

Lactobacillusjohnsonii L531 Protects against Salmonella Infantis-Induced Intestinal Damage by Regulating the NOD Activation, Endoplasmic Reticulum Stress, and Autophagy
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. L. johnsonii L531 Is Protective in the Prevention of S. Infantis-Induced Intestinal Inflammation and Damage to the Small Intestine
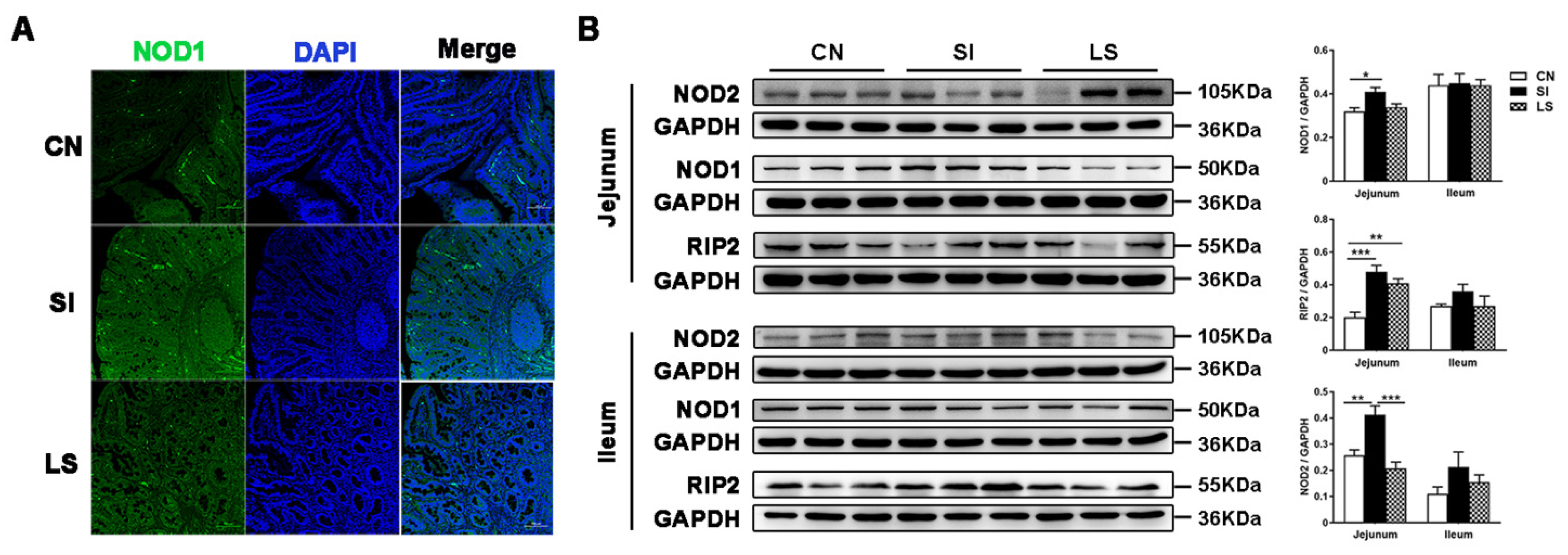
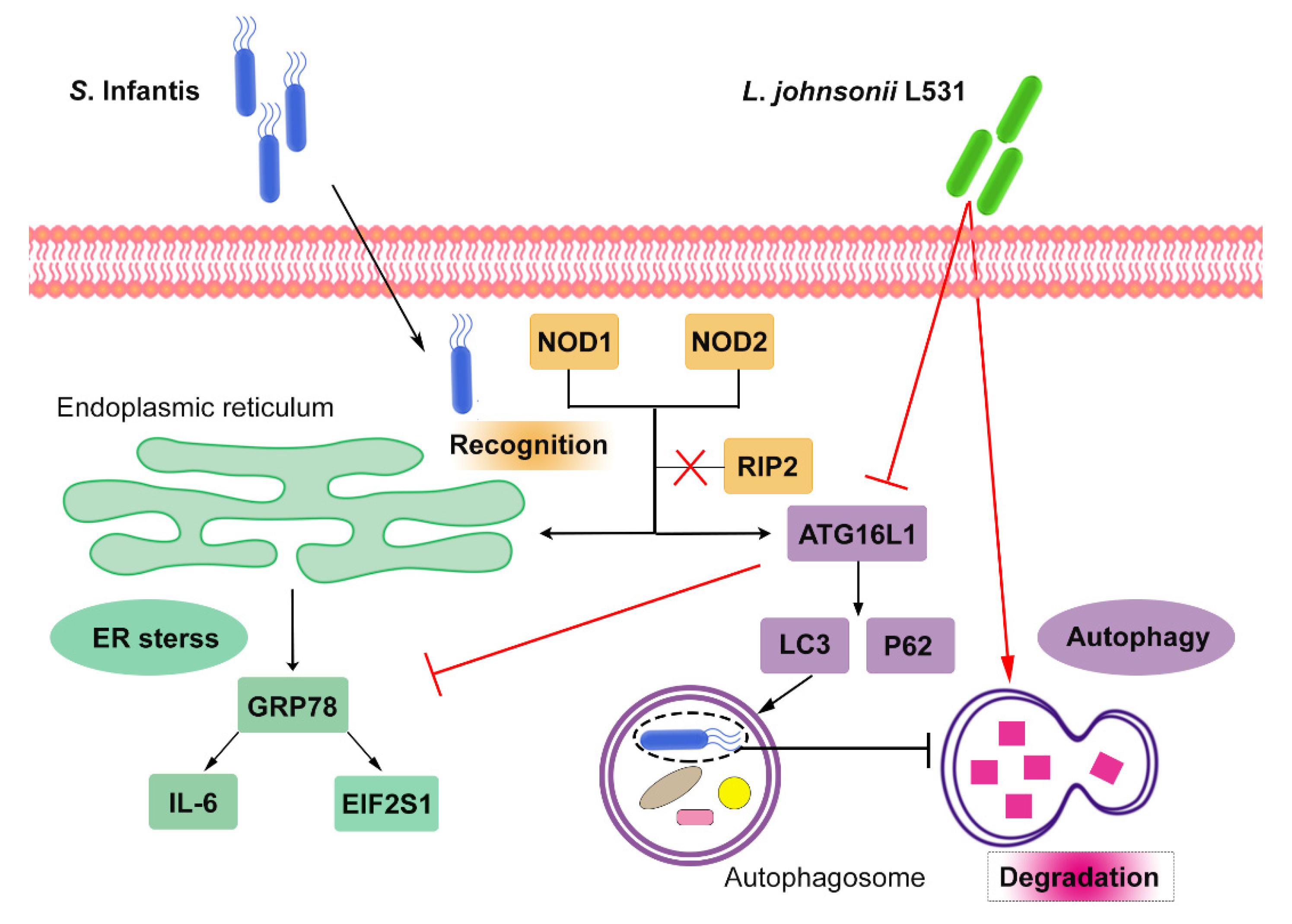
2.2. Pretreatment with L. johnsonii L531 Suppresses S. Infantis-Induced Activation of the NOD Pathway in the Small Intestine
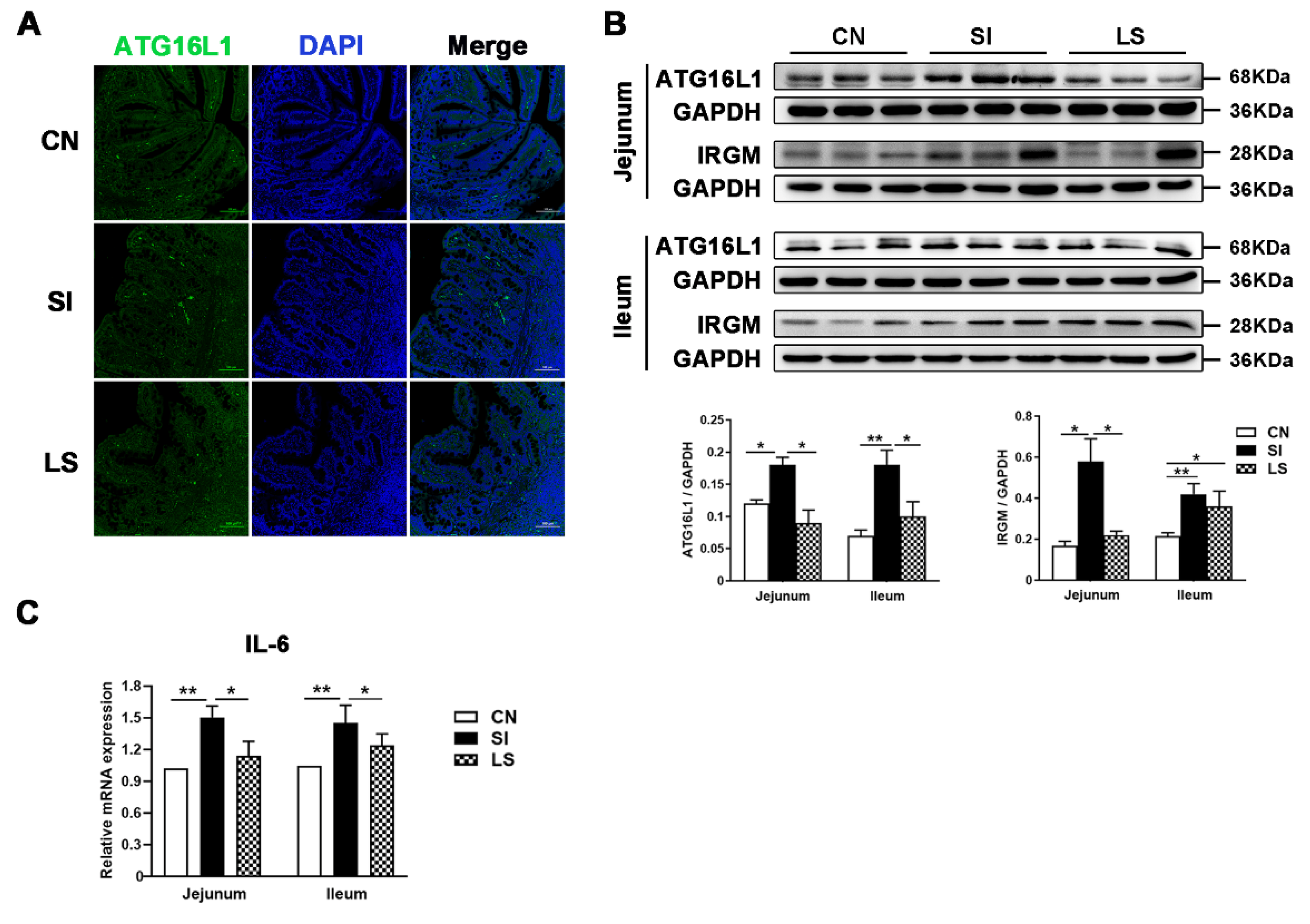
2.3. Pretreatment with L. johnsonii L531 Attenuates S. Infantis-Induced Autophagy in the Small Intestine
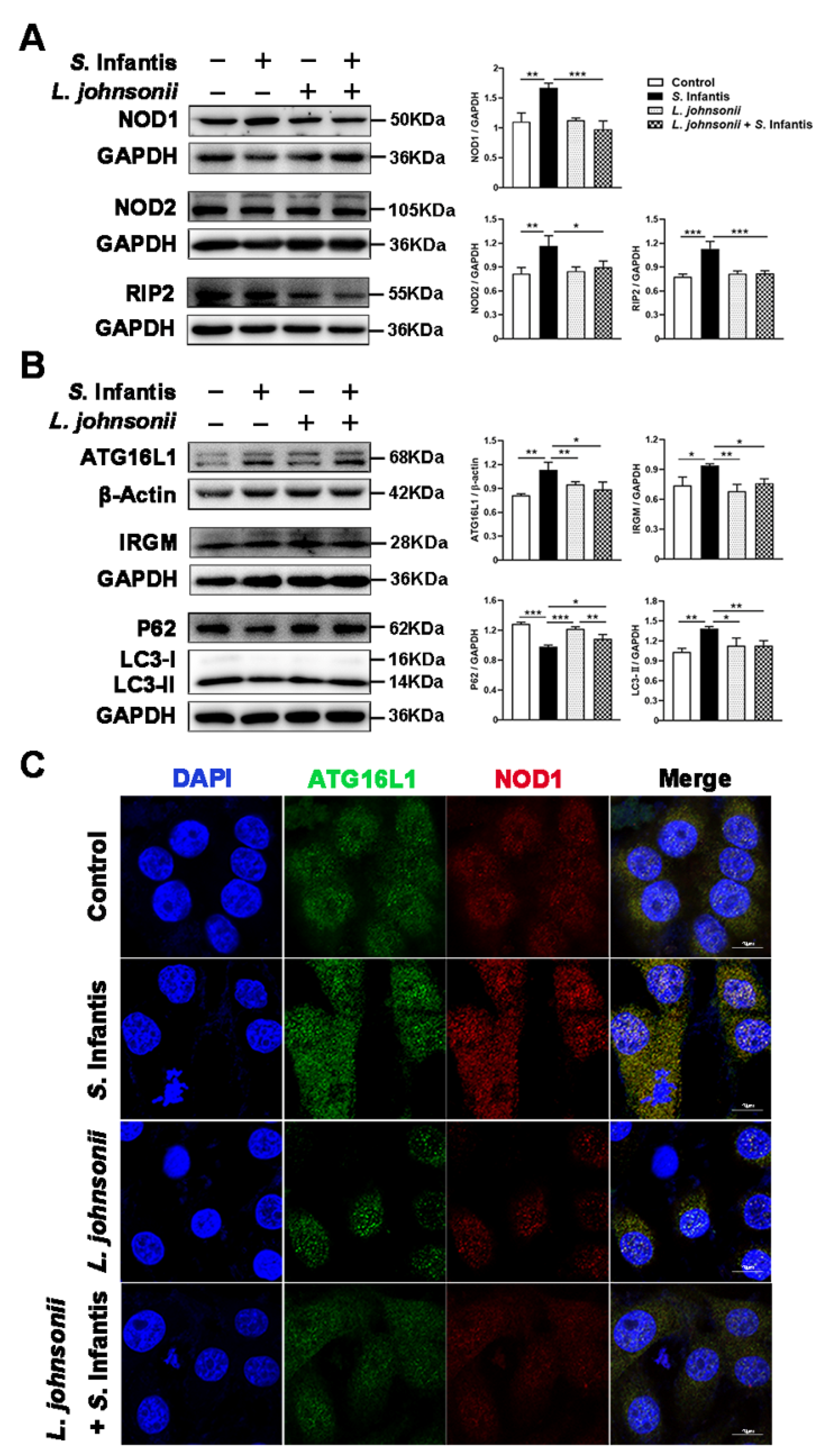
2.4. L. johnsonii L531 Represses S. Infantis-Induced NOD Activation and Modulates Autophagy in Caco2 Cells
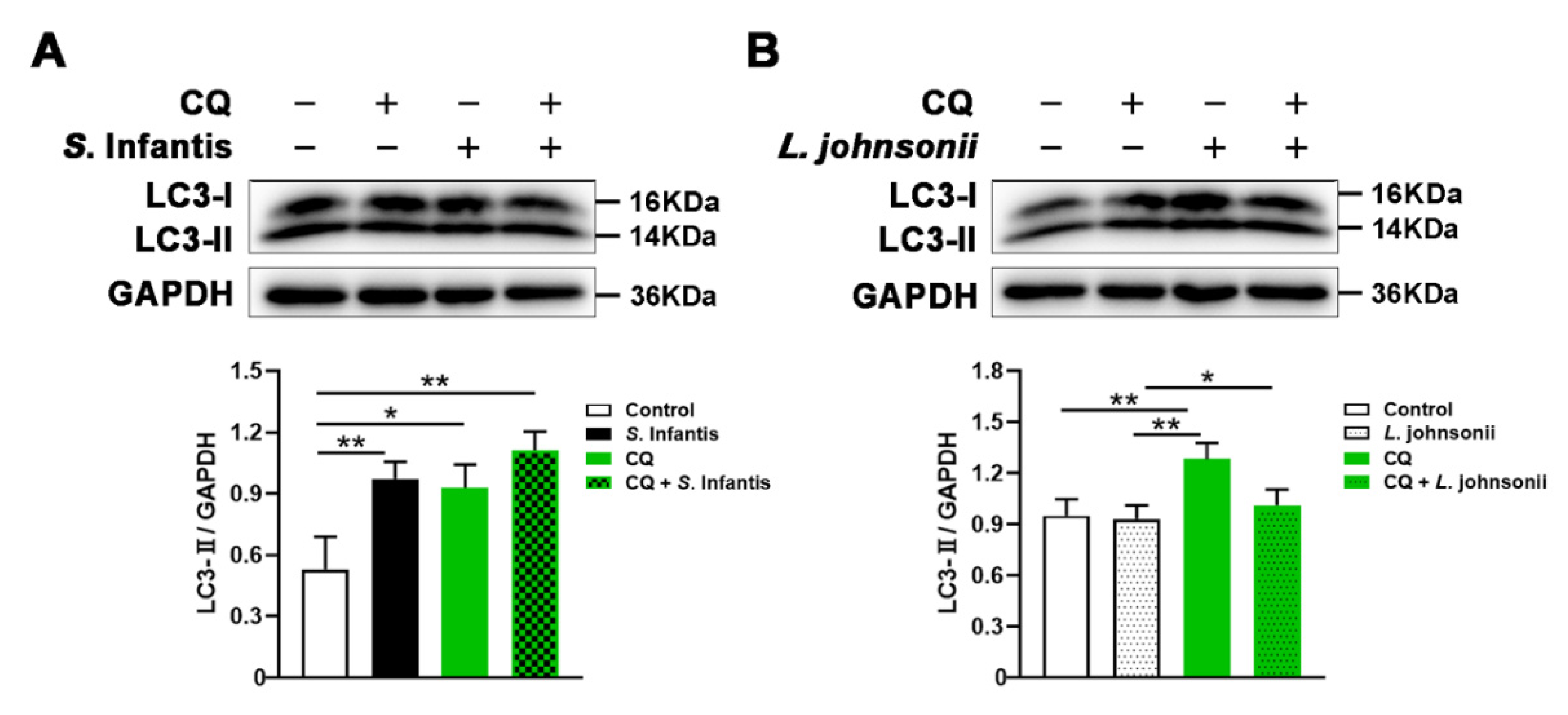
2.5. L. johnsonii L531 Enhances the Resistance of Caco2 Cells to S. Infantis by Promoting Autophagy Degradation
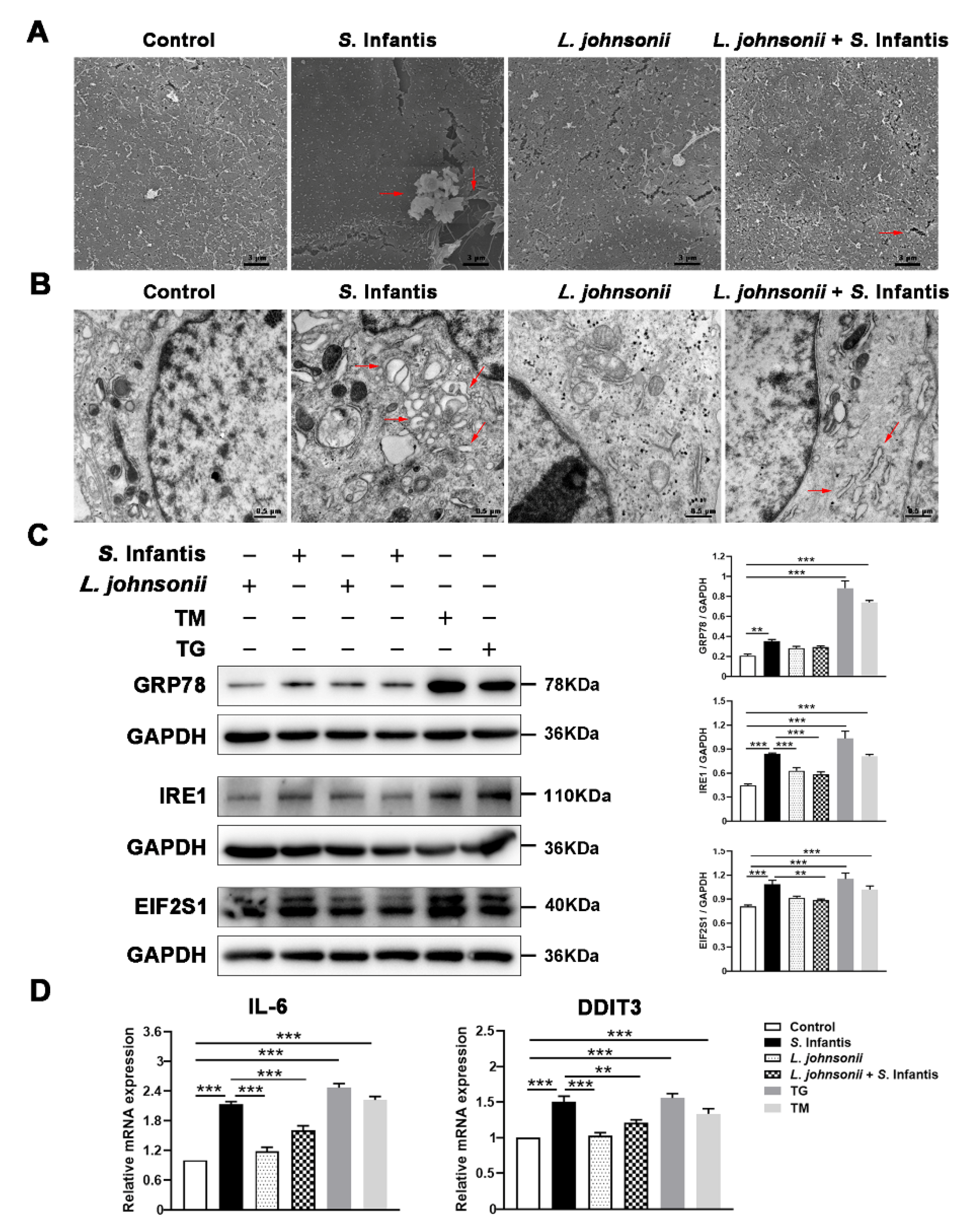
2.6. L. johnsonii L531 Alleviates Cell Damage by Regulating ER Stress in Response to S. Infantis
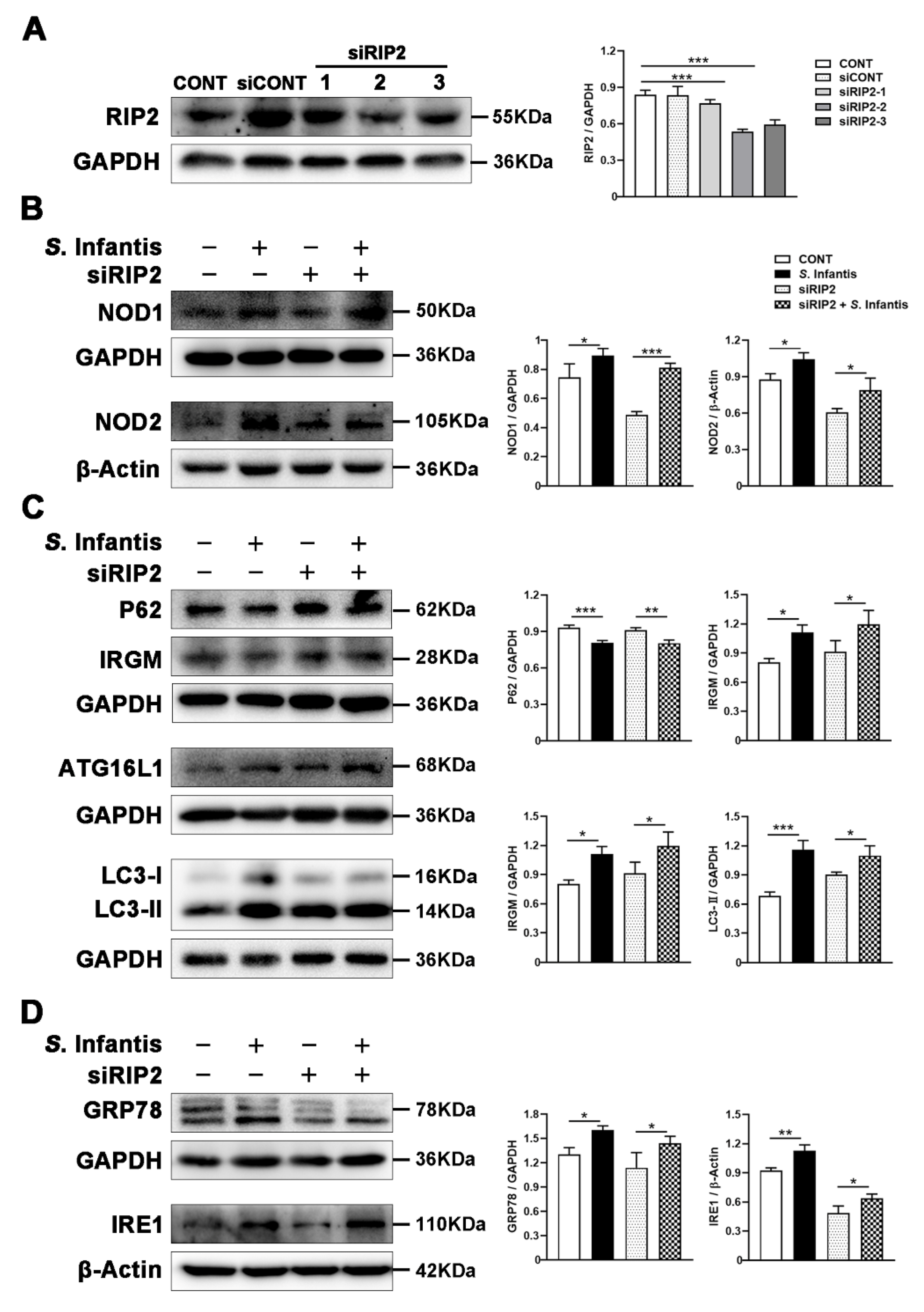
2.7. RIP2 Inhibition Did Not Alter S. Infantis-Induced Autophagy and ER Stress in Caco2 Cells
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Bacterial Strains and Preparation
4.3. Animals and Specimens
4.4. Regents and Antibodies
4.5. In Vitro Caco2 Cell Infection Model
4.6. RNA Interference
4.7. Western Blotting
4.8. Quantitative Real-Time PCR
4.9. Scanning Electron Microscopy and Transmission Electron Microscopy
