
Hcp of the Type VI Secretion System (T6SS) in Acidovorax citrulli Group II Strain Aac5 Has a Dual Role as a Core Structural Protein and an Effector Protein in Colonization, Growth Ability, Competition, Biofilm Formation, and Ferric Iron Absorption
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
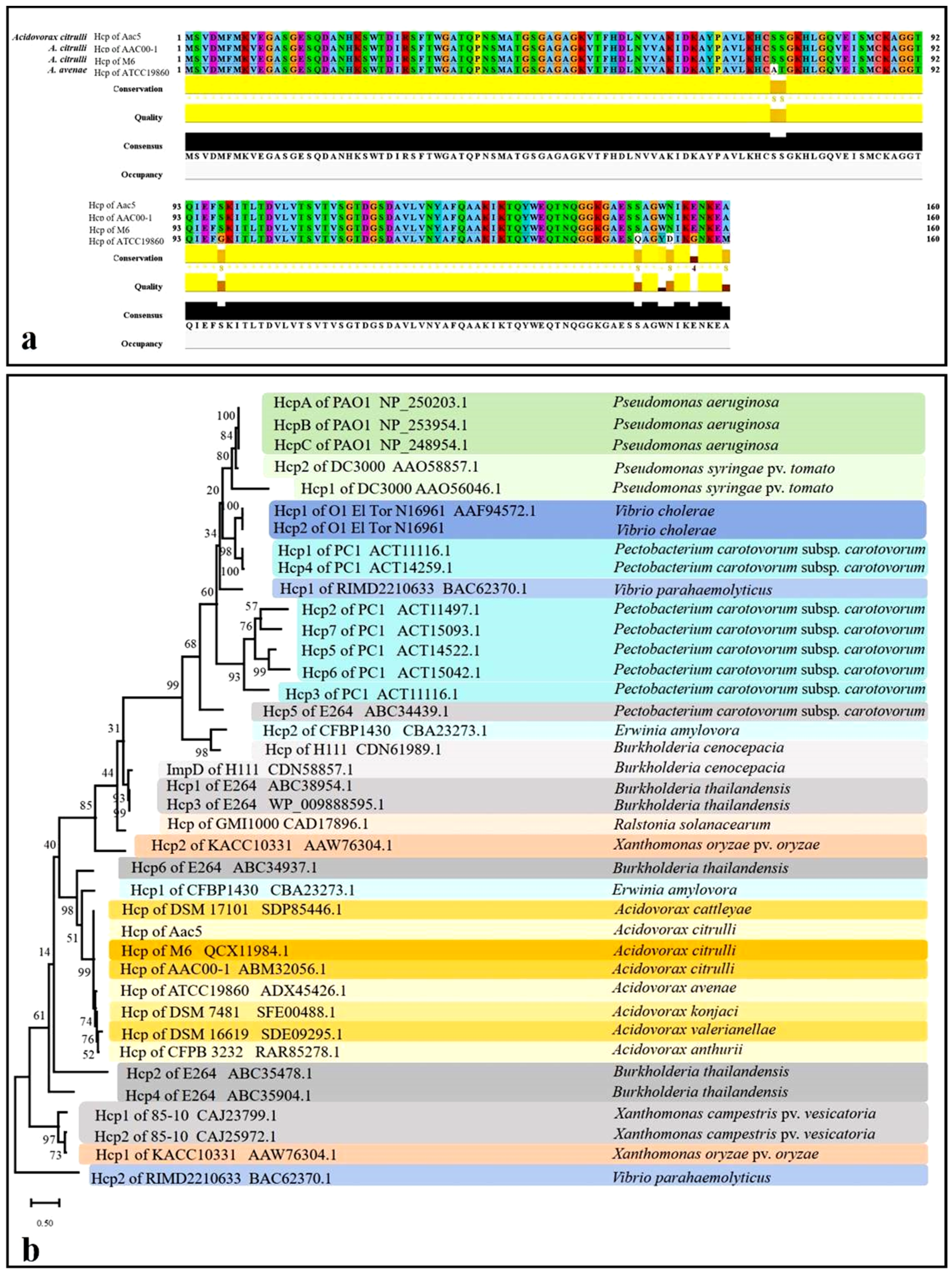
2.1. Sequence Analysis of the T6SS and Hcp Homolog in A. citrulli
2.2. Construction of Δhcp, Δhcpcomp, Δtssm, and ΔhcpΔtssm
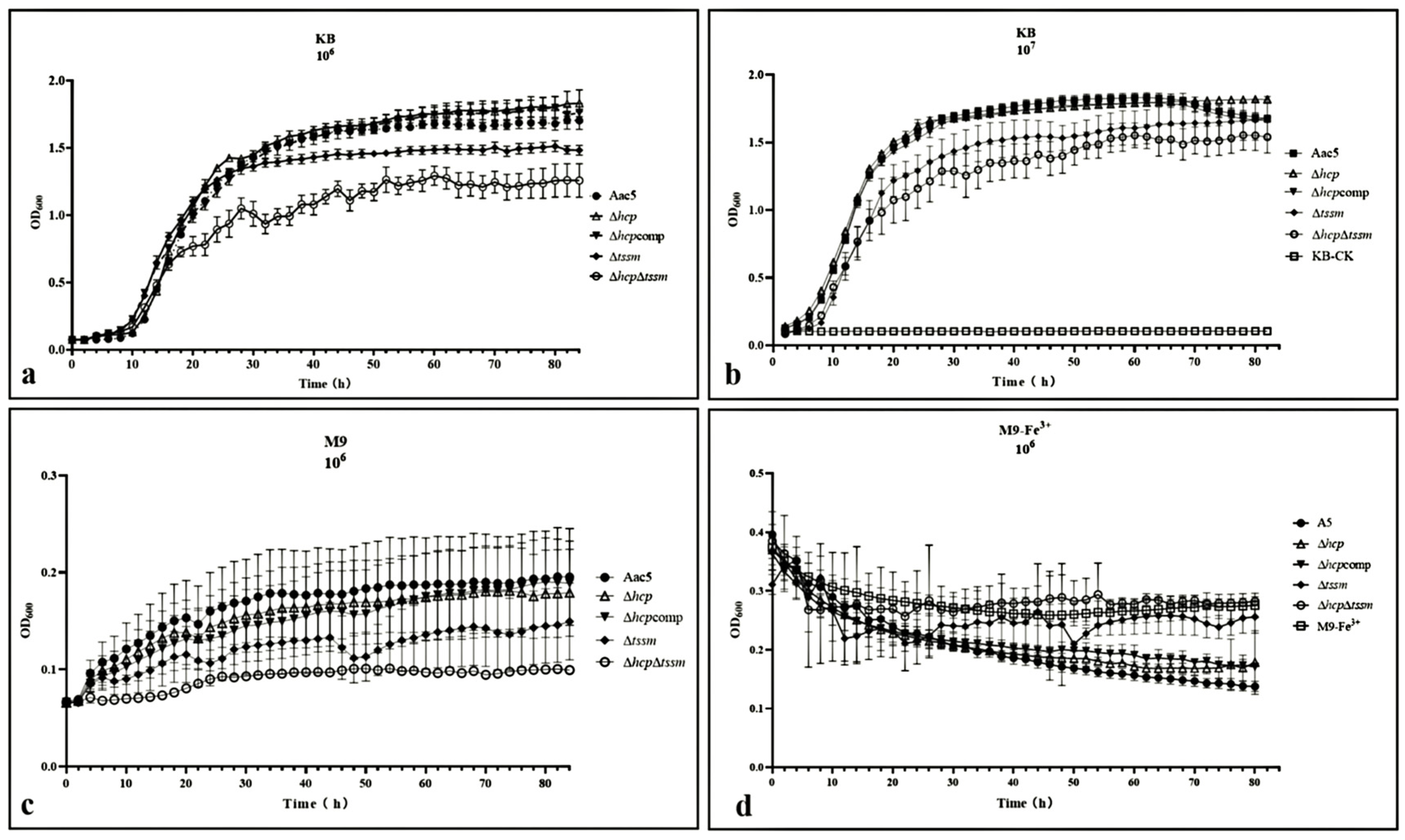
2.3. T6SS Affects the In-Vitro Growth of A. citrulli and Its Ability to Absorb Ferric Iron
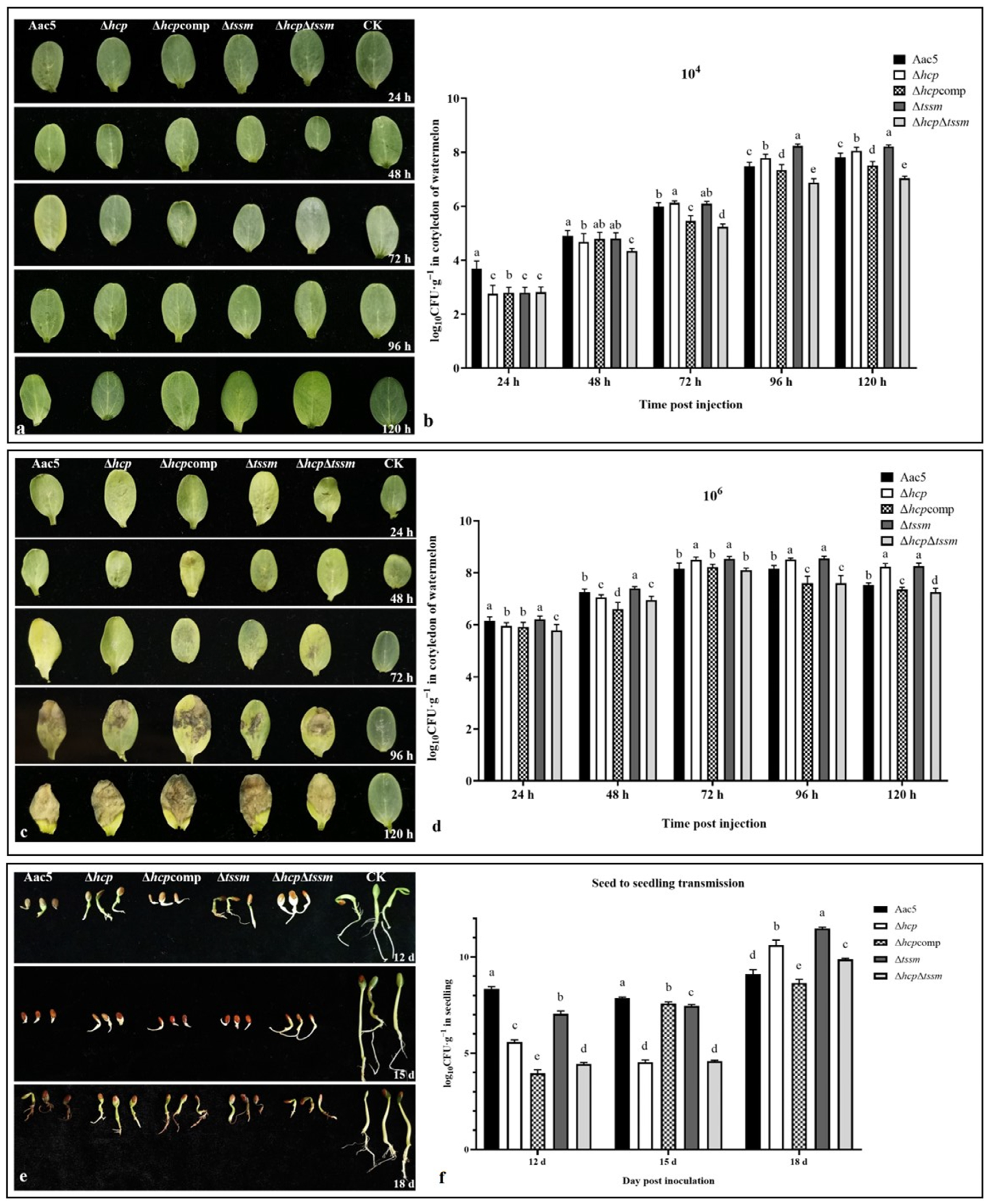
2.4. Δhcp, Δtssm, and ΔhcpΔtssm Strains Showed Differences in Colonization Ability but Not in Inducing Symptoms
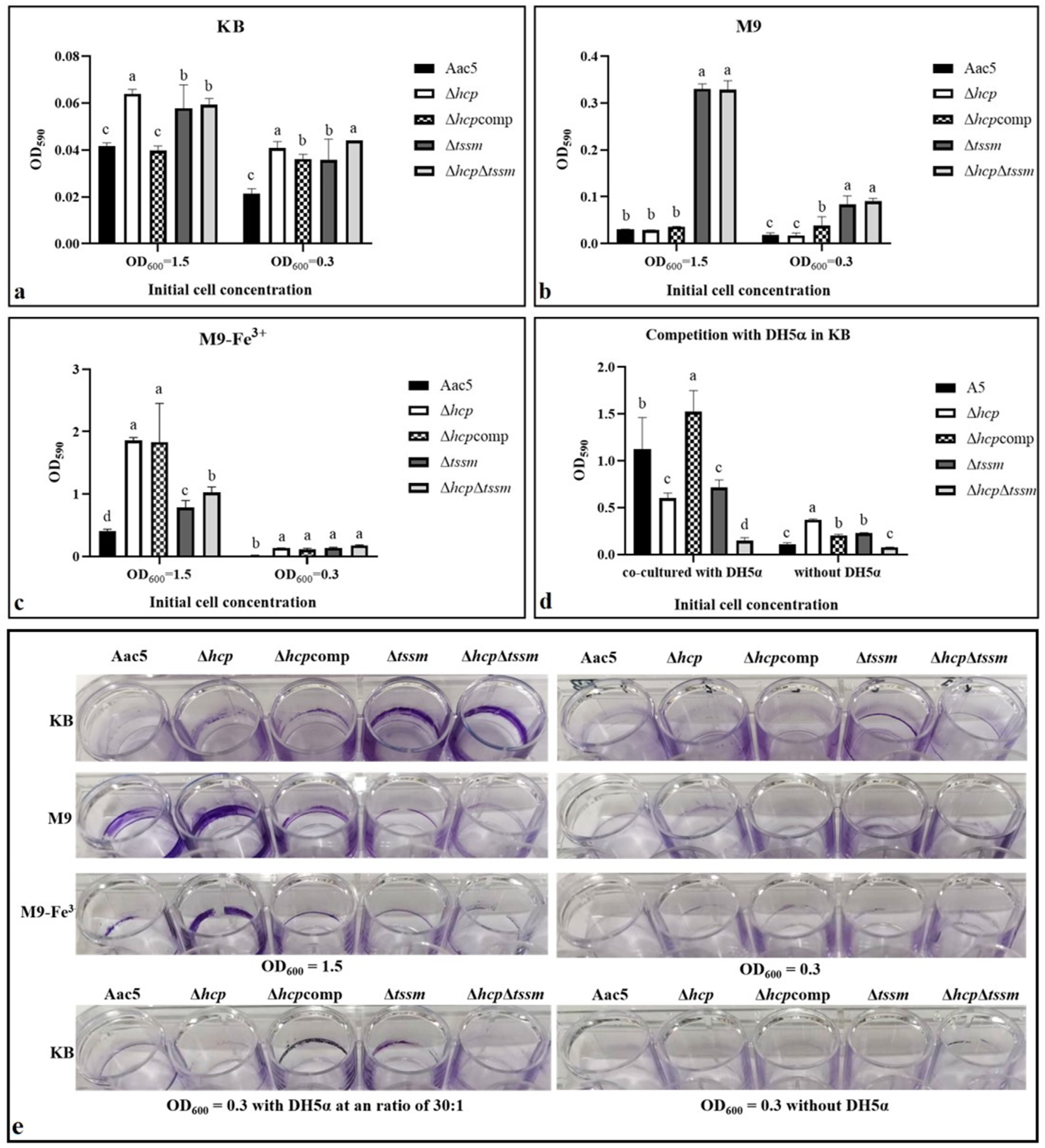
2.5. T6SS Affected the Biofilm Formation Ability
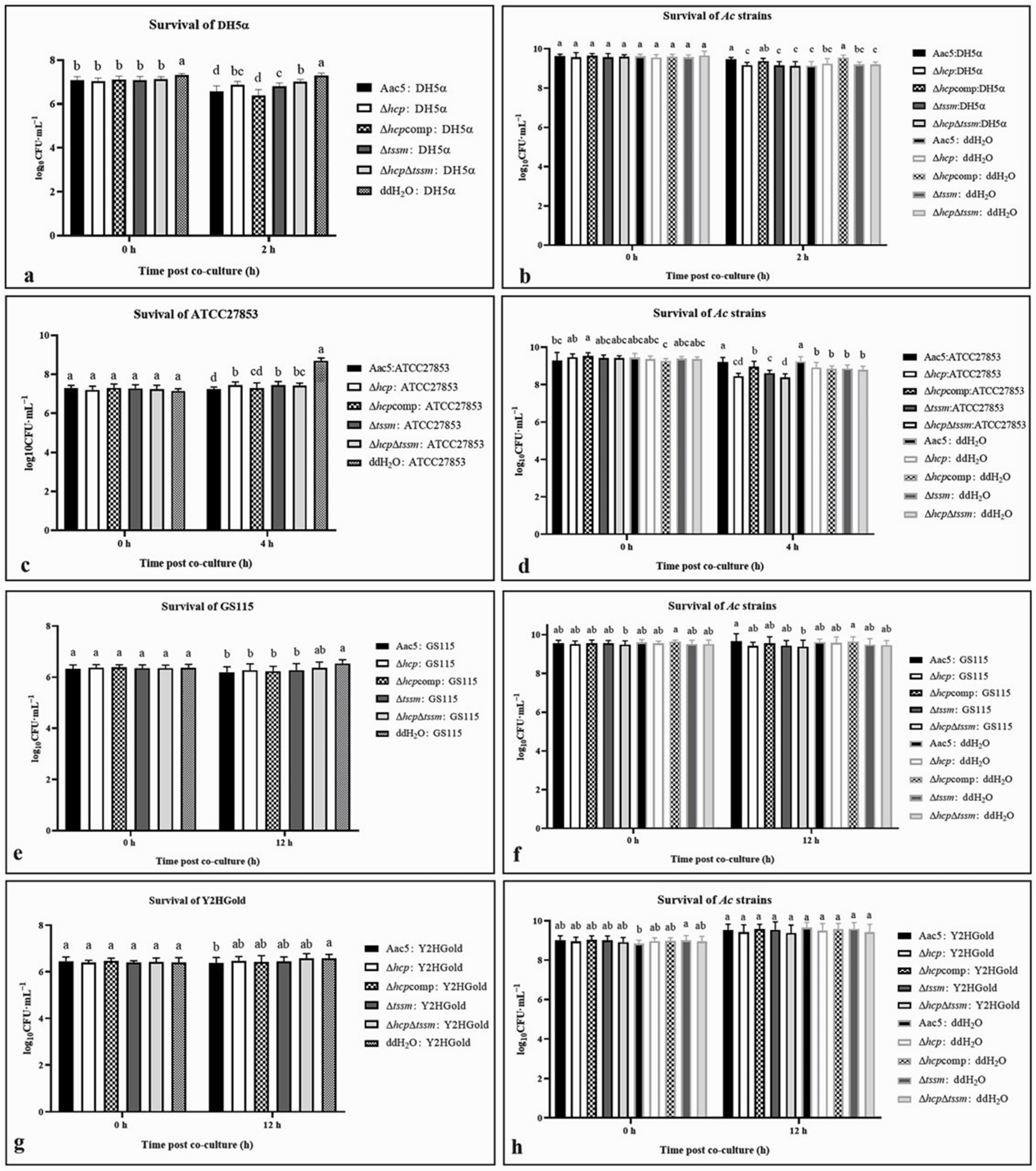
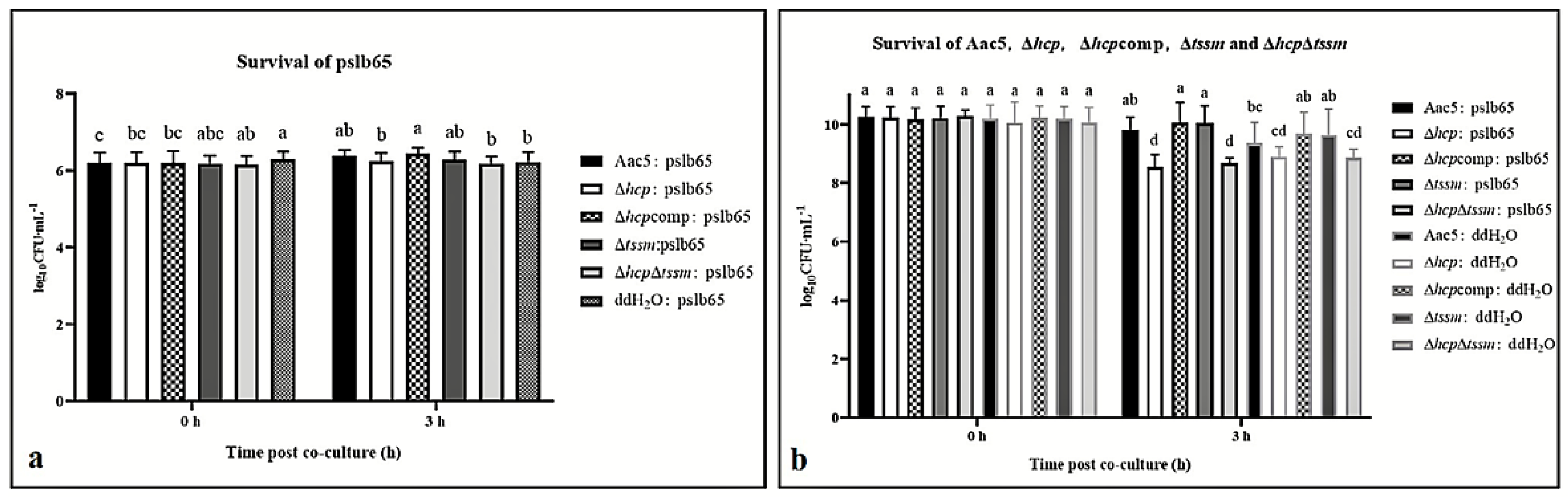
2.6. Acidovorax citrulli T6SS Is Involved in Interspecies and Intraspecies Competition
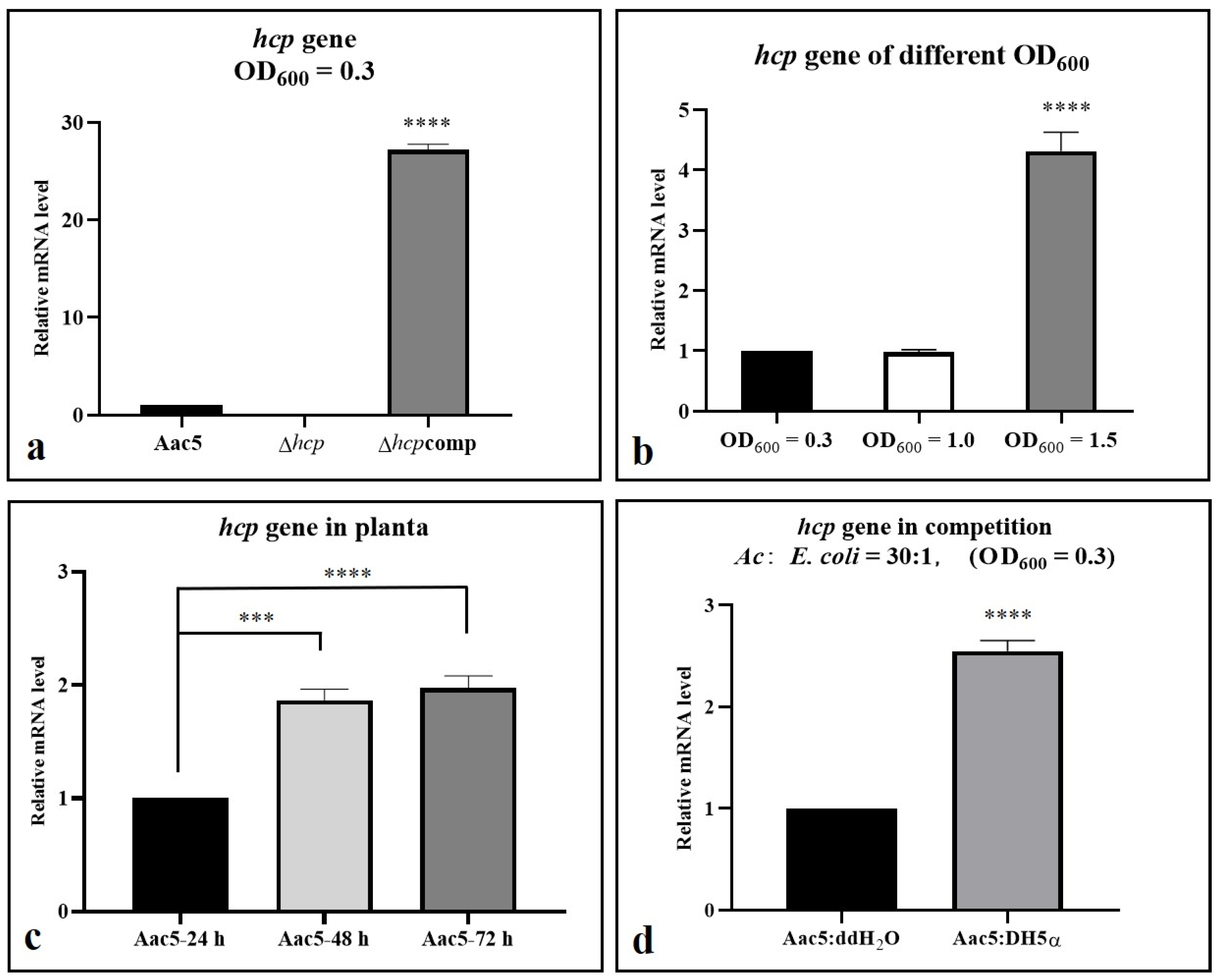
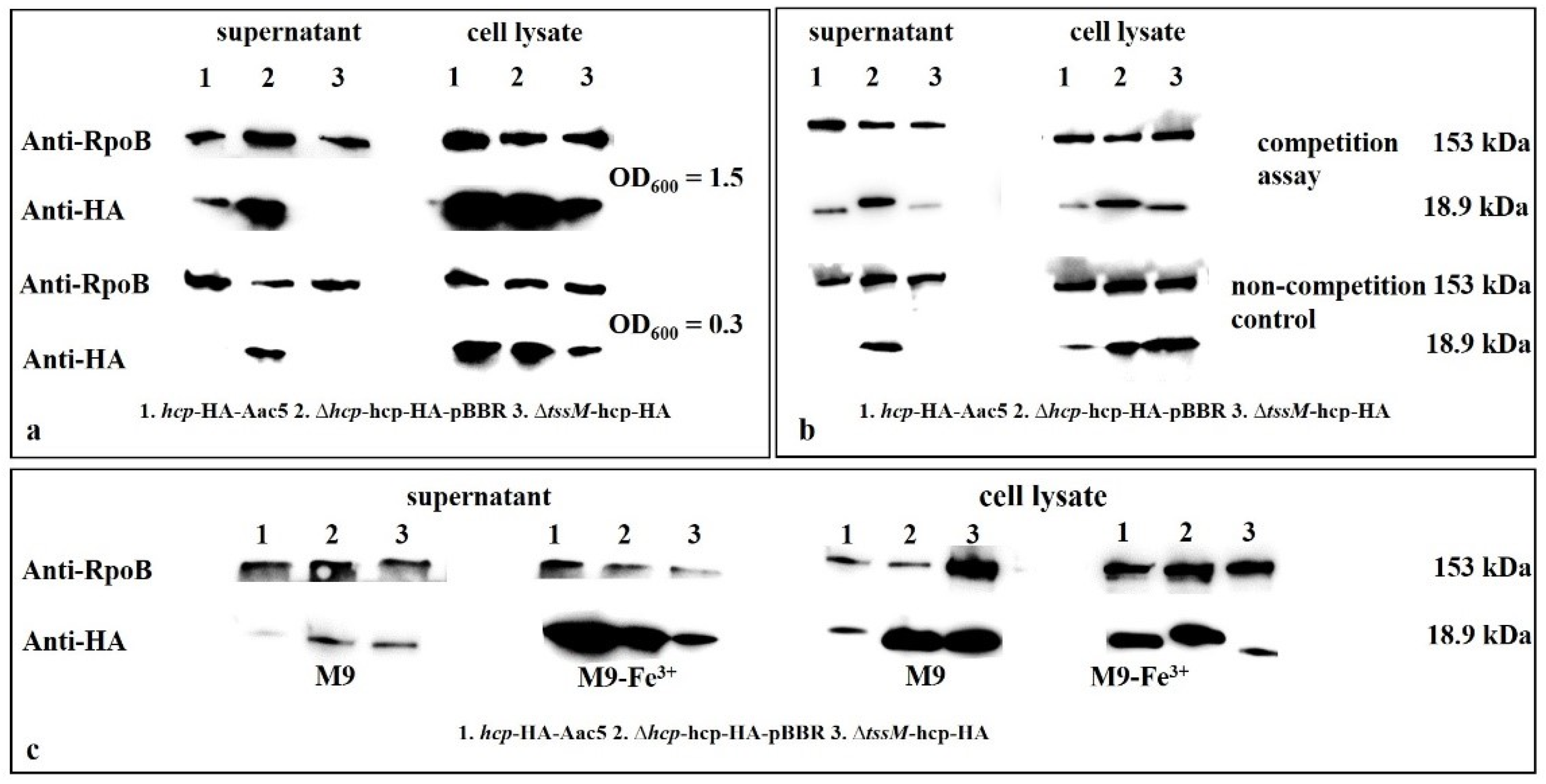
2.7. Hcp Can Be Activated by Cell Density, Ferric Iron, Competition, and the Host
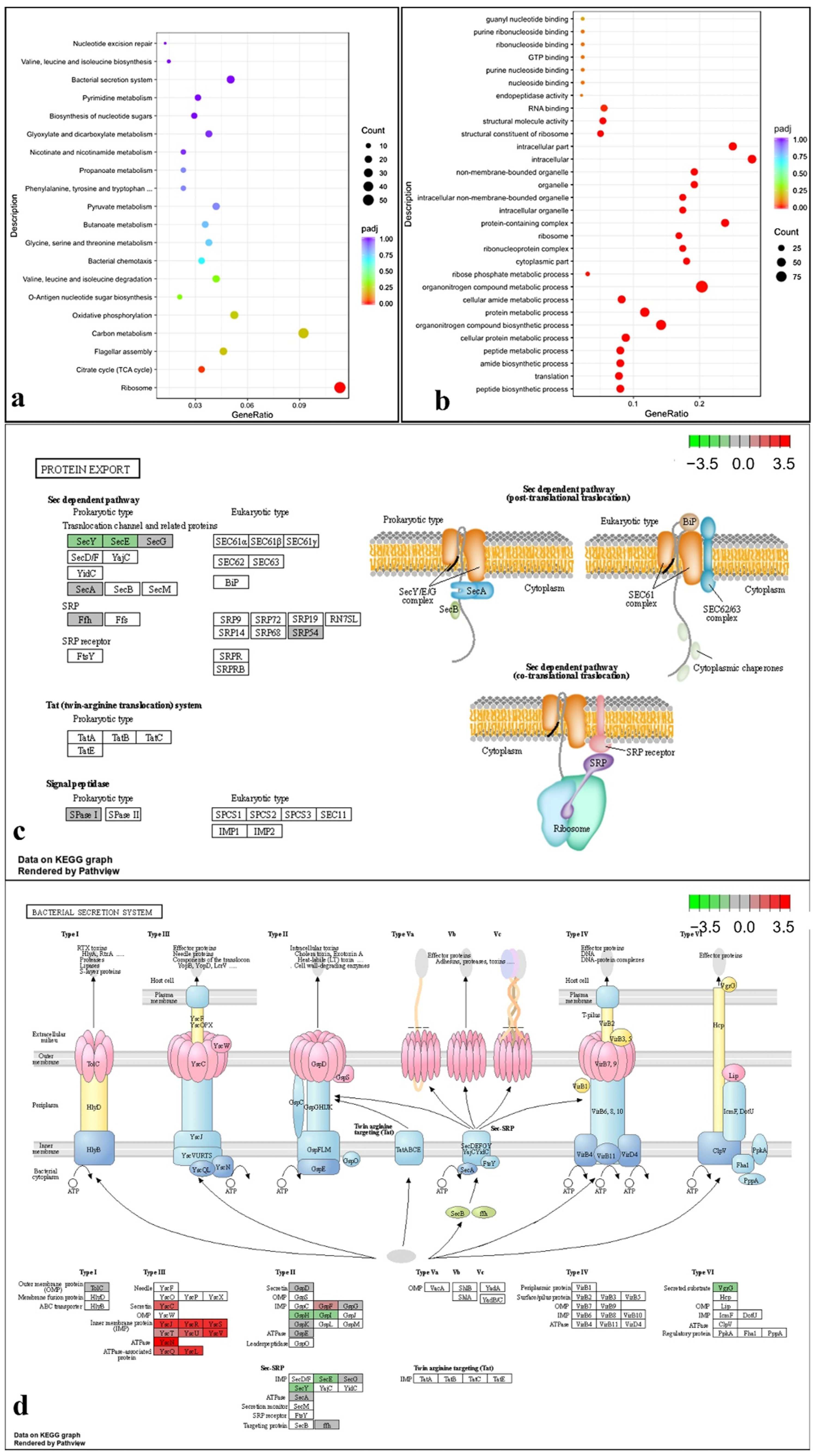
2.8. Transcriptome Analysis under T6SS Activating Conditions Indicated That the T6SS Is Related to Multiple Pathways
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Plasmids, Media, and Culture Conditions
4.2. Bioinformatics Analysis
4.3. Construction of hcp and tssm Marker-Less Deletion Mutants, hcp Complementation
4.4. Virulence and Seed-to-Seedling Transmission
4.5. Biofilm Formation Assay
4.6. Growth Ability Assay
4.7. Interspecies and Intraspecies Competition Assays
4.8. Quantitative Reverse-Transcription (qRT-PCR)
4.9. Western Blotting
4.10. Transcriptome Sequencing and Analysis under T6SS Activation Conditions
