C.; Caliogna, L.; Nappi, R.E.; Laforenza, U. HPV Infection Affects Human Sperm Functionality by Inhibition of Aquaporin-8. Cells 2020, 9, 1241. [Google Scholar] [CrossRef]
- Mori, S.; Kurimoto, T.; Miki, A.; Maeda, H.; Kusuhara, S.; Nakamura, M. Aqp9 Gene Deletion Enhances Retinal Ganglion Cell (RGC) Death and Dysfunction Induced by Optic Nerve Crush: Evidence That Aquaporin 9 Acts as an Astrocyte-to-Neuron Lactate Shuttle in Concert with Monocarboxylate Transporters To Support RGC Function and Survival. Mol. Neurobiol. 2020, 57, 4530–4548. [Google Scholar] [CrossRef]
- Rambow, J.; Wu, B.; Rönfeldt, D.; Beitz, E. Aquaporins with Anion/Monocarboxylate Permeability: Mechanisms, Relevance for Pathogen-host Interactions. Front. Pharmacol. 2014, 5, 199. [Google Scholar] [CrossRef] [Green Version]
- Jesus, T.T.; Bernardino, R.L.; Martins, A.D.; Sá, R.; Sousa, M.; Alves, M.G.; Oliveira, P.F. Aquaporin-9 Is Expressed in Rat Sertoli Cells and Interacts with the Cystic Fibrosis Transmembrane Conductance Regulator. IUBMB Life 2014, 66, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Pietrement, C.; Da Silva, N.; Silberstein, C.; James, M.; Marsolais, M.; Van Hoek, A.; Brown, D.; Pastor-Soler, N.; Ameen, N.; Laprade, R.; et al. Role of NHERF1, Cystic Fibrosis Transmembrane Conductance Regulator, and CAMP in the Regulation of Aquaporin 9. J. Biol. Chem. 2008, 283, 2986–2996. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.H.; Leung, C.T.; Leung, G.P.H.; Wong, P.Y.D. Synergistic Effects of Cystic Fibrosis Transmembrane Conductance Regulator and Aquaporin-9 in the Rat Epididymis. Biol. Reprod. 2003, 68, 1505–1510. [Google Scholar] [CrossRef] [PubMed]
- Bieth, E.; Hamdi, S.M.; Mieusset, R. Genetics of the Congenital Absence of the Vas Deferens. Hum. Genet. 2021, 140, 59–76. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, T.M.; van Beek, L.; Shilliday, F.; Debreczeni, J.É.; Phillips, C. Cryo-EM: The Resolution Revolution and Drug Discovery. SLAS Discov. 2021, 26, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Abir-Awan, M.; Kitchen, P.; Salman, M.M.; Conner, M.T.; Conner, A.C.; Bill, R.M. Inhibitors of Mammalian Aquaporin Water Channels. Int. J. Mol. Sci. 2019, 20, 1589. [Google Scholar] [CrossRef]
- Verkman, A.S.; Anderson, M.O.; Papadopoulos, M.C. Aquaporins: Important but Elusive Drug Targets. Nat. Rev. Drug Discov. 2014, 13, 259–277. [Google Scholar] [CrossRef]
- Soveral, G.; Casini, A. Aquaporin Modulators: A Patent Review (2010–2015). Expert Opin. Ther. Pat. 2017, 27, 49–62. [Google Scholar] [CrossRef]
- Pimpão, C.; Wragg, D.; da Silva, I.V.; Casini, A.; Soveral, G. Aquaglyceroporin Modulators as Emergent Pharmacological Molecules for Human Diseases. Front. Mol. Biosci. 2022, 9, 845237. [Google Scholar] [CrossRef]
- Salman, M.M.; Kitchen, P.; Yool, A.J.; Bill, R.M. Recent Breakthroughs and Future Directions in Drugging Aquaporins. Trends Pharmacol. Sci. 2022, 43, 30–42. [Google Scholar] [CrossRef]
- Girsch, S.J.; Peracchia, C. Calmodulin Interacts with a C-Terminus Peptide from the Lens Membrane Protein MIP26. Curr. Eye Res. 1991, 10, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.-J.; Jang, H.-J.; Park, E.; Tingskov, S.J.; Nørregaard, R.; Jung, H.J.; Kwon, T.-H. Sorting Nexin 27 Regulates the Lysosomal Degradation of Aquaporin-2 Protein in the Kidney Collecting Duct. Cells 2020, 9, 1208. [Google Scholar] [CrossRef] [PubMed]
- Connors, N.C.; Kofuji, P. Potassium Channel Kir4.1 Macromolecular Complex in Retinal Glial Cells. Glia 2006, 53, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Fort, P.E.; Sene, A.; Pannicke, T.; Roux, M.J.; Forster, V.; Mornet, D.; Nudel, U.; Yaffe, D.; Reichenbach, A.; Sahel, J.A.; et al. Kir4.1 and AQP4 Associate with Dp71- and Utrophin-DAPs Complexes in Specific and Defined Microdomains of Müller Retinal Glial Cell Membrane. Glia 2008, 56, 597–610. [Google Scholar] [CrossRef] [PubMed]