4.10. Immunofluorescence
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, G.Y.; Yu, J.; Su, J.H.; Jiao, L.G.; Liu, X.; Zhu, Y.H. Oral Administration of Lactobacillus rhamnosus GG ameliorates Salmonella Infantis-induced inflammation in a pig model via activation of the IL-22BP/IL-22/STAT3 pathway. Front. Cell. Infect. Microbiol. 2017, 7, 323. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Bernstein, C.N.; Gearry, R.; Hviid, A.; Kolho, K.L.; Kronman, M.P.; Shaw, S.; Van Kruiningen, H.; Colombel, J.F.; Atreja, A. Antibiotics associated with increased risk of new-onset Crohn’s disease but not ulcerative colitis: A meta-analysis. Am. J. Gastroenterol. 2014, 109, 1728–1738. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Zhu, Y.H.; Yu, J.; Xia, B.; Liu, X.; Yang, G.Y.; Su, J.H.; Guo, L.; Wang, M.L.; Wang, J.F. Lactobacillus johnsonii L531 reduces pathogen load and helps maintain short-chain fatty acid levels in the intestines of pigs challenged with Salmonella enterica Infantis. Vet. Microbiol. 2019, 230, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Yu, J.; He, T.; Liu, X.; Su, J.; Wang, M.; Wang, J.; Zhu, Y. Lactobacillus johnsonii L531 ameliorates enteritis via elimination of damaged mitochondria and suppression of SQSTM1-dependent mitophagy in a Salmonella Infantis model of piglet diarrhea. FASEB J. 2020, 34, 2821–2839. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Y.; Xia, B.; Su, J.H.; He, T.; Liu, X.; Guo, L.; Zhang, S.; Zhu, Y.H.; Wang, J.F. Anti-inflammatory effects of Lactobacillus johnsonii L531 in a pig model of Salmonella Infantis infection involves modulation of CCR6+ T cell