4.11. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Webb, R.E.; Goth, R.W. A seedborne bacterium isolated from watermelon. Plant Dis. Rep. 1965, 49, 818–821. [Google Scholar]
- Tian, Y.; Zhao, Y.; Zhou, J.; Sun, T.; Luo, X.; Kurowski, C.; Gong, W.; Hu, B.; Walcott, R.R. Prevalence of Acidovorax citrulli in Commercial Cucurbit Seedlots during 2010–2018 in China. Plant Dis. 2020, 104, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Schaad, N.W.; Sowell, G.; Goth, R.W.; Colwell, R.R.; Webb, R.W. Pseudomonas pseudoalcaligenes subsp. citrulli subsp. nov. Int. J. Syst. Evol. Microbiol. 1978, 28, 117–125. [Google Scholar] [CrossRef]
- Schaad, N.W.; Postnikova, E.; Sechler, A.; Claflin, L.E.; Vidaver, A.K.; Jones, J.B.; Agarkova, I.; Ignatov, A.; Dickstein, E.; Ramundo, B.A. Reclassification of subspecies of Acidovorax avenae as A. Avenae (Manns 1905) emend., A. cattleyae (Pavarino, 1911) comb. nov., A. citrulli Schaad et al., 1978) comb. nov., and proposal of A. oryzae sp. nov. Syst. Appl. Microbiol. 2008, 31, 434–446. [Google Scholar] [CrossRef]
- Walcott, R.R.; Fessehaie, A.; Castro, A.C. Differences in Pathogenicity between two Genetically Distinct Groups of Acidovorax avenae subsp. citrulli on Cucurbit Hosts. Cucurbit Hosts. J. Phytopathol. 2004, 152, 277–285. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, M.; Zhang, L.; Qiao, P.; Bai, X.; Zhang, X.; Walcott, R.; Guan, W.; Zhao, T. Development of a multiplex PCR assay based on the pilA gene sequences to detect different types of Acidovorax citrulli. J. Microbiol. Methods 2019, 158, 93–98. [Google Scholar] [CrossRef]
- Yan, S.; Yang, Y.; Wang, T.; Zhao, T.; Schaad, N.W. Genetic diversity analysis of Acidovorax citrulli in China. Eur. J. Plant Pathol. 2012, 136, 171–181. [Google Scholar] [CrossRef]