- Nicchia, G.P.; Cogotzi, L.; Rossi, A.; Basco, D.; Brancaccio, A.; Svelto, M.; Frigeri, A. Expression of Multiple AQP4 Pools in the Plasma Membrane and Their Association with the Dystrophin Complex. J. Neurochem. 2008, 105, 2156–2165. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AQP | Interacting Partner | AQP Region Involved in PPI | Detection Method |
|---|---|---|---|
| AQP0 | CaM | C-term [27,191] | CL-EM [23]; MST [25]; CL-MS [27] |
| ezrin | C-term [31] | CL-MS [31] | |
| BFSP1 (filensin) | C-term [35,36] | AFC [35] CL-MS [36] | |
| BFSP2 (CP49) | C-term [35] | AFC [35] | |
| connexins | C-term [40] | IP [40] | |
| AKAP2 | C-term [28] | coIP [28] | |
| crystallins | C-term [46] | IP, YTH [46]; CL-MS [47] | |
| AQP1 | Lin7 | ND | IP [48] |
| FAK | ND | IP [49] | |
| ß-catenin | ND | IP [49] | |
| APP | N-term [56] | IP, OVL [56] | |
| AQP2 | ezrin | C-term [88] | IP [88] |
| Hsc70 | C-term [66] | IP, PDA [66,67] | |
| Hsp70 | C-term [68] | IP [67,68] | |
| Clathrin | ND | IP [66] | |
| AP2 | ND | IP [66] | |
| Dynamin | ND | IP [66] | |
| Sipa1l1 | C-term [69] | IP [69] | |
| 14-3-3 θ/ζ | C-term [74] | IP [74] | |
| NEDD4 | ND | IP [76] | |
| NEDFIP 1/2 | ND | IP [76] | |
| USP4 | ND | IP [75] | |
| LIP5 | C-term [78] | FS [79], MST [25,78], YTH [77] | |
| SNX27 | C-term [192] | IP, PDA [192] | |
| TRPV4 | ND | IP [90] | |
| SK3 | ND | IP [90] | |
| Caveolin-1 | ND | IP [81] | |
| MAL | C-term [80] | IP [80] | |
| Spa1 | ND | IP [70] | |
| AKAP220 | ND | YTH [72] | |
| Annexin II | C-term [67] | IP [67] | |
| actin | C-term [85] | IP [82,84] | |
| TM5b | C-term [86] | IP [67] | |
| AQP5 | ND | IP [92] | |
| AQP3 | ClC3 | ND | IP [102] |
| PDL2 | ND | IP [105] | |
| Perilipin-1 | ND | BN-PAGE [107] | |
| Caveolin-1 | ND | IP [106] | |
| AQP4 | AQP4 | IC loop D, TM2 bottom [6] | FRET, MDS [6] |
| CaM | C-term [111,126]; N-term [126] | MST [111]; NMR [126] | |
| DGC | ND | IP [193,194,195] | |
| Kir4.1 | ND | IP [193] | |
| TRPV4 | ND | IP [119] | |
| TRPM4 | ND | IP, FRET [123] | |
| SUR1 | ND | IP, FRET [123] | |
| CFTR | ND | IP [135] | |
| GLT1 | C-term | IP, FRET [133] | |
| MOR | ND | IP [133] | |
| Na,K-ATPase | N-term [134] | IP, FRET [134] | |
| mGluR5 | N-term [134] | IP, FRET [134] | |
| µAP 2/3 | C-term [136] | YTH [136] | |
| AQP5 | PIP | C-term [150,151] | PDA [150], PLA, MST [151] |
| ezrin | C-term [152] | SILAC-IP-MS, PLA [152] | |
| NKCC1 | ND | IP [147] | |
| TRPV4 | N-term [148] | IP [148] | |
| Mucin 5AC | ND | IP [149] | |
| AE2 | ND | IP [147] | |
| AQP6 | CaM | N-term [157] | PDA [157] |
| AQP7 | Perilipin-1 | N-term [108] | PDA, PLA [108] |
| AQP8 | HPVL1 | ND | IP [178] |
| AQP9 | CFTR | ND | IP [181] |
| NHERF1 | C-term [182] | PDA, OVL [182] | |
| MCTs | ND | IP [179] | |
| AQP10 | ND | ND | - |
| AQP11 | ND | ND | - |
| AQP12 | ND | ND | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Törnroth-Horsefield, S.; Chivasso, C.; Strandberg, H.; D’Agostino, C.; O’Neale, C.V.T.; Schey, K.L.; Delporte, C. Insight into the Mammalian Aquaporin Interactome. Int. J. Mol. Sci. 2022, 23, 9615. https://doi.org/10.3390/ijms23179615
Törnroth-Horsefield S, Chivasso C, Strandberg H, D’Agostino C, O’Neale CVT, Schey KL, Delporte C. Insight into the Mammalian Aquaporin Interactome. International Journal of Molecular Sciences. 2022; 23(17):9615. https://doi.org/10.3390/ijms23179615
Chicago/Turabian StyleTörnroth-Horsefield, Susanna, Clara Chivasso, Helin Strandberg, Claudia D’Agostino, Carla V. T. O’Neale, Kevin L. Schey, and Christine Delporte. 2022. "Insight into the Mammalian Aquaporin Interactome" International Journal of Molecular Sciences 23, no. 17: 9615. https://doi.org/10.3390/ijms23179615