responses and ER stress. Vet. Res. 2020, 51, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Mu, T.; Yang, Y.; Zhang, J.; Ren, F.; Wu, Z. Lactobacillus johnsonii Attenuates Citrobacter rodentium-induced colitis by regulating inflammatory responses and endoplasmic reticulum stress in mice. J. Nutr. 2021, 151, 3391–3399. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sun, L.; Zha, W.; Studer, E.; Gurley, E.; Chen, L.; Wang, X.; Hylemon, P.B.; Pandak, W.M., Jr.; Sanyal, A.J.; et al. HIV protease inhibitors induce endoplasmic reticulum stress and disrupt barrier integrity in intestinal epithelial cells. Gastroenterology 2010, 138, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.P.; da Rocha, A.L.; Kohama, E.B.; Gaspar, R.C.; Simabuco, F.M.; Frantz, F.G.; de Moura, L.P.; Pauli, J.R.; Cintra, D.E.; Ropelle, E.R.; et al. Exhaustive acute exercise-induced ER stress is attenuated in IL-6-knockout mice. J. Endocrinol. 2019, 240, 181–193. [Google Scholar] [CrossRef]
- Li, Y.; Schwabe, R.F.; DeVries-Seimon, T.; Yao, P.M.; Gerbod-Giannone, M.C.; Tall, A.R.; Davis, R.J.; Flavell, R.; Brenner, D.A.; Tabas, I. Free cholesterol-loaded macrophages are an abundant source of tumor necrosis factor-alpha and interleukin-6: Model of NF-kappaB- and map kinase-dependent inflammation in advanced atherosclerosis. J. Biol. Chem. 2005, 280, 21763–21772. [Google Scholar] [CrossRef]
- Zhang, Z.; Qiao, D.; Zhang, Y.; Chen, Q.; Chen, Y.; Tang, Y.; Que, R.; Chen, Y.; Zheng, L.; Dai, Y.; et al. Portulaca oleracea L. Extract Ameliorates Intestinal Inflammation by Regulating Endoplasmic Reticulum Stress and Autophagy. Mol. Nutr. Food Res. 2022, 66, e2100791. [Google Scholar] [CrossRef]