- Eckshtain-Levi, N.; Shkedy, D.; Gershovits, M.; Da Silva, G.M.; Tamir-Ariel, D.; Walcott, R.; Pupko, T.; Burdman, S. Insights from the Genome Sequence of Acidovorax citrulli M6, a Group I Strain of the Causal Agent of Bacterial Fruit Blotch of Cucurbits. Front. Microbiol. 2016, 7, 430. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Y.; Zhao, M.; Yang, L.; Jiang, J.; Walcott, R.; Yang, S.; Zhao, T. Acidovorax citrulli Type III Effector AopP Suppresses Plant Immunity by Targeting the Watermelon Transcription Factor WRKY6. Front. Plant Sci. 2020, 11, 579218. [Google Scholar] [CrossRef]
- Bahar, O.; Goffer, T.; Burdman, S. Type IV Pili Are Required for Virulence, Twitching Motility, and Biofilm Formation of Acidovorax avenae subsp. citrulli. Mol. Plant-Microbe Interact. 2009, 22, 909–920. [Google Scholar] [CrossRef]
- Wang, T.; Guan, W.; Huang, Q.; Yang, Y.; Yan, W.; Sun, B.; Zhao, T. Quorum-sensing contributes to virulence, twitching motility, seed attachment and biofilm formation in the wild type strain Aac-5 of Acidovorax citrulli. Microb. Pathog. 2016, 100, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shavit, R.; Lebendiker, M.; Pasternak, Z.; Burdman, S.; Helman, Y. The vapB–vapC Operon of Acidovorax citrulli Functions as a Bona-fide Toxin–Antitoxin Module. Front. Microbiol. 2016, 6, 1499. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Wang, T.; Huang, Q.; Zhao, M.; Tian, E.; Liu, Y.; Liu, B.; Yang, Y.; Zhao, T. Transcriptomic and Functional Analyses Reveal Roles of AclR, a luxR-type Global Regular, in Regulating Motility and Virulence of Acidovorax citrulli. Mol. Plant-Microbe Interact. 2021, 34, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Tian, Y.; Zhao, Y.; Zeng, R.; Chen, B.; Hu, B.; Walcott, R.R. Ferric Uptake Regulator (FurA) is Required for Acidovorax citrulli Virulence on Watermelon. Phytopathology 2019, 109, 1997–2008. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lee, J.; Heo, L.; Lee, S.J.; Han, S.-W. Proteomic and Phenotypic Analyses of a Putative Glycerol-3-Phosphate Dehydrogenase Required for Virulence in Acidovorax citrulli. Plant Pathol. J. 2021, 37, 36–46. [Google Scholar] [CrossRef]