- Tang, C.; Meng, F.; Pang, X.; Chen, M.; Zhou, L.; Lu, Z.; Lu, Y. Protective effects of Lactobacillus acidophilus NX2-6 against oleic acid-induced steatosis, mitochondrial dysfunction, endoplasmic reticulum stress and inflammatory responses. J. Funct. Foods 2020, 74, 104206. [Google Scholar] [CrossRef]
- Tang, C.; Kong, L.; Shan, M.; Lu, Z.; Lu, Y. Protective and ameliorating effects of probiotics against diet-induced obesity: A review. Food Res. Int. 2021, 147, 110490. [Google Scholar] [CrossRef]
- Keestra-Gounder, A.M.; Byndloss, M.X.; Seyffert, N.; Young, B.M.; Chavez-Arroyo, A.; Tsai, A.Y.; Cevallos, S.A.; Winter, M.G.; Pham, O.H.; Tiffany, C.R.; et al. NOD1 and NOD2 signaling links ER stress with inflammation. Nature 2016, 532, 394–397. [Google Scholar] [CrossRef]
- Irving, A.T.; Mimuro, H.; Kufer, T.A.; Lo, C.; Wheeler, R.; Turner, L.J.; Thomas, B.J.; Malosse, C.; Gantier, M.P.; Casillas, L.N.; et al. The immune receptor NOD1 and kinase RIP2 interact with bacterial peptidoglycan on early endosomes to promote autophagy and inflammatory signaling. Cell Host Microbe 2014, 15, 623–635. [Google Scholar] [CrossRef]
- Dufner, A.; Duncan, G.S.; Wakeham, A.; Elford, A.R.; Hall, H.T.; Ohashi, P.S.; Mak, T.W. CARD6 is interferon inducible but not involved in nucleotide-binding oligomerization domain protein signaling leading to NF-kappaB activation. Mol. Cell. Biol. 2008, 28, 1541–1552. [Google Scholar] [CrossRef]