- Pukatzki, S.; Ma, A.T.; Sturtevant, D.; Krastins, B.; Sarracino, D.; Nelson, W.C.; Heidelberg, J.F.; Mekalanos, J.J. Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl. Acad. Sci. USA 2006, 103, 1528–1533. [Google Scholar] [CrossRef]
- Mougous, J.D.; Cuff, M.E.; Raunser, S.; Shen, A.; Zhou, M.; Gifford, C.A.; Goodman, A.L.; Joachimiak, G.; Ordoñez, C.L.; Lory, S.; et al. A Virulence Locus of Pseudomonas aeruginosa Encodes a Protein Secretion Apparatus. Science 2006, 312, 1526–1530. [Google Scholar] [CrossRef]
- Chen, L.; Zou, Y.; She, P.; Wu, Y. Composition, function, and regulation of T6SS in Pseudomonas aeruginosa. Microbiol. Res. 2015, 172, 19–25. [Google Scholar] [CrossRef]
- Sana, T.G.; Hachani, A.; Bucior, I.; Soscia, C.; Garvis, S.; Termine, E.; Engel, J.; Filloux, A.; Bleves, S. The Second Type VI Secretion System of Pseudomonas aeruginosa Strain PAO1 Is Regulated by Quorum Sensing and Fur and Modulates Internalization in Epithelial Cells. J. Biol. Chem. 2012, 287, 27095–27105. [Google Scholar] [CrossRef]
- Tian, Y.; Zhao, Y.; Wu, X.; Liu, F.; Hu, B.; Walcott, R.R. The type VI protein secretion system contributes to biofilm formation and seed-to-seedling transmission of Acidovorax citrulli on melon. Mol. Plant Pathol. 2014, 16, 38–47. [Google Scholar] [CrossRef]
- Bernal, P.; Llamas, M.A.; Filloux, A. Type VI secretion systems in plant-associated bacteria. Environ. Microbiol. 2018, 20, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, S.; West, T.E.; Boyer, F.; Chiang, W.-C.; Carl, M.A.; Hood, R.D.; Rohmer, L.; Tolker-Nielsen, T.; Skerrett, S.J.; Mougous, J.D. Burkholderia Type VI Secretion Systems Have Distinct Roles in Eukaryotic and Bacterial Cell Interactions. PLOS Pathog. 2010, 6, e1001068. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Salas Gonzalez, I.; Mittelviefhaus, M.; Clingenpeel, S.; Harrera Paredes, S.; Miao, J.; Wang, K.; Devescovi, G.; Stillman, K.; Monteiro, F.; et al. Genomic features of bacterial adaptation to plants. Nat. Genet. 2018, 50, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Pei, T.-T.; Kan, Y.; Wang, Z.-H.; Tang, M.-X.; Li, H.; Yan, S.; Cui, Y.; Zheng, H.-Y.; Luo, H.; Liang, X.; et al. Delivery of an Rhs-family nuclease effector reveals direct penetration of the gram-positive cell envelope by a type VI secretion system in Acidovorax citrulli. mLife 2022, 1, 66–78. [Google Scholar] [CrossRef]
- Silverman, J.M.; Agnello, D.M.; Zheng, H.; Andrews, B.T.; Li, M.; Catalano, C.E.; Gonen, T.; Mougous, J.D. Haemolysin Coregulated Protein Is an Exported Receptor and Chaperone of Type VI Secretion Substrates. Mol. Cell 2013, 51, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Lien, Y.-W.; Lai, E.-M. Type VI Secretion Effectors: Methodologies and Biology. Front. Cell. Infect. Microbiol. 2017, 7, 254. [Google Scholar] [CrossRef]
- Cianfanelli, F.R.; Monlezun, L.; Coulthurst, S.J. Aim, Load, Fire: The Type VI Secretion System, a Bacterial Nanoweapon. Trends Microbiol. 2015, 24, 51–62. [Google Scholar] [CrossRef]
- Wu, C.-F.; Lien, Y.-W.; Bondage, D.; Lin, J.-S.; Pilhofer, M.; Shih, Y.-L.; Chang, J.H.; Lai, E.-M. Effector loading onto the VgrG carrier activates type VI secretion system assembly. EMBO Rep. 2019, 21, e47961. [Google Scholar] [CrossRef]
- Ma, L.-S.; Hachani, A.; Lin, J.-S.; Filloux, A.; Lai, E.-M. Agrobacterium tumefaciens Deploys a Superfamily of Type VI Secretion DNase Effectors as Weapons for Interbacterial Competition in Planta. Cell Host Microbe 2014, 16, 94–104. [Google Scholar] [CrossRef]