- Anand, P.K.; Tait, S.W.; Lamkanfi, M.; Amer, A.O.; Nunez, G.; Pagès, G.; Pouysségur, J.; McGargill, M.A.; Green, D.R.; Kanneganti, T.D. TLR2 and RIP2 pathways mediate autophagy of Listeria monocytogenes via extracellular signal-regulated kinase (ERK) activation. J. Biol. Chem. 2011, 286, 42981–42991. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Pantoom, S.; Wu, Y.W. Elucidation of the anti-autophagy mechanism of the Legionella effector RavZ using semisynthetic LC3 proteins. eLife 2017, 6, e23905. [Google Scholar] [CrossRef]
- Liu, W.; Zhou, Y.; Peng, T.; Zhou, P.; Ding, X.; Li, Z.; Zhong, H.; Xu, Y.; Chen, S.; Hang, H.C.; et al. N(epsilon)-fatty acylation of multiple membrane-associated proteins by Shigella IcsB effector to modulate host function. Nat. Microbiol. 2018, 3, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Zhu, Y.; Su, J.; Xia, B.; Zou, Y.; Nie, J.; Zhang, W.; Wang, J. Butyrate-mediated autophagy inhibition limits cytosolic Salmonella Infantis replication in the colon of pigs treated with a mixture of Lactobacillus and Bacillus. Vet. Res. 2020, 51, 99. [Google Scholar] [CrossRef]
- Travassos, L.H.; Carneiro, L.A.; Ramjeet, M.; Hussey, S.; Kim, Y.G.; Magalhaes, J.G.; Yuan, L.; Soares, F.; Chea, E.; Le Bourhis, L.; et al. Nod1 and Nod2 direct autophagy by recruiting ATG16L1 to the plasma membrane at the site of bacterial entry. Nat. Immunol. 2010, 11, 55–62. [Google Scholar] [CrossRef]
- Lu, Y.; Zheng, Y.; Coyaud, E.; Zhang, C.; Selvabaskaran, A.; Yu, Y.; Xu, Z.; Weng, X.; Chen, J.S.; Meng, Y.; et al. Palmitoylation of NOD1 and NOD2 is required for bacterial sensing. Science 2019, 366, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, Y.; Chu, B.; Yuan, L.; Liu, N.; Zhu, Y.; Wang, J. Lactobacillus johnsonii L531 Alleviates the Damage Caused by Salmonella Typhimurium via Inhibiting TLR4, NF-kappaB, and NLRP3 Inflammasome Signaling Pathways. Microorganisms 2021, 9, 1983. [Google Scholar] [CrossRef] [PubMed]
- Homer, C.R.; Kabi, A.; Marina-Garcia, N.; Sreekumar, A.; Nesvizhskii, A.I.; Nickerson, K.P.; Chinnaiyan, A.M.; Nunez, G.; McDonald, C. A dual role for receptor-interacting protein kinase 2 (RIP2) kinase activity in nucleotide-binding oligomerization domain 2 (NOD2)-dependent autophagy. J. Biol. Chem. 2012, 287, 25565–25576. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.W.; Wilkins, A.; Asara, J.M.; Cantley, L.C. The Crohn’s disease protein, NOD2, requires RIP2 in order to induce ubiquitinylation of a novel site on NEMO. Curr. Biol. 2004, 14, 2217–2227. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Y.H.; Yang, G.Y.; Liu, X.; Xia, B.; Hu, X.; Su, J.H.; Wang, J.F. Lactobacillus rhamnosus GG affects Microbiota and suppresses autophagy in the intestines of pigs challenged with Salmonella Infantis. Front. Microbiol. 2017, 8, 2705. [Google Scholar] [CrossRef]
- Sundaram, B.; Kanneganti, T.D. Advances in Understanding Activation and Function of the NLRC4 Inflammasome. Int. J. Mol. Sci. 2021, 22, 1048. [Google Scholar] [CrossRef]
- Keestra, A.M.; Winter, M.G.; Klein-Douwel, D.; Xavier, M.N.; Winter, S.E.; Kim, A.; Tsolis, R.M.; Baumler, A.J. A Salmonella virulence factor activates the NOD1/NOD2 signaling pathway. mBio 2011, 2, e00266-11. [Google Scholar]
- Gomes, L.C.; Dikic, I. Autophagy in antimicrobial immunity. Mol. Cell 2014, 54, 224–233. [Google Scholar] [CrossRef]