- Flaugnatti, N.; Le, T.T.H.; Canaan, S.; Aschtgen, M.-S.; Nguyen, V.S.; Blangy, S.; Kellenberger, C.; Roussel, A.; Cambillau, C.; Cascales, E.; et al. A phospholipase A1 antibacterial Type VI secretion effector interacts directly with the C-terminal domain of the VgrG spike protein for delivery. Mol. Microbiol. 2016, 99, 1099–1118. [Google Scholar] [CrossRef]
- Cianfanelli, F.R.; Alcoforado Diniz, J.; Guo, M.; De Cesare, V.; Trost, M.; Coulthurst, S.J. VgrG and PAAR Proteins Define Distinct Versions of a Functional Type VI Secretion System. PLOS Pathog. 2016, 12, e1005735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- English, G.; Trunk, K.; Rao, V.A.; Srikannathasan, V.; Hunter, W.N.; Coulthurst, S.J. New secreted toxins and immunity proteins encoded within the T ype VI secretion system gene cluster of Serratia marcescens. Mol. Microbiol. 2012, 86, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Hood, R.D.; Singh, P.; Hsu, F.; Güvener, T.; Carl, M.A.; Trinidad, R.R.S.; Silverman, J.M.; Ohlson, B.B.; Hicks, K.G.; Plemel, R.L.; et al. A Type VI Secretion System of Pseudomonas aeruginosa Targets a Toxin to Bacteria. Cell Host Microbe 2010, 7, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Ringel, P.D.; Hu, D.; Basler, M. The Role of Type VI Secretion System Effectors in Target Cell Lysis and Subsequent Horizontal Gene Transfer. Cell Rep. 2017, 21, 3927–3940. [Google Scholar] [CrossRef]
- Pazhani, G.P.; Chowdhury, G.; Ramamurthy, T. Adaptations of Vibrio parahaemolyticus to Stress During Environmental Survival, Host Colonization, and Infection. Front. Microbiol. 2021, 12, 737299. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Thompson, C.M. Evaluation of Citrullus sp. Germ Plasm for Resistance to Acidovorax avenae subsp. citrulli. Plant Dis. 2002, 86, 61–64. [Google Scholar] [CrossRef]
- Bahar, O.; Kritzman, G.; Burdman, S. Bacterial fruit blotch of melon: Screens for disease tolerance and role of seed transmission in pathogenicity. Eur. J. Plant Pathol. 2008, 123, 71–83. [Google Scholar] [CrossRef]
- Bendtsen, J.D.; Kiemer, L.; Fausbøll, A.; Brunak, S. Non-classical protein secretion in bacteria. BMC Microbiol. 2005, 5, 58. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Li, J.; Yao, Y.; Xu, H.H.; Hao, L.; Deng, Z.; Rajakumar, K.; Ou, H.-Y. SecReT6: A web-based resource for type VI secretion systems found in bacteria. Environ. Microbiol. 2015, 17, 2196–2202. [Google Scholar] [CrossRef]
- Journet, L.; Cascales, E. The Type VI Secretion System in Escherichia coli and Related Species. EcoSal Plus 2016, 7, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, A.; Kostiuk, B.; Rogers, A.; Teschler, J.; Pukatzki, S.; Yildiz, F.H. Rules of Engagement: The Type VI Secretion System in Vibrio cholerae. Trends Microbiol. 2016, 25, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Burtnick, M.N.; Brett, P.J. Burkholderia mallei and Burkholderia pseudomallei Cluster 1 Type VI Secretion System Gene Expression Is Negatively Regulated by Iron and Zinc. PLoS ONE 2013, 8, e76767. [Google Scholar] [CrossRef]
- Haapalainen, M.; Mosorin, H.; Dorati, F.; Wu, R.-F.; Roine, E.; Taira, S.; Nissinen, R.; Mattinen, L.; Jackson, R.; Pirhonen, M.; et al. Hcp2, a Secreted Protein of the Phytopathogen Pseudomonas syringae pv. Tomato DC3000, Is Required for Fitness for Competition against Bacteria and Yeasts. J. Bacteriol. 2012, 194, 4810–4822. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Zhang, X.; Zhao, M.; Yan, J.; Yang, L.; Yang, Y.; Guan, W.; Walcott, R.; Zhao, T. Involvement of hrpX and hrpG in the Virulence of Acidovorax citrulli Strain Aac5, Causal Agent of Bacterial Fruit Blotch in Cucurbits. Front. Microbiol. 2018, 9, 507. [Google Scholar] [CrossRef]