- Kishi-Itakura, C.; Ktistakis, N.T.; Buss, F. Ultrastructural insights into pathogen clearance by autophagy. Traffic 2020, 21, 310–323. [Google Scholar] [CrossRef]
- Huang, J.; Brumell, J.H. Autophagy in immunity against intracellular bacteria. Curr. Top. Microbiol. Immunol. 2009, 335, 189–215. [Google Scholar]
- Chauhan, S.; Mandell, M.A.; Deretic, V. IRGM governs the core autophagy machinery to conduct antimicrobial defense. Mol. Cell 2015, 58, 507–521. [Google Scholar] [CrossRef]
- Akiyama, T.; Oishi, K.; Wullaert, A. Bifidobacteria Prevent Tunicamycin-Induced Endoplasmic Reticulum Stress and Subsequent Barrier Disruption in Human Intestinal Epithelial Caco-2 Monolayers. PLoS ONE 2016, 11, e0162448. [Google Scholar] [CrossRef] [Green Version]
- Raciti, M.; Lotti, L.V.; Valia, S.; Pulcinelli, F.M.; Di Renzo, L. JNK2 is activated during ER stress and promotes cell survival. Cell Death Dis. 2012, 3, e429. [Google Scholar] [CrossRef]
- Bernales, S.; Schuck, S.; Walter, P. ER-phagy: Selective autophagy of the endoplasmic reticulum. Autophagy 2007, 3, 285–287. [Google Scholar] [CrossRef]
- Huang, J.; Brumell, J.H. Bacteria-autophagy interplay: A battle for survival. Nat. Rev. Microbiol. 2016, 12, 101–114. [Google Scholar] [CrossRef] [PubMed]
- García-Gil, A.; Galán-Enríquez, C.S.; Pérez-López, A.; Nava, P.; Alpuche-Aranda, C.; Ortiz-Navarrete, V. SopB activates the Akt-YAP pathway to promote Salmonella survival within B cells. Virulence 2018, 9, 1390–1402. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer name | Direction | Sequence (5′→3′) | GenBank Accession |
|---|---|---|---|