- Guan, W.; Wang, T.; Huang, Q.; Tian, E.; Liu, B.; Yang, Y.; Zhao, T. A LuxR-type regulator, AcrR, regulates flagellar assembly and contributes to virulence, motility, biofilm formation, and growth ability of Acidovorax citrulli. Mol. Plant Pathol. 2020, 21, 489–501. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential Expression of RNA-Seq Data at the Gene Level—The DESeq Package. 2012. Available online: http://www.bioconductor.org/packages/2.11/bioc/vignettes/DESeq/inst/doc/DESeq.pdf (accessed on 9 June 2022).
- Pan, Y.; Liang, F.; Li, R.-J.; Qian, W. MarR-Family Transcription Factor HpaR Controls Expression of the vgrR-vgrS Operon of Xanthomonas campestris pv. campestris. Mol. Plant-Microbe Interact. 2018, 31, 299–310. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, N.; Ji, W.; Yang, L.; Yu, C.; Qiao, P.; Yan, J.; Guan, W.; Yang, Y.; Zhao, T. Hcp of the Type VI Secretion System (T6SS) in Acidovorax citrulli Group II Strain Aac5 Has a Dual Role as a Core Structural Protein and an Effector Protein in Colonization, Growth Ability, Competition, Biofilm Formation, and Ferric Iron Absorption. Int. J. Mol. Sci. 2022, 23, 9632. https://doi.org/10.3390/ijms23179632
Fei N, Ji W, Yang L, Yu C, Qiao P, Yan J, Guan W, Yang Y, Zhao T. Hcp of the Type VI Secretion System (T6SS) in Acidovorax citrulli Group II Strain Aac5 Has a Dual Role as a Core Structural Protein and an Effector Protein in Colonization, Growth Ability, Competition, Biofilm Formation, and Ferric Iron Absorption. International Journal of Molecular Sciences. 2022; 23(17):9632. https://doi.org/10.3390/ijms23179632
Chicago/Turabian StyleFei, Nuoya, Weiqin Ji, Linlin Yang, Chunyan Yu, Pei Qiao, Jianpei Yan, Wei Guan, Yuwen Yang, and Tingchang Zhao. 2022. "Hcp of the Type VI Secretion System (T6SS) in Acidovorax citrulli Group II Strain Aac5 Has a Dual Role as a Core Structural Protein and an Effector Protein in Colonization, Growth Ability, Competition, Biofilm Formation, and Ferric Iron Absorption" International Journal of Molecular Sciences 23, no. 17: 9632. https://doi.org/10.3390/ijms23179632