| GAPDH | F | GATTCCACCCACGGCAAGTTCC | NM_001206359 |
| R | AGCACCAGCATCACCCCATTTG | ||
| IL-6 | F | ATAAGGGAAATGTCGAGGCTGTGC | NM_214399 |
| R | GGGTGGTGGCTTTGTCTGGATTC |
| Primer name | Direction | Sequence (5′→3′) | GenBank Accession |
|---|---|---|---|
| GAPDH | F | GGAGCGAGATCCCTCCAAAAT | NM_001289746.2 |
| R | GGCTGTTGTCATACTTCTCATGG | ||
| DDIT3 | F | TCTGGCTTGGCTGACTGAGGAG | NM_001195056.1 |
| R | TTTCCGTTTCCTGGGTCTTCTTTGG | ||
| IL-6 | F | GACAGCCACTCACCTCTTCAGAAC | NM_000600.5 |
| R | GCCTCTTTGCTGCTTTCACACATG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wang, J.-F.; Liu, N.; Wang, X.; Wang, J.; Yang, G.-H.; Yang, G.-Y.; Zhu, Y.-H. Lactobacillusjohnsonii L531 Protects against Salmonella Infantis-Induced Intestinal Damage by Regulating the NOD Activation, Endoplasmic Reticulum Stress, and Autophagy. Int. J. Mol. Sci. 2022, 23, 10395. https://doi.org/10.3390/ijms231810395
Yang L, Wang J-F, Liu N, Wang X, Wang J, Yang G-H, Yang G-Y, Zhu Y-H. Lactobacillusjohnsonii L531 Protects against Salmonella Infantis-Induced Intestinal Damage by Regulating the NOD Activation, Endoplasmic Reticulum Stress, and Autophagy. International Journal of Molecular Sciences. 2022; 23(18):10395. https://doi.org/10.3390/ijms231810395
Chicago/Turabian StyleYang, Lan, Jiu-Feng Wang, Ning Liu, Xue Wang, Jing Wang, Guang-Hui Yang, Gui-Yan Yang, and Yao-Hong Zhu. 2022. "Lactobacillusjohnsonii L531 Protects against Salmonella Infantis-Induced Intestinal Damage by Regulating the NOD Activation, Endoplasmic Reticulum Stress, and Autophagy" International Journal of Molecular Sciences 23, no. 18: 10395. https://doi.org/10.3390/ijms